Abstract
Glyceollins, isoflavonoid-derived antimicrobial metabolites, are the major phytoalexins in soybean (Glycine max). They play essential roles in providing resistance to the soil-borne pathogen Phytophthora sojae and have unconventional anticancer and neuroprotective activities that render them desirable for pharmaceutical development. Our previous studies revealed that the transcription factors GmMYB29A2 and GmNAC42-1 have essential roles in activating glyceollin biosynthesis, yet each cannot activate the transcription of all biosynthesis genes in the absence of a pathogen elicitor treatment. Here, we report that co-overexpressing both transcription factors is also insufficient to activate glyceollin biosynthesis. To understand this insufficiency, we compared the transcriptome profiles of hairy roots overexpressing each transcription factor with glyceollin-synthesizing roots treated with wall glucan elicitor (WGE) from P. sojae. GmMYB29A2 upregulated most of the WGE-regulated genes that encode enzymatic steps spanning from primary metabolism to the last step of glyceollin biosynthesis. By contrast, GmNAC42-1 upregulated glyceollin biosynthesis genes only when overexpressed in the presence of WGE treatment. This is consistent with our recent discovery that, in the absence of WGE, GmNAC42-1 is bound by GmJAZ1 proteins that inhibit its transactivation activity. WGE, and not GmMYB29A2 or GmNAC42-1, upregulated the heat shock family gene GmHSF6-1, the homolog of Arabidopsis HSFB2a that directly activated the transcription of several glyceollin biosynthesis genes. Our results provide important insights into what biosynthesis genes will need to be upregulated to activate the entire glyceollin biosynthetic pathway.
1. Introduction
Phytoalexins are plant defense metabolites that are biosynthesized in response to pathogens [1,2,3]. Each plant species produces distinct phytoalexin metabolites that, collectively, represent molecules from all categories of specialized metabolism [4,5,6]. Their chemical diversity and roles as plant defense metabolites may be what affords them a wide range of biological activities in human cells [7]. The most well-known phytoalexins that are clinical drugs include the anticancer metabolite taxol [8], the antidiabetic berberine [9], and the antimalarial artemisinin [10]. Many other phytoalexins are in (pre)clinical testing [5,11,12], including the glyceollins from soybean that inhibit the survival and proliferation of several cancer cell and tumor types by mechanisms that are distinct from conventional therapeutics [13,14,15]. Additionally, glyceollins hold promise as natural antimicrobials in the swine industry [16,17], yet exhibit neuroprotective activities in mammalian cells [18]. All phytoalexins share one commonality: hey are produced at a low level in response to pathogens. Since phytoalexin biosynthesis is primarily regulated at the level of transcription of the biosynthesis genes [19,20,21], this feature makes the transcription factors that regulate those genes of prime importance as genetic targets for improving phytoalexin yields [21,22,23].
The regulation of phytoalexin biosynthesis is complex as it involves both positive and redundant, negative regulatory mechanisms, some of which were recently discovered to be conserved in regulating diverse phytoalexin biosynthetic pathways among different plant species (reviewed by [22]). In Arabidopsis, WRKY33 is a well-characterized transcription factor that directly positive regulates camalexin biosynthesis genes [20,24,25]. The phosphorylation of WRKY33 by the protein kinases MPK3/MPK6 and CPK5/CPK6 enhances its DNA binding and transactivation activities, respectively, and is needed for full activation of camalexin biosynthesis [26,27]. The ethylene response factor 1 (ERF1) interacts with WRKY33 and directly binds camalexin biosynthesis genes to mediate synergy between the ethylene, jasmonate, and map kinase (MAPK), signaling pathways in activating camalexin biosynthesis [28]. ERF72 is also phosphorylated by MPK3/MPK6 and physically interacts with WRKY33 to directly regulate camalexin biosynthesis genes [29]. WRKY33 directly represses ABA signaling genes, including JAZ1, and ABA suppresses camalexin biosynthesis [30]. RNAi silencing of JAZ1 in Arabidopsis calli was recently shown to partially activate the biosynthesis of camalexin in the absence of an elicitor treatment [31]. Similarly, RNAi silencing VvJAZ9 (the grapevine homolog of JAZ1) in grapevine cell cultures increases the production of the stilbene phytoalexin resveratrol [32]. By RNA-seq analyses of transcripts up and downregulated with glyceollin biosynthesis, we recently identified GmNAC42-1 (Glyma.02G284300) as the first transcription factor that is an essential and direct activator of glyceollin phytoalexin biosynthesis in soybean [33]. The Arabidopsis homolog of GmNAC42-1 was previously shown to be required for the activation of camalexin biosynthesis in Arabidopsis [34], yet it still remains unknown whether it directly regulates camalexin biosynthesis genes.
Transcription factors of the MYB family have not yet been found to directly regulate camalexin biosynthesis genes. However, the R2R3-MYB-type transcription factors VvMYB14 and VvMYB15 from grapevine (Vitis vinifera) directly regulate shikimate, phenylpropanoid, and stilbene synthase genes, the latter being specific to stilbene phytoalexin biosynthesis [35,36]. VvWRKY8 physically interacts with VvMYB14, inhibiting its interaction with VvSTS15/21 promoters to suppress stilbene biosynthesis [37]. By gene overexpression and silencing experiments in soybean hairy roots, we recently discovered that GmMYB29A2 (Glyma.02G005600), the soybean homolog of VvMYB14, is essential for activating glyceollin I phytoalexin biosynthesis in soybean and provides race-specific resistance to the pathogen Phytophthora sojae [2]. Like GmNAC42-1, GmMYB29A2 directly binds the promoter regions of glyceollin biosynthesis genes (i.e., IFS2 and G4DT) in electrophoretic mobility shift and yeast one-hybrid assays [2,33]. Since these transcription factors directly activate the expressions of two genes that are required for glyceollin biosynthesis, they are prime candidates to engineer to fully activate glyceollin biosynthesis for stable production of those molecules. However, overexpressing either GmNAC42-1 or GmMYB29A2 without WGE treatment failed to activate the transcription of several glyceollin biosynthesis genes, resulting in no accumulation of glyceollins.
