

The Distribution of Phytoecdysteroids among Terrestrial Vascular Plants: A Comparison of Two Databases and Discussion of the Implications for Plant/Insect Interactions and Plant Protection

Abstract
1. Introduction
2. Comparison of the Two Databases
2.1. General
2.2. Sources of the Data
2.3. Coverage
2.3.1. Lycophytes and Pteridophytes
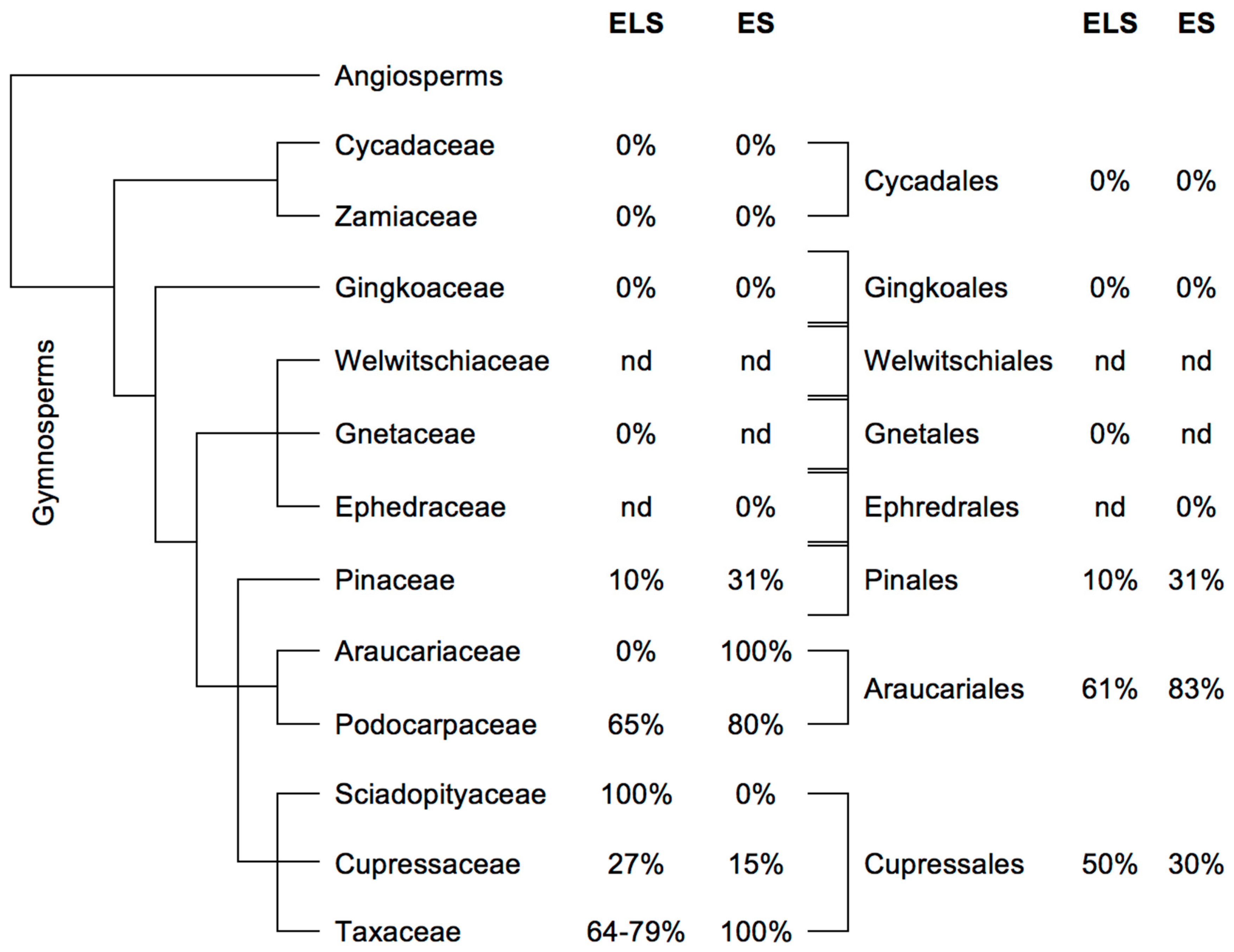
2.3.2. Gymnosperms
2.3.3. Angiosperms
2.4. Taxonomic Distribution of Phytoecdysteroids
2.4.1. Lycophytes and Pteridophytes
2.4.2. Gymnosperms
2.4.3. Angiosperms
2.4.4. Estimate of the Overall % Occurrence of Ecdysteroids in Terrestrial Vascular Plants
3. Discussion of the Implications for Plant/Insect Interactions and Plant Protection
3.1. Co-Evolution of Plants and Insects
3.2. Implications for Plant Protection
3.3. Prospects and Applications
3.4. Possible Relevance to Other Classes of Triterpenoids
4. Materials and Methods
4.1. Databases
4.1.1. Ecdybase Literature Survey (ELS)
4.1.2. Exeter Survey (ES)
4.2. Processing of Data
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Christenhusz, M.J.M.; Byng, J.W. The number of known plant species in the world and its annual increase. Phytotaxa 2016, 261, 201–217. [Google Scholar] [CrossRef]
- Antonelli, A.; Fry, C.; Smith, R.J.; Simmonds, M.S.J.; Kersey, P.J.; Pritchard, H.W.; Abbo, M.S.; Acedo, C.; Adams, J.; Ainsworth, A.M.; et al. State of the World’s Plants and Fungi. Royal Botanic Gardens: Kew, UK, 2020. [Google Scholar] [CrossRef]
- Fürstenberg-Hägg, J.; Zagrobelny, M.; Bak, S. Plant defense against insect herbivores. Int. J. Mol. Sci. 2013, 14, 10242–10297. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, K.; Koreeda, M.; Sasaki, S.; Chang, M.L.; Hsu, H.Y. Insect hormones. The structure of ponasterone A, an insect-moulting hormone from the leaves of Podocarpus nakaii Hay. J. Chem. Soc. Chem. Commun. 1966, 24, 915–917. [Google Scholar] [CrossRef]
- Galbraith, M.N.; Horn, D.H.S. An insect-moulting hormone from a plant. J. Chem. Soc. Chem. Commun. 1966, 24, 905–906. [Google Scholar] [CrossRef]
