
Early Canopy Management Practices Differentially Modulate Fruit Set, Fruit Yield, and Berry Composition at Harvest Depending on the Grapevine Cultivar

 , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Vine Vegetative Growth
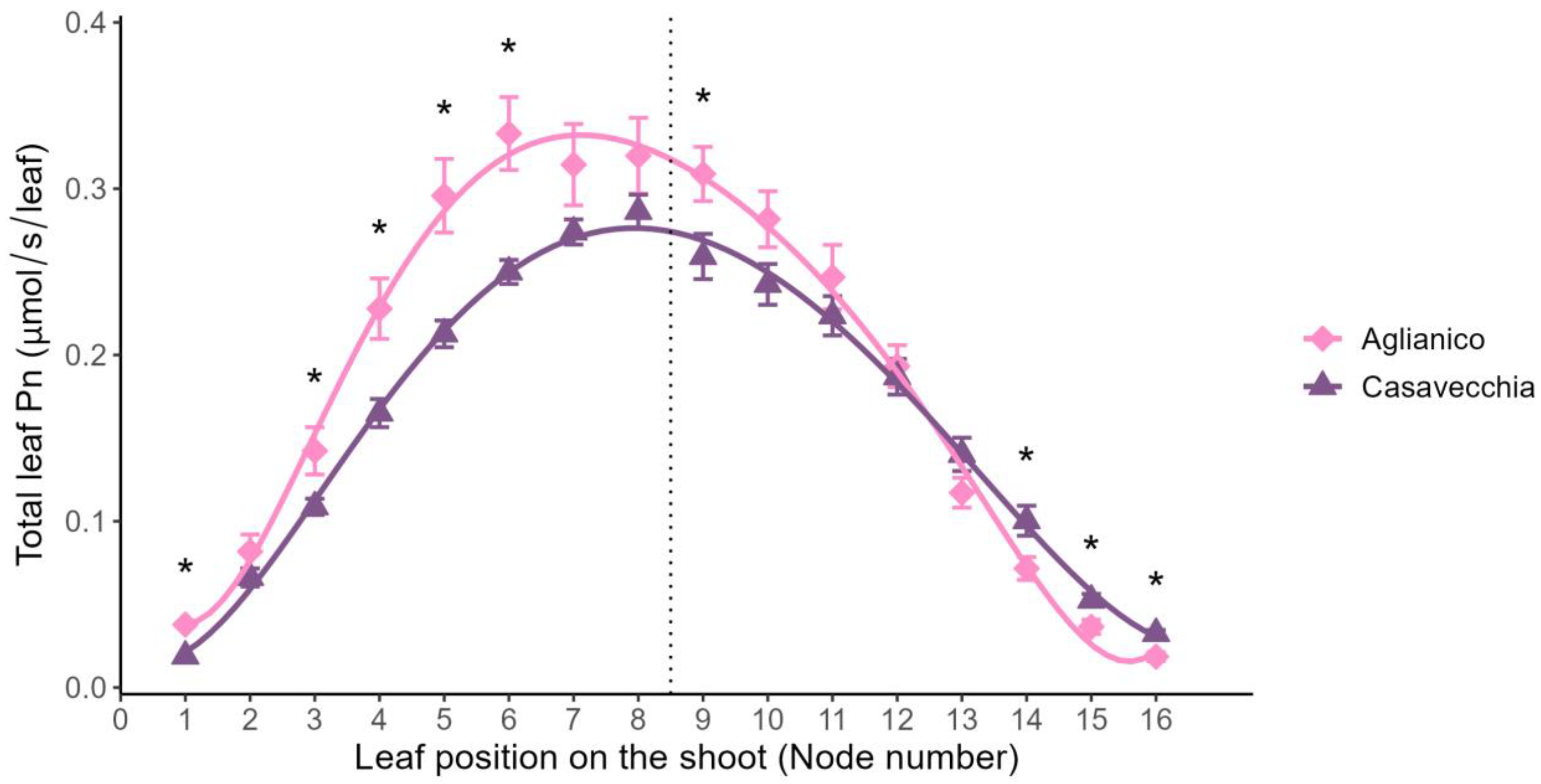
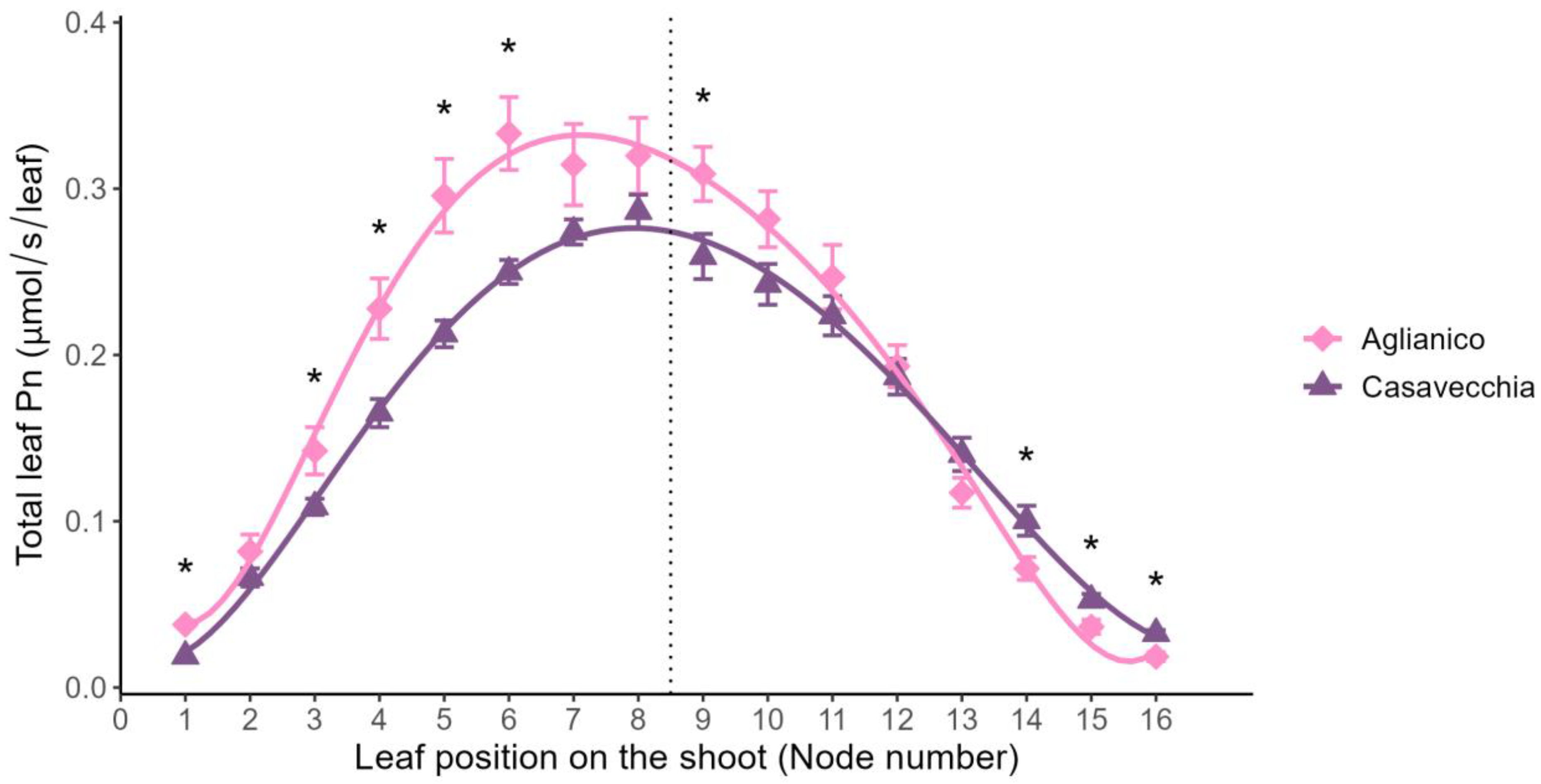
2.2. Leaf Net Photosynthetic Rate
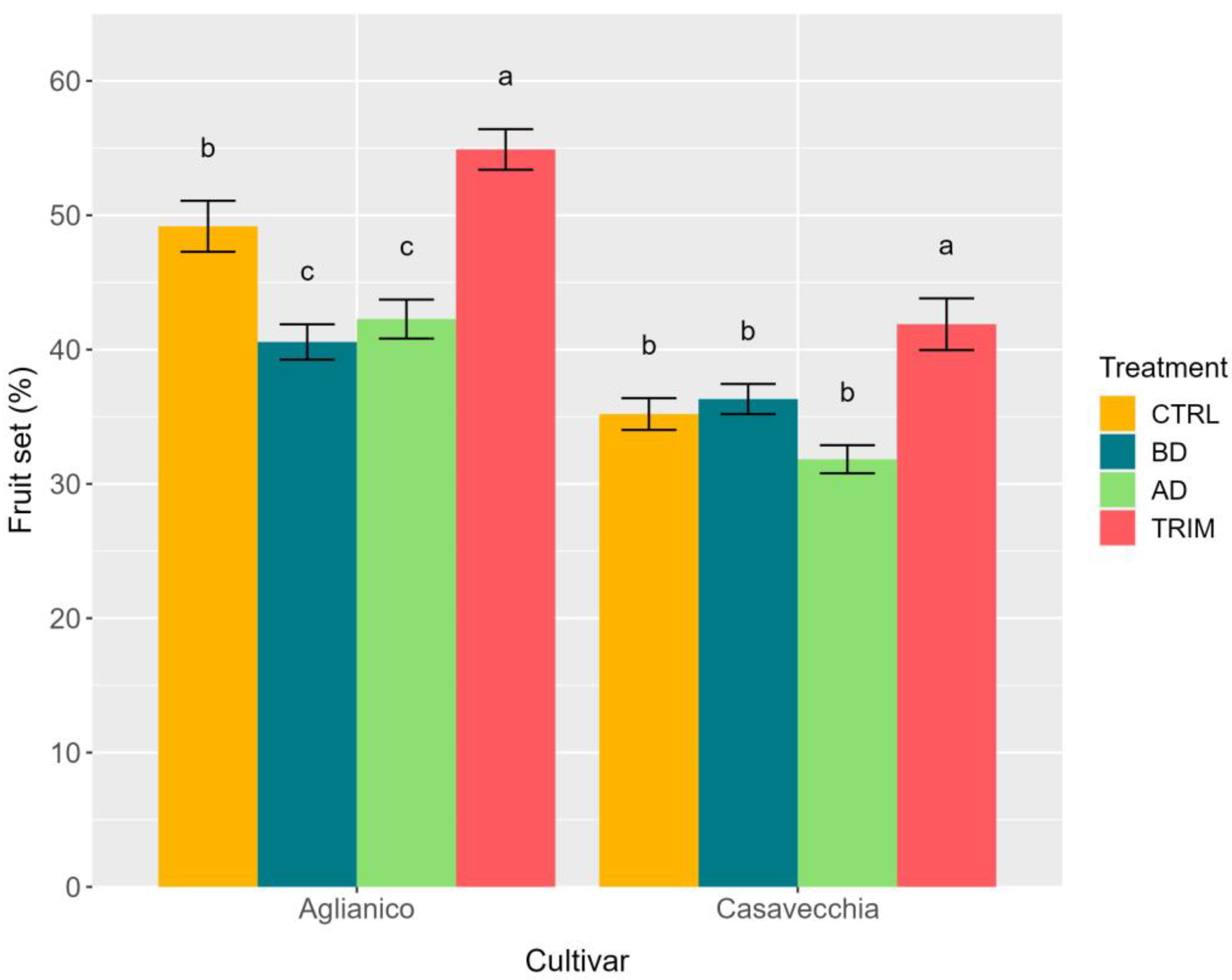
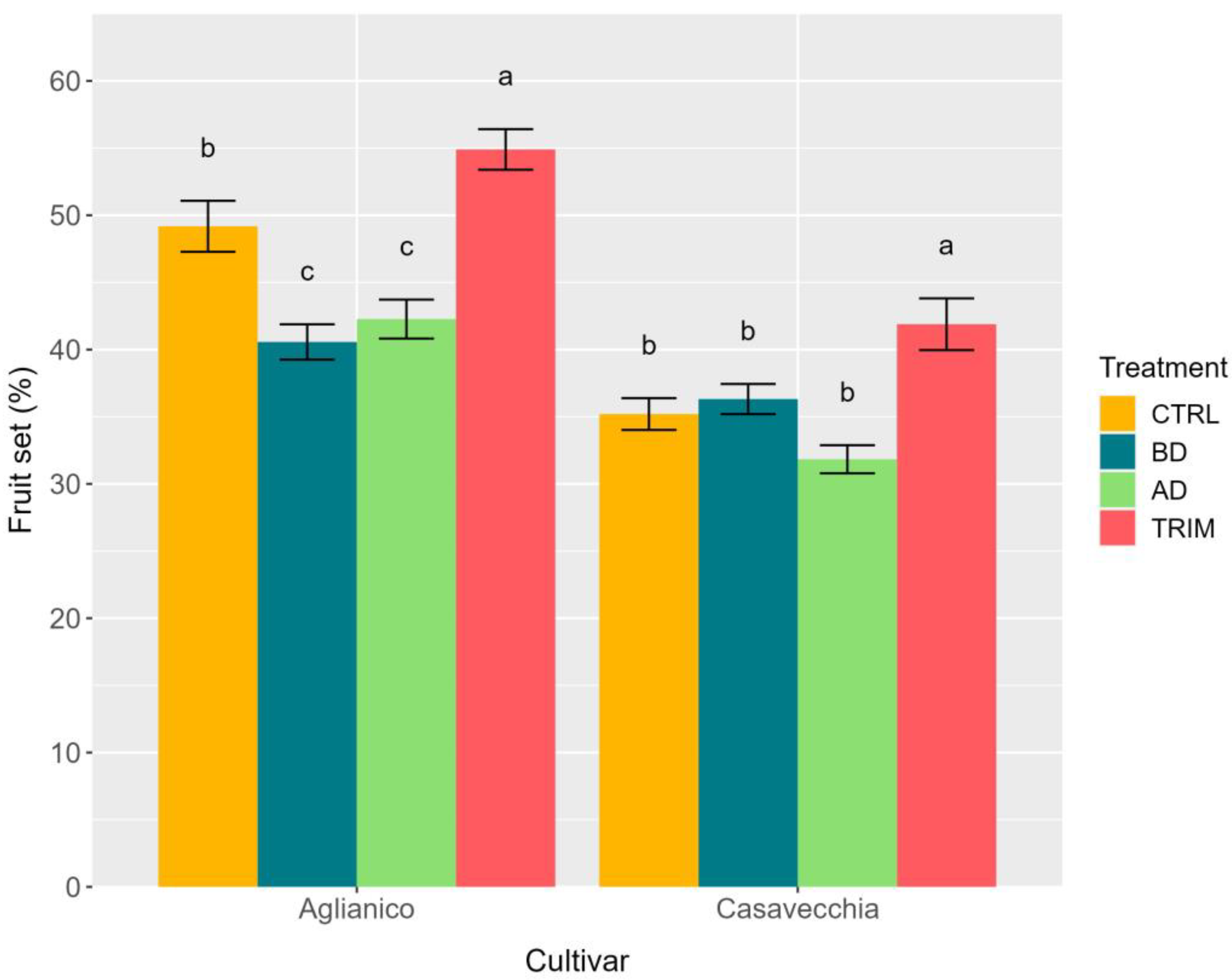
2.3. Inflorescence Fertility, Fruit Set, and Bunch Architecture at Harvest
2.4. Bud Potential Fertility, Fruit Yield Components, and Berry Composition at Harvest
3. Discussion
4. Materials and Methods
4.1. Experimental Site and Plant Material
4.2. Experimental Design and Treatments
4.3. Vegetative Growth Measurements
4.4. Leaf Photosynthetic Rate Measurements
4.5. Vine Fertility
4.6. Fruit Yield Components, Fruit Set, and Berry Composition at Harvest
4.7. Meteorological Data
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cangi, R.; Yagcı, A.; Akgul, S.; Kesgin, M.; Yanar, Y. Effects of shading and covering material application for delaying harvest on gray mold disease severity. Afr. J. Biotechnol. 2011, 10, 12182–12187. [Google Scholar]
- Intrigliolo, D.S.; Llacer, E.; Revert, J.; Esteve, M.D.; Climent, M.D.; Palau, D.; Gomez, I. Early defoliation reduces cluster compactness and improves grape composition in Mandó, an autochthonous cultivar of Vitis vinifera from southeastern Spain. Sci. Hortic. 2014, 167, 71–75. [Google Scholar] [CrossRef]
- Molitor, D.; Baron, N.; Sauerwein, T.; André, C.M.; Kicherer, A.; Döring, J.; Stoll, M.; Beyer, M.; Hoffmann, L.; Evers, D. Postponing first shoot topping reduces grape cluster compactness and delays bunch rot epidemic. Am. J. Enol. Vitic. 2015, 66, 164–176. [Google Scholar] [CrossRef]
- Tello, J.; Ibáñez, J. What do we know about grapevine bunch compactness? A state-of-the-art review. Aust. J. Grape Wine Res. 2018, 24, 6–23. [Google Scholar] [CrossRef]
- Fournier, E.; Gladieux, P.; Giraud, T. The ‘D r J ekyll and M r H yde fungus’: Noble rot versus gray mold symptoms of Botrytis cinerea on grapes. Evol. Appl. 2013, 6, 960–969. [Google Scholar] [CrossRef]
- Steel, C.C.; Blackman, J.W.; Schmidtke, L.M. Grapevine bunch rots: Impacts on wine composition, quality, and potential procedures for the removal of wine faults. J. Agric. Food Chem. 2013, 61, 5189–5206. [Google Scholar] [CrossRef]
- Reshef, N.; Walbaum, N.; Agam, N.; Fait, A. Sunlight modulates fruit metabolic profile and shapes the spatial pattern of compound accumulation within the grape cluster. Front. Plant Sci. 2017, 8, 70. [Google Scholar] [CrossRef]
- Tello, J.; Aguirrezábal, R.; Hernáiz, S.; Larreina, B.; Montemayor, M.I.; Vaquero, E.; Ibáñez, J. Multicultivar and multivariate study of the natural variation for grapevine bunch compactness. Aust. J. Grape Wine Res. 2015, 21, 277–289. [Google Scholar] [CrossRef]
- Bondada, B.; Covarrubias, J.I.; Tessarin, P.; Boliani, A.C.; Marodin, G.; Rombolà, A.D. Postveraison shoot trimming reduces cluster compactness without compromising fruit quality attributes in organically grown Sangiovese grapevines. Am. J. Enol. Vitic. 2016, 67, 206–211. [Google Scholar] [CrossRef]
- Basile, B.; Caccavello, G.; Giaccone, M.; Forlani, M. Effects of early shading and defoliation on bunch compactness, yield components, and berry composition of Aglianico grapevines under warm climate conditions. Am. J. Enol. Vitic. 2015, 66, 234–243. [Google Scholar] [CrossRef]
- Basile, B.; DeJong, T.M. Control of fruit tree vigor induced by dwarfing rootstocks. Hortic Rev 2018, 46, 39–97. [Google Scholar]
- Domingos, S.; Nobrega, H.; Raposo, A.; Cardoso, V.; Soares, I.; Ramalho, J.C.; Leitão, A.E.; Oliveira, C.M.; Goulao, L.F. Light management and gibberellic acid spraying as thinning methods in seedless table grapes (Vitis vinifera L.): Cultivar responses and effects on the fruit quality. Sci. Hortic. 2016, 201, 68–77. [Google Scholar] [CrossRef]
- Acimovic, D.; Tozzini, L.; Green, A.; Sivilotti, P.; Sabbatini, P. Identification of a defoliation severity threshold for changing fruitset, bunch morphology and fruit composition in Pinot Noir. Aust. J. Grape Wine Res. 2016, 22, 399–408. [Google Scholar] [CrossRef]
- Caccavello, G.; Giaccone, M.; Scognamiglio, P.; Forlani, M.; Basile, B. Influence of intensity of post-veraison defoliation or shoot trimming on vine physiology, yield components, berry and wine composition in Aglianico grapevines. Aust. J. Grape Wine Res. 2017, 23, 226–239. [Google Scholar] [CrossRef]
- Frioni, T.; Acimovic, D.; VanderWeide, J.; Tombesi, S.; Palliotti, A.; Gatti, M.; Poni, S.; Sabbatini, P. Whole-canopy source-sink balance at bloom dictates fruit set in cv. Pinot noir subjected to early leaf removal. Am. J. Enol. Vitic. 2019, 70, 411–419. [Google Scholar] [CrossRef]
- Palliotti, A.; Gatti, M.; Poni, S. Early leaf removal to improve vineyard efficiency: Gas exchange, source-to-sink balance, and reserve storage responses. Am. J. Enol. Vitic. 2011, 62, 219–228. [Google Scholar] [CrossRef]
- Poni, S.; Casalini, L.; Bernizzoni, F.; Civardi, S.; Intrieri, C. Effects of early defoliation on shoot photosynthesis, yield components, and grape composition. Am. J. Enol. Vitic. 2006, 57, 397–407. [Google Scholar] [CrossRef]
- Risco, D.; Pérez, D.; Yeves, A.; Castel, J.R.; Intrigliolo, D.S. Early defoliation in a temperate warm and semi-arid Tempranillo vineyard: Vine performance and grape composition. Aust. J. Grape Wine Res. 2014, 20, 111–122. [Google Scholar] [CrossRef]
- Bergqvist, J.; Dokoozlian, N.; Ebisuda, N. Sunlight exposure and temperature effects on berry growth and composition of Cabernet Sauvignon and Grenache in the Central San Joaquin Valley of California. Am. J. Enol. Vitic. 2001, 52, 1–7. [Google Scholar] [CrossRef]
- Spayd, S.E.; Tarara, J.M.; Mee, D.L.; Ferguson, J. Separation of sunlight and temperature effects on the composition of Vitis vinifera cv. Merlot berries. Am. J. Enol. Vitic. 2002, 53, 171–182. [Google Scholar] [CrossRef]
- Cataldo, E.; Salvi, L.; Paoli, F.; Fucile, M.; Mattii, G.B. Effects of defoliation at fruit set on vine physiology and berry composition in cabernet sauvignon grapevines. Plants 2021, 10, 1183. [Google Scholar] [CrossRef]
- Filippetti, I.; Movahed, N.; Allegro, G.; Valentini, G.; Pastore, C.; Colucci, E.; Intrieri, C. Effect of post-veraison source limitation on the accumulation of sugar, anthocyanins and seed tannins in Vitis vinifera cv. S angiovese berries. Aust. J. Grape Wine Res. 2015, 21, 90–100. [Google Scholar] [CrossRef]
- Tessarin, P.; Parpinello, G.P.; Rombolà, A.D. Physiological and enological implications of postveraison trimming in an organically-managed Sangiovese vineyard. Am. J. Enol. Vitic. 2018, 69, 59–69. [Google Scholar] [CrossRef]
- Frioni, T.; Acimovic, D.; Tombesi, S.; Sivilotti, P.; Palliotti, A.; Poni, S.; Sabbatini, P. Changes in within-shoot carbon partitioning in Pinot noir grapevines subjected to early basal leaf removal. Front. Plant Sci. 2018, 9, 1122. [Google Scholar] [CrossRef]
- Poni, S.; Palliotti, A. Traditional and innovative summer pruning techniques for vineyard management. Tradit. Innov. Summer Pruning Tech. Vineyard Manag. 2011, 25, 151–163. [Google Scholar]
- Quinlan, J.D.; Weaver, R.J. Modification of pattern of photosynthate movement within and between shoots of Vitis vinifera L. Plant Physiol. 1970, 46, 527–530. [Google Scholar] [CrossRef] [PubMed]
- Poni, S.; Giachino, E. Growth, photosynthesis and cropping of potted grapevines (Vitis vinifera L. cv. Cabernet Sauvignon) in relation to shoot trimming. Aust. J. Grape Wine Res. 2000, 6, 216–226. [Google Scholar] [CrossRef]
- Zhang, P.; Wu, X.; Needs, S.; Liu, D.; Fuentes, S.; Howell, K. The influence of apical and basal defoliation on the canopy structure and biochemical composition of Vitis vinifera cv. Shiraz grapes and wine. Front. Chem. 2017, 5, 48. [Google Scholar] [CrossRef]
- Bobeica, N.; Poni, S.; Hilbert, G.; Renaud, C.; Gomès, E.; Delrot, S.; Dai, Z. Differential responses of sugar, organic acids and anthocyanins to source-sink modulation in Cabernet Sauvignon and Sangiovese grapevines. Front. Plant Sci. 2015, 6, 382. [Google Scholar] [CrossRef]
- Vaillant-Gaveau, N.; Wojnarowiez, G.; Petit, A.-N.; Jacquens, L.; Panigai, L.; Clement, C.; Fontaine, F. Relationships between carbohydrates and reproductive development in Chardonnay grapevine: Impact of defoliation and fruit removal treatments during four successive growing seasons. OENO One 2014, 48, 219–229. [Google Scholar] [CrossRef]
- Caccavello, G.; Giaccone, M.; Scognamiglio, P.; Mataffo, A.; Teobaldelli, M.; Basile, B. Vegetative, yield, and berry quality response of aglianico to shoot-trimming applied at three stages of berry ripening. Am. J. Enol. Vitic. 2019, 70, 351–359. [Google Scholar] [CrossRef]
- Intrieri, C.; Filippetti, I.; Allegro, G.; Centinari, M.; Poni, S. Early defoliation (hand vs mechanical) for improved crop control and grape composition in Sangiovese (Vitis vinifera L.). Aust. J. Grape Wine Res. 2008, 14, 25–32. [Google Scholar] [CrossRef]
- Keller, M. The Science of Grapevines; Academic Press: Cambridge, MA, USA, 2020. [Google Scholar]
- Li, Y.-M.; Forney, C.; Bondada, B.; Leng, F.; Xie, Z.-S. The molecular regulation of carbon sink strength in grapevine (Vitis vinifera L.). Front. Plant Sci. 2021, 11, 606918. [Google Scholar] [CrossRef]
- Girona, J.; Marsal, J.; Mata, M.; Del Campo, J.; Basile, B. Phenological sensitivity of berry growth and composition of Tempranillo grapevines (Vitis vinifera L.) to water stress. Aust. J. Grape Wine Res. 2009, 15, 268–277. [Google Scholar] [CrossRef]
- Basile, B.; Marsal, J.; Mata, M.; Vallverdú, X.; Bellvert, J.; Girona, J. Phenological sensitivity of Cabernet Sauvignon to water stress: Vine physiology and berry composition. Am. J. Enol. Vitic. 2011, 62, 452–461. [Google Scholar] [CrossRef]
- Rolle, L.; Torchio, F.; Zeppa, G.; Gerbi, V. Anthocyanin extractability assesment of grape skins by texture analysis. OENO One 2008, 42, 157–162. [Google Scholar] [CrossRef]
- Romero-Cascales, I.; Ortega-Regules, A.; López-Roca, J.M.; Fernández-Fernández, J.I.; Gómez-Plaza, E. Differences in anthocyanin extractability from grapes to wines according to variety. Am. J. Enol. Vitic. 2005, 56, 212–219. [Google Scholar] [CrossRef]
- VanderWeide, J.; Frioni, T.; Ma, Z.; Stoll, M.; Poni, S.; Sabbatini, P. Early leaf removal as a strategy to improve ripening and lower cluster rot in cool climate (Vitis vinifera L.) Pinot Grigio. Am. J. Enol. Vitic. 2020, 71, 70–79. [Google Scholar] [CrossRef]
- Rossouw, G.C.; Smith, J.P.; Barril, C.; Deloire, A.; Holzapfel, B.P. Carbohydrate distribution during berry ripening of potted grapevines: Impact of water availability and leaf-to-fruit ratio. Sci. Hortic. 2017, 216, 215–225. [Google Scholar] [CrossRef]
- Kuhn, N.; Guan, L.; Dai, Z.W.; Wu, B.-H.; Lauvergeat, V.; Gomès, E.; Li, S.-H.; Godoy, F.; Arce-Johnson, P.; Delrot, S. Berry ripening: Recently heard through the grapevine. J. Exp. Bot. 2013, 65, 4543–4559. [Google Scholar] [CrossRef]
- Soil Surey Stuff. Keys to Soil Taxonomy, XIII ed.; USDA-Natural Resources Conservation Service: Washington, DC, USA, 2022. [Google Scholar]
- Baggiolini, M. Les stades repérés des arbres fruitiers à noyau. Rev. Rom. Agric. Vitic. Arboric 1952, 8, 3–4. [Google Scholar]
- Jones, G.V.; Duff, A.A.; Hall, A.; Myers, J.W. Spatial analysis of climate in winegrape growing regions in the western United States. Am. J. Enol. Vitic. 2010, 61, 313–326. [Google Scholar] [CrossRef]
- De Micheaux, P.L.; Drouilhet, R.; Liquet, B. The R Software; Springer: New York, NY, USA, 2013. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Main Shoot Length (cm) | Number of Main Leaves on the Shoot | Internode Length of the Main Shoot (cm) | Area of the Main Leaves (cm2/leaf) | Number of Lateral Shoots on the Main Shoot | Lateral Shoot Length (cm/lateral) | Number of Leaves per Lateral |
|---|---|---|---|---|---|---|---|
| Cultivar (CV) | |||||||
| ‘Aglianico’ | 100.0 ± 2.2 | 16.4 ± 0.2 | 6.1 ± 0.1 | 110.8 ± 2.9 | 7.4 ± 0.3 | 3.8 ± 0.1 | 2.2 ± 0.1 |
| ‘Casavecchia’ | 131.2 ± 3.2 | 16.0 ± 0.2 | 8.2 ± 0.2 | 97.0 ± 2.0 | 8.9 ± 0.3 | 5.5 ± 0.1 | 2.5 ± 0.1 |
| Significance | *** | n.s. | *** | *** | *** | *** | * |
| Source of Variation | No Flowers/INFLORESCENCE | Tot Pn per Shoot (Main + Laterals) after trt (μmol/s/shoot) | No Berries/Bunch | Fruit Set (%) | Total Rachis Length (cm) | Berry Fresh Weight (g/berry) | BCI-1 (No berries/cm) | BCI-2 (g berries/cm) |
|---|---|---|---|---|---|---|---|---|
| Cultivar (CV) | ||||||||
| ‘Aglianico’ | 239 ± 9 b | 2.80 ± 0.17 b | 114 ± 5 a | 47.0 ± 0.9 a | 36.9 ± 1.6 b | 1.86 ± 0.04 a | 3.26 ± 0.12 a | 5.50 ± 0.18 a |
| ‘Casavecchia’ | 309 ± 15 a | 3.61 ± 0.2 a | 112 ± 6 a | 36.1 ± 0.8 b | 84.1 ± 3.8 a | 1.90 ± 0.03 a | 1.34 ± 0.04 b | 2.48 ± 0.09 b |
| Significance | *** | *** | n.s. | *** | *** | n.s. | *** | *** |
| Canopy management (CM) | ||||||||
| Control | 274 ± 21 a | 4.8 ± 0.29 a | 119 ± 10 ab | 42.9 ± 1.6 b | 56.5 ± 4.9 a | 1.96 ± 0.07 a | 2.35 ± 0.12 b | 4.17 ± 0.29 ab |
| Basal Defoliation | 270 ± 19 a | 2.64 ± 0.2 b | 103 ± 7 b | 38.7 ± 0.9 c | 59.4 ± 6.3 a | 1.96 ± 0.07 a | 2.28 ± 0.19 b | 4.31 ± 0.35 ab |
| Apical Defoliation | 259 ± 13 a | 2.62 ± 0.18 b | 96 ± 4 b | 37.6 ± 1.2 c | 55.3 ± 4.2 a | 1.89 ± 0.05 ab | 2.10 ± 0.14 b | 3.65 ± 0.24 b |
| Shoot Trimming | 272 ± 14 a | 2.74 ± 0.24 b | 135 ± 8 a | 49.8 ± 1.5 a | 57.9 ± 4.8 a | 1.72 ± 0.03 b | 2.99 ± 0.25 a | 4.68 ± 0.32 a |
| Significance | n.s. | *** | *** | *** | n.s. | ** | *** | * |
| CV × CM | ||||||||
| Significance | n.s. | n.s. | n.s. | ** | n.s. | n.s. | n.s. | n.s. |
| Source of Variation | Bud Potential Fertility Index (No bunches/bud) | Fruit Yield (kg/vine) | No Bunches per Vine | Bunch Fresh Weight (g/bunch) | Total Soluble Solids (°Brix) | pH | Titratable Acidity (% Tartaric Acid) |
|---|---|---|---|---|---|---|---|
| Cultivar (CV) | |||||||
| ‘Aglianico’ | 1.64 ± 0.06 a | 6.47 ± 0.40 a | 33 ± 1 a | 194.7 ± 9.3 a | 21.3 ± 0.1 b | 3.23 ± 0.01 b | 0.77 ± 0.01 a |
| ‘Casavecchia’ | 1.21 ± 0.05 b | 3.83 ± 0.25 b | 19 ± 1 b | 205.4 ± 9.3 a | 22.6 ± 0.1 a | 3.48 ± 0.01 a | 0.45 ± 0.01 b |
| Significance | *** | *** | *** | n.s. | *** | *** | *** |
| Canopy management (CM) | |||||||
| Control | 1.36 ± 0.09 a | 5.35 ± 0.69 a | 24 ± 3 a | 222.7 ± 18.2 a | 21.8 ± 0.2 b | 3.37 ± 0.03 ab | 0.62 ± 0.03 ab |
| Basal Defoliation | 1.47 ± 0.10 a | 5.21 ± 0.55 a | 26 ± 2 a | 197.8 ± 11.2 a | 21.8 ± 0.2 b | 3.33 ± 0.02 b | 0.60 ± 0.03 ab |
| Apical Defoliation | 1.42 ± 0.10 a | 4.69 ± 0.40 a | 27 ± 2 a | 179.3 ± 9.7 a | 22.4 ± 0.2 a | 3.40 ± 0.03 a | 0.57 ± 0.02 b |
| Shoot Trimming | 1.44 ± 0.09 a | 5.35 ± 0.61 a | 26 ± 2 a | 200.3 ± 10.4 a | 21.7 ± 0.2 b | 3.32 ± 0.02 b | 0.64 ± 0.03 a |
| Significance | n.s. | n.s. | n.s. | n.s. | ** | ** | * |
| CV × CM | |||||||
| Significance | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mataffo, A.; Scognamiglio, P.; Molinaro, C.; Corrado, G.; Basile, B. Early Canopy Management Practices Differentially Modulate Fruit Set, Fruit Yield, and Berry Composition at Harvest Depending on the Grapevine Cultivar. Plants 2023, 12, 733. https://doi.org/10.3390/plants12040733
Mataffo A, Scognamiglio P, Molinaro C, Corrado G, Basile B. Early Canopy Management Practices Differentially Modulate Fruit Set, Fruit Yield, and Berry Composition at Harvest Depending on the Grapevine Cultivar. Plants. 2023; 12(4):733. https://doi.org/10.3390/plants12040733
Chicago/Turabian StyleMataffo, Alessandro, Pasquale Scognamiglio, Carlo Molinaro, Giandomenico Corrado, and Boris Basile. 2023. "Early Canopy Management Practices Differentially Modulate Fruit Set, Fruit Yield, and Berry Composition at Harvest Depending on the Grapevine Cultivar" Plants 12, no. 4: 733. https://doi.org/10.3390/plants12040733
APA StyleMataffo, A., Scognamiglio, P., Molinaro, C., Corrado, G., & Basile, B. (2023). Early Canopy Management Practices Differentially Modulate Fruit Set, Fruit Yield, and Berry Composition at Harvest Depending on the Grapevine Cultivar. Plants, 12(4), 733. https://doi.org/10.3390/plants12040733