
Detection, Identification and Molecular Characterization of the 16SrII-V Subgroup Phytoplasma Strain Associated with Pisum sativum and Parthenium hysterophorus L.
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
2.1. Green Pea and Parthenium Weed Exhibited Typical Phytoplasma-Infected Symptoms
2.2. Identification of the Phytoplasma Associated with the Symptomatic Green Pea and Parthenium Weed
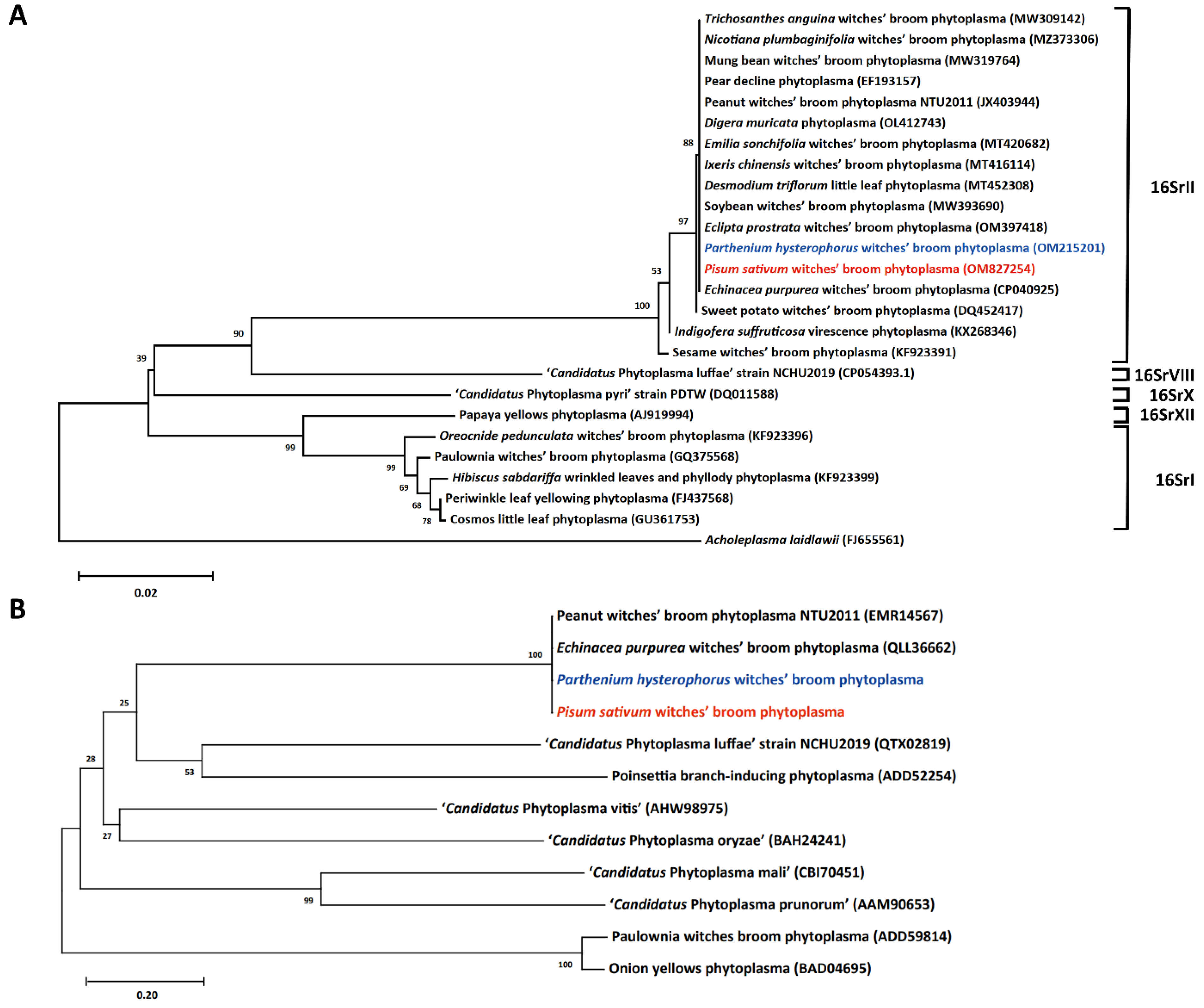
2.3. Classification of the Phytoplasma Associated with the Symptomatic Green Pea and Parthenium Weed
2.4. Identification of Putative Effectors Responsible for the Phyllody and Virescence Symptoms Associated with PsWB and PhWB Diseases
3. Materials and Methods
3.1. Field Sampling
3.2. Polymerase Chain Reaction (PCR) and Nested PCR
3.3. Transmission Electron Microscopy (TEM) Assay
3.4. Western Blotting
3.5. Phylogenetic Tree Construction
3.6. iPhyClassifier Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, I.-M.; Robert, E.D.; Gundersen-Rindal, D.E. Phytoplasma: Phytopathogenic Mollicutes. Annu. Rev. Microbiol. 2000, 54, 221–255. [Google Scholar] [CrossRef] [PubMed]
- Hogenhout, S.A.; Oshima, K.; Ammar, E.-D.; Kakizawa, S.; Kingdom, H.N.; Namba, S. Phytoplasmas: Bacteria that Manipulate Plants and Insects. Mol. Plant Pathol. 2008, 9, 403–423. [Google Scholar] [CrossRef] [PubMed]
- Trivellone, V.; Dietrich, C.H. Evolutionary Diversification in Insect Vector–Phytoplasma–Plant Associations. Ann. Entomol. Soc. Am. 2020, 114, 137–150. [Google Scholar] [CrossRef]
- Christensen, N.M.; Axelsen, K.B.; Nicolaisen, M.; Schulz, A. Phytoplasmas and Their Interactions with Hosts. Trends Plant Sci. 2005, 10, 526–535. [Google Scholar] [CrossRef]
- Namba, S. Molecular and Biological Properties of Phytoplasmas. Proc. Jpn. Acad. Ser. B Biol. Sci. 2019, 95, 401–418. [Google Scholar] [CrossRef]
- Doi, Y.; Teranaka, M.; Yora, K.; Asuyama, H. Mycoplasma- or PLT Group-like Microorganisms Found in the Phloem Elements of Plants Infected with Mulberry Dwarf, Potato Witches’ Broom, Aster Yellows, or Paulownia Witches’ Broom. Ann. Phytopathol. Soc. Jpn. 1967, 33, 259–266. [Google Scholar] [CrossRef]
- Tomkins, M.; Kliot, A.; Marée, A.F.M.; Hogenhout, S.A. A Multi-Layered Mechanistic Modelling Approach to Understand How Effector Genes Extend Beyond Phytoplasma to Modulate Plant Hosts, Insect Vectors and the Environment. Curr. Opin. Plant Biol. 2018, 44, 39–48. [Google Scholar] [CrossRef]
- Chang, S.H.; Tan, C.M.; Wu, C.-T.; Lin, T.-H.; Jiang, S.-Y.; Liu, R.-C.; Tsai, M.-C.; Su, L.-W.; Yang, J.-Y. Alterations of Plant Architecture and Phase Transition by the Phytoplasma Virulence Factor SAP11. J. Exp. Bot. 2018, 69, 5389–5401. [Google Scholar] [CrossRef]
- Hoshi, A.; Oshima, K.; Kakizawa, S.; Ishii, Y.; Ozeki, J.; Hashimoto, M.; Komatsu, K.; Kagiwada, S.; Yamaji, Y.; Namba, S. A Unique Virulence Factor for Proliferation and Dwarfism in Plants Identified from a Phytopathogenic Bacterium. Proc. Natl. Acad. Sci. USA 2009, 106, 6416–6421. [Google Scholar] [CrossRef]
- Huang, W.; MacLean, A.M.; Sugio, A.; Maqbool, A.; Busscher, M.; Cho, S.-T.; Kamoun, S.; Kuo, C.-H.; Immink, R.G.H.; Hogenhout, S.A. Parasitic Modulation of Host Development by Ubiquitin-Independent Protein Degradation. Cell 2021, 184, 5201–5214.e12. [Google Scholar] [CrossRef]
- MacLean, A.M.; Sugio, A.; Makarova, O.V.; Findlay, K.C.; Grieve, V.M.; Tóth, R.; Nicolaisen, M.; Hogenhout, S.A. Phytoplasma Effector SAP54 Induces Indeterminate Leaf-Like Flower Development in Arabidopsis Plants. Plant Physiol. 2011, 157, 831–841. [Google Scholar] [CrossRef] [PubMed]
- Maejima, K.; Iwai, R.; Himeno, M.; Komatsu, K.; Kitazawa, Y.; Fujita, N.; Ishikawa, K.; Fukuoka, M.; Minato, N.; Yamaji, Y.; et al. Recognition of Floral Homeotic MADS Domain Transcription Factors by a Phytoplasmal Effector, Phyllogen, Induces Phyllody. Plant J. 2014, 78, 541–554. [Google Scholar] [CrossRef] [PubMed]
- Sugio, A.; Kingdom, H.N.; MacLean, A.M.; Grieve, V.M.; Hogenhout, S.A. Phytoplasma Protein Effector SAP11 Enhances Insect Vector Reproduction by Manipulating Plant Development and Defense Hormone Biosynthesis. Proc. Natl. Acad. Sci. USA 2011, 108, E1254–E1263. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.M.; Li, C.-H.; Tsao, N.-W.; Su, L.-W.; Lu, Y.-T.; Chang, S.H.; Lin, Y.Y.; Liou, J.-C.; Hsieh, L.-C.; Yu, J.-Z.; et al. Phytoplasma SAP11 Alters 3-Isobutyl-2-Methoxypyrazine Biosynthesis in Nicotiana benthamiana by Suppressing NbOMT1. J. Exp. Bot. 2016, 67, 4415–4425. [Google Scholar] [CrossRef]
- Guindon, M.F.; Cazzola, F.; Palacios, T.; Gatti, I.; Bermejo, C.; Cointry, E. Biofortification of Pea (Pisum sativum L.): A Review. J. Sci. Food Agric. 2021, 101, 3551–3563. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.-H.; Chiang, C.-H.; Chang, C.-A. First Report of Lettuce Mosaic Virus Infecting Pea in Taiwan. Plant Dis. 2019, 103, 2971. [Google Scholar] [CrossRef]
- Wang, P.-H.; Lin, Y.-S. Ecology of Pea Wilt and Root Rot Pathogens in Drained Paddy Field. Plant Prot. Bull. 1985, 21, 317–324. [Google Scholar]
- Mao, R.; Shabbir, A.; Adkins, S. Parthenium hysterophorus: A Tale of Global Invasion Over Two Centuries, Spread and Prevention Measures. J. Environ. Manag. 2021, 279, 111751. [Google Scholar] [CrossRef]
- Ojija, F. Eco-Friendly Management of Parthenium hysterophorus. Sci. Prog. 2022, 105, 00368504221118234. [Google Scholar] [CrossRef]
- Kaur, L.; Malhi, D.S.; Cooper, R.; Kaur, M.; Sohal, H.S.; Mutreja, V.; Sharma, A. Comprehensive Review on Ethnobotanical Uses, Phytochemistry, Biological Potential and Toxicology of Parthenium hysterophorus L.: A Journey from Noxious Weed to a Therapeutic Medicinal Plant. J. Ethnopharmacol. 2021, 281, 114525. [Google Scholar] [CrossRef]
- Adkins, S.; Shabbir, A. Biology, Ecology and Management of the Invasive Parthenium Weed (Parthenium hysterophorus L.). Pest Manag. Sci. 2014, 70, 1023–1029. [Google Scholar] [CrossRef] [PubMed]
- Chung, W.-C.; Chen, L.-L.; Lo, W.-S.; Lin, C.-P.; Kuo, C.-H. Comparative Analysis of the Peanut Witches’-Broom Phytoplasma Genome Reveals Horizontal Transfer of Potential Mobile Units and Effectors. PLoS ONE 2013, 8, e62770. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-T.; Cho, S.-T.; Lin, Y.-C.; Tan, C.-M.; Chiu, Y.-C.; Yang, J.-Y.; Kuo, C.-H. Comparative Genome Analysis of ‘Candidatus Phytoplasma luffae’ Reveals the Influential Roles of Potential Mobile Units in Phytoplasma Evolution. Front. Microbiol. 2022, 13, 773608. [Google Scholar] [CrossRef] [PubMed]
- Bau, H.J.; Hung, S.C.; Chang, W.C.; Chen, Y.K. First Report of Group 16SrXII Phytoplasma Associated with Papaya Yellows in Taiwan. Plant Dis. 2011, 95, 1581. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.-L.; Chen, C.-C.; Lin, C.-P. Detection and Identification of the Phytoplasma Associated with Pear Decline in Taiwan. Eur. J. Plant Pathol. 2007, 117, 281–291. [Google Scholar] [CrossRef]
- Kumari, S.; Nagendran, K.; Rai, A.B.; Singh, B.; Rao, G.P.; Bertaccini, A. Global Status of Phytoplasma Diseases in Vegetable Crops. Front. Microbiol. 2019, 10, 1349. [Google Scholar] [CrossRef]
- Naik, D.V.K.; Reddy, B.V.B.; Rani, J.S.; Jayalakshmi Devi, R.S.; Prasad, K.V.H. Molecular Characterization of Phytoplasma Associated with Crops, Weeds and Forest Trees in Andhra Pradash, India. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 781–791. [Google Scholar] [CrossRef]
- Cai, H.; Wang, L.C.; Yang, Z.X.; Wan, Q.L.; Wei, W.; Davis, R.E.; Zhao, Y. Evidence for the Role of an Invasive Weed in Widespread Occurrence of Phytoplasma Diseases in Diverse Vegetable Crops: Implications from Lineage-Specific Molecular Markers. Crop Prot. 2016, 89, 193–201. [Google Scholar] [CrossRef]
- Tessema, T.; Obermeier, C.; Einhorn, G.; Seemüller, E.; Büttner, C. Phyllody Disease of Parthenium Weed in Ethiopia. Pest Mat. J. Ethiop. 2004, 8, 39–50. [Google Scholar]
- Tan, C.M.; Lin, Y.-C.; Li, J.-R.; Chien, Y.-Y.; Wang, C.-J.; Chou, L.; Wang, C.-W.; Chiu, Y.-C.; Kuo, C.-H.; Yang, J.-Y. Accelerating Complete Phytoplasma Genome Assembly by Immunoprecipitation-Based Enrichment and MinION-Based DNA Sequencing for Comparative Analyses. Front. Microbiol. 2021, 12, 766221. [Google Scholar] [CrossRef]
- Kakizawa, S.; Oshima, K.; Namba, S. Diversity and Functional Importance of Phytoplasma Membrane Proteins. Trends Microbiol. 2006, 14, 254–256. [Google Scholar] [CrossRef] [PubMed]
- Konnerth, A.; Krczal, G.; Boonrod, K. Immunodominant Membrane Proteins of Phytoplasmas. Microbiology 2016, 162, 1267–1273. [Google Scholar] [CrossRef] [PubMed]
- Iwabuchi, N.; Kitazawa, Y.; Maejima, K.; Koinuma, H.; Miyazaki, A.; Matsumoto, O.; Suzuki, T.; Nijo, T.; Oshima, K.; Namba, S.; et al. Functional Variation in Phyllogen, a Phyllody-Inducing Phytoplasma Effector Family, Attributable to a Single Amino Acid Polymorphism. Mol. Plant Pathol. 2020, 21, 1322–1336. [Google Scholar] [CrossRef] [PubMed]
- Pecher, P.; Moro, G.; Canale, M.C.; Capdevielle, S.; Singh, A.; MacLean, A.; Sugio, A.; Kuo, C.-H.; Lopes, J.R.S.; Hogenhout, S.A. Phytoplasma SAP11 Effector Destabilization of TCP Transcription Factors Differentially Impact Development and Defence of Arabidopsis versus Maize. PLoS Pathog. 2019, 15, e1008035. [Google Scholar] [CrossRef]
- Lee, I.-M.; Hammond, R.W.; Davis, R.E.; Gundersen, D.E. Universal Amplification and Analysis of Pathogen 16S rDNA for Classification and Identification of Mycoplasmalike Organisms. Phytopathology 1993, 83, 834–842. [Google Scholar] [CrossRef]
- Deng, S.; Hiruki, C. Amplification of 16S rRNA Genes from Culturable and Nonculturable Mollicutes. J. Microbiol. Methods 1991, 14, 53–61. [Google Scholar] [CrossRef]
- Schneider, B.; Seemüller, E.; Smart, C.; Kirkpatrick, B. Phylogenetic classification of plant pathogenic mycoplasma-like organisms or phytoplasmas. In Molecular and Diagnostic Procedures in Mycoplasmology; Razin, S., Tully, J., Eds.; Academic Press: San Diego, CA, USA, 1995; Volume 1, pp. 369–380. [Google Scholar]
- Gundersen, D.; Lee, I.-M. Ultrasensitive Detection of Phytoplasmas by Nested-PCR Assays Using Two Universal Primer Pairs. Phytopathol. Mediterr. 1996, 35, 144–151. [Google Scholar]
- Chien, Y.-Y.; Tan, C.M.; Kung, Y.-C.; Lee, Y.-C.; Chiu, Y.-C.; Yang, J.-Y. Lilac Tasselflower (Emilia sonchifolia) is a New Host for Peanut Witches’ Broom Phytoplasma, a 16SrII-V Subgroup Strain in Taiwan. Plant Dis. 2020, 105, 211. [Google Scholar] [CrossRef]
- Buxa, S.V.; Degola, F.; Polizzotto, R.; De Marco, F.; Loschi, A.; Kogel, K.-H.; di Toppi, L.S.; van Bel, A.J.E.; Musetti, R. Phytoplasma Infection in Tomato is Associated with Re-Organization of Plasma Membrane, ER Stacks, and Actin Filaments in Sieve Elements. Front. Plant Sci. 2015, 6, 650. [Google Scholar] [CrossRef]
- Hu, S.-F.; Huang, Y.-H.; Lin, C.-P.; Liu, L.-Y.D.; Hong, S.-F.; Yang, C.-Y.; Lo, H.-F.; Tseng, T.-Y.; Chen, W.-Y.; Lin, S.-S. Development of a Mild Viral Expression System for Gain-Of-Function Study of Phytoplasma Effector In Planta. PLoS ONE 2015, 10, e0130139. [Google Scholar] [CrossRef]
- Zhao, Y.; Wei, W.; Lee, I.-M.; Shao, J.; Suo, X.; Davis, R.E. The iPhyClassifier, an Interactive Online Tool for Phytoplasma Classification and Taxonomic Assignment. In Phytoplasma: Methods and Protocols; Dickinson, M., Hodgetts, J., Eds.; Humana Press: Totowa, NJ, USA, 2013; pp. 329–338. [Google Scholar]
- Liao, P.-Q.; Chiu, Y.-C.; Mejia, H.M.; Tan, C.M.; Chen, Y.-K.; Yang, J.-Y. First Report of ‘Candidatus Phytoplasma aurantifolia’ Associated with the Invasive Weed Eclipta prostrata (L.) in Taiwan. Plant Dis. 2022. [Google Scholar] [CrossRef] [PubMed]
- Chien, Y.-Y.; Tan, C.M.; Kung, Y.-C.; Lee, Y.-C.; Chiu, Y.-C.; Yang, J.-Y. Ixeris chinensis is a New Host for Peanut Witches’ Broom Phytoplasma, a 16SrII-V Subgroup Strain, in Taiwan. Plant Dis. 2021, 105, 210. [Google Scholar] [CrossRef] [PubMed]
- Mejia, H.M.; Liao, P.-Q.; Chen, Y.-K.; Lee, Y.-C.; Tan, C.M.; Chiu, Y.-C.; Yang, J.-Y. Detection, Identification and Molecular Characterization of the 16SrII-V Subgroup Phytoplasma Strain Associated with Digera muricata (L.) Mart. in Taiwan. Plant Dis. 2022, 106, 1788–1792. [Google Scholar] [CrossRef]
- Liao, P.-Q.; Chen, Y.-K.; Mejia, H.M.; Chien, Y.-Y.; Lee, Y.-C.; Tan, C.M.; Chiu, Y.-C.; Yang, J.-Y. Detection, Identification and Molecular Characterization of a 16SrII-V Subgroup Phytoplasma Associated with Nicotiana plumbaginifolia Viviani. Plant Dis. 2021, 106, 805–809. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiu, Y.-C.; Liao, P.-Q.; Mejia, H.M.; Lee, Y.-C.; Chen, Y.-K.; Yang, J.-Y. Detection, Identification and Molecular Characterization of the 16SrII-V Subgroup Phytoplasma Strain Associated with Pisum sativum and Parthenium hysterophorus L. Plants 2023, 12, 891. https://doi.org/10.3390/plants12040891
Chiu Y-C, Liao P-Q, Mejia HM, Lee Y-C, Chen Y-K, Yang J-Y. Detection, Identification and Molecular Characterization of the 16SrII-V Subgroup Phytoplasma Strain Associated with Pisum sativum and Parthenium hysterophorus L. Plants. 2023; 12(4):891. https://doi.org/10.3390/plants12040891
Chicago/Turabian StyleChiu, Yi-Ching, Pei-Qing Liao, Helen Mae Mejia, Ya-Chien Lee, Yuh-Kun Chen, and Jun-Yi Yang. 2023. "Detection, Identification and Molecular Characterization of the 16SrII-V Subgroup Phytoplasma Strain Associated with Pisum sativum and Parthenium hysterophorus L." Plants 12, no. 4: 891. https://doi.org/10.3390/plants12040891
APA StyleChiu, Y.-C., Liao, P.-Q., Mejia, H. M., Lee, Y.-C., Chen, Y.-K., & Yang, J.-Y. (2023). Detection, Identification and Molecular Characterization of the 16SrII-V Subgroup Phytoplasma Strain Associated with Pisum sativum and Parthenium hysterophorus L. Plants, 12(4), 891. https://doi.org/10.3390/plants12040891