Glyceollin biosynthesis involves around 43 metabolic steps starting from glucose (Figure 1). In primary metabolism, D-glucose is transformed into D-erythrose 4-phosphate through the pentose phosphate pathway, followed by enzymatic condensation with pyruvate, oxidative cyclization, and dehydration to generate shikimate [38,39,40]. Shikimate is initially converted into L-phenylalanine, followed by reductive deamination and hydroxylation to generate 4-coumarate throughout the phenylpropanoid pathway [41,42]. In the isoflavonoid pathway, isoliquiritigenin is generated by condensation of 4-coumarate with CoA and 3-molonyl-CoA and transformed into daidzein. Daidzein is reduced and hydroxylated to produce (3R)-2′-hydroxydihydrodaidzein, the precursor of glycinol [33,43,44]. Finally, glyceollins I, II, and III are produced through the condensation of (-)-glycinol and prenyl diphosphate. The addition of prenyl diphosphate in the position 4 of (-)-glycinol is needed to produce glyceollin I, while the addition in position 2 for glyceollins II and III [45,46,47].
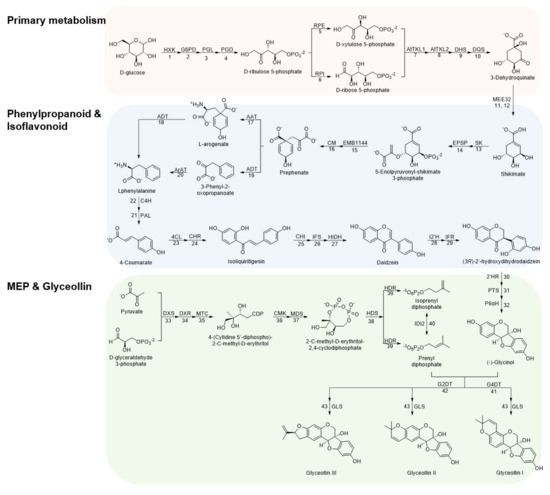
Figure 1.
Glyceollin biosynthesis from D−glucose. All 43 enzymatic steps from primary metabolism, to phenylpropanoid and isoflavonoid pathways, to MEP and glyceollin biosynthesis are shown. HXK, hexokinase; G6PD, glucose−6−phosphate dehydrogenase; PGL, 6−phosphogluconolactonase; PGD, 6−phosphogluconate dehydrogenase; RPE, ribulose−phosphate 3−epimerase; RPI, ribose−5−phosphate isomerase; AtTKL1, Arabidopsis thaliana transketolase 1; AtTKL2, Arabidopsis thaliana transketolase 2; DHS, 3−deoxy−7−phosphoheptulonate synthase; DQS, 3−dehydroquinate synthase; MEE32, shikimate dehydrogenase; SK, shikimate kinase; EPSP, 3−phosphoshikimate 1−carboxyvinyltransferase; EMB1144, chorismate synthase; CM, chorismate mutase; AAT, L−glutamate:prephenate aminotransferase; ADT, arogenate dehydratase; ArAT, aromatic aminotransferase; PAL, phenylalanine ammonia−lyase; C4H, cinnamic acid 4−hydroxylase; 4CL, 4−coumarate−coenzyme A ligase; CHR, chalcone reductase; CHI, chalcone isomerase; IFS, isoflavone synthase; HIDH, 2−hydroxyisoflavanone dehydratase; I2′H, isoflavone 2′−hydroxylase; IFR, isoflavone reductase; 2′HR, (3R)−2′−hydroxyisoflavanone reductase; PTS, pterocarpan synthase; P6αH, dihydroxypterocarpan− 6α−hydroxylase; DXS, 1−deoxy−D−xylulose 5−phosphate synthase; DXR, 1−deoxy−D−xylulose 5−phosphate reductoisomerase; MTC, 2−C−methyl−D−erythritol 4−phosphate cytidylyltransferase; CMK, 4−(cytidine 5′−diphospho)−2−C−methyl−D−erythritol kinase; MDS, 2−C−methyl−D−erythritol 2,4−cyclodiphosphate synthase; HDS, (E)−4−hydroxy−3−methylbut−2−enyl−diphosphate synthase; HDR, 4−hydroxy−3−methylbut−2−en−1−yl diphosphate reductase; IDI2, isopentenyl−diphosphate Δ−isomerase; G4DT, trihydroxypterocarpan dimethylallyltransferase; G2DT, dimethylallylpyrophosphate:trihydroxypterocarpan dimethylallyl transferase; GLS, glyceollin synthase.
2. Results
2.1. Co-Overexpressing the Transcription Factors GmMYB29A2 and GmNAC42-1 Fails to Activate Glyceollin Biosynthesis in Absence of an Elicitor Treatment
Since GmMYB29A2 and GmNAC42-1 failed to activate glyceollin biosynthesis when each gene was overexpressed individually in the absence of an elicitor [2,33], we tested whether co-overexpressing both transcription factors together could activate glyceollin biosynthesis. GmMYB29A2 and GmNAC42-1 were cloned upstream of the p35S and pFMV viral promoters of the pGEMINI vector [48] and were co-overexpressed 26.6- and 45-fold, respectively, in mock-treated roots (Figure 2A). The co-overexpression resulted in a modest (1.2-fold) upregulation of HIDH and G4DT by qRT-PCR, whereas C4H, IFS2, and I2′H expressions were not upregulated. Glyceollin metabolite amounts also remained unchanged (Figure 2B). In contrast, co-overexpressing GmMYB29A2 and GmNAC42-1 3.5- and five-fold in WGE-treated roots upregulated I2′H and G4DT 1.2- to 1.6-fold, respectively, and resulted in a 1.8-fold increase in the amounts of glyceollin metabolites (Figure 2A,B).
