
Ruta angustifolia Pers. (Narrow-Leaved Fringed Rue): Pharmacological Properties and Phytochemical Profile


Abstract
1. Introduction
2. Pharmacological Activities of R. angustifolia Extracts
3. Bioactive Phytochemicals from R. angustifolia Pers.
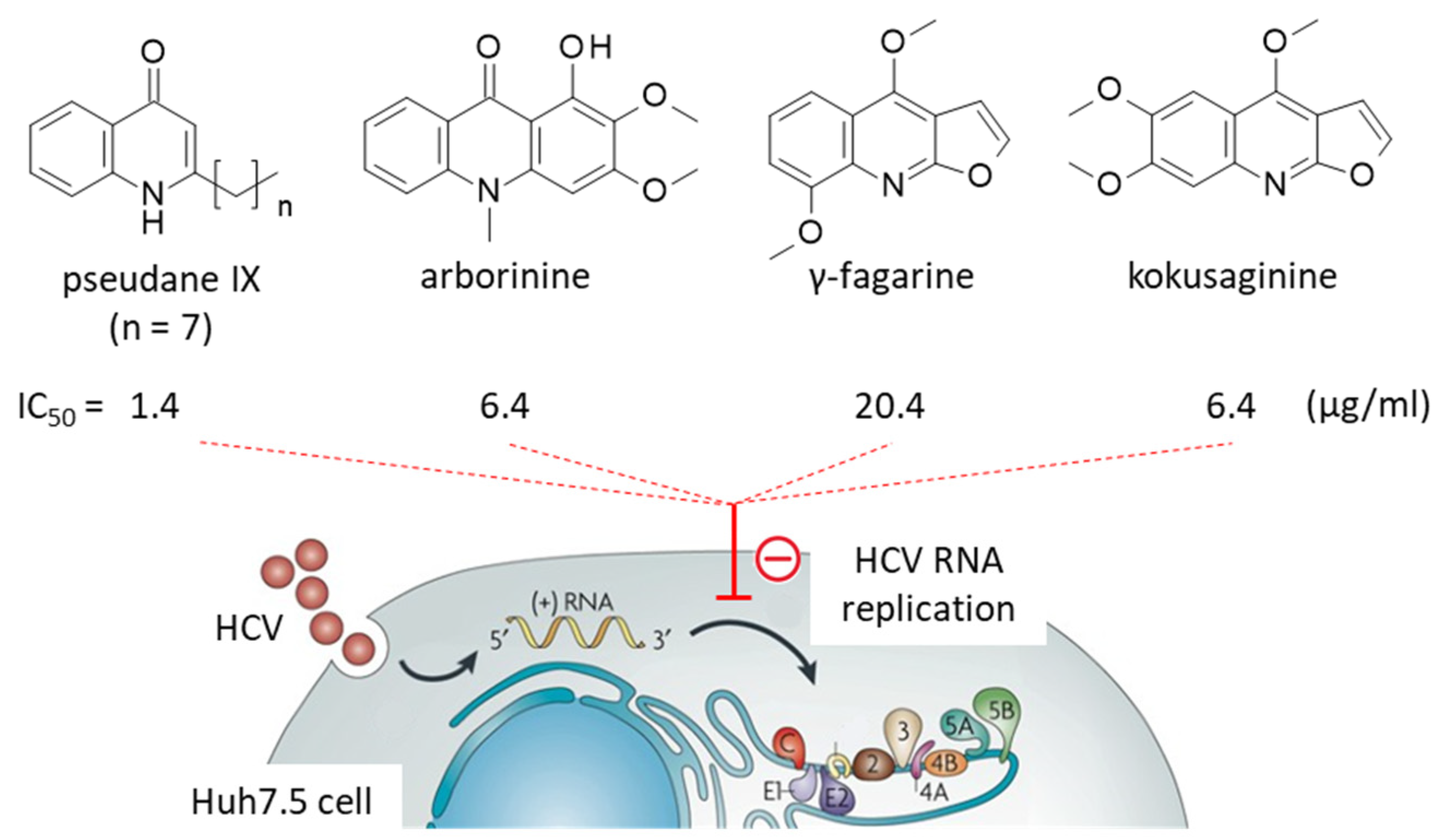
3.1. Quinoline Alkaloids
3.2. Furocoumarins
3.3. Other Compounds
4. Discussion
Funding
Data Availability Statement
Conflicts of Interest
References
- Appelhans, M.S.; Bayly, M.J.; Heslewood, M.M.; Groppo, M.; Verboom, A.; Forster, P.I.; Kallunki, J.A.; Duretto, M.F. A new subfamily classification of the Citrus family (Rutaceae) based on six nuclear and plastid markers. Taxon 2021, 70, 1035–1061. [Google Scholar] [CrossRef]
- Gao, L.; Zhang, H.; Yuan, C.H.; Zeng, L.H.; Xiang, Z.; Song, J.F.; Wang, H.G.; Jiang, J.P. Citrus aurantium ‘Changshan-huyou’—An ethnopharmacological and phytochemical review. Front. Pharmacol. 2022, 13, 983470. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Wang, P.; Pang, X.; Zhang, L.; Qian, L.; Jia, X.; Chen, W.; Ruan, S.; Sun, L. The new exploration of pure total flavonoids extracted from Citrus maxima (Burm.) Merr. as a new therapeutic agent to bring health benefits for people. Front. Nutr. 2022, 9, 958329. [Google Scholar] [CrossRef] [PubMed]
- Liang, P.L.; Chen, X.L.; Gong, M.J.; Xu, Y.; Tu, H.S.; Zhang, L.; Liao, B.S.; Qiu, X.H.; Zhang, J.; Huang, Z.H.; et al. Guang Chen Pi (the pericarp of Citrus reticulata Blanco’s cultivars ‘Chachi’) inhibits macrophage-derived foam cell formation. J. Ethnopharmacol. 2022, 293, 115328. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Pu, Z.J.; Zhang, Z.Y.; Zhou, G.S.; Zou, D.Q.; Guo, S.; Li, C.; Zhan, Z.L.; Duan, J.A. Research on Biomarkers of Different Growth Periods and Different Drying Processes of Citrus wilsonii Tanaka Based on Plant Metabolomics. Front. Plant. Sci. 2021, 12, 700367. [Google Scholar] [CrossRef]
- Nahar, L.; El-Seedi, H.R.; Khalifa, S.A.M.; Mohammadhosseini, M.; Sarker, S.D. Ruta Essential Oils: Composition and Bioactivities. Molecules 2021, 26, 4766. [Google Scholar] [CrossRef] [PubMed]
- Keswara, Y.D.; Handayani, S.R. Uji Aktivitas Analgetik Ekstrak Etanol Daun Inggu (Ruta angustifolia [L.] Pers) Pada Tikus Putih Jantan. J. Syifa. Sci. Clin. Res. 2019, 1, 57–69. [Google Scholar] [CrossRef]
- Noer, S.; Abinawanto, A.; Basith, A. Molecular identification of the medicinal herb plant Ruta angustifolia from Lembang Bandung using chloroplast DNA markers. AIP Conf. Proc. 2018, 2023, 020113. [Google Scholar] [CrossRef]
- Coimbra, A.T.; Ferreira, S.; Duarte, A.P. Genus Ruta: A natural source of high value products with biological and pharmacological properties. J. Ethnopharmacol. 2020, 260, 113076. [Google Scholar] [CrossRef]
- Pollio, A.; De Natale, A.; Appetiti, E.; Aliotta, G.; Touwaide, A. Continuity and change in the Mediterranean medical tradition: Ruta spp. (rutaceae) in Hippocratic medicine and present practices. J. Ethnopharmacol. 2008, 116, 469–482. [Google Scholar] [CrossRef]
- Shahrajabian, M.H. A Candidate for Health Promotion, Disease Prevention and Treatment, Common Rue (Ruta graveolens L.), an Important Medicinal plant in Traditional Medicine. Curr. Rev. Clin. Exp. Pharmacol. 2022; online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Jiao, M.; Liu, X.; Ren, Y.; Wang, Y.; Cheng, L.; Liang, Y.; Li, Y.; Zhang, T.; Wang, W.; Mei, Z. Comparison of Herbal Medicines Used for Women’s Menstruation Diseases in Different Areas of the World. Front. Pharmacol. 2022, 12, 751207. [Google Scholar] [CrossRef] [PubMed]
- Richardson, J.S.; Sethi, G.; Lee, G.S.; Malek, S.N. Chalepin: Isolated from Ruta angustifolia L. Pers induces mitochondrial mediated apoptosis in lung carcinoma cells. BMC Complement. Altern. Med. 2016, 16, 389. [Google Scholar] [CrossRef]
- Kamal, L.Z.M.; Hassan, N.M.; Taib, N.M.; Soe, M. Graveoline from Ruta angustifolia (L.) Pers. and Its Antimicrobial Synergistic Potential in Erythromycin or Vancomycin Combinations. Sains Malays. 2018, 47, 2429–2435. [Google Scholar] [CrossRef]
- Shuib, N.A.; Iqbal, A.; Sulaiman, F.A.; Razak, I.; Susanti, D. Antioxidant and antibacterial activities of Ruta angustifolia extract. J. Teknol. 2015, 77, 101–105. [Google Scholar] [CrossRef]
