

Changes in Plant and Grain Quality of Winter Oat (Avena sativa L.) Varieties in Response to Silicon and Sulphur Foliar Fertilisation under Abiotic Stress Conditions

 , ,
, ,  ,
,  , , and
, , and 
Abstract
1. Introduction
- to investigate the effect of treatments on the plant and grain quality parameters of winter oats;
- to examine whether significant differences are found in the mineral composition of the varieties;
- to examine whether there is an interaction between the varieties and the treatments, i.e., whether the varieties respond in the same way to the treatments in terms of protein and element content.
2. Results
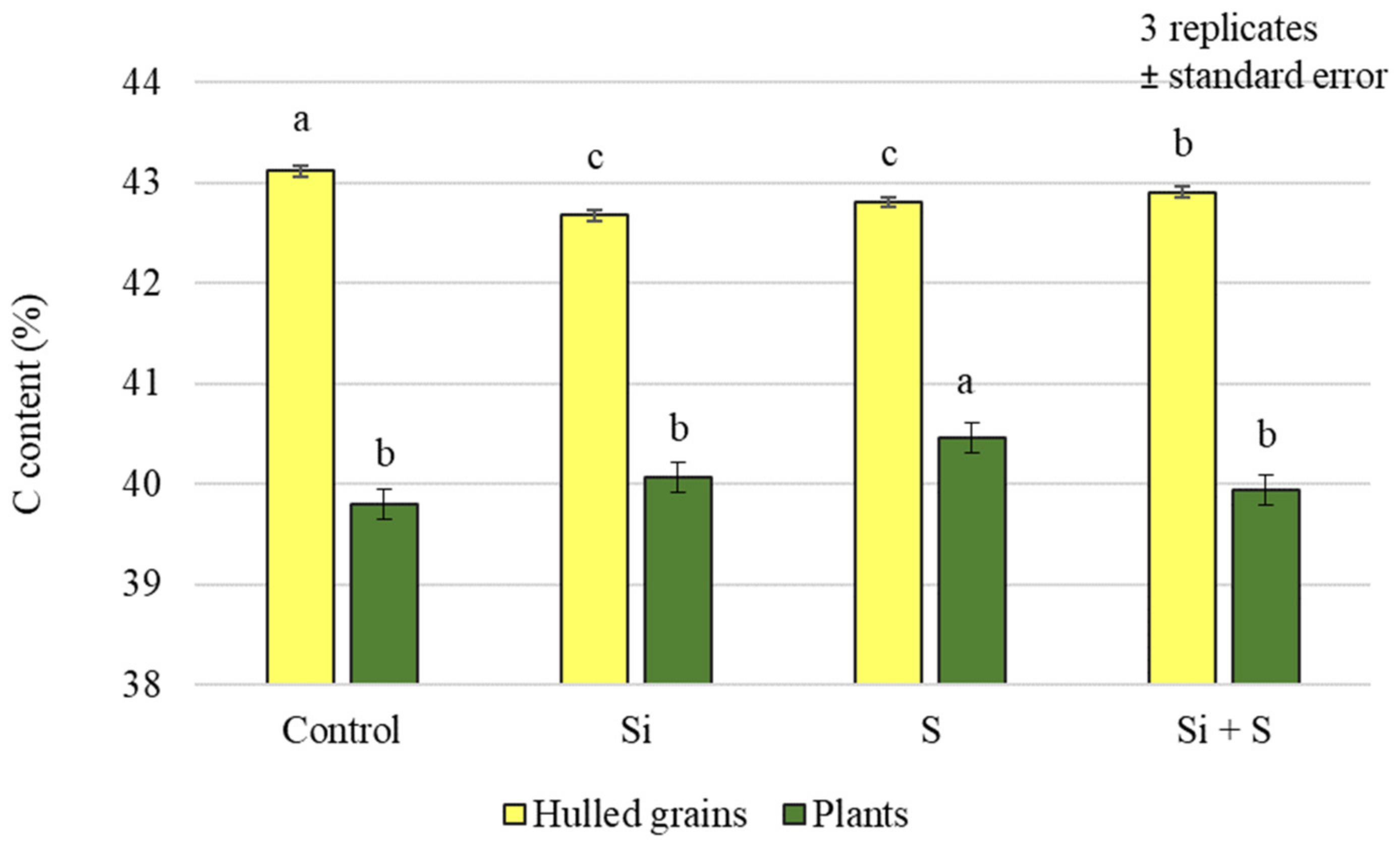
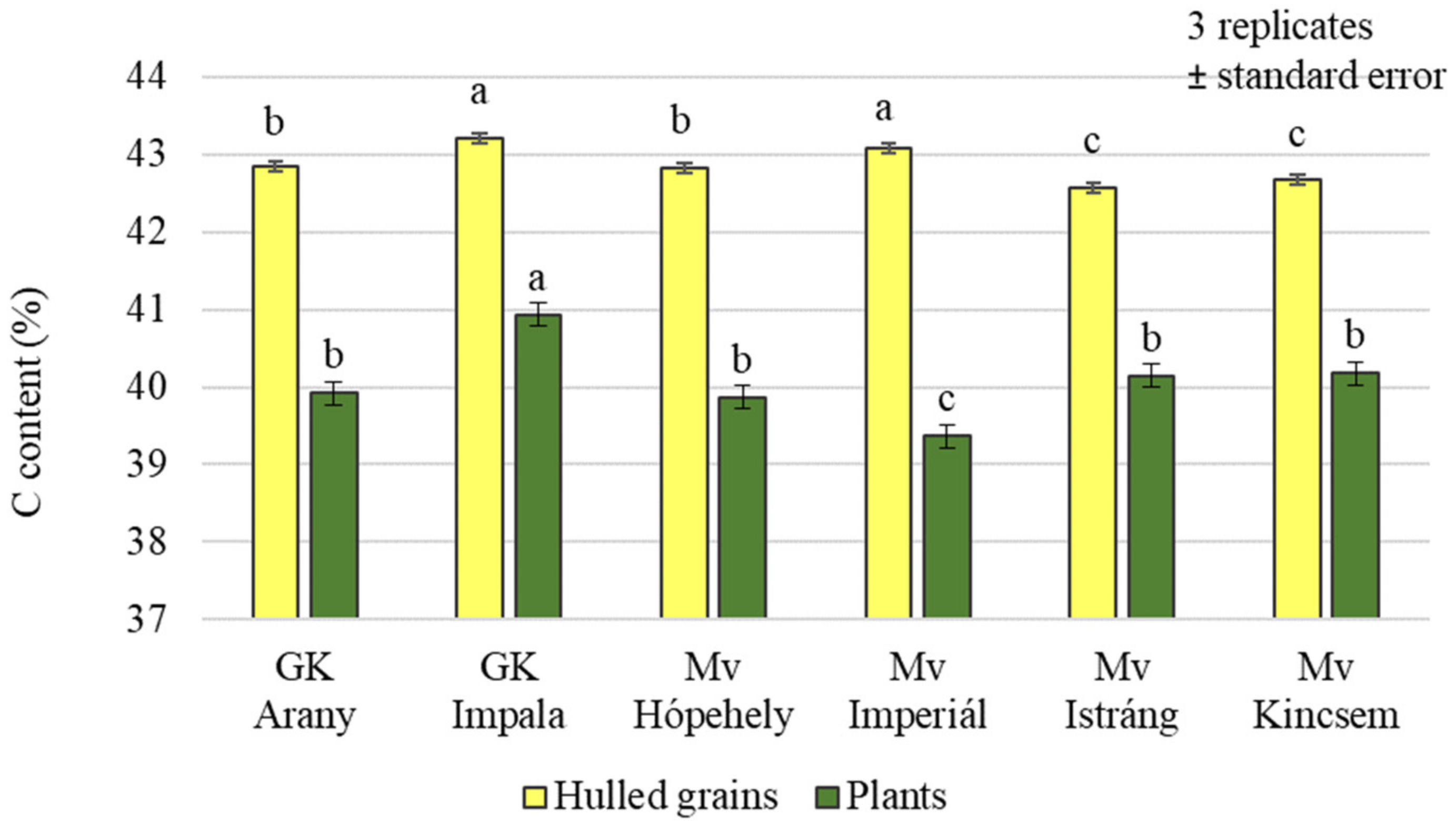
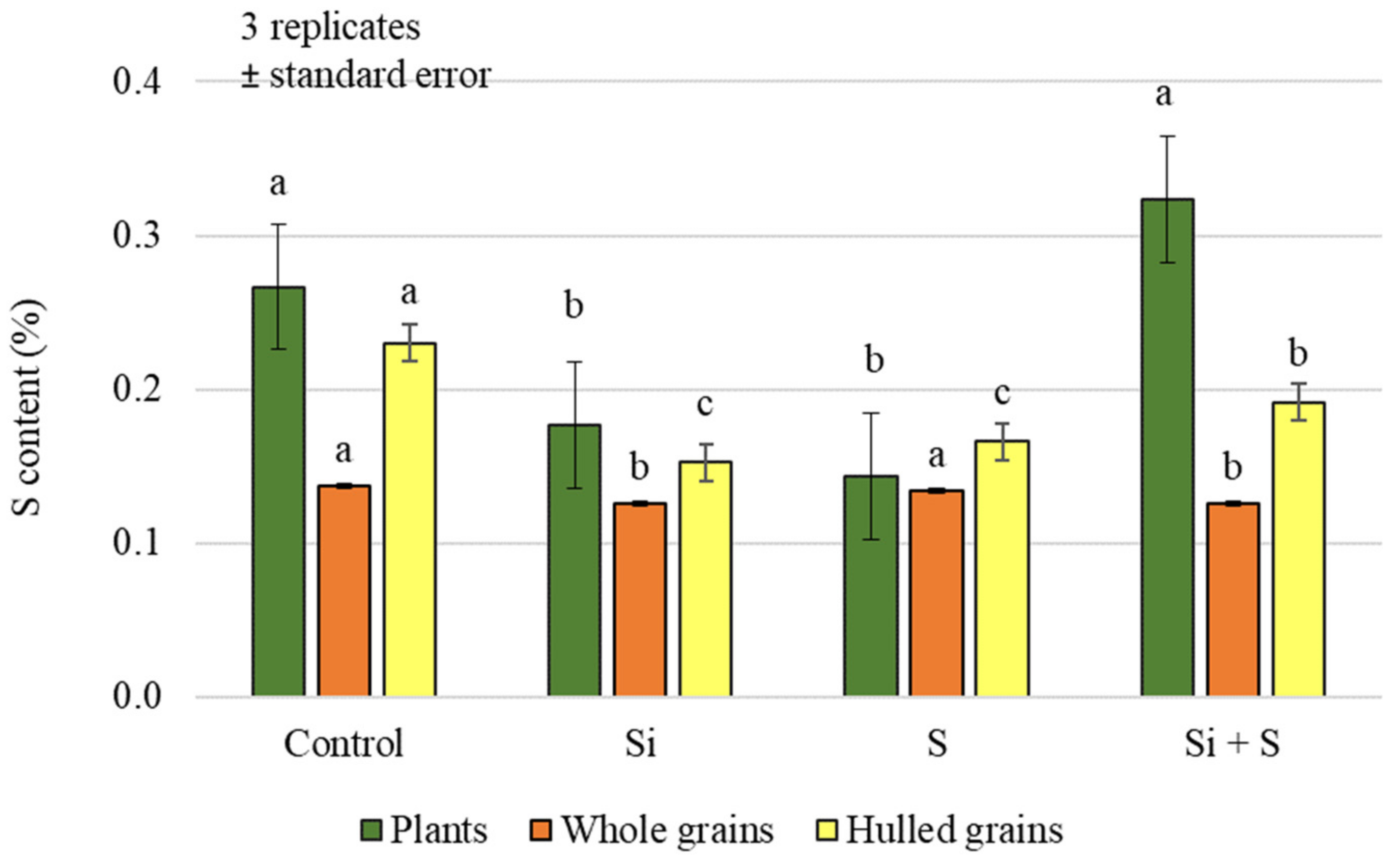
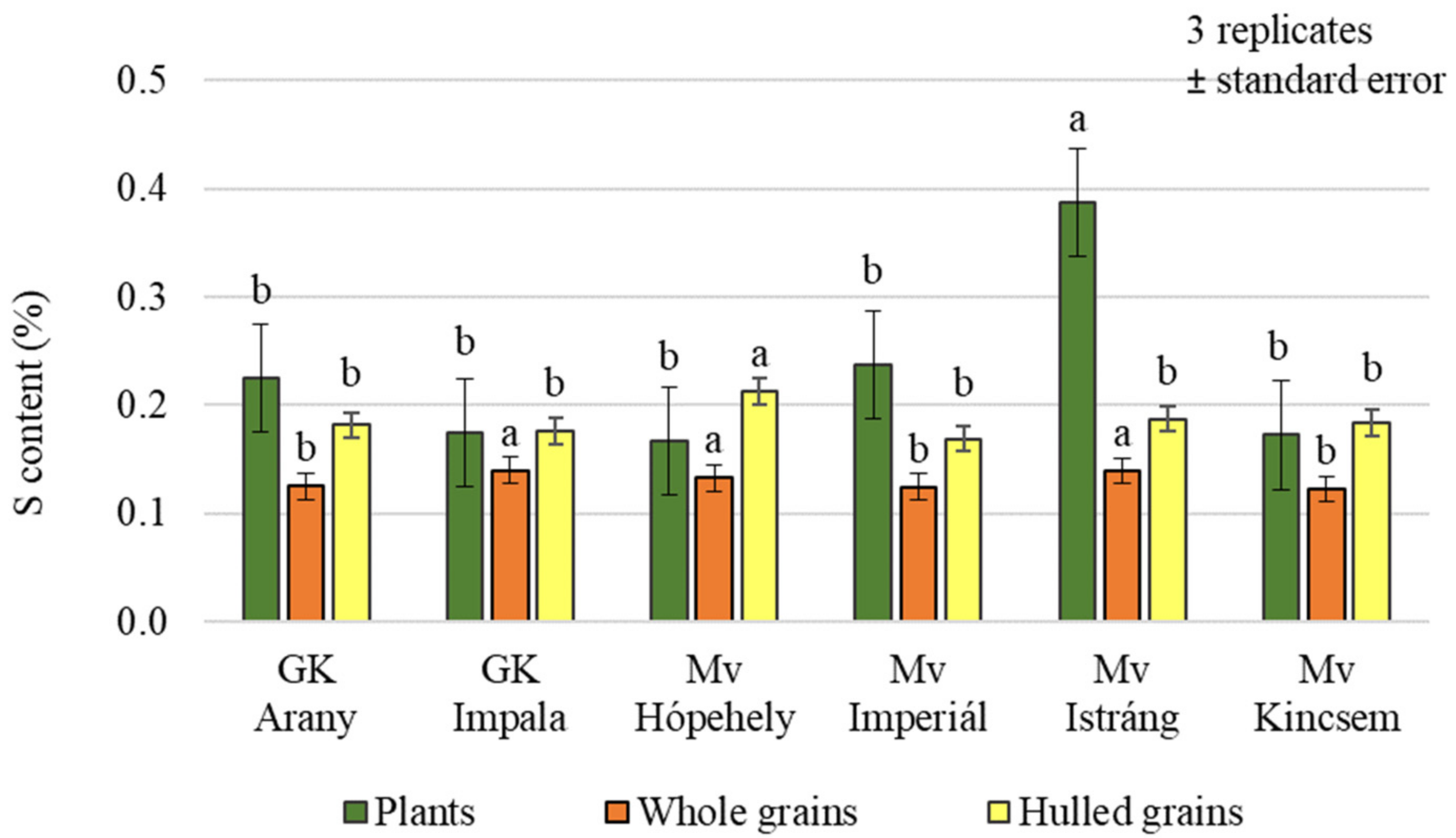
2.1. Effect of Silicon and Sulphur Treatments on the Carbon, Nitrogen, and Sulphur Content of Oat Plants and Hulled Grains
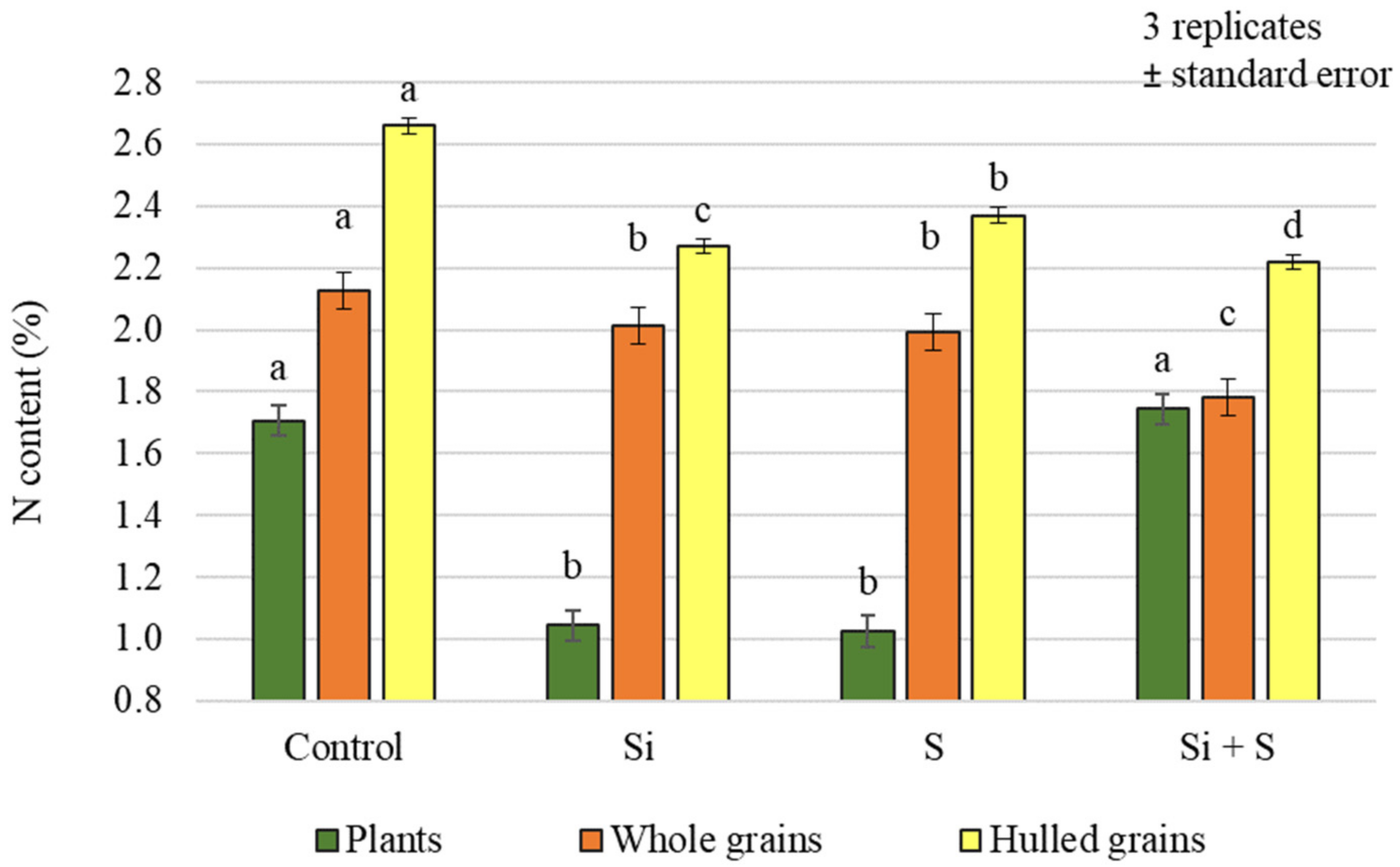
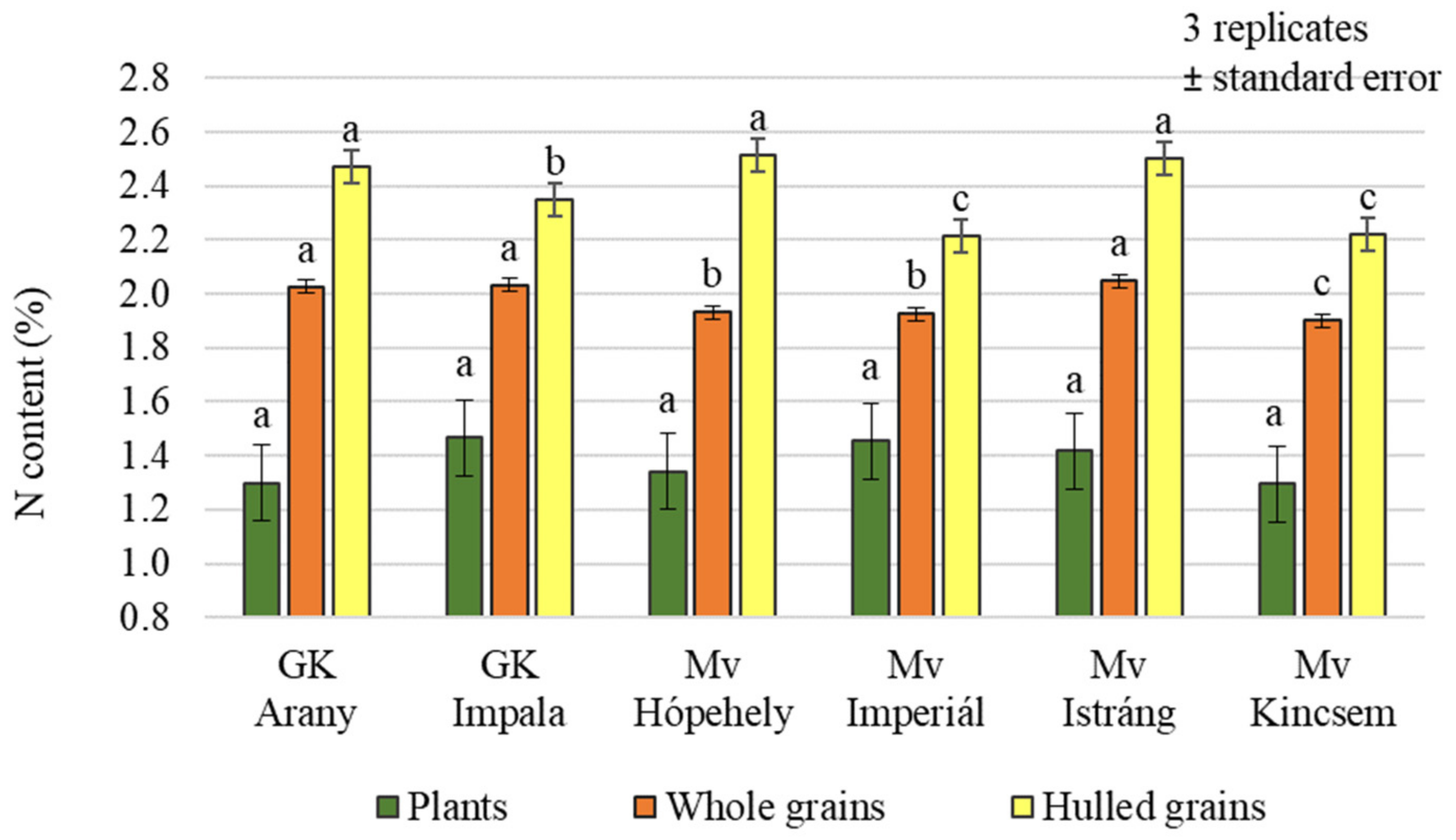
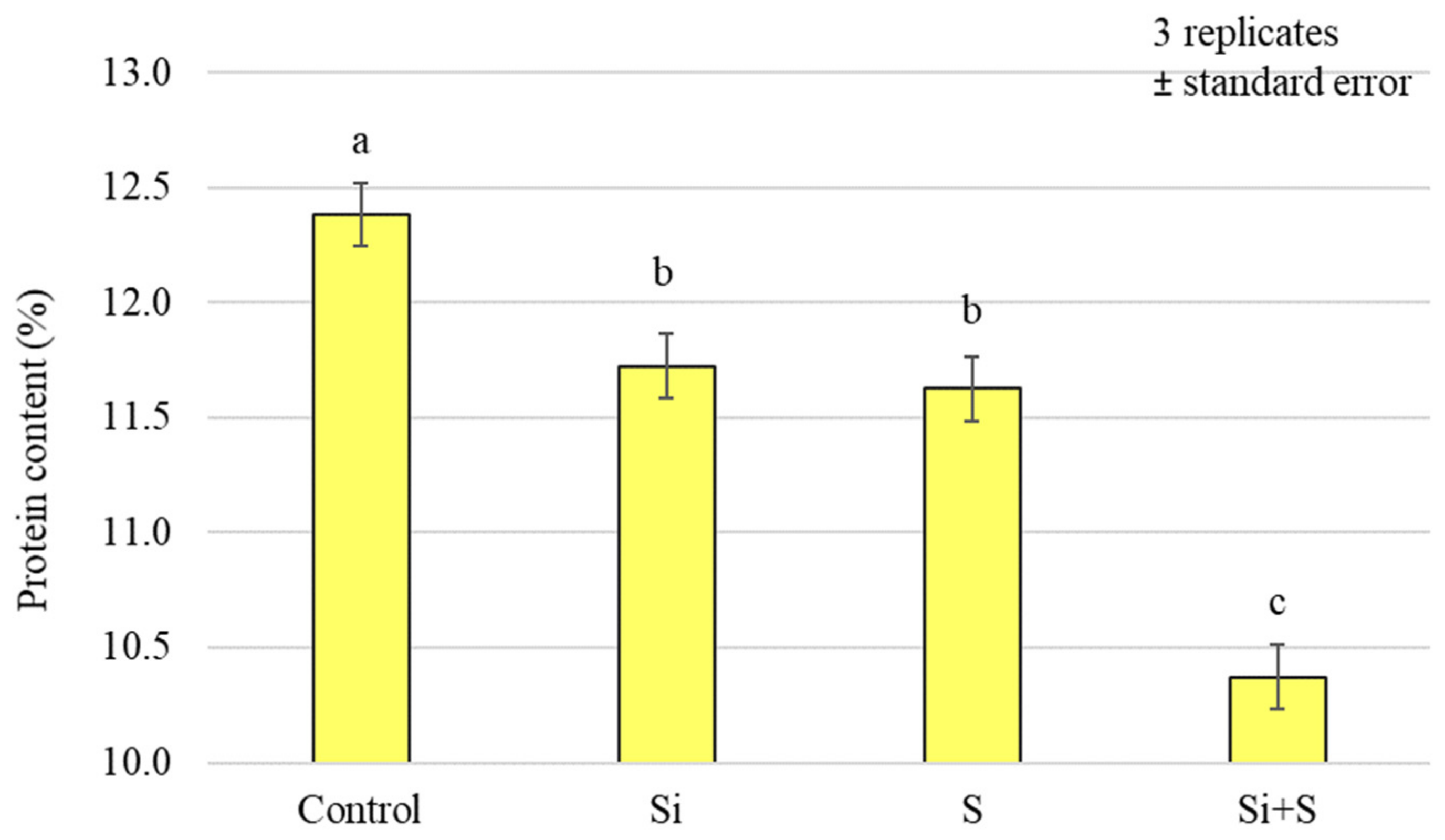
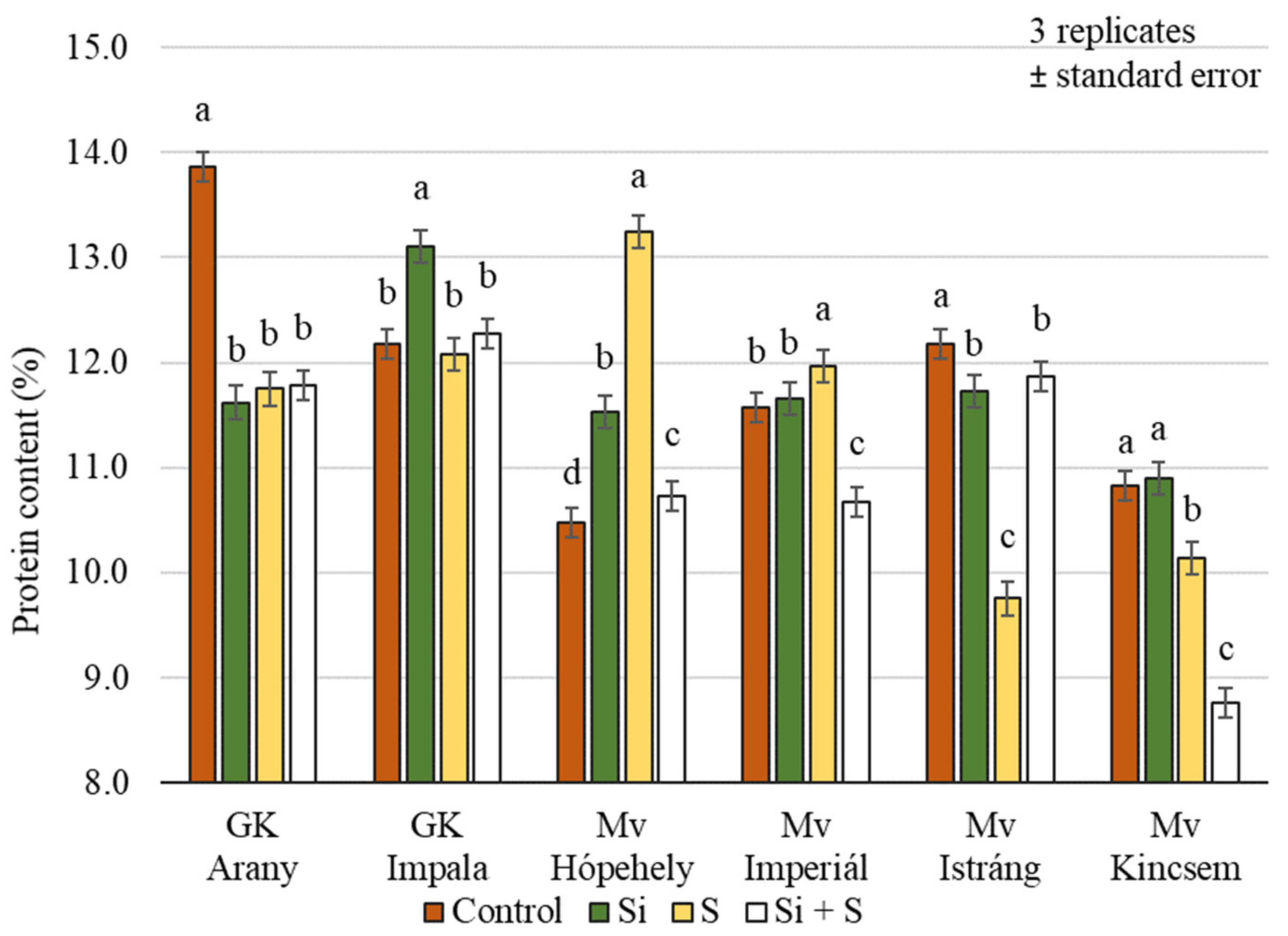
2.2. Effect of Silicon and Sulphur Treatments on the Protein Content of Oat Whole Grains
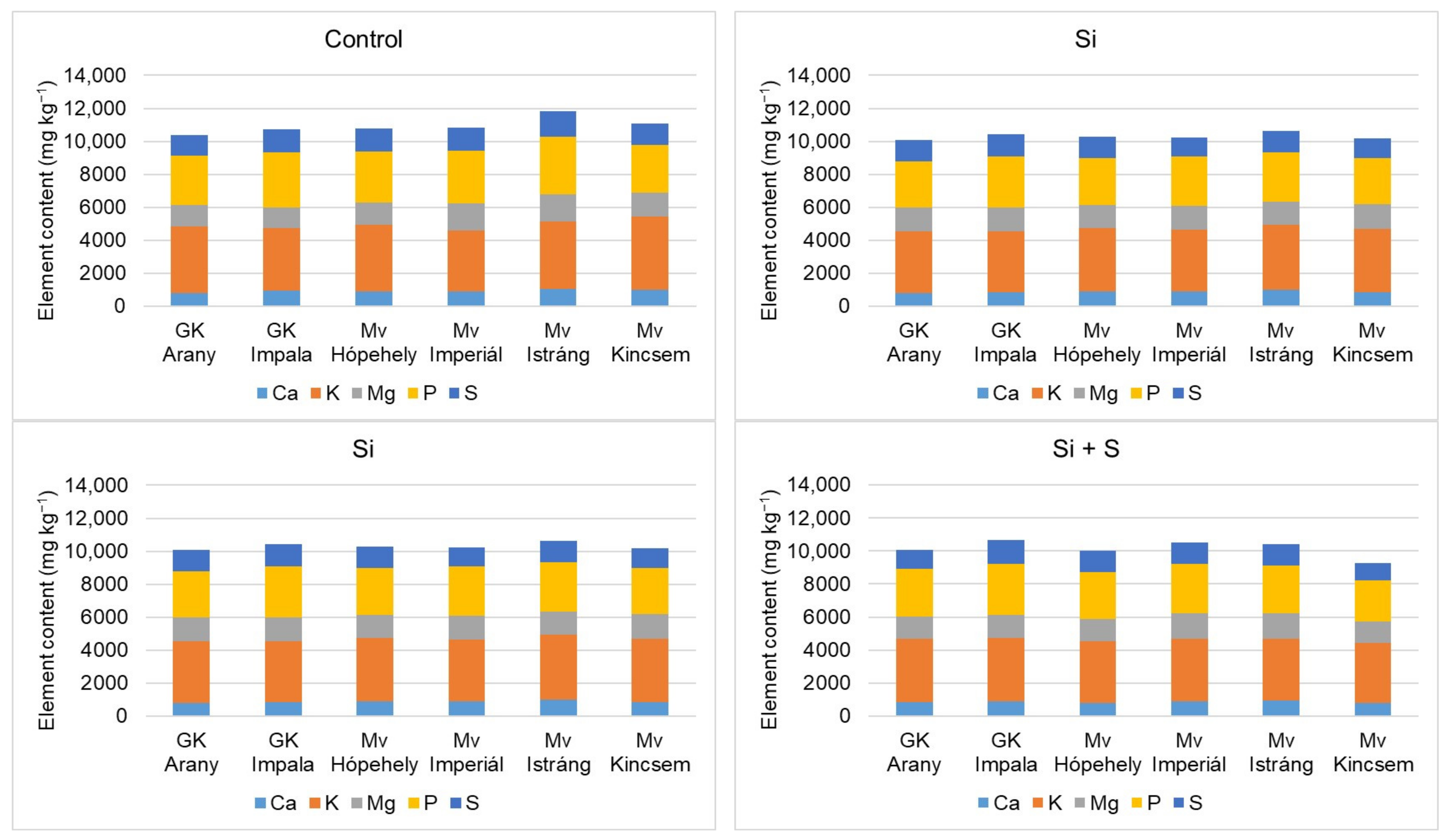
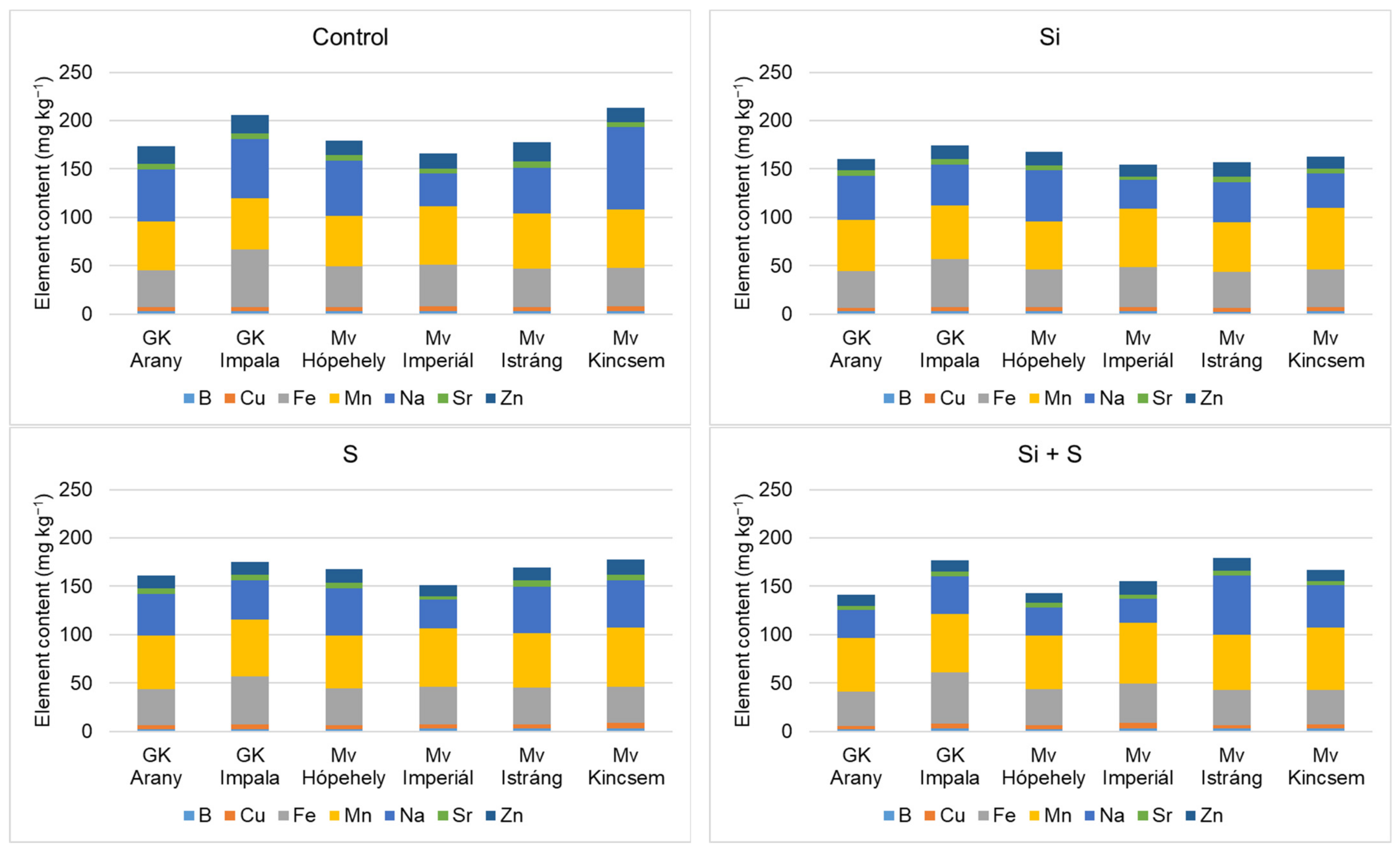
2.3. Effect of Silicon and Sulphur Treatments on the Element Contents of Oat Whole Grains
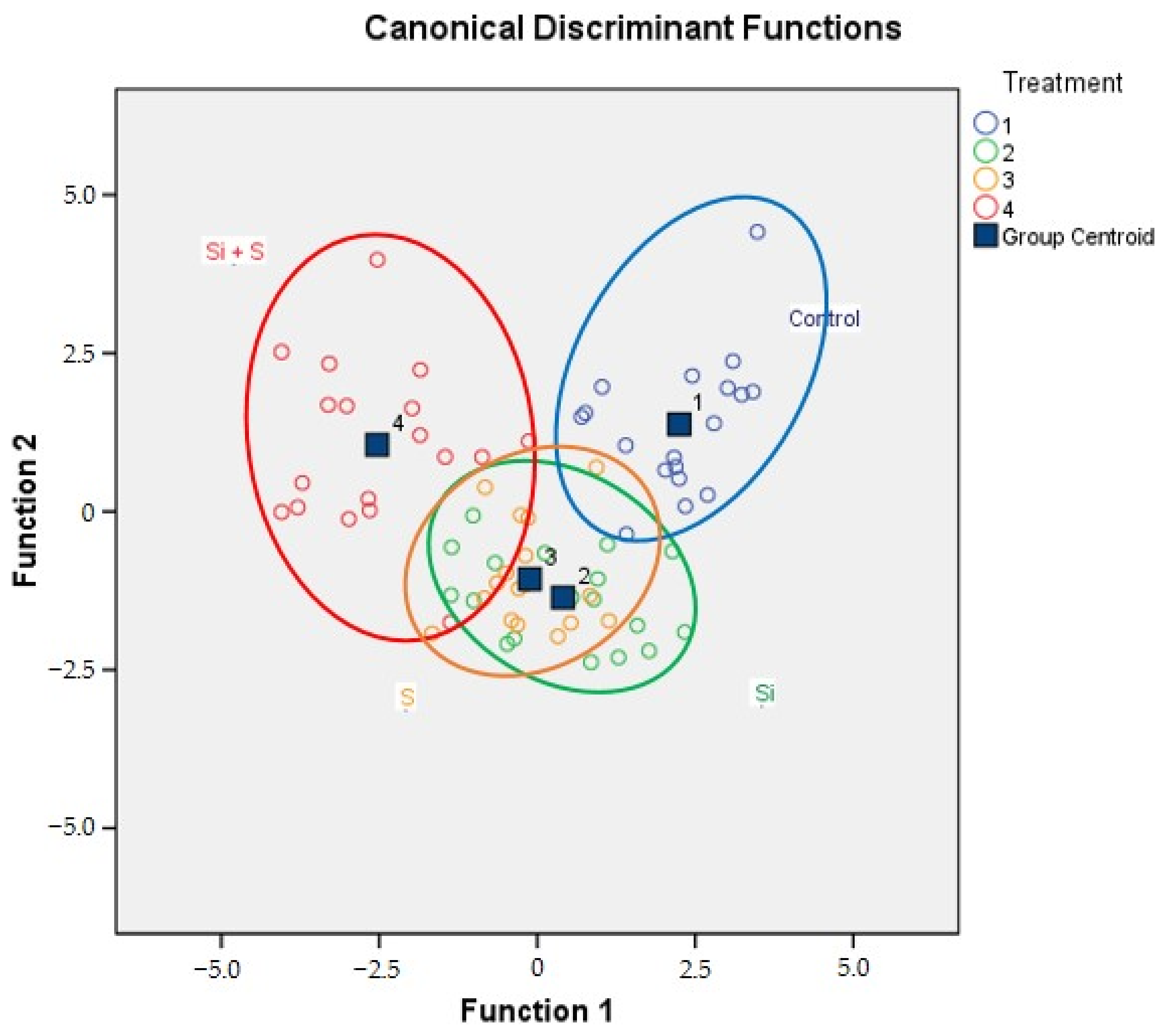
2.4. Results of the Discriminant Analysis
3. Discussion
4. Materials and Methods
4.1. Soil Characteristics of the Experimental Site
4.2. Climatic Conditions
4.3. Experimental Setup
- Control, without foliar fertilisation
- Silicon fertilisation (Si) 3.0 L ha−1
- Sulphur fertilisation (S) 5.0 L ha−1
- Silicon + sulphur fertilisation (Si + S) 3.0 + 5.0 L ha−1
- Sulphur fertiliser: liquid foliar fertiliser with high content sulphur (lignosulfonate formulation) 1000 g L−1 SO3, 30 g L−1 N, 30 g L−1 MgO, 27 g L−1 B, 0.003 g L−1 Mo
- Silicon fertiliser: (potassium silicate formulation) 1.4 m/m% Si, 10.5 m/m% K2O
| 1 December 2020 | BBCH13 (3 leaves unfolded) |
| 10 May 2021 | BBCH39 (flag leaf stage) |
| 18 June 2021 | BBCH77 (late milk) |
4.4. Measurements, Calculations, and Their Methodology
4.4.1. Plant Carbon, Nitrogen, and Sulphur Contents
4.4.2. Grain Carbon, Nitrogen and Sulphur Content
4.4.3. Grain Protein Content by Kjeldahl Method
4.4.4. Grain Element Content