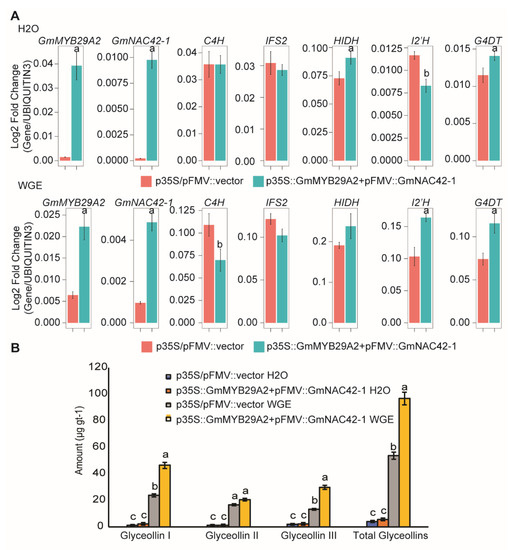
Figure 2.
Co−overexpressing the transcription factors GmMYB29A2 and GmNAC42−1 fails to activate glyceollin biosynthesis in absence of an elicitor treatment. (A) Gene expressions in 24 h mock (H2O)−treated and P. sojae wall glucan elicitor (WGE)−treated hairy roots co−overexpressing GmMYB29A2 and GmNAC42−1. The significance test was performed by paired students t−test, which is indicated by different letters (p < 0.05). (B) Amounts of glyceollin metabolites from hairy roots overexpressing GmMYB29A2 and GmNAC42−1 under mock or WGE treatment. The significance test was performed by single factor ANOVA, Tukey post hoc test, which is indicated by different letters (p < 0.01). Error bars represent SE (n ≥ 3).
In summary, while co-overexpressing GmMYB29A2 and GmNAC42-1 increases glyceollin biosynthesis in WGE-elicited hairy roots, it is insufficient to drive glyceollin biosynthesis in the absence of an elicitor treatment. The observed results raised the question of the relative roles of WGE and these two transcription factors in reprogramming soybean genome expressions.
2.2. Comparative Transcriptome Analyses Identify Common and Distinct Gene Sets Regulated by WGE, GmMYB29A2, and GmNAC42-1
To gain insight into the relative roles of WGE, GmMYB29A2 and GmNAC42-1, we compared the transcriptomes of WGE-treated empty-vector-transformed hairy roots that were biosynthesizing glyceollins to mock-treated hairy roots overexpressing GmMYB29A2 or GmNAC42-1 that were not biosynthesizing those metabolites. The WGE-treated hairy roots upregulated 3567 genes while downregulated 3567 genes (Figure 3A). Similarly, overexpressing GmMYB29A2 2.9-fold upregulated 3140 genes and downregulated 2926. However, overexpressing GmNAC42-1 115-fold only upregulated 890 and downregulated 403 genes. Of the genes that were upregulated by WGE, 765 and 534 were also upregulated by p35S::GmMYB29A2 and p35S::GmNAC42-1, respectively, with 93 genes being upregulated in all three genotypes (Figure 3A, Supplementary Tables S1–S3). The latter gene-set included the transcription factor GmMYB29A1 (Glyma.10G006600), that putatively has a role in promoting glyceollin turnover and/or competing biosynthetic pathways [2]. Of the WGE-downregulated genes, 571 and 102 were also downregulated in p35S::GmMYB29A2 and p35S::GmNAC42-1, respectively, with 52 genes being downregulated in all three genotypes (Supplementary Tables S4–S6).
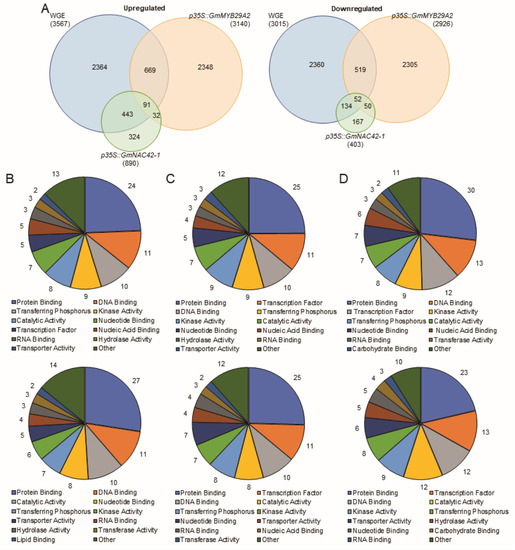
Figure 3.
A comparison of differentially regulated genes among P. sojae wall glucan elicitor (WGE)-treated W82 hairy roots and mock-treated hairy roots overexpressing the transcription factors GmMYB29A2 and GmNAC42-1. (A) Venn diagrams of up and downregulated genes. (B) Percentage of ontologies of genes upregulated (top) and downregulated (bottom) by WGE. (C) Percentage of ontologies of genes upregulated (top) and downregulated (bottom) by p35S::GmMYB29A2. (D) Percentage of ontologies of genes upregulated (top) and downregulated (bottom) by p35S::GmNAC42-1. Ontology analysis was conducted using the (https://soybase.org/, accessed on 20 December 2022) Gene model Data Mining and Analysis tool.
Ontological analysis of the genes up and downregulated by WGE found ‘Protein Binding’ and ‘DNA binding’ were the most highly represented functionalities in those gene-sets (Figure 3B). They were also most highly represented in p35S::GmMYB29A2 and p35S::GmNAC42-1 gene-sets (Figure 3C,D). Perhaps not surprisingly, a similar analysis of the genes commonly regulated by WGE and p35S::GmMYB29A2, WGE and p35S::GmNAC42-1, and all three genotypes demonstrated similar distributions of ontologies (Supplementary Figure S1). This demonstrated that the WGE, p35S::GmMYB29A2, and p35S::GmNAC42-1 broadly regulated genome functions in a proportionally similar manner. The ontology ‘Catalytic Activity’ comprised 8–12% of the downregulated genes in each genotype (Figure 3B–D). In contrast, ‘Catalytic Activity’ represented 7% of the genes that were upregulated by each genotype. These included glyceollin biosynthesis genes.