- Takemoto, T.; Ogawa, S.; Nishimoto, N. Studies on the constituents of Achyranthis radix. II. Isolation of insect moulting hormones. Yakugaku Zasshi 1967, 87, 1469–1473. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Heinrich, G.; Hoffmeister, H. Ecdyson als Begleitsubstanz des Ecdysterons in Polypodium vulgare L. Experientia 1967, 23, 995. [Google Scholar] [CrossRef]
- Kaplanis, J.N.; Thompson, M.J.; Robbins, W.E.; Bryce, B.M. Insect hormones: Alpha ecdysone and 20-hydroxyecdysone in bracken fern. Science 1967, 157, 1436–1438. [Google Scholar] [CrossRef]
- Jizba, J.; Herout, V.; Šorm, F. Isolation of ecdysterone (crustecdysone) from Polypodium vulgare L. rhizomes. Tetrahedron Lett. 1967, 18, 1689–1691. [Google Scholar] [CrossRef]
- Jizba, J.; Herou, T.V.; Šorm, F. Polypodine B–A novel ecdysone-like substance from plant material. Tetrahedron Lett. 1967, 51, 5139–5143. [Google Scholar] [CrossRef]
- Fukuzawa, A.; Kumagai, Y.; Masamune, T.; Furusaki, A.; Katayama, C.; Matsumoto, T. Acetylpinnasterol and pinnasterol, ecdysone-like metabolites from the marine red alga Laurencia pinnata Yamada. Tetrahedron Lett. 1981, 22, 4085–4086. [Google Scholar] [CrossRef]
- Fukuzawa, A.; Miyamoto, M.; Kumagai, Y.; Masamune, T. Ecdysone-like metabolites, 14α-hydroxypinnasterols from the red alga Laurencia pinnata. Phytochemistry 1986, 25, 1305–1307. [Google Scholar] [CrossRef]
- Dziwornu, G.A.; Caira, M.R.; de la Mare, J.-A.; Edkins, A.L.; Bolton, J.J.; Beukes, D.R.; Sunassee, S.N. Isolation, characterization and antiproliferative activity of new metabolites from the South African endemic red algal species Laurencia alfredensis. Molecules 2017, 22, 513. [Google Scholar] [CrossRef]
- Bandara, B.M.R.; Jayasinghe, L.; Karunaratne, V.; Wannigama, G.P.; Bokel, M.; Kraus, W.; Sotheeswaran, S. Ecdysterone from stem of Diploclisia glaucescens. Phytochemistry 1989, 28, 1073–1075. [Google Scholar] [CrossRef]
- Wang, J.-L.; Ruan, D.-C.; Cheng, Z.-Y.; Yang, C.-R. The dynamic variations of 20-hydroxyecdysone in Cyanotis arachnoidea. Acta Bot. Yunnanica 1996, 18, 459–464. [Google Scholar]
- Lafont, R.; Harmatha, J.; Marion-Poll, F.; Dinan, L.; Wilson, I.D. The Ecdysone Handbook, 3rd ed.; on-line; Cybersales s.a.: Prague, Czech Republic, 2002; Available online: https://ecdybase.org (accessed on 11 December 2022).