- Haddouchi, F.; Chaouche, T.M.; Zaouali, Y.; Ksouri, R.; Attou, A.; Benmansour, A. Chemical composition and antimicrobial activity of the essential oils from four Ruta species growing in Algeria. Food Chem. 2013, 141, 253–258. [Google Scholar] [CrossRef]
- Dob, T.; Dahmane, D.; Gauriat-Desrdy, B.; Daligault, V. Volatile Constituents of the Essential Oil of Ruta chalepensis L. subsp. Angustifolia (Pers.) P. Cout. J. Essent. Oil Res. 2008, 20, 306–309. [Google Scholar] [CrossRef]
- Bennaoum, Z.; Benhassaini, H.; Falconieri, D.; Piras, A.; Porcedda, S. Chemical variability in essential oils from Ruta species among seasons, and its taxonomic and ecological significance. Nat. Prod. Res. 2017, 31, 2329–2334. [Google Scholar] [CrossRef]
- Kamal, L.Z.M.; Adam, M.A.A.; Shahpudin, S.N.M.; Shuib, A.N.; Sandai, R.; Hassan, N.M.; Tabana, Y.; Basri, D.F.; Than, L.T.L.; Sandai, D. Identification of Alkaloid Compounds Arborinine and Graveoline from Ruta angustifolia (L.) Pers for their Antifungal Potential against Isocitrate lyase (ICL1) gene of Candida albicans. Mycopathologia 2021, 186, 221–236. [Google Scholar] [CrossRef]
- Noer, S.; Abinawanto, A.; Bachtiar, B.M.; Bowolaksono, A. Inhibition of Candida albicans hypha formation in biofilm formation by Ruta angustifolia extract. AIP Conf. Proc. 2021, 2331, 050003. [Google Scholar] [CrossRef]
- Wahyuni, T.S.; Widyawaruyanti, A.; Lusida, M.I.; Fuad, A.; Soetjipto; Fuchino, H.; Kawahara, N.; Hayashi, Y.; Aoki, C.; Hotta, H. Inhibition of hepatitis C virus replication by chalepin and pseudane IX isolated from Ruta angustifolia leaves. Fitoterapia 2014, 99, 276–283. [Google Scholar] [CrossRef]
- González, J.A.; García-Barriuso, M.; Gordaliza, M.; Amich, F. Traditional plant-based remedies to control insect vectors of disease in the Arribes del Duero (western Spain): An ethnobotanical study. J. Ethnopharmacol. 2011, 138, 595–601. [Google Scholar] [CrossRef]
- Salama, M.G.M.; Mead, H.M.I.; Sabry, H.M.; Abo-Ghalia, A.H.A.; Semida, F.M.M. Biocidal effect of Ergosterol-Propyl Ester Isolated from Ruta angustifolia (Pers.) on Spodopetra littoralis (Boisd). Catrina 2020, 21, 37–45. [Google Scholar] [CrossRef]
- Tampe, J.; Parra, L.; Huaiquil, K.; Quiroz, A. Potential repellent activity of the essential oil of ruta chalepensis (linnaeus) from chile against aegorhinus superciliosus (guérin) (Coleoptera: Curculionidae). J. Soil Sci. Plant Nutr. 2016, 16, 48–59. [Google Scholar] [CrossRef]
- Najem, M.; Bammou, M.; Bachiri, L.; Bouiamrine, E.H.; Ibijbijen, J.; Nassiri, L. Ruta chalepensis L. Essential Oil Has a Biological Potential for a Natural Fight against the Pest of Stored Foodstuffs: Tribolium castaneum Herbst. Evid. Based Complement. Alternat. Med. 2020, 2020, 5739786. [Google Scholar] [CrossRef] [PubMed]
- Ortu, E.; Sanna, G.; Scala, A.; Pulina, G.; Caboni, P.; Battacone, G. In vitro anthelmintic activity of active compounds of the fringed rue Ruta chalepensis against dairy ewe gastrointestinal nematodes. J. Helminthol. 2017, 91, 447–453. [Google Scholar] [CrossRef]
- Ntalli, N.G.; Manconi, F.; Leonti, M.; Maxia, A.; Caboni, P. Aliphatic ketones from Ruta chalepensis (Rutaceae) induce paralysis on root knot nematodes. J. Agric. Food Chem. 2011, 59, 7098–7103. [Google Scholar] [CrossRef]
- Faria, J.M.; Barbosa, P.; Bennett, R.N.; Mota, M.; Figueiredo, A.C. Bioactivity against Bursaphelenchus xylophilus: Nematotoxics from essential oils, essential oils fractions and decoction waters. Phytochemistry 2013, 94, 220–228. [Google Scholar] [CrossRef]
- Vasudevan, T.N.; Luckner, M. Alkaloids from Ruta angustifolia Pers., Ruta chalepensis L., Ruta graveolens L. and Ruta montana Mill. Pharmazie 1968, 23, 520–521. [Google Scholar]
- Adamska-Szewczyk, A.; Glowniak, K.; Baj, T. Furochinoline alkaloids in plants from Rutaceae family—A review. Curr. Issues Pharm. Med. Sci. 2016, 29, 33–38. [Google Scholar] [CrossRef]
- Kim, W.J.; Kim, Y.O.; Kim, J.H.; Nam, B.H.; Kim, D.G.; An, C.M.; Lee, J.S.; Kim, P.S.; Lee, H.M.; Oh, J.S.; et al. Liquid Chromatography-Mass Spectrometry-Based Rapid Secondary-Metabolite Profiling of Marine Pseudoalteromonas sp. M2. Mar. Drugs 2016, 14, 24. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.E.; Jung, I.; Lee, J.S.; Na, J.Y.; Kim, W.J.; Kim, Y.O.; Park, Y.D.; Lee, J.S. Pseudane-VII Isolated from Pseudoalteromonas sp. M2 Ameliorates LPS-Induced Inflammatory Response In Vitro and In Vivo. Mar. Drugs 2017, 15, 336. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.E.; Jung, I.; Na, J.Y.; Lee, Y.; Lee, J.; Lee, J.S.; Lee, J.S. Pseudane-VII Regulates LPS-Induced Neuroinflammation in Brain Microglia Cells through the Inhibition of iNOS Expression. Molecules 2018, 23, 3196. [Google Scholar] [CrossRef]
- Ritter, C.; Luckner, M. Biosynthesis of 2-n-alkyl-4-hydroxyquinoline derivates (pseudane) in Pseudomonas aeruginosa. Eur. J. Biochem. 1971, 18, 391–400. [Google Scholar] [CrossRef]
- Bandatmakuru, S.R.; Arava, V.R. Novel synthesis of graveoline and graveolinine. Synth. Commun. 2018, 8, 2635–2641. [Google Scholar] [CrossRef]
- Singh, S.; Nerella, S.; Pabbaraja, S.; Mehta, G. Access to 2-Alkyl/Aryl-4-(1H)-Quinolones via Orthogonal “NH3” Insertion into o -Haloaryl Ynones: Total Synthesis of Bioactive Pseudanes, Graveoline, Graveolinine, and Waltherione, F. Org. Lett. 2020, 22, 1575–1579. [Google Scholar] [CrossRef]
- Chen, H.; Li, S.; Wang, S.; Li, W.; Bao, N.; Ai, W. The inhibitory effect of kokusaginine on the growth of human breast cancer cells and MDR-resistant cells is mediated by the inhibition of tubulin assembly. Bioorg. Med. Chem. Lett. 2018, 28, 2490–2492. [Google Scholar] [CrossRef]
- Sichaem, J.; Rojpitikul, T.; Sawasdee, P.; Lugsannangarm, K.; Santi, T.P. Furoquinoline Alkaloids from the Leaves of Evodia lepta as Potential Cholinesterase Inhibitors and their Molecular Docking. Nat. Prod. Commun. 2015, 10, 1359–1362. [Google Scholar] [CrossRef] [PubMed]
- Senol Deniz, F.S.; Eren, G.; Orhan, I.E.; Sener, B.; Ozgen, U.; Aldaba, R.; Calis, I. Outlining In Vitro and In Silico Cholinesterase Inhibitory Activity of Twenty-Four Natural Products of Various Chemical Classes: Smilagenin, Kokusaginine, and Methyl Rosmarinate as Emboldening Inhibitors. Molecules 2021, 26, 2024. [Google Scholar] [CrossRef]
- Belén Valdez, M.; Bernal Giménez, D.M.; Fernández, L.R.; Musikant, A.D.; Ferri, G.; Sáenz, D.; Di Venosa, G.; Casas, A.; Avigliano, E.; Edreira, M.M.; et al. Antiparasitic Derivatives of the Furoquinoline Alkaloids Kokusaginine And Flindersiamine. ChemMedChem 2022, 17, e202100784. [Google Scholar] [CrossRef]
- Zuo, Y.; Pu, J.; Chen, G.; Shen, W.; Wang, B. Study on the activity and mechanism of skimmianine against human non-small cell lung cancer. Nat. Prod. Res. 2019, 33, 759–762. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Kang, L.; Shi, S.M.; Li, B.J.; Zhang, Y.; Zhang, X.Z.; Guo, X.W.; Fu, G.; Zheng, G.N.; Hao, H.; et al. Skimmianine as a novel therapeutic agent suppresses proliferation and migration of human esophageal squamous cell carcinoma via blocking the activation of ERK1/2. Neoplasma 2022, 69, 571–582. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Q.; Xuan, W.L.; Huang, Y.; Ren, S.; Wulan, T.Y.; Song, Y.; Xue, D.B.; Zhang, W.H. Dictamnine inhibits pancreatic cancer cell growth and epithelial-mesenchymal transition by blocking the PI3K/AKT signaling pathway. Neoplasma 2022, 69, 603–619. [Google Scholar] [CrossRef]
- El Sayed, K.; Al-Said, M.S.; El-Feraly, F.S.; Ross, S.A. New quinoline alkaloids from Ruta chalepensis. J. Nat. Prod. 2000, 63, 995–997. [Google Scholar] [CrossRef]
- Hale, A.L.; Meepagala, K.M.; Oliva, A.; Aliotta, G.; Duke, S.O. Phytotoxins from the leaves of Ruta graveolens. J. Agric. Food Chem. 2004, 52, 3345–3349. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, O.M.; Vieira, L.C.C.; Bellete, B.S.; King-Diaz, B.; Lotina-Hennsen, B.; da Silva, M.F.D.G.F.; Veiga, T.A.M. Evaluation of Alkaloids Isolated from Ruta graveolens as Photosynthesis Inhibitors. Molecules 2018, 23, 2693. [Google Scholar] [CrossRef]
- Ghosh, S.; Bishayee, K.; Khuda-Bukhsh, A.R. Graveoline isolated from ethanolic extract of Ruta graveolens triggers apoptosis and autophagy in skin melanoma cells: A novel apoptosis-independent autophagic signaling pathway. Phytother. Res. 2014, 28, 1153–1162. [Google Scholar] [CrossRef]
- Liu, H.; Guo, S.; Xi, S. A high-resolution accurate mass approach to identification of graveoline metabolites using ultra-high-performance liquid chromatography combined with a photo diode array detector and quadrupole/time-of-flight tandem mass spectrometry. Biomed. Chromatogr. 2022, 37, e5511. [Google Scholar] [CrossRef]
- An, Z.Y.; Yan, Y.Y.; Peng, D.; Ou, T.M.; Tan, J.H.; Huang, S.L.; An, L.K.; Gu, L.Q.; Huang, Z.S. Synthesis and evaluation of graveoline and graveolinine derivatives with potent anti-angiogenesis activities. Eur. J. Med. Chem. 2010, 45, 3895–3903. [Google Scholar] [CrossRef]
- Luo, W.; Lv, J.W.; Wang, T.; Zhang, Z.Y.; Guo, H.Y.; Song, Z.Y.; Wang, C.J.; Ma, J.; Chen, Y.P. Synthesis, in vitro and in vivo biological evaluation of novel graveolinine derivatives as potential anti-Alzheimer agents. Bioorg. Med. Chem. 2020, 28, 115190. [Google Scholar] [CrossRef]
- Rodrigues, T.; Reker, D.; Kunze, J.; Schneider, P.; Schneider, G. Revealing the Macromolecular Targets of Fragment-Like Natural Products. Angew. Chem. Int. Ed. Engl. 2015, 54, 10516–10520. [Google Scholar] [CrossRef] [PubMed]
- Ombito, J.O.; Chi, G.F.; Wansi, J.D. Ethnomedicinal uses, phytochemistry, and pharmacology of the genus Vepris (Rutaceae): A review. J. Ethnopharmacol. 2021, 267, 113622. [Google Scholar] [CrossRef] [PubMed]
- Langat, M.K.; Kami, T.; Cheek, M. Chemistry, taxonomy and ecology of the potentially chimpanzee-dispersed Vepris teva sp.nov. (Rutaceae) endangered in coastal thicket in the Congo Republic. PeerJ 2022, 10, e13926. [Google Scholar] [CrossRef] [PubMed]
- Noulala, C.G.T.; Ouete, J.L.N.; Atangana, A.F.; Mbahbou, G.T.B.; Fotso, G.W.; Stammler, H.G.; Lenta, B.N.; Happi, E.N.; Sewald, N.; Ngadjui, B.T. Soyauxinine, a New Indolopyridoquinazoline Alkaloid from the Stem Bark of Araliopsis soyauxii Engl. (Rutaceae). Molecules 2022, 27, 1104. [Google Scholar] [CrossRef] [PubMed]
- Al-Majmaie, S.; Nahar, L.; Rahman, M.M.; Nath, S.; Saha, P.; Talukdar, A.D.; Sharples, G.P.; Sarker, S.D. Anti-MRSA Constituents from Ruta chalepensis (Rutaceae) Grown in Iraq, and In Silico Studies on Two of Most Active Compounds, Chalepensin and 6-Hydroxy-rutin 3′,7-Dimethyl ether. Molecules 2021, 26, 1114. [Google Scholar] [CrossRef]
- Réthy, B.; Zupkó, I.; Minorics, R.; Hohmann, J.; Ocsovszki, I.; Falkay, G. Investigation of cytotoxic activity on human cancer cell lines of arborinine and furanoacridones isolated from Ruta graveolens. Planta Med. 2007, 73, 41–48. [Google Scholar] [CrossRef]
- Réthy, B.; Hohmann, J.; Minorics, R.; Varga, A.; Ocsovszki, I.; Molnár, J.; Juhász, K.; Falkay, G.; Zupkó, I. Antitumour properties of acridone alkaloids on a murine lymphoma cell line. Anticancer Res. 2008, 28, 2737–2743. [Google Scholar]
- Kuete, V.; Sandjo, L.P.; Seukep, J.A.; Zeino, M.; Mbaveng, A.T.; Ngadjui, B.; Efferth, T. Cytotoxic compounds from the fruits of Uapaca togoensis towards multifactorial drug-resistant cancer cells. Planta Med. 2015, 81, 32–38. [Google Scholar] [CrossRef]
- Piboonprai, K.; Khumkhrong, P.; Khongkow, M.; Yata, T.; Ruangrungsi, N.; Chansriniyom, C.; Iempridee, T. Anticancer activity of arborinine from Glycosmis parva leaf extract in human cervical cancer cells. Biochem. Biophys. Res. Commun. 2018, 500, 866–872. [Google Scholar] [CrossRef]
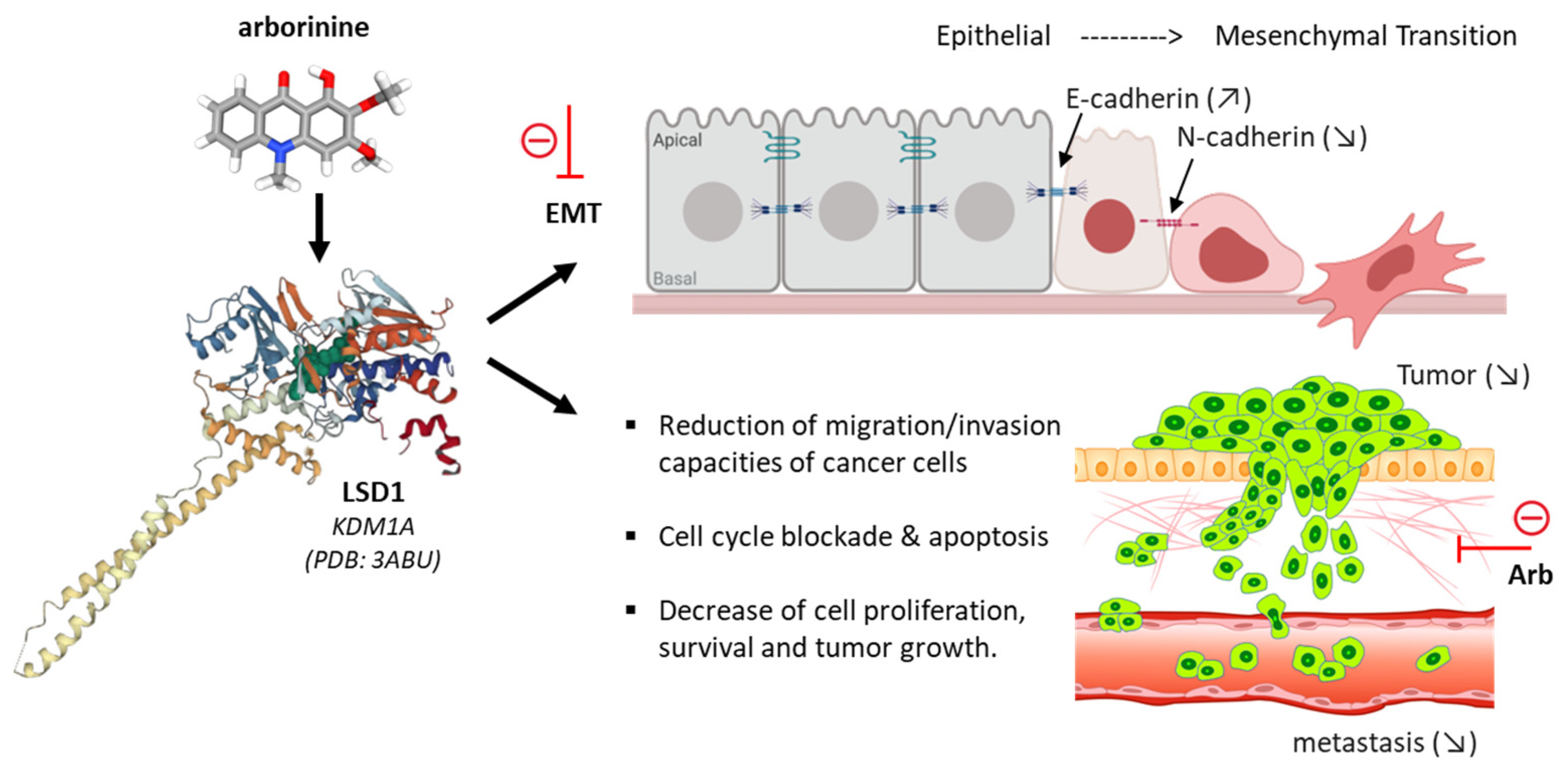
- Chu, Y.; Xiao, Z.; Jing, N.; Yan, W.; Wang, S.; Ma, B.; Zhang, J.; Li, Y. Arborinine, a potential LSD1 inhibitor, inhibits epithelial-mesenchymal transition of SGC-7901 cells and adriamycin-resistant gastric cancer SGC-7901/ADR cells. Investig. New Drugs 2021, 39, 627–635. [Google Scholar] [CrossRef]
- Li, N.; Yang, L.; Zuo, H. Arborinine suppresses ovarian cancer development through inhibition of LSD1. Life Sci. 2022, 291, 120275. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Gong, L.; Wang, J. Arborinine from Glycosmis parva leaf extract inhibits clear-cell renal cell carcinoma by inhibiting KDM1A/UBE2O signaling. Food Nutr. Res. 2022, 66, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Mehndiratta, S.; Liou, J.P. Histone lysine specific demethylase 1 inhibitors. RSC Med. Chem. 2020, 11, 969–981. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, X.; Wu, T.; Yin, W.; Yan, J.; Sun, Y.; Zhao, D. Therapeutic potential of targeting LSD1/ KDM1A in cancers. Pharmacol. Res. 2022, 175, 105958. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.C.; Liu, Y.J.; Gao, Y.; Wang, B.; Liu, H.M. An Update of Lysine Specific Demethylase 1 Inhibitor: A Patent Review (2016–2020). Recent Pat. Anticancer Drug Discov. 2022, 17, 9–25. [Google Scholar]
- Gill, H. Lysine-Specific Demethylase 1 (LSD1/KDM1A) Inhibition as a Target for Disease Modification in Myelofibrosis. Cells 2022, 11, 2107. [Google Scholar] [CrossRef]
- Rapanelli, M.; Williams, J.B.; Ma, K.; Yang, F.; Zhong, P.; Patel, R.; Kumar, M.; Qin, L.; Rein, B.; Wang, Z.J.; et al. Targeting histone demethylase LSD1 for treatment of deficits in autism mouse models. Mol. Psychiatry 2022, 27, 3355–3366. [Google Scholar] [CrossRef]
- Maiques-Diaz, A.; Somervaille, T.C. LSD1: Biologic roles and therapeutic targeting. Epigenomics 2016, 8, 1103–1116. [Google Scholar] [CrossRef]
- Mullard, A. Merck & Co. pays US$1.35 billion for LSD1 inhibitors. Nat. Rev. Drug Discov. 2022; online ahead of print. [Google Scholar] [CrossRef]
- Pal, C.; Kundu, M.K.; Bandyopadhyay, U.; Adhikari, S. Synthesis of novel heme-interacting acridone derivatives to prevent free heme-mediated protein oxidation and degradation. Bioorg. Med. Chem. Lett. 2011, 21, 3563–3567. [Google Scholar] [CrossRef]
- Laprasert, C.; Chansriniyom, C.; Limpanasithikul, W. S-deoxydihydroglyparvin from Glycosmis parva inhibits lipopolysaccharide induced murine macrophage activation through inactivating p38 mitogen activated protein kinase. J. Adv. Pharm. Technol. Res. 2021, 12, 32–39. [Google Scholar]
- Suhaimi, S.A.; Hong, S.L.; Abdul Malek, S.N. Rutamarin, an Active Constituent from Ruta angustifolia Pers., Induced Apoptotic Cell Death in the HT29 Colon Adenocarcinoma Cell Line. Pharmacogn. Mag. 2017, 13, S179–S188. [Google Scholar] [PubMed]
- Kozioł, E.; Luca, S.V.; Ağalar, H.G.; Sağlık, B.N.; Demirci, F.; Marcourt, L.; Wolfender, J.L.; Jóźwiak, K.; Skalicka-Woźniak, K. Rutamarin: Efficient Liquid-Liquid Chromatographic Isolation from Ruta graveolens L. and Evaluation of Its In Vitro and In Silico MAO-B Inhibitory Activity. Molecules 2020, 25, 2678. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Wang, L.; González-Molleda, L.; Wang, Y.; Xu, J.; Yuan, Y. Antiviral activity of (+)-rutamarin against Kaposi’s sarcoma-associated herpesvirus by inhibition of the catalytic activity of human topoisomerase II. Antimicrob. Agents Chemother. 2014, 58, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Bazaldúa-Rodríguez, A.F.; Quintanilla-Licea, R.; Verde-Star, M.J.; Hernández-García, M.E.; Vargas-Villarreal, J.; Garza-González, J.N. Furanocoumarins from Ruta chalepensis with Amebicide Activity. Molecules 2021, 26, 3684. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Wang, Y.; Yuan, Y. Antiviral activity of topoisomerase II catalytic inhibitors against Epstein-Barr virus. Antiviral Res. 2014, 107, 95–101. [Google Scholar] [CrossRef]
- Lin, Y.; Wang, Q.; Gu, Q.; Zhang, H.; Jiang, C.; Hu, J.; Wang, Y.; Yan, Y.; Xu, J. Semisynthesis of (-)-Rutamarin Derivatives and Their Inhibitory Activity on Epstein-Barr Virus Lytic Replication. J. Nat. Prod. 2017, 80, 53–60. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, H.; Yao, X.G.; Shen, H.; Chen, J.; Li, C.; Chen, L.; Zheng, M.; Ye, J.; Hu, L.; et al. (+)-Rutamarin as a dual inducer of both GLUT4 translocation and expression efficiently ameliorates glucose homeostasis in insulin-resistant mice. PLoS ONE 2012, 7, e31811. [Google Scholar] [CrossRef]
- Orlita, A.; Sidwa-Gorycka, M.; Kumirska, J.; Maliński, E.; Siedlecka, E.M.; Gajdus, J.; Lojkowska, E.; Stepnowski, P. Identification of Ruta graveolens L. metabolites accumulated in the presence of abiotic elicitors. Biotechnol. Prog. 2008, 24, 128–133. [Google Scholar] [CrossRef]
- Tamene, D.; Endale, M. Antibacterial Activity of Coumarins and Carbazole Alkaloid from Roots of Clausena anisata. Adv. Pharmacol. Sci. 2019, 2019, 5419854. [Google Scholar] [CrossRef]
- Pavão, F.; Castilho, M.S.; Pupo, M.T.; Dias, R.L.; Correa, A.G.; Fernandes, J.B.; da Silva, M.F.; Mafezoli, J.; Vieira, P.C.; Oliva, G. Structure of Trypanosoma cruzi glycosomal glyceraldehyde-3-phosphate dehydrogenase complexed with chalepin, a natural product inhibitor, at 1.95 A resolution. FEBS Lett. 2002, 520, 13–17. [Google Scholar] [CrossRef]
- Menezes, I.R.; Lopes, J.C.; Montanari, C.A.; Oliva, G.; Pavão, F.; Castilho, M.S.; Vieira, P.C.; Pupo, M.T. 3D QSAR studies on binding affinities of coumarin natural products for glycosomal GAPDH of Trypanosoma cruzi. J. Comput. Aided Mol. Des. 2003, 17, 277–290. [Google Scholar] [CrossRef] [PubMed]
- de Marchi, A.A.; Castilho, M.S.; Nascimento, P.G.; Archanjo, F.C.; del Ponte, G.; Oliva, G.; Pupo, M.T. New 3-piperonylcoumarins as inhibitors of glycosomal glyceraldehyde-3-phosphate dehydrogenase (gGAPDH) from Trypanosoma cruzi. Bioorg. Med. Chem. 2004, 12, 4823–4833. [Google Scholar] [CrossRef]
- Olorunsogo, O.O.; Emerole, G.O.; Malomo, S.O.; Thabrew, M.I. Inhibition of mitochondrial respiration by chalepin, a naturally occurring furocoumarin. Toxicol. Lett. 1983, 15, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Olorunsogo, O.O.; Uwaifo, A.O.; Malomo, S.O. Comparative effects of three naturally occurring furanocoumarins on mitochondrial bioenergetics. Chem. Biol. Interact. 1990, 74, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Richardson, J.S.M.; Aminudin, N.; Abd Malek, S.N. Chalepin: A Compound from Ruta angustifolia L. Pers Exhibits Cell Cycle Arrest at S phase, Suppresses Nuclear Factor-Kappa B (NF-κB) Pathway, Signal Transducer and Activation of Transcription 3 (STAT3) Phosphorylation and Extrinsic Apoptotic Pathway in Non-small Cell Lung Cancer Carcinoma (A549). Pharmacogn. Mag. 2017, 13, S489–S498. [Google Scholar]
- Fakai, M.I.; Abd Malek, S.N.; Karsani, S.A. Induction of apoptosis by chalepin through phosphatidylserine externalisations and DNA fragmentation in breast cancer cells (MCF7). Life Sci. 2019, 220, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Del Castillo, J.B.; Luis, F.R.; Secundino, M. Angustifolin, a coumarin from Ruta angustifolia. Phytochemistry 1984, 23, 2095–2096. [Google Scholar] [CrossRef]
- Jiang, Y.; Xu, J.; Huang, P.; Yang, L.; Liu, Y.; Li, Y.; Wang, J.; Song, H.; Zheng, P. Scoparone Improves Nonalcoholic Steatohepatitis Through Alleviating JNK/Sab Signaling Pathway-Mediated Mitochondrial Dysfunction. Front. Pharmacol. 2022, 13, 863756. [Google Scholar] [CrossRef]
- Wei, M.; Li, T.; Cao, H.; He, H.; Yang, C.; Yin, Y.; Lu, H.; Novák, P.; Zhang, K.; Gao, Y. The effects of scoparone on alcohol and high-fat diet-induced liver injury revealed by RNA sequencing. Biomed. Pharmacother. 2022, 155, 113770. [Google Scholar] [CrossRef]
- Park, K.R.; Kim, B.; Lee, J.Y.; Moon, H.J.; Kwon, I.K.; Yun, H.M. Effects of Scoparone on differentiation, adhesion, migration, autophagy and mineralization through the osteogenic signalling pathways. J. Cell. Mol. Med. 2022, 26, 4520–4529. [Google Scholar] [CrossRef]
- Li, N.; Yang, F.; Liu, D.Y.; Guo, J.T.; Ge, N.; Sun, S.Y. Scoparone inhibits pancreatic cancer through PI3K/Akt signaling pathway. World J. Gastrointest. Oncol. 2021, 13, 1164–1183. [Google Scholar] [CrossRef] [PubMed]
- Na, Z.; Xiang, W.; Niu, X.M.; Mei, S.X.; Lin, Z.W.; Li, C.M.; Sun, H.D. Diterpenoids from Isodon enanderianus. Phytochemistry 2002, 60, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.B.; Zhao, A.H.; Zhang, J.X.; Lu, Y.; Zhang, L.L.; Zheng, Q.T.; Sun, H.D. Cytotoxic constituents of Isodon rubescens var. lushiensis. J. Nat. Prod. 2003, 66, 1391–1394. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.G.; Qin, G.W.; Xie, Y.Y.; Cheng, K.F.; Lin, Z.W.; Sun, H.D.; Kang, Y.H.; Han, B.H. Lignans from Kadsura angustifolia. J. Asian Nat. Prod. Res. 1998, 1, 125–131. [Google Scholar] [CrossRef]
- Wysocka, W.; Przybyt, A. (+)-Angustifoline: A Minor Alkaloid from Lupinus albus1. Planta Med. 1993, 59, 289. [Google Scholar] [CrossRef]
- Cely-Veloza, W.; Quiroga, D.; Coy-Barrera, E. Quinolizidine-Based Variations and Antifungal Activity of Eight Lupinus Species Grown under Greenhouse Conditions. Molecules 2022, 27, 305. [Google Scholar] [CrossRef]
- Aslam, H.; Khan, A.U.; Qazi, N.G.; Ali, F.; Hassan, S.S.U.; Bungau, S. Pharmacological basis of bergapten in gastrointestinal diseases focusing on H+/K+ ATPase and voltage-gated calcium channel inhibition: A toxicological evaluation on vital organs. Front. Pharmacol. 2022, 13, 1005154. [Google Scholar] [CrossRef]
- Quetglas-Llabrés, M.M.; Quispe, C.; Herrera-Bravo, J.; Catarino, M.D.; Pereira, O.R.; Cardoso, S.M.; Dua, K.; Chellappan, D.K.; Pabreja, K.; Satija, S.; et al. Pharmacological Properties of Bergapten: Mechanistic and Therapeutic Aspects. Oxid. Med. Cell Longev. 2022, 2022, 8615242. [Google Scholar] [CrossRef]
- Gao, S.; Zou, X.; Wang, Z.; Shu, X.; Cao, X.; Xia, S.; Shao, P.; Bao, X.; Yang, H.; Xu, Y.; et al. Bergapten attenuates microglia-mediated neuroinflammation and ischemic brain injury by targeting Kv1.3 and Carbonyl reductase 1. Eur. J. Pharmacol. 2022, 933, 175242. [Google Scholar] [CrossRef]
- Yang, Y.; Han, J.; Lilly, R.G.; Yang, Q.; Guo, Y. Bergapten mediated inflammatory and apoptosis through AMPK/eNOS/AKT signaling pathway of isoproterenol-induced myocardial infarction in Wistar rats. J. Biochem. Mol. Toxicol. 2022, 36, e23143. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Xie, L.; Liu, K.; Cao, Y.; Dai, X.; Wang, X.; Lu, J.; Zhang, X.; Li, X. Bergapten: A review of its pharmacology, pharmacokinetics, and toxicity. Phytother. Res. 2021, 35, 6131–6147. [Google Scholar] [CrossRef]
- Santoro, M.; Guido, C.; De Amicis, F.; Sisci, D.; Cione, E.; Vincenza, D.; Donà, A.; Panno, M.L.; Aquila, S. Bergapten induces metabolic reprogramming in breast cancer cells. Oncol. Rep. 2016, 35, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.P.; Lin, C.S.; Lin, H.H.; Li, K.T.; Kao, S.H.; Tsao, S.M. Bergapten induces G1 arrest and pro-apoptotic cascade in colorectal cancer cells associating with p53/p21/PTEN axis. Environ. Toxicol. 2019, 34, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Del Castillo, J.B.; Secundino, M.; Luis, F.R. Four aromatic derivatives from Ruta angustifolia. Phytochemistry 1986, 25, 2209–2210. [Google Scholar] [CrossRef]
- Ulubelen, A.; Tan, N. A moskachan from roots of Ruta chalepensis. Phytochemistry 1990, 29, 3991–3992. [Google Scholar] [CrossRef]
- Joulain, D.; Laurent, R.; Fourniol, J.P.; Yaacob, K.B. Novel Moskachan Related Compounds in the Essential Oil of Ruta angustifolia Pers. from Malaysia. J. Essential Oil Res. 1991, 3, 355–357. [Google Scholar] [CrossRef]
- Salvo, G.; Ho, S.Y.; Rosenbaum, G.; Ree, R.; Conti, E. Tracing the temporal and spatial origins of island endemics in the Mediterranean region: A case study from the citrus family (Ruta L., Rutaceae). Syst. Biol. 2010, 59, 705–722. [Google Scholar] [CrossRef]
- Tounsi, M.S.; Wannes, W.A.; Ouerghemmi, I.; Msaada, K.; Smaoui, A.; Marzouk, B. Variation in essential oil and fatty acid composition in different organs of cultivated and growing wild Ruta chalepensis L. Indus. Crops Prod. 2011, 33, 617–623. [Google Scholar] [CrossRef]
- Ouerghemmi, I.; Rebey, I.B.; Rahali, F.Z.; Bourgou, S.; Pistelli, L.; Ksouri, R.; Marzouk, B.; Tounsi, M.S. Antioxidant and antimicrobial phenolic compounds from extracts of cultivated and wild-grown Tunisian Ruta chalepensis. J. Food Drug Anal. 2017, 25, 350–359. [Google Scholar] [CrossRef]
- Álvarez-Caballero, J.M.; Cuca-Suárez, L.E.; Coy-Barrera, E. Bio-Guided Fractionation of Ethanol Extract of Leaves of Esenbeckia alata Kunt (Rutaceae) Led to the Isolation of Two Cytotoxic Quinoline Alkaloids: Evidence of Selectivity against Leukemia Cells. Biomolecules 2019, 9, 585. [Google Scholar] [CrossRef] [PubMed]
- Coy Barrera, C.A.; Coy Barrera, E.D.; Granados Falla, D.S.; Delgado Murcia, G.; Cuca Suarez, L.E. Seco-limonoids and quinoline alkaloids from Raputia heptaphylla and their antileishmanial activity. Chem. Pharm. Bull. 2011, 59, 855–859. [Google Scholar] [CrossRef] [PubMed]
- Nganou, B.K.; Mbaveng, A.T.; Fobofou, S.A.T.; Fankam, A.G.; Bitchagno, G.T.M.; Simo Mpetga, J.D.; Wessjohann, L.A.; Kuete, V.; Efferth, T.; Tane, P. Furoquinolines and dihydrooxazole alkaloids with cytotoxic activity from the stem bark of Araliopsis soyauxii. Fitoterapia 2019, 133, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Shi, F.; Pan, H.; Cui, B.; Li, Y.; Huang, L.; Lu, Y. Dictamnine-induced hepatotoxicity in mice: The role of metabolic activation of furan. Toxicol. Appl. Pharmacol. 2019, 364, 68–76. [Google Scholar] [CrossRef]
- Feng, W.D.; Wang, Y.; Luo, T.; Jia, X.; Cheng, C.Q.; Wang, H.J.; Zhang, M.Q.; Li, Q.Q.; Wang, X.J.; Li, Y.Y.; et al. Scoparone suppresses mitophagy-mediated NLRP3 inflammasome activation in inflammatory diseases. Acta Pharmacol. Sin. 2022. online ahead of print. [Google Scholar] [CrossRef]
- Szewczyk, A.; Paździora, W.; Ekiert, H. The Influence of Exogenous Phenylalanine on the Accumulation of Secondary Metabolites in Agitated Shoot Cultures of Ruta graveolens L. Molecules 2023, 28, 727. [Google Scholar] [CrossRef]
- Szewczyk, A.; Marino, A.; Molinari, J.; Ekiert, H.; Miceli, N. Phytochemical Characterization, and Antioxidant and Antimicrobial Properties of Agitated Cultures of Three Rue Species: Ruta chalepensis, Ruta corsica, and Ruta graveolens. Antioxidants 2022, 11, 592. [Google Scholar] [CrossRef]
- Qahtan, A.A.; Faisal, M.; Alatar, A.A.; Abdel-Salam, E.M. Callus-Mediated High-Frequency Plant Regeneration, Phytochemical Profiling, Antioxidant Activity and Genetic Stability in Ruta chalepensis L. Plants 2022, 11, 1614. [Google Scholar] [CrossRef]
- Ereifej, K.I.; Feng, H.; Rababah, T.; Almajwal, A.; Alu’datt, M.; Gammoh, S.I.; Oweis, L.I. Chemical Composition, Phenolics, Anthocyanins Concentration and Antioxidant Activity of Ten Wild Edible Plants. Food Nutr. Sci. 2015, 6, 56083. [Google Scholar] [CrossRef]
- Parray, S.A.; Bhat, J.U.; Ahmad, G.; Jahan, N.; Sofi, G.; Iqbal, S.M.F. Ruta graveolens: From Traditional System of Medicine to Modern Pharmacology: An Overview. Am. J. PharmTech. Res. 2012, 2, 239–252. [Google Scholar]
- Kannan, R.; Babu, U.V. Identity and pharmacognosy of Ruta graveolens Linn. Anc. Sci. Life 2012, 32, 16–19. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Salas, J.; Gutiérrez-García, L.; Labrador-Moreno, J.; Ruiz-Té, T. Wild Plants Potentially Used in Human Food in the Protected Area “Sierra Grande de Hornachos” of Extremadura (Spain). Sustainability 2019, 11, 456. [Google Scholar] [CrossRef]
- Hanane, S.; Amhaouch, Z. Seasonal pattern in browsing trees and shrubs by reintroduced Cuvier’s gazelles: A study in Morocco with management implications. Eur. J. Wildl. Res. 2021, 67, 53. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, N.; Akram, M.; Yaniv-Bachrach, Z.; Daniyal, M. Is it safe to consume traditional medicinal plants during pregnancy? Phytother. Res. 2021, 35, 1908–1924. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytochemical Categories and Names | Activities | References |
|---|---|---|
| Alkaloids arborinine, γ-fagarine, graveoline, graveolinine, kokusaginine, pseudane IX. | Anticancer, antiviral, anti-inflammatory, antiparasitic | [21,38,39,44,49,56,57,58,59] |
| Coumarins angustifolin, bergapten, chalepin, chalepensin, rutamarine, scoparone, trihydroxycoumarin. | Anticancer, antioxidant, antiviral | [74,75,76,77,78,88] |
| Methyl ketones 2-decanone, 2-nonanone, 2-undecanone. | Antifungal, anti-inflammatory, insect repellent | [17,18,19,20,24,25] |
| Sterol ergosterol propyl ester. | Biocide | [23] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bailly, C. Ruta angustifolia Pers. (Narrow-Leaved Fringed Rue): Pharmacological Properties and Phytochemical Profile. Plants 2023, 12, 827. https://doi.org/10.3390/plants12040827
Bailly C. Ruta angustifolia Pers. (Narrow-Leaved Fringed Rue): Pharmacological Properties and Phytochemical Profile. Plants. 2023; 12(4):827. https://doi.org/10.3390/plants12040827
Chicago/Turabian StyleBailly, Christian. 2023. "Ruta angustifolia Pers. (Narrow-Leaved Fringed Rue): Pharmacological Properties and Phytochemical Profile" Plants 12, no. 4: 827. https://doi.org/10.3390/plants12040827
APA StyleBailly, C. (2023). Ruta angustifolia Pers. (Narrow-Leaved Fringed Rue): Pharmacological Properties and Phytochemical Profile. Plants, 12(4), 827. https://doi.org/10.3390/plants12040827