2.3. Overexpressing GmMYB29A2 or GmNAC42-1 in the Absence of an Elicitor Treatment Fails to Upregulate Several Glyceollin Biosynthesis Genes That Are Regulated by WGE
To understand how overexpressing p35S::GmMYB29A2 or p35S::GmNAC42-1 in the absence of an elicitor treatment fails to activate glyceollin biosynthesis, we conducted an in-depth comparison of biosynthetic gene expressions among mock-treated roots overexpression those transcription factors and empty vector hairy roots treated with WGE. We compiled a list of 338 genes annotated to encode enzymes spanning the 43 enzyme-catalyzed steps from D-glucose to the formation of glyceollins (Figure 1, Supplementary Table S7). This large number of genes was justifiable due to the palaeopolyploid genome of soybean [49], where several genes encode most enzymes. For example, there are 12 chalcone isomerase (CHI) genes and a similar number of chalcone synthase (CHS) genes with distinct expression profiles, subcellular localizations, and sub-functionalities [50,51]. WGE upregulated 87 genes for 33 of those enzymes (Figure 4A). The nine enzymes that WGE did not regulate were PGL, TKL1, DQS, and MEE32 from primary metabolism, EPSP, CM and ArAT from phenylpropanoid metabolism, and MTC and G2DT from the MEP and glyceollin pathways, respectively.
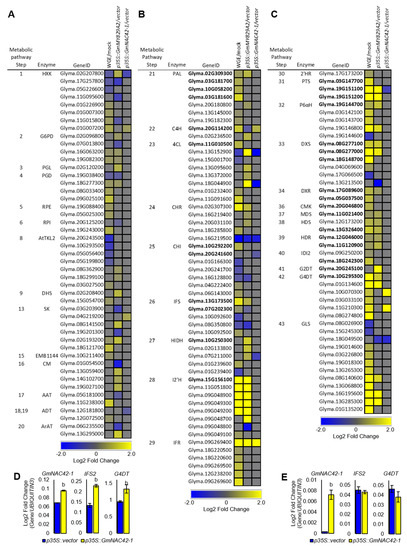
Figure 4.
Glyceollin biosynthesis genes that were differentially regulated in soybean W82 hairy roots by P. sojae wall glucan elicitor (WGE), p35S::GmMYB29A2, and p35S::GmNAC42−1. (A) Heat map of primary metabolism genes corresponding to enzymatic steps 1–20 (see Figure 1). (B) Heat map of phenylpropanoid and isoflavonoid genes corresponding to enzymatic steps 21–29. (C) Heat map of the gene regulation from the methylerythritol phosphate (MEP) and glyceollin pathways (steps 30–43). (D) Gene expression levels in 24 h WGE−treated hairy roots of GmNAC42−1. (E) Gene expression levels in 24 h mock−treated (H2O) hairy roots GmNAC42−1.
Genes for four primary metabolism enzymes were upregulated by WGE but not by p35S::GmMYB29A2 or p35S::GmNAC42-1. These were RPE, RPI, EMB1144, and ADT (Figure 4A). p35S::GmMYB29A2 upregulated genes for most of the same primary enzymes as WGE, yet it also upregulated genes for PGL and CM. In contrast, p35S::GmNAC42-1 upregulated only genes for HXK and SK, which WGE and p35S::GmMYB29A2 also upregulated.
WGE upregulated genes encoding most phenylpropanoid and isoflavonoid pathway enzymes (Figure 4B). p35S::GmMYB29A2 upregulated most of those enzymes except CHI. It also upregulated ArAT, which catalyzes an alternative route to L-phenylalanine that was not upregulated by WGE. In contrast, p35S::GmNAC42-1 upregulated only genes for C4H and IFR, and downregulated genes for 4Cl, CHR, and HIDH.
WGE upregulated genes for the vast majority of MEP and glyceollin I biosynthetic enzymes, including several monooxygenases that are candidates of glyceollin synthase (GLS) (Figure 4C). However, WGE did not upregulate the gene for G2DT, which is the branchpoint enzyme for glyceollin II biosynthesis [52]. p35S::GmMYB29A2 upregulated G2DT and genes for all other WGE-regulated enzymes except 2′HR, CMK, and IDI2. The p35S::GmNAC42-1, in contrast, failed to upregulated most MEP and glyceollin genes. It upregulated only two G4DT-like genes and downregulated genes for PTS and GLS.
Since GmNAC42-1 has an essential role in activating glyceollin biosynthesis, we tested whether p35S::GmNAC42-1 could upregulate glyceollin biosynthesis genes when roots were exposed to WGE, as previously reported [33]. Overexpressing GmNAC42-1 1.4-fold in WGE-treated hairy roots upregulates IFS2 and G4DT 1.7- and 1.4-fold, respectively (Figure 4D). However, overexpressing GmNAC42-1 22.6-fold in mock-treated roots failed to upregulate both biosynthesis genes (Figure 4E), validating the RNA-seq results presented here (Figure 4C).
Thus, we observed that overexpressing GmMYB29A2 in mock-treated roots upregulated genes for most, but not all enzymes that were upregulated by WGE treatment. In contrast, overexpressing GmNAC42-1 in mock-treated roots failed to upregulate the vast majority of those genes, even though it could upregulate some of them in elicited roots. Our results demonstrate that overexpressing GmMYB29A2 and GmNAC42-1 in the absence of an elicitor is insufficient to activate the transcription of all glyceollin biosynthesis genes, raising the possibility that one or more additional transcription factors are needed to activate the entire glyceollin biosynthesis pathway.
2.4. The WGE-Upregulated Transcription Factor GmHSF6-1 Directly Activates the Expression of Biosynthesis Genes HIDH and I2′H
Of the 2364 genes that were upregulated exclusively by WGE, 11% (260 genes) were annotated as DNA binding transcription factors (Figure 3A). To search for additional activators of glyceollin biosynthesis, we compared those genes to our previously published RNA-seq datasets [2,33]. We narrowed down this list to 100 transcription factor genes by including only those transcription factors that were also significantly upregulated by WGE treatment in Harosoy63 seeds [2] (Supplementary Table S8). Finally, we reduced the list to 22 transcription factor genes by including only those also upregulated by acidity stress and downregulated by dehydration, which represent an elicitor and a suppressor of glyceollin biosynthesis, respectively [33]. Of those 22 genes, the heat shock family gene GmHSF6-1 (Glyma.03G135800) was selected for functional characterization since it demonstrated homology to VIT_208s0007g08750 that is co-regulated with stilbene phytoalexin biosynthesis genes in grapevine [36].