- Lafont, R. Phytoecdysteroids in the world flora: Diversity, distribution, biosynthesis and evolution. Russ. J. Plant Physiol. 1998, 45, 276–295. [Google Scholar]
- Dinan, L. Phytoecdysteroids: Biological aspects. Phytochemistry 2001, 57, 325–339. [Google Scholar] [CrossRef]
- Sláma, K. Ecdysteroids: Insect hormones, plant defensive factors, or human medicine. Phytoparasitica 1993, 21, 3–8. [Google Scholar] [CrossRef]
- Macháčková, I.; Vágner, M.; Sláma, K. Comparison between the effects of 20-hydroxyecdysone and phytohormones on growth and development in plants. Eur. J. Entomol. 1995, 92, 309–316. [Google Scholar]
- Dinan, L.; Harmatha, J.; Volodin, V.; Lafont, R. Phytoecdysteroids: Diversity, Biosynthesis and Distribution. In Ecdysone: Structures and Functions; Smagghe, G., Ed.; Springer Science + Business Media BV: Berlin/Heidelberg, Germany, 2009; pp. 3–45. [Google Scholar] [CrossRef]
- Dinan, L.; Lafont, R. Ecdysteroids as defensive chemicals. Adv. Insect Physiol. 2022, 63, 107–154. [Google Scholar] [CrossRef]
- Dinan, L.; Whiting, P.; Scott, A.J. Taxonomic distribution of phytoecdysteroids in seeds of members of the Chenopodiaceae. Biochem. Syst. Ecol. 1998, 26, 553–576. [Google Scholar] [CrossRef]
- Zibareva, L.; Volodin, V.; Saatov, Z.; Savchenko, T.; Whiting, P.; Lafont, R.; Dinan, L. Distribution of phytoecdysteroids in the Caryophyllaceae. Phytochemistry 2003, 64, 499–517. [Google Scholar] [CrossRef] [PubMed]
- Christenhusz, M.J.M.; Reveal, J.L.; Farjon, A.; Garder, M.F.; Mill, R.R.; Chase, M.W. A new classification and linear sequence of extant gymnosperms. Phytotaxa 2011, 19, 55–70. [Google Scholar] [CrossRef]
- PPG1. A community-derived classification for extant lycophytes and ferns. J. Syst. Evol. 2016, 54, 563–603. [Google Scholar] [CrossRef]
- APGIV. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef]
- Kariñho-Betancourt, E. Plant-herbivore interactions and secondary metabolites of plants: Ecological and evolutionary perspectives. Bot. Sci. 2018, 96, 35–51. [Google Scholar] [CrossRef]
- Wink, M. Evolution of secondary metabolites from an ecological and molecular phylogenetic perspective. Phytochemistry 2003, 64, 3–19. [Google Scholar] [CrossRef]
- Nabavi, S.M.; Šamec, D.; Tomczyk, M.; Milella, L.; Russo, D.; Habtemariam, S.; Suntar, I.; Rastrelli, L.; Daglia, M.; Xiao, J.; et al. Flavonoid biosynthetic pathways in plants: Versatile targets for metabolic engineering. Biotechnol. Adv. 2020, 38, 107316. [Google Scholar] [CrossRef]
- Bell, E.A.; Watson, A.A.; Nash, R.J. Non-protein amino acids: A review of the biosynthesis and taxonomic significance. Nat. Prod. Commun. 2008, 3, 93–110. [Google Scholar] [CrossRef]
- Steyn, P.S.; van Heerden, F.R. Bufadienolides of plant and animal origin. Nat. Prod. Rep. 1998, 15, 397–413. [Google Scholar] [CrossRef]
- Agrawal, A.A.; Petschenka, G.; Bingham, R.A.; Weber, M.G.; Rasmann, S. Tansley review: Toxic cardenolides: Chemical ecology and coevolution of specialized plant-herbivore interactions. New Phytol. 2012, 194, 28–45. [Google Scholar] [CrossRef]
- Morsy, N. Cardiac glycosides in medicinal plants. In Aromatic and Medicinal Plants–Back to Nature; El-Shemy, H.A., Ed.; IntechOpen: London, UK, 2017; Chapter 2. [Google Scholar] [CrossRef]
- Chomicki, G.; Schaefer, H.; Renner, S.S. Tansley review: Origin and domestication of Cucurbitaceae crops: Insights from phylogenies, genomics and archaeology. New Phytol. 2020, 226, 1240–1255. [Google Scholar] [CrossRef]
- Sparg, S.G.; Light, M.E.; van Staden, J. Biological activities and distribution of plant saponins. J. Ethnopharmacol. 2004, 94, 219–243. [Google Scholar] [CrossRef] [PubMed]
- Dhar, N.; Razdan, S.; Rana, S.; Bhat, W.W.; Vishwakarma, R.; Lattoo, S.K. A decade of molecular understanding of withanolide biosynthesis and in vitro studies in Withania somnifera (L.) Dunal: Prospects and perspectives for pathway engineering. Front. Plant Sci. 2015, 6, 1031. [Google Scholar] [CrossRef]
- Hikino, H.; Okuyama, T.; Jin, H.; Takemoto, T. Screening of Japanese ferns for phytoecdysones. I. Chem. Pharm. Bull. 1973, 21, 2292–2302. [Google Scholar] [CrossRef][Green Version]
- Yen, K.-Y.; Yang, L.-L.; Okuyama, T.; Hikino, H.; Takemoto, T. Screening of Formosan ferns for phytoecdysones. I. Chem. Pharm. Bull. 1974, 22, 805–808. [Google Scholar] [CrossRef]
- Dinan, L.; Savchenko, T.; Whiting, P. On the distribution of phytoecdysteroids in plants. Cell. Mol. Life Sci. 2001, 58, 1121–1132. [Google Scholar] [CrossRef]
- Schmelz, E.A.; Grebenok, R.J.; Ohnmeiss, T.E.; Bowers, W.S. Interactions between Spinacia oleracea and Bradysia impatiens: A role for phytoecdysteroids. Arch. Insect Biochem. Physiol. 2002, 51, 204–221. [Google Scholar] [CrossRef]
- Lafont, R.; Bouthier, A. Phytoecdysteroids: Structures, occurrence, biosynthesis and possible ecological significance. In Insect Chemical Ecology; Hrdý, I., Ed.; Academia: Prague, Czech Republic, 1991; pp. 197–214. [Google Scholar]
- Dinan, L. A strategy for the identification of ecdysteroid receptor agonists and antagonists from plants. Eur. J. Entomol. 1995, 92, 271–283. [Google Scholar]
- Futuyma, D.J. Some current approaches to the evolution of plant-herbivore interactions. Plant Species Biol. 2000, 15, 1–9. [Google Scholar] [CrossRef]
- Speed, M.P.; Fenton, A.; Jones, M.G.; Ruxton, G.D.; Brockhurst, M.A. Coevolution can explain defensive secondary metabolite diversity in plants. New Phytol. 2015, 208, 1251–1263. [Google Scholar] [CrossRef] [PubMed]
- DNP. Dictionary of Natural Products Version 31.1; CRC Press, Taylor & Francis Group, Informa PLC: London, UK, 2022. Available online: dnp.chemnetbase.com/faces/chemical/ChemicalSearch.html (accessed on 8 December 2022).
- Firn, R.D.; Jones, C.G. Natural products–A simple model to explain chemical diversity. Nat. Prod. Rep. 2003, 20, 382–391. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, P.R.; Raven, P.H. Butterflies and plants: A study in coevolution. Evolution 1964, 18, 586–608. [Google Scholar] [CrossRef]
- Occhipinti, A. Plant coevolution: Evidences and new challenges. J. Plant Interact. 2013, 8, 188–196. [Google Scholar] [CrossRef]
- Dawkins, R.; Krebs, J.R. Arms races between and within species. Proc. R. Soc. Lond. B Biol. Sci. 1979, 205, 489–511. [Google Scholar] [CrossRef] [PubMed]
- Janz, N.; Nylin, S. Butterflies and plants: A phylogenetic study. Evolution 1998, 52, 486–502. [Google Scholar] [CrossRef] [PubMed]
- Becerra, J.X. The impact of herbivore-plant coevolution on plant community structure. Proc. Natl. Acad. Sci. USA 2007, 104, 7483–7488. [Google Scholar] [CrossRef]
- Sharma, A.; Rana, C.; Shiwan, I.K. Important insect pests of cucurbits and their management. In Handbook of Cucurbits: Growth, Cultural Practices, and Physiology; CRC Press, Taylor & Francis Group: London, UK, 2016; Chapter 19; pp. 327–359. [Google Scholar]
- Engel, M.S. Insect Evolution. Curr. Biol. 2015, 25, R868–R872. [Google Scholar] [CrossRef]
- Bronstein, J.L.; Alarcón, R.; Geber, M. Tansley review:The evolution of plant-insect mutualisms. New Phytol. 2006, 172, 412–428. [Google Scholar] [CrossRef]
- Dinan, L.; Lafont, R. Compilation of the Literature Reports for the Screening of Vascular Plants, Algae, Fungi and Non-Arthropod Invertebrates for the Presence of Ecdysteroids; Version 11; Cybersales s.a.: Prague, Czech Republic, 2002; Available online: www.ecdybase.org (accessed on 26 September 2021).
- Savolainen, V.; Wuest, J.; Lafont, R.; Connat, J.-L. Effects of ingested phytoecdysteroids in the female soft tick Ornithodorus moubata. Experientia 1995, 51, 596–600. [Google Scholar] [CrossRef]
- Jones, C.G.; Firn, R.D. The role of phytoecdysteroids in bracken fern, Pteridium aquilinum (L.) Kuhn as a defence against phytophagous insect attack. J. Chem. Ecol. 1978, 4, 117–138. [Google Scholar] [CrossRef]
- Cooper-Driver, G.A. Defense strategies in bracken, Pteridium aquilinum (L.) Kuhn. Ann. Mo. Bot. Gard. 1990, 77, 281–286. [Google Scholar] [CrossRef]
- Alonso-Amelot, M.E. The chemistry and toxicology of bioactive compounds in bracken fern (Pteridium spp.), with special reference to chemical ecology and carcinogenesis. Stud. Nat. Prod. Chem. 2002, 26, 685–739. [Google Scholar]
- Der, J.P.; Thomson, J.A.; Stratford, J.K.; Wolf, P.G. Global chloroplast phylogeny and biogeography of bracken (Pteridium; Dennstaedtiaceae). Am. J. Bot. 2009, 96, 1041–1049. [Google Scholar] [CrossRef] [PubMed]
- Robinson, G.S.; Ackery, P.R.; Kitching, I.J.; Beccaloni, G.W.; Hernández, L.M. HOSTS–A Database of the World's Lepidopteran Hostplants; Natural History Museum: London, UK, 2010; Available online: http://www.nhm.ac.uk/hosts (accessed on 2 December 2022).
- DBIF. Database of Insects and Their Food Plants; Biological Records Centre: Wallingford, UK, 2008; Available online: dbif.brc.ac.uk (accessed on 2 December 2022).
- CAB Direct. Centre for Agriculture and Bioscience International. Available online: www.cabdirect.org/cabdirect (accessed on 2 December 2022).
- Dinan, L.; Dioh, W.; Veillet, S.; Lafont, R. 20-Hydroxyecdysone, from plant extracts to clinical use: Therapeutic potential for the treatment of neuromuscular, cardio-metabolic and respiratory diseases. Biomedicines 2021, 9, 492. [Google Scholar] [CrossRef]
- Volodin, V.; Chadin, I.; Whiting, P.; Dinan, L. Screening plants of European North-East Russia for ecdysteroids. Biochem. Syst. Ecol. 2002, 30, 525–578. [Google Scholar] [CrossRef]
- Volodin, V.V.; Loan, V.T.; Volodina, S.O.; Kuznetsov, A.N. Ecdysteroid-containing plants of the National Park Cuc Phuong (Northern Vietnam). Rep. Komi Res. Cent. 2018, 35, 46–53. [Google Scholar] [CrossRef]
- Zhang, Y.; Deng, T.; Lu, S.; Landis, J.; Moore, M.; Wang, H.; Wang, Y.; Hao, X.; Chen, J.; Li, S.X.M.; et al. Phylogenetic patterns suggest frequent multiple origins of secondary metabolites across the seed plant “tree of life”. Natl. Sci. Rev. 2021, 8, 12. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Whiting, P.; Šik, V.; Rees, H.H.; Dinan, L. Ecdysteroids and bufadienolides from Helleborus torquatus (Ranunculaceae). Phytochemistry 2001, 57, 401–407. [Google Scholar] [CrossRef]
- Yokosuka, A.; Inomata, M.; Yoshizawa, Y.; Iguchi, T.; Mimaki, Y. Bufadienolides and ecdysteroids from the whole plants of Helleborus niger and their cytotoxicity. J. Nat. Med. 2021, 75, 393–402. [Google Scholar] [CrossRef]
- Sawai, S.; Saito, K. Triterpenoid biosynthesis and engineering in plants. Front. Plant Sci. 2011, 2, 25. [Google Scholar] [CrossRef]
- Noushahi, H.A.; Khan, A.H.; Noushahi, U.F.; Hussain, M.; Javed, T.; Zafar, M.; Batool, M.; Ahmed, U.; Liu, K.; Harrison, M.T.; et al. Biosynthetic pathways of triterpenoids and strategies to improve their biosynthetic efficiency. Plant Growth Regul. 2022, 97, 439–454. [Google Scholar] [CrossRef] [PubMed]
- Chambliss, O.L.; Jones, C.M. Cucurbitacins: Specific insect attractants in Cucurbitaceae. Science 1996, 153, 1392–1393. [Google Scholar] [CrossRef]
- Metcalf, R.L. Coevolutionary adaptations of rootworm beetles (Coleoptera: Chrysomelidae) to cucurbitacins. J. Chem. Ecol. 1986, 12, 1109–1124. [Google Scholar] [CrossRef] [PubMed]
- Dinan, L.; Whiting, P.; Girault, J.P.; Lafont, R.; Dhadialla, T.S.; Cress, D.E.; Mugat, B.; Antoniewski, C.; Lepesant, J.A. Cucurbitacins are insect steroid hormone antagonists acting at the ecdysteroid receptor. Biochem. J. 1997, 327, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Dinan, L. The Exeter Survey of Plant Species for Ecdysteroid Agonist and Antagonist Activities (1995–2002); Version 1; Cybersales s.a.: Prague, Czech Republic, 2021. [Google Scholar]
- Dinan, L. The analysis of phytoecdysteroids in single (pre-flowering) specimens of fat hen, Chenopodium album. Phytochem. Anal. 1992, 3, 132–138. [Google Scholar] [CrossRef]
- Clément, C.Y.; Bradbrook, D.A.; Lafont, R.; Dinan, L. Assessment of a microplate-based bioassay for the detection of ecdysteroid-like or antiecdysteroid activities. Insect Biochem. Mol. Biol. 1993, 23, 187–193. [Google Scholar] [CrossRef]
- Brummitt, R.K. Vascular Plant Families and Genera; Royal Botanic Gardens: Kew, UK, 1992; p. 804, Print ISSN 2730-6259. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| % of All Species Assessed | |||
|---|---|---|---|
| No. Species # | ELS | ES | |
| Gymnosperms | 990 | 9.0% | 8.7% |
| Lycophytes | 1338 | 1.9% * | |
| Pteridophytes | 10,578 | 4.4% * | |
| Angiosperms | 295,383 | 0.77% | 1.37% |
| Class | Pathway | Plant Families | References |
|---|---|---|---|
| Alkaloids | Shikimic acid pathway | Fabaceae, Liliaceae, Solanaceae, Papaveraceae, Apocynaceae, Amaryllidaceae, Rununculaceae | [29] |
| Cyanogenic glycosides | Shikimic acid pathway | Most vascular plants: gymnosperms and angiosperms | [3] |
| Flavonoids | Phenylpropanoid pathway | All plants | [28,30] |
| Glucosinolates | Shikimic acid pathway | Brassicaceae, Capparidaceae, Tropaolaceae | [3] |
| Latex and resins | Various pathways | Ca. 10% of angiosperms | [3,28] |
| Mono- and diterpenes | MEP pathway | Lamiaceae | [29] |
| Non-protein amino acids | Modified AA pathways and novel metabolic routes | Fabaceae | [29,31] |
| Phenolics | Shikimic acid and/or malonic acid pathways | Widely distributed in vascular plants | [3] |
| Triterpenoids | MVA pathway | ||
| Bufadienolides | Liliaceae, Crassulaceae, Iridaceae, Melanthiaceae, Ranunculaceae, Santalaceae | [32] | |
| Cardenolides | Apocynaceae, Liliaceae, Ranunculaceae, Moraceae, Fabaceae, Scrophulariaceae, Cruciferae, Sterculaceae, Euphorpiaceae, tiliaceae, Celastraceae | [33,34] | |
| Cucurbitacins | Cucurbitaceae | [35] | |
| Phytoecdysteroids | Wide distribution | This report | |
| Steroidal alkaloids | Buxaceae, Liliaceae, Apocynaceae, Solanaceae | [29] | |
| Steroidal saponins | Monocotoledonous angiosperms (Agavaceae, Alliaceae, Asparagaceae, Dioscoreaceae, Liliaceae, Taccaceae) + Solanaceae | [36] | |
| Triterpenoid saponins | Dicotyledonous angiosperms (Amaranthaceae, Apiaceae, Caryophyllaceae, Fabaceae, Ranunculaceae) | [36] | |
| Withanolides | Mainly from certain genera in the Solanaceae (e.g., Iochroma, Datura, Jabrosa, Physalis, Salpichroa, Withania), but also certain species in the Taccaceae, Fabaceae, Dioscoraceae, Myrtaceae and Lamiaceae | [37] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dinan, L.; Lafont, F.; Lafont, R. The Distribution of Phytoecdysteroids among Terrestrial Vascular Plants: A Comparison of Two Databases and Discussion of the Implications for Plant/Insect Interactions and Plant Protection. Plants 2023, 12, 776. https://doi.org/10.3390/plants12040776
Dinan L, Lafont F, Lafont R. The Distribution of Phytoecdysteroids among Terrestrial Vascular Plants: A Comparison of Two Databases and Discussion of the Implications for Plant/Insect Interactions and Plant Protection. Plants. 2023; 12(4):776. https://doi.org/10.3390/plants12040776
Chicago/Turabian StyleDinan, Laurence, Françoise Lafont, and René Lafont. 2023. "The Distribution of Phytoecdysteroids among Terrestrial Vascular Plants: A Comparison of Two Databases and Discussion of the Implications for Plant/Insect Interactions and Plant Protection" Plants 12, no. 4: 776. https://doi.org/10.3390/plants12040776
APA StyleDinan, L., Lafont, F., & Lafont, R. (2023). The Distribution of Phytoecdysteroids among Terrestrial Vascular Plants: A Comparison of Two Databases and Discussion of the Implications for Plant/Insect Interactions and Plant Protection. Plants, 12(4), 776. https://doi.org/10.3390/plants12040776