4.5. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kutasy, E.; Buday-Bódi, E.; Virág, I.C.; Forgács, F.; Melash, A.A.; Zsombik, L.; Nagy, A.; Csajbók, J. Mitigating the Negative Effect of Drought Stress in Oat (Avena sativa L.) with Silicon and Sulphur Foliar Fertilization. Plants 2022, 11, 30. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; De, S.; Belkheir, A. Avena sativa (Oat), a potential neutraceutical and therapeutic agent: An overview. Crit. Rev. Food Sci. Nutr. 2013, 53, 126–144. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Li, M.; Chen, Y.; Zhou, Q.; Liu, W.; Liang, G.; Jia, Z. Important Physiological Changes Due to Drought Stress on Oat. Front. Ecol. Evol. 2021, 9, 644726. [Google Scholar] [CrossRef]
- Chen, O.; Mah, E.; Dioum, E.; Marwaha, A.; Shanmugam, S.; Malleshi, N.; Sudha, V.; Gayathri, R.; Unnikrishnan, R.; Anjana, R.M.; et al. The Role of Oat Nutrients in the Immune System: A Narrative Review. Nutrients 2021, 13, 1048. [Google Scholar] [CrossRef] [PubMed]
- Rasane, P.; Jha, A.; Sabikhi, L.; Kumar, A.; Unnikrishnan, V.S. Nutritional advantages of oats and opportunities for its processing as value added foods—A review. J. Food Sci. Technol. 2015, 52, 662–675. [Google Scholar] [CrossRef]
- Isidro-Sánchez, J.; Prats, E.; Howarth, C.; Langdon, T.; Montilla-Bascón, G. Genomic Approaches for Climate Resilience Breeding in Oats. In Genomic Designing of Climate-Smart Cereal Crops; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2020; pp. 133–169. [Google Scholar] [CrossRef]
- Gell, G.; Bugyi, Z.; Florides, C.G.; Birinyi, Z.; Réder, D.; Szegő, Z.; Mucsi, E.; Schall, E.; Ács, K.; Langó, B.; et al. Investigation of Protein and Epitope Characteristics of Oats and Its Implications for Celiac Disease. Front. Nutr. 2021, 8, 702352. [Google Scholar] [CrossRef]
- Martínez-Villaluenga, C.; Peñas, E. Health benefits of oat: Current evidence and molecular mechanisms. Curr. Opin. Food Sci. 2017, 14, 26–31. [Google Scholar] [CrossRef]
- Pavlovic, J.; Kostic, L.; Bosnic, P.; Kirkby, E.A.; Nikolic, M. Interactions of Silicon with Essential and Beneficial Elements in Plants. Front. Plant Sci. 2021, 12, 697592. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, W.; Lv, Y.; Li, T.; Tang, J.; Yang, X.; Bai, J.; Jin, X.; Zhou, H. Effects of drought stress during critical periods on the photosynthetic characteristics and production performance of Naked oat (Avena nuda L.). Sci. Rep. 2022, 12, 11199. [Google Scholar] [CrossRef]
- Gebrewahid, L.; Mengistu, D.K.; Tsehaye, Y.; Aberha, A.; Aberra, D.A. Variability among Ethiopian durum wheat genotypes grown under different climatic conditions of Tigray for some agronomic and grain-quality traits. J. Crop Improv. 2021, 35, 184–203. [Google Scholar] [CrossRef]
- Asseng, S.; Martre, P.; Maiorano, A.; Rötter, R.P.; O’Leary, G.J.; Fitzgerald, G.J.; Girousse, C.; Motzo, R.; Giunta, F.; Babar, M.A.; et al. Climate change impact and adaptation for wheat protein. Glob. Chang. Biol. 2019, 25, 155–173. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.M.; Qiu, S.N.; Li, Q.Y.; Di, G.L.; Wang, J.L.; Sun, H.X. Simulated global climate change benefits the nutritive value of oat grass. Plant Biol. J. 2022, 24, 846–853. [Google Scholar] [CrossRef] [PubMed]
- Bista, D.R.; Heckathorn, S.A.; Jayawardena, D.M.; Mishra, S.; Boldt, J.K. Effects of Drought on Nutrient Uptake and the Levels of Nutrient-Uptake Proteins in Roots of Drought-Sensitive and -Tolerant Grasses. Plants 2018, 7, 28. [Google Scholar] [CrossRef] [PubMed]
- Melash, A.A.; Ábrahám, É.B. Barriers and levers to enhance end-use functional properties of durum wheat (Triticum turgidum L.) grain: An agronomic implication. Heliyon 2022, 8, e09542. [Google Scholar] [CrossRef]
- Sallam, A.; Alqudah, A.M.; Dawood, M.F.A.; Baenziger, P.S.; Börner, A. Drought Stress Tolerance in Wheat and Barley: Advances in Physiology, Breeding and Genetics Research. Int. J. Mol. Sci. 2019, 20, 3137. [Google Scholar] [CrossRef]
- Krishan, K.V.; Xiu-Peng, S.; Dan-Dan, T.; Munna, S.; Chhedi, L.V.; Vishnu, D.R.; Singh, R.K.; Sharma, A.; Singh, P.; Malviya, M.K.; et al. Investigation of Defensive Role of Silicon during Drought Stress Induced by Irrigation Capacity in Sugarcane: Physiological and Biochemical Characteristics. ACS Omega 2021, 6, 19811–19821. [Google Scholar] [CrossRef]
- Wang, M.; Wang, R.; Mur, L.A.; Jianyun, R.; Qirong, S.; Shiwei, G. Functions of Silicon in plant drought stress responses. Hortic. Res. 2021, 8, 254. [Google Scholar] [CrossRef]
- Kovács, S.; Kutasy, E.; Csajbók, J. The Multiple Role of Silicon Nutrition in Alleviating Environmental Stresses in Sustainable Crop Production. Plants 2022, 11, 1223. [Google Scholar] [CrossRef]
- Lux, A.; Lukačová, Z.; Vaculík, M.; Švubová, R.; Kohanová, J.; Soukup, M.; Martinka, M.; Bokor, B. Silicification of Root Tissues. Plants 2020, 9, 111. [Google Scholar] [CrossRef]
- Marylyn, M.; Hussein, S.; Mark, D.L.; Toi, J.; Isack, M. Breeding for Silicon-use efficiency, protein content and drought tolerance in bread wheat (Triticum aestivum L.): A review. Acta Agric. Scand. Sect. B-Soil Plant Sci. 2022, 72, 17–29. [Google Scholar] [CrossRef]
- Hanna, K.; Marek, M. Sulphur and Nitrogen Fertilization as a Potential Means of Agronomic Biofortification to Improve the Content and Uptake of Microelements in Spring Wheat Grain DM. J. Chem. 2018, 2018, 9326820. [Google Scholar] [CrossRef]
- Barczak, B.; Klikocka, H.; Kozera, W.; Knapowski, T. Assessment of the effect of sulphur fertilisation on oat grain yield and micronutrient uptake. J. Elem. 2018, 23, 45–56. [Google Scholar] [CrossRef]
- Meruyert, K.; Tolganai, S.; Zhanar, T.; Tursynbek, K.; Adil, K.; Shermakhan, S.; Aigul, M.; Mukhambetkali, B.; Nadezhda, B. Influence of new sulfur-containing fertilizers on performance of wheat yield. Saudi J. Biol. Sci. 2021, 28, 4644–4655. [Google Scholar] [CrossRef]
- Peterson, D.M. Oat antioxidants. J. Cereal Sci. 2001, 33, 115–129. [Google Scholar] [CrossRef]
- Flander, L.; Salmenkallio-Marttila, M.; Suortti, T.; Autio, K. Optimization of ingredients and baking process for improved wholemeal oat bread quality. LWT-Food Sci. Technol. 2007, 40, 860–870. [Google Scholar] [CrossRef]
- Liu, L.; Song, Z.; Yu, C.; Yu, G.; Ellam, R.M.; Liu, H.; Singh, B.P.; Wang, H. Silicon Effects on Biomass Carbon and Phytolith-Occluded Carbon in Grasslands Under High-Salinity Conditions. Front. Plant Sci. 2020, 11, 657. [Google Scholar] [CrossRef] [PubMed]
- Wiggans, S.C.; Frey, K.J. Nitrogen Uptake in Oats. Proc. Iowa Acad. Sci. 1956, 63, 266–273. [Google Scholar]
- Redaelli, R.; Scalfati, G.; Ciccoritti, R.; Cacciatori, P.; Stefanis, E.; Sgrulletta, D. Effects of Genetic and Agronomic Factors on GrainComposition in Oats. Cereal Res. Commun. 2015, 43, 144–154. [Google Scholar] [CrossRef]
- Helal, N.M.; Khattab, H.I.; Emam, M.M.; Niedbała, G.; Wojciechowski, T.; Hammami, I.; Alabdallah, N.M.; Darwish, D.B.E.; El-Mogy, M.M.; Hassan, H.M. Improving Yield Components and Desirable Eating Quality of Two Wheat Genotypes Using Si and NanoSi Particles under Heat Stress. Plants 2022, 11, 1819. [Google Scholar] [CrossRef]
- Tao, Z.; Chang, X.; Wang, D.; Wang, Y.; Ma, S.; Yang, Y.; Zhao, G. Effects of sulfur fertilization and short-term high temperature on wheat grain production and wheat flour proteins. Crop J. 2018, 6, 413–425. [Google Scholar] [CrossRef]
- Zenda, T.; Liu, S.; Dong, A.; Duan, H. Revisiting Sulphur—The Once Neglected Nutrient: It’s Roles in Plant Growth, Metabolism, Stress Tolerance and Crop Production. Agriculture 2021, 11, 626. [Google Scholar] [CrossRef]
- Alemayehu, G.F.; Forsido, S.F.; Tola, Y.B.; Teshager, M.A.; Assegie, A.A.; Amare, E. Proximate, mineral and anti-nutrient compositions of oat grains (Avena sativa) cultivated in Ethiopia: Implications for nutrition and mineral bioavailability. Heliyon 2021, 7, e07722. [Google Scholar] [CrossRef]
- Jinger, D.; Dhar, S.; Dass, A.; Sharma, V.K.; Paramesh, V.; Parihar, M.; Joshi, E.; Singhal, V.; Gupta, G.; Prasad, D.; et al. Co-fertilization of Silicon and Phosphorus Influences the Dry Matter Accumulation, Grain Yield, Nutrient Uptake, and Nutrient-Use Efficiencies of Aerobic Rice. Silicon 2022, 14, 4683–4697. [Google Scholar] [CrossRef]
- Shivay, Y.S.; Prasad, R.; Pal, M. Zinc Fortification of Oat Grains Through Zinc Fertilisation. Agric. Res. 2013, 2, 375–381. [Google Scholar] [CrossRef]
- Özcan, M.A.; Bagci, A.; Dursun, N.; Gezgin, S.; Hamurcu, M.; Dumlupinar, Z.; Uslu, N. Macro and Micro Element Contents of Several Oat (Avena sativa L.) Genotype and Variety Grains. Iran. J. Chem. Chem. Eng. 2017, 36, 73–79. [Google Scholar] [CrossRef]
- Bityutskii, N.; Loskutov, I.; Yakkonen, K.; Konarev, A.; Shelenga, T.; Khoreva, V.; Blinova, E.; Rymin, A. Screening of Avena sativa cultivars for iron, zinc, manganese, protein and oil contents and fatty acid composition in whole grains. Cereal Res. Commun. 2020, 48, 87–94. [Google Scholar] [CrossRef]
- Loskutov, I.G.; Khlestkina, E.K. Wheat, Barley, and Oat Breeding for Health Benefit Components in Grain. Plants 2021, 10, 86. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Hou, S.; Sun, Y.; Dang, H.; Song, Q.; Jiang, L.; Cao, W.; Wang, H.; He, X.; Wang, Z. Regional wheat grain manganese and its potential risks affected by soil pH and precipitation. J. Clean. Prod. 2020, 264, 121677. [Google Scholar] [CrossRef]
- Li, Z.; Song, Z.; Yang, X.; Song, A.; Yu, C.; Wang, T.; Xia, S.; Liang, Y. Impacts of silicon on biogeochemical cycles of carbon and nutrients in croplands. J. Integr. Agric. 2018, 17, 2182–2195. [Google Scholar] [CrossRef]
- Artyszak, A. Effect of Silicon Fertilization on Crop Yield Quantity and Quality—A Literature Review in Europe. Plants 2018, 7, 54. [Google Scholar] [CrossRef]
- Walsh, O.S.; Shafian, S.; McClintick-Chess, J.R.; Belmont, K.M.; Blanscet, S.M. Potential of Silicon Amendment for Improved Wheat Production. Plants 2018, 7, 26. [Google Scholar] [CrossRef] [PubMed]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014, International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports: Rome, Italy, 2015. [Google Scholar]
- Meier, U. Growth Stages of Mono and Dicotyledonous Plants. BBCH Monograph; Julius Kühn-Institut (JKI): Quedlinburg, Germany, 2018. [Google Scholar] [CrossRef]
- Matejovic, I. Total nitrogen in plant material determinated by means of dry combustion: A possible alternative to determination by Kjeldahl digestion. Commun. Soil. Sci. Plant Anal. 1995, 26, 2217–2229. [Google Scholar] [CrossRef]
- Nagy, P.T. Égetéses elven működő elemanalizátor alkalmazhatósága talaj- és növényvizsgálatokban. (Application of dry combustion element analyser in soil and crop testing). Agrokémia és Talajtan 2000, 49, 521–534. (In Hungarian) [Google Scholar]
- Capouchová, I.; Petr, J.; Tlaskalová-Hogenová, H.; Michalík, I.; Faměra, O.; Urminská, D.; Tučková, L.; Knoblochová, H.; Borovská, D. Protein fractions of oats and possibilities of oat utilisation for patients with coeliac disease. Czech J. Food Sci. 2004, 22, 151–162. [Google Scholar] [CrossRef]
- Sharma, G.M.; Pereira, M.; Wang, S.S.; Chirtel, S.J.; Whitaker, T.B.; Wehling, P.; Arlinghaus, M.; Canida, T.; Jackson, L.S.; Williams, K.M. Evaluation of sampling plans for measurement of gluten in oat groats. Food Control. 2020, 114, 107241. [Google Scholar] [CrossRef]
- Decker, E.; Rose, D.; Stewart, D. Processing of oats and the impact of processing operations on nutrition and health benefits. Br. J. Nutr. 2014, 112, S58–S64. [Google Scholar] [CrossRef]
- Sunilkumar, B.A.; Tareke, E. Review of analytical methods for measurement of oat proteins: The need for standardized methods. Crit. Rev. Food Sci. Nutr. 2019, 59, 1467–1485. [Google Scholar] [CrossRef] [PubMed]
- Kjeldahl, J. Neue Methode zur Bestimmung des Stickstoffs in organischen Körpern (New method for the determination of nitrogen in organic substances). Z. Anal. Chem. 1883, 22, 366–383. [Google Scholar] [CrossRef]
- Jones, D.B. Factors for converting percentages of nitrogen in foods and feeds into percentages of protein. USDA Circ. 1931, 183, 1–21. [Google Scholar]
- Kovács, B.; Győri, Z.; Prokisch, J.; Loch, J.; Dániel, P. A study of plant sample preparation and inductively coupled plasma emission spectrometry parameters. Commun. Soil. Sci. Plant Anal. 1996, 27, 1177–1198. [Google Scholar] [CrossRef]
- Forgács, F.Z.; Kutasy, E.T. Szilícium és kén állománykezelés hatása az őszi zab genotípusok levélterületére és produktivitására eltérő évjáratokban. (Effect of silicon and sulphur fertilisation on leaf area and productivity of winter oat genotypes in different years). In 11th Interdisciplinary Doctoral Conference 2022 Conference Book, Pécs, Hungary, 25–26 November 2022; Kajos, L.F., Bali, C., Puskás, T., Szabó, R., Eds.; Doctoral Student Association of the University of Pécs: Pécs, Hungary, 2023; pp. 169–180, (In Hungarian). ISBN 978-963-626-070-5. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Si | S | Si + S | |
|---|---|---|---|---|
| Al | 1.396 ± 0.162 a | 1.017 ± 0.188 b | 0.972 ± 0.130 b | 0.671 ± 0.132 c |
| B | 3.070 ± 0.071 a | 2.833 ± 0.085 a | 2.783 ± 0.117 a | 2.882 ± 0.129 a |
| Ba | 0.895 ± 0.057 c | 1.007 ± 0.081 b | 1.121 ± 0.114 a | 1.072 ± 0.097 b |
| Ca | 943.00 ± 23.73 a | 894.00 ± 18.85 b | 903.83 ± 18.81 b | 854.67 ± 16.94 c |
| Cu | 4.373 ± 0.078 a | 4.082 ± 0.095 b | 4.394 ± 0.149 a | 4.048 ± 0.193 b |
| Fe | 43.456 ± 1.889 a | 40.689 ± 1.312 b | 39.778 ± 1.041 b | 39.822 ± 1.527 b |
| K | 4002.61 ± 69.15 a | 3792.06 ± 27.46 b | 3947.33 ± 59.20 a | 3779.06 ± 26.87 b |
| Li | 0.134 ± 0.004 b | 0.136 ± 0.005 b | 0.138 ± 0.008 b | 0.142 ± 0.009 a |
| Mg | 1444.89 ± 44.20 a | 1422.56 ± 28.27 a | 1453.94 ± 25.33 a | 1399.83 ± 30.99 a |
| Mn | 55.794 ± 1.017 c | 55.561 ± 1.320 c | 57.750 ± 1.118 b | 59.194 ± 1.153 a |
| Mo | 1.416 ± 0.059 c | 1.534 ± 0.052 c | 1.572 ± 0.086 b | 1.869 ± 0.081 a |
| Na | 56.461 ± 4.012 a | 41.167 ± 2.205 b | 43.250 ± 1.955 b | 37.706 ± 3.243 c |
| Ni | 1.492 ± 0.075 a | 1.306 ± 0.073 c | 1.527 ± 0.069 a | 1.395 ± 0.100 b |
| P | 3168.94 ± 50.40 a | 2926.89 ± 55.05 c | 3015.50 ± 40.57 b | 2869.72 ± 53.22 c |
| Pb | 0.377 ± 0.057 a | 0.169 ± 0.042 c | 0.134 ± 0.039 c | 0.230 ± 0.050 b |
| S | 1375.39 ± 28.40 a | 1258.44 ± 22.29 b | 1342.00 ± 27.40 a | 1254.33 ± 32.21 b |
| Sr | 5.600 ± 0.186 a | 5.226 ± 0.193 a | 5.535 ± 0.227 a | 4.727 ± 0.110 b |
| Zn | 17.083 ± 0.565 a | 12.913 ± 0.426 b | 13.456 ± 0.404 b | 11.921 ± 0.394 c |
| Ca | Cu | Mo | Fe | K | Mg | P | S | Zn | Ni | Na | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Ca | 1 | 0.213 | −0.409 ** | 0.100 | 0.528 ** | 0.235 * | 0.646 ** | 0.539 ** | 0.540 ** | 0.075 | 0.320 ** |
| Cu | 0.213 | 1 | −0.007 | 0.423 ** | 0.242 * | 0.109 | 0.325 ** | 0.342 ** | 0.317 ** | 0.706 ** | 0.132 |
| Mo | −0.409 ** | −0.007 | 1 | 0.169 | −0.455 ** | 0.251 | −0.177 | −0.159 | −0.465 ** | 0.242 | −0.655 ** |
| Fe | 0.100 | 0.423 ** | 0.169 | 1 | −0.227 | −0.018 | 0.476 ** | 0.398 ** | 0.287 * | 0.360 ** | 0.095 |
| K | 0.528 ** | 0.242 * | −0.455 ** | −0.227 | 1 | 0.070 | 0.317 ** | 0.331 ** | 0.405 ** | 0.127 | 0.506 ** |
| Mg | 0.235 * | 0.109 | 0.251 | −0.018 | 0.070 | 1 | 0.351 ** | 0.369 ** | 0.228 | 0.224 | −0.156 |
| P | 0.646 ** | 0.325 ** | −0.177 | 0.476 ** | 0.317 ** | 0.351 ** | 1 | 0.774 ** | 0.683 ** | 0.195 | 0.143 |
| S | 0.539 ** | 0.432 ** | −0.159 | 0.398 ** | 0.331 ** | 0.369 ** | 0.774 ** | 1 | 0.575 ** | 0.114 | 0.208 |
| Zn | 0.540 ** | 0.317 ** | −0.465 ** | 0.287 * | 0.405 ** | 0.228 | 0.683 ** | 0.575 ** | 1 | 0.126 | 0.420 ** |
| Ni | 0.075 | 0.706 ** | 0.242 | 0.360 ** | 0.127 | 0.224 | 0.195 | 0.114 | 0.126 | 1 | −0.019 |
| Na | 0.320 ** | 0.132 | −0.655 ** | 0.095 | 0.506 ** | −0.156 | 0.143 | 0.208 | 0.420 ** | −0.019 | 1 |
| Treatment | Predicted Group Membership | |||
|---|---|---|---|---|
| Control | Si | S | Si + S | |
| Control | 94.4% | 0% | 5.6% | 0% |
| Si | 0% | 77.8% | 22.2% | 0% |
| S | 0% | 11.1% | 83.3% | 5.6% |
| Si + S | 0% | 11.1% | 0% | 88.9% |
| Layer 0–20 cm | Layer 20–40 cm | Layer 40–60 cm | |
|---|---|---|---|
| pH (H2O) | 8.30 | 8.36 | 8.43 |
| KA | 38 | 38 | 38 |
| CaCO3 (%) | 8.1 | 8.1 | 8.1 |
| Humus (%) | 3.66 | 2.92 | 2.70 |
| NO3 + NO2 (mg kg−1) | 1.71 | 2.95 | 3.18 |
| NH4 (mg kg−1) | 0.84 | 1.02 | 3.18 |
| P2O5 (AL) (mg kg−1) | 1671.6 | 1376.1 | 1076.8 |
| K2O (AL) (mg kg−1) | 658.9 | 648.2 | 525.5 |
| SO4 (mg kg−1) | 3.07 | 6.00 | 7.81 |
| Zn (EDTA) (mg kg−1) | 2.77 | 2.24 | 1.84 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kutasy, E.; Diósi, G.; Buday-Bódi, E.; Nagy, P.T.; Melash, A.A.; Forgács, F.Z.; Virág, I.C.; Vad, A.M.; Bytyqi, B.; Buday, T.; et al. Changes in Plant and Grain Quality of Winter Oat (Avena sativa L.) Varieties in Response to Silicon and Sulphur Foliar Fertilisation under Abiotic Stress Conditions. Plants 2023, 12, 969. https://doi.org/10.3390/plants12040969
Kutasy E, Diósi G, Buday-Bódi E, Nagy PT, Melash AA, Forgács FZ, Virág IC, Vad AM, Bytyqi B, Buday T, et al. Changes in Plant and Grain Quality of Winter Oat (Avena sativa L.) Varieties in Response to Silicon and Sulphur Foliar Fertilisation under Abiotic Stress Conditions. Plants. 2023; 12(4):969. https://doi.org/10.3390/plants12040969
Chicago/Turabian StyleKutasy, Erika, Gerda Diósi, Erika Buday-Bódi, Péter Tamás Nagy, Anteneh Agezew Melash, Fanni Zsuzsa Forgács, István Csaba Virág, Attila Miklós Vad, Bekir Bytyqi, Tamás Buday, and et al. 2023. "Changes in Plant and Grain Quality of Winter Oat (Avena sativa L.) Varieties in Response to Silicon and Sulphur Foliar Fertilisation under Abiotic Stress Conditions" Plants 12, no. 4: 969. https://doi.org/10.3390/plants12040969
APA StyleKutasy, E., Diósi, G., Buday-Bódi, E., Nagy, P. T., Melash, A. A., Forgács, F. Z., Virág, I. C., Vad, A. M., Bytyqi, B., Buday, T., & Csajbók, J. (2023). Changes in Plant and Grain Quality of Winter Oat (Avena sativa L.) Varieties in Response to Silicon and Sulphur Foliar Fertilisation under Abiotic Stress Conditions. Plants, 12(4), 969. https://doi.org/10.3390/plants12040969