The qRT-PCR confirmed that GmHSF6-1 was upregulated by WGE treatment (Figure 5A). The G. max RNA-seq Atlas (https://soybase.org/, accessed on 20 December 2022) demonstrated that the expression of GmHSF6-1 did not follow developmentally regulated patterns of HIDH or the isoflavonoid regulator GmMYB176 and was more similar to G4DT and the glyceollin regulators GmMYB29A2, GmNAC42-1 (Figure 5B). The predicted peptide sequence of GmHSF6-1 consisted of 231 amino acids and a three-component HSF DNA binding domain (DBD) (Figure 5C). Phylogenetic analysis demonstrated that GmHSF6-1 and four other soybean proteins clustered as a subgroup with the Arabidopsis proteins AtHSFB2a and AtHSFB2b (Figure 5D). The peptide had 55% and 49% similarity to the Arabidopsis proteins HSFA2a (AT5G62020) and HSFA2b (AT4G11660), respectively, which notably have roles in regulating biotic and environmental stress responses [53,54].
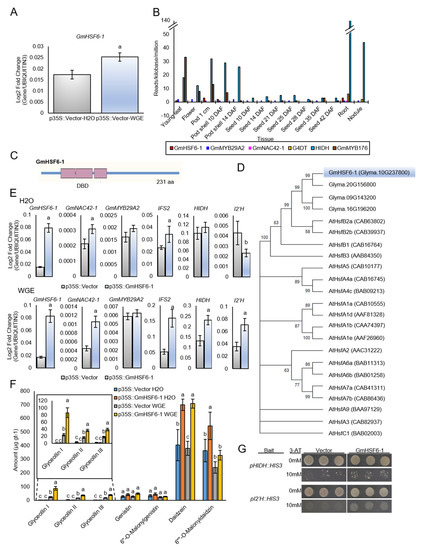
Figure 5.
The P. sojae wall glucan elicitor (WGE)−upregulated transcription factor GmHSF6−1 directly activates the expression of glyceollin biosynthesis genes. (A) Expression level of GmHSF6−1 gene in W82 hairy roots elicited for 24 h with WGE. (B) Gene expressions in developing soybean organs from the RNA−Seq Atlas of G. max (https://soybase.org/, accessed on 20 December 2022). GmMYB29A2, GmNAC42−1, and G4DT are markers of stress inducible glyceollin biosynthesis, whereas GmMYB176 and HIDH are markers of developmentally regulated isoflavonoids. (C) Schematic diagram of GmHSF6−1 gene indicating the DNA binding domain (DBD) region and coding sequence. (D) Cluster analysis of deduced amino sequences of GmHSFs with Arabidopsis AtHsfs. (E) Gene expressions in p35S::GmHSF6−1 W82 hairy roots elicited for 24 h with H2O and WGE. a Significantly greater and b significantly less than control, paired students t−test (p < 0.01). Error bars represent SE (n ≥ 3). (F) Amounts of glyceollins and isoflavonoid metabolites from hairy roots overexpressing GmHSF6−1 24 h after treatment with WGE or H2O. Letters indicate significantly different levels determined by single factor ANOVA, Tukey post hoc test, p ≤ 0.01. (G) Yeast one−hybrid assays of YM4271 yeast transformed with GmHSF6−1 and HIDH/I2′H promoters on selection medium (SD−Leu−His+3AT). Representative values of two independent experiments are shown.
To assess the potential involvement of GmHSF6-1 in regulating glyceollin biosynthesis, the coding sequence was cloned from the cDNA of WGE-treated W82 hairy roots and was transferred downstream of the p35S viral promoter in the plant gene expression vector. Overexpressing GmHSF6-1 5.1-fold in mock-treated W82 hairy roots resulted in a 1.5-fold upregulation of GmNAC42-1, but this was not accompanied by any changes in the expression of five glyceollin biosynthesis genes and GmMYB29A2 by qRT-PCR (Figure 5E). Metabolite measurements also found no difference in the total amounts of glyceollins, but the roots over accumulated their biosynthetic intermediate daidzein 1.7-fold compared to the empty vector control (Figure 5F). In contrast, overexpressing GmHSF6-1 4.8-fold in WGE-treated hairy roots resulted in a 1.7- to 3.1-fold upregulation of the biosynthesis genes IFS2, HIDH, and I2′H (Figure 5E) and a 2.2- to 3.6-fold increase in the amounts of glyceollin I, II, and III (Figure 5F). To investigate whether GmHSF6-1 is a direct regulator of glyceollin biosynthesis genes, we conducted yeast one-hybrid analysis by fusing GmHSF6-1 to the yeast Gal4 activation domain (AD) and by integrating the promoters of HIDH and I2′H upstream of the HIS3 gene in the yeast genome. Both yeast strains expressing AD-GmHSF6-1 fusions could grow on medium lacking histidine (Figure 5G), indicating that GmHSF6-1 was physically binding those promoters. The results demonstrate that GmHSF6-1 is a direct activator of those glyceollin biosynthesis genes.
3. Discussion
The development of stable approaches for the production of phytoalexins has been ongoing for decades. These approaches include (semi)synthesis, culturing microbes that ectopically express phytoalexin biosynthesis genes in large bioreactors, combining elicitor treatments to elicit maximum biosynthesis in plant tissues, and overexpressing rate-limiting biosynthetic genes in plants [55,56,57]. Yet, each of these has suffered shortcomings [56,58,59]. In plants, the biosynthesis of phytoalexins is primarily regulated at the level of transcription of the biosynthetic genes [19,20,21]. This feature makes the transcription factors that regulate those genes of prime importance for enhancing phytoalexin biosynthesis [21,22,23]. However, only with the advent of next generation sequencing approaches, such as transcriptome sequencing (RNA-seq) and chromatin immunoprecipitation genome-wide sequencing (ChIP-seq), can we begin to understand how phytoalexin gene regulatory networks can be reprogrammed to unlock phytoalexin biosynthesis [22].
Our research has been focusing on understanding the control the glyceollin biosynthesis in soybean as a model phytoalexin gene regulatory network. We recently identified two essential activators of glyceollin biosynthesis, namely GmMYB29A2 and GmNAC42-1 [2,33]. RNAi silencing in soybean hairy roots demonstrated that two transcription factors were required to activate glyceollin biosynthesis and they were found to directly bind the promoters of essential biosynthesis genes, namely G4DT and IFS2, in vitro and in the yeast one-hybrid system. However, independently overexpressing each gene failed to activate glyceollin biosynthesis without an elicitor treatment [2,33]. In this study, we tested whether co-overexpressing both transcription factors could activate glyceollin biosynthesis in the absence of elicitation, but it could not (Figure 2). To understand why co-overexpressing both transcription factors could not activate glyceollin biosynthesis without elicitation, we compared the transcriptomes of WGE-treated empty-vector-transformed hairy roots that were biosynthesizing glyceollins to mock-treated hairy roots overexpressing GmMYB29A2 or GmNAC42-1 (Figure 2 and Figure 3). WGE upregulated 87 genes for 33 of the biosynthetic enzymes (Figure 4). This suggested that upregulating genes for these 33 enzymes may be needed to achieve glyceollin biosynthesis. Our results demonstrated that overexpressing GmMYB29A2 in mock-treated roots upregulated genes for all but eight enzymes that were regulated by WGE. These were for the primary metabolism enzymes RPE, RPI, EMB1144, and ADT, the isoflavonoid enzyme CHI, and the glyceollin/MEP pathway enzymes 2′HR, CMK, and IDI2 (Figure 4). Thus, overexpressing GmMYB29A2 alone activates most, but not all genes needed to activate glyceollin biosynthesis. The current model comparing WGE versus GmMYB29A2 regulations is shown in Figure 6.
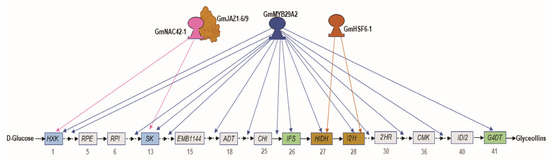
Figure 6.
A schematic diagram of WGE-regulated glyceollin biosynthesis genes that are, and are not, upregulated by the transcription factors GmMYB29A2, GmNAC42-1, and GmHSF6-1 when each is overexpressed in the absence of an elicitor treatment. Solid and dotted black arrows indicate single and multiple enzyme-coding genes, respectively. Blue and pink arrows indicate genes that are upregulated by GmMYB29A2 and GmNAC42-1 via RNA-seq. Red arrows indicate genes that were upregulated by GmHSF6-1 by qRT-PCR. Genes that have been characterized to directly interact with GmMYB29A2, GmNAC42-1, and GmHSF6-1 are shown as blue, purple, and orange boxes, respectively. Note, only those genes have been assessed for protein DNA interactions. Gray boxes represent WGE-regulated genes that were not upregulated by overexpressing GmMYB29A2, GmNAC42-1, or GmHSF6-1. The interaction of GmJAZ1-6/-9 inhibits GmNAC42-1′s interaction with gene targets.
The transcription factor VvMYB14 from grapevine (Vitis vinifera) was recently reported to regulate biosynthetic steps for stilbene phytoalexin biosynthesis, including primary metabolism genes [36]. GmMYB29A2 shows the highest amino acid similarity to VvMYB14 among all grapevine proteins [2,35]. Additionally, genes encoding those two are syntenic with malate dehydrogenase on chromosomes 2 and 7 of soybean and grapevine, respectively. Together, this demonstrates that GmMYB29A2 and VvMYB14 are homologs. DNA affinity purification sequencing (DAP-Seq) combined with RNA-seq gene co-expression networks (GCN) analysis identified direct targets of VvMYB14 [36]. Those targets included biosynthesis genes that were also upregulated by GmMYB29A2 overexpression, such as the primary metabolism genes for shikimate kinase (SK, biosynthetic step 13 in Figure 1) and chorismate mutase (CM, step 16), and phenylpropanoid pathway enzymes phenylalanine ammonia-lyase (PAL, step 21), cinnamate 4-hydroxylase (C4H, step 22) and 4-coumarate-CoA ligase (4CL, step 23) (Figure 4). However, our yeast one-hybrid and electrophoretic mobility shift assays (EMSAs) have shown that GmMYB29A2 directly binds the promoters of isoflavone synthase (IFS) and glycinol-4-dimethylallyltransferase (G4DT) [2], which have no obvious homologs in grapevine which does not biosynthesize glyceollins. Likewise, VvMYB14 directly binds stilbene synthase (STS) gene promoters in grapevine [35,36], and there have been no reports that soybean biosynthesizes stilbenoids. Thus, taken together with [36], our results strongly suggest that GmMYB29A2 and VvMYB14 have maintained roles in regulating several primary and early phenylpropanoid genes, but also have coopted distinct downstream genes for glyceollin and stilbene biosynthesis, respectively.
Since isoflavonoids and phenylpropanoids are the most abundant specialized metabolites that accumulate without stress in soybean and grapevine, GmMYB29A2 and VvMYB14 likely faced evolutionary pressure to coopt biosynthesis genes that could convert available metabolic intermediates into phytoalexins that are toxic to pathogens. The R2 R3 DNA binding domains of GmMYB29A2 and VvMYB14 are 89% similar/identical at the amino acid level (Supplementary Figure S2), so the consensus DNA binding sequence bound by each factor is likely highly similar. DAP-seq suggested the VvMYB14-binding sequence [36]. However, VvMYB13, VvMYB14, and VvMYB15 have identical DNA binding domains and do not regulate identical gene-sets [36]. Thus, gene targets of VvMYB14-type proteins such as GmMYB29A2 are likely influenced by their interactions with other proteins, including other transcription factors. This could explain why GmMYB29A2 positively regulates G4DT for the biosynthesis of glyceollin I during elicitation [2], but instead regulates G2DT for the biosynthesis of glyceollin II in the absence of an elicitor (Figure 4).
Results on GmNAC42-1 suggest a similar scenario. GmNAC42-1 upregulated IFS2 and G4DT when overexpressed in WGE-treated hairy roots (Figure 4D). However, it failed to upregulate those genes in the absence of WGE treatment (Figure 3E). The latter is consistent with our RNA-seq results, where overexpressing GmNAC42-1 without an elicitor failed to upregulate most glyceollin biosynthesis genes (Figure 3C). In a parallel study in our lab searching for genes that were oppositely regulated compared to glyceollin biosynthesis, we identify GmJAZ1-6 and GmJAZ1-9 as negative regulators of glyceollin biosynthesis [60]. These two proteins physically interact with GmNAC42-1 and block its activation of glyceollin biosynthesis gene promoters in the yeast three-hybrid and promoter-luciferase reporter systems [60]. Notably, silencing GmJAZ1s results in some accumulation of glyceollins in the absence of an elicitor. Thus, GmJAZ1 genes likely need to be silenced for GmNAC42-1 overexpression to activate most glyceollin biosynthesis gene targets. Despite that p35S::GmNAC42-1 regulated numerous genes (Figure 2), only two primary metabolism genes that contribute to glyceollin biosynthesis were upregulated in the absence of an elicitor treatment (Figure 6). Thus, GmNAC42-1 likely has very different roles in the presence and absence proteins expressed in response to an elicitor treatment.
Unlocking the function of GmNAC42-1 by silencing GmJAZ1 gene expressions may be insufficient to activate all glyceollin biosynthesis genes, and additional transcriptional activators regulated by WGE may be required. The combinatorial action of GmMYB176 (Glyma.19g214900) and GmbZIP5 (Glyma.05G122400) controls the biosynthesis of developmentally regulated isoflavonoids in soybean [61] and GmMYB133 (Glyma.07G066100) modulates the biosynthetic pathway [62]. Yet, neither WGE, p35S::GmMYB29A2, or p35S::GmNAC42-1 upregulated those genes in W82 hairy roots (Supplementary Tables S1, S2 and S8). This study identified the WGE-upregulated GmHSF6-1 as a direct positive regulator of HIDH and I2′H (Figure 5). Thus, GmHSF6-1 likely cooperates with GmMYB29A2 to fully activate the expression of those genes.
GmHSF6-1 belongs to the heat shock factor family of transcription factors and compared to Arabidopsis proteins it shows highest similarity to HsfB2a (Figure 5). Plant HSFs of subgroup B are generally involved in non-heat shock functions [63]. HsfB2a positively regulates the expression of defensin genes in response to insects and microbial pathogens [64,65]. In vitro and agroinfiltration-based kinase assays have demonstrated that the Ca2+-dependent protein kinases (CPK3 and CPK13) phosphorylate HsfB2a and that is needed for full transcriptional activation of its defensin gene targets [64,65]. The same study demonstrated that CPK3 also phosphorylates ethylene response factor ERF1. In Arabidopsis upon pathogen infection, ERF1 functions to integrate ethylene and jasmonate (JA) signaling by directly activating camalexin phytoalexin biosynthesis genes [28]. ERF1 needs to interact physically with WRKY33 to directly bind and upregulate camalexin biosynthetic genes [28]. Homologs of both ERF1 and WRKY33 were found to be upregulated by WGE but not p35S::GmMYB29A2 or p35S::GmNAC42-1 (Supplementary Table S8), indicating that they may be upstream of those transcription factors, if involved in activating glyceollin biosynthesis. We have discovered that GmNAC42-1- and GmJAZ1-type proteins have conserved roles in regulating glyceollin biosynthesis [33,60], as their homologs do in regulating camalexin biosynthesis in Arabidopsis [31,34]. Thus, HSF6-1 could be another member of a conserved transcription factor network that regulates distinct phytoalexin biosynthetic pathways among plant species.
4. Materials and Methods
4.1. Chemicals
Stocks (50 mg/mL) of the antibiotics including kanamycin, timentin, hygromycin-B, and ampicillin (Gold Biotechnology, Olivette, MO, USA) were prepared in MilliQ-purified water. Acetosyringone stock (Sigma-Aldrich, St. Louis, MO, USA) was 100 mM in dimethyl sulfoxide. (−)-Glyceollin I was purchased from Dr. Paul Erhardt (University of Toledo). Isoflavone standards were purchased from Extrasynthese (Genay, France). Other reactants were purchased from Sigma-Aldrich. Ultra-performance liquid chromatography (UPLC) solvents were liquid chromatography-mass spectrometry grade from Fisher (Hampton, NH, USA). WGE was extracted from race 1 Phytophthora sojae cultured on lima bean medium for three weeks as described previously [66].
4.2. Plant Materials
Soybean seeds of Williams 82 (W82) were obtained from the USDA-GRIN Soybean Germplasm Collection (Beltsville, MD, USA). The surface of W82 seeds (16–20 per batch) were sterilized with [70% isopropanol (v/v; 30 s), 6% sodium hypochlorite (v/v; 5 min), and distilled water (rinsed three times)] and imbibed overnight before use. For hairy roots, seeds were grown for 7 days [3 days at dark; 4 days under cool white T5 fluorescent lights (100 μEm2/s)] on germination and co-cultivation (GC) medium followed by the removal of the seed coat, and the distal end (~2 to 3 mm) of the cotyledons were excised as described by [2].
4.3. RNA Extraction and Gene Expression Measurements by qRT-PCR
Total RNA from soybean tissue was extracted using the Spectrum Plant Total RNA Kit (Sigma-Aldrich, St. Louis, MO, USA) following the manufacturer’s protocol. RNA-seq analysis was described in [2,33]. DNase I (Amplification grade, Invitrogen, Waltham, MA, USA) treatment of the total RNA (500 ng) was completed to remove genomic DNA contamination before complementary DNA (cDNA) synthesis, and SuperScript II Reverse Transcriptase (Invitrogen, Waltham, MA, USA) was used to synthesize cDNA following the manufacturers protocol. SsoAdvanced Universal SYBR Green Supermix (BioRad, Hercules, CA, USA) was used to carry out qRT-PCR of diluted cDNA templates as described previously [66]. Briefly, reactions (5 μL) consisted of 2 μL of first-strand cDNA (or untreated RNA), 0.5 μL of 5 mM forward and reverse primers, 0.5 μL of RNase free H2O, and 2 μL of the iQ SYBR Green Supermix (BioRad, Hercules, CA, USA). CFX Opus 96 Real-Time PCR System (BioRad, Hercules, CA, USA) was used to perform qRT-PCR on cDNA from three biological replicates or RNA that were not reverse transcribed to measure for genomic DNA contamination. The conditions of the qRT-PCR were as follows: initial denaturation at 95 °C for 3 min, followed by 39 cycles of 95 °C for 15 s and 60 °C for 30 s. Normalized gene expression was calculated based on cycle threshold (Ct) values using the formula expression = 2^ − [Ct(gene) − Ct(UBIQUITIN3)]. To verify the specificity of the qRT-qPCR reactions, melting curves were determined after each reaction. Primers used in this study are listed in Supplementary Table S9.
4.4. Transcriptome Data Analysis
The RNA sequencing and analysis of four biological replicates of WGE- and water-treated tissues of W82 hairy roots was reported in [2]. The data are available in the Gene Expression Omnibus (GEO) database at the National Center for Biotechnology Information (NCBI) under the accession number GSE131686. The same method from [2] was used to analyze three biological replicates of H2O-treated W hairy roots of p35S::GmMYB29A2, p35S::GmNAC42-1, and the p35S empty vector (pGWB2) control. Those data are available in GEO under the accession number GSE221901.
For analysis of glyceollin biosynthesis gene expressions, the enzyme and their corresponding Glyma gene accession numbers were obtained from the PlantCyc database of the Plant Metabolic Network (PMN), except glyceollin synthase (GLS). Cytochrome p450 monooxygenases [67] that were upregulated by WGE treatment were considered candidates of GLS. Any glyceollin biosynthesis gene that were significantly up or downregulated (adjusted p < 0.05) in any RNA-seq comparison (p35S::GmMYB29A2/vector, p35S::GmNAC42-1/vector, or WGE/mock), were compared by heat maps.
4.5. Cloning
The GmMYB29A2, GmNAC42-1, and GmHSF6-1 coding sequences were PCR amplified from the cDNA of WGE-treated Willliams 82 hairy roots and BP cloned into pDONR221. Primers are listed in Supplementary Table S9. For co-overexpression, the donor clones were simultaneously LR recombined into the dual overexpression vector pGEMINI [48]. GmHSF6-1-pDONR221 was LR cloned into pGWB2 and pDEST-GADT7 for overexpression and yeast one-hybrid assays. HIDH and I2′H promoter regions were cloned into the BsaI site of the pGG vector that is flanked by attL4 and attR1 recombination sites and LR recombined into the destination vector pMW2.
4.6. Hairy Root Transformation and Elicitation
Agrobacterium rhizogenes strain K599 containing the empty vector and the overexpression constructs were used to transform soybean cotyledons using a previously described protocol [33] with slight modifications. Briefly, A. rhizogenes harboring the construct was grown overnight on plates containing Luria-Bertani (LB) medium, hygromycin, and kanamycin (50 mg/L), and cells from the plate were resuspended in phosphate buffer (pH 7.5) containing 100 μM acetosyringone to an OD600 of 0.5–0.8 before use. Soybean cotyledons were inoculated through several 1 mM-deep cuts on the adaxial surface of the cotyledon with a scalpel dipped in the A. rhizogenes suspension. The cotyledons were grown for 3 days under a 16 h photoperiod (~65 μE) in GC medium containing 100 μM acetosyringone and then transferred into hairy root growth (HRG) medium containing timentin (500 mg/L). Primary roots with secondary roots (2–3 cm) were grown for 5 to 7 days in HRG plates containing kanamycin and hygromycin (50 mg/L). Transgenic hairy roots (secondary roots that grew 3 to 6 cm on selection HRG plates) were harvested and cut into 1 cm pieces for WGE or water treatments. Secondary roots from a single primary root represented one biological replicate since each primary root is derived from a distinct transformation event. The excised 1 cm pieces of secondary roots were placed in HRG plates lacking antibiotics and then overlaid with 80 μL of WGE (20 mg/mL) or water. The hairy roots were then incubated for 24 h under cool white T5 fluorescent lights (500 μEm2/s). Treated roots were used directly for metabolite extractions or were harvested into liquid nitrogen and lyophilized for 3 to 5 d (BenchTop Pro, SP Scientific, Warminster, PA, USA) before storage at −80 °C for RNA extraction.
4.7. Metabolite Analyses
Treated roots (90–110 mg) were harvested and extracted according to [66]. The extracts were analyzed using a Vanquish UPLC system (Thermo Scientific, Waltham, MA, USA) configured with a Quaternary Pump F, Split Sampler FT Autosampler, and Diode Array Detector HL (DAD). The UPLC-DAD method and quantification of isoflavonoids using Chromeleon 7.2.10 software (Thermo Scientific, Waltham, MA, USA) was conducted according to [33]. Isoflavonoid amounts were measured from five biological replicates. Secondary roots from a single primary root represented one biological replicate since each primary root is derived from a distinct transformation event. Hairy roots for all treatments were generated from the same batch of seeds to avoid differences caused by the seed age. Roots harboring gene overexpression constructs and their corresponding empty vector controls were treated and harvested at the same time to prevent inter-day experimental variation.
4.8. Yeast One-Hybrid
Bait strains of Saccharomyces cerevisiae YM4271 (MATa, ura3–52, lys2–801, ade2–101, ade5, trp1–901, leu2–3, 112, tyr1–501, gal4D, gal80D, ade5::hisG) that integrate HIDH and I2′H promoter regions upstream of the HIS3 gene were prepared as described by [33] and transformed with either pDEST-GADT7 or GmHSF6-1-pDEST-GADT7. Bait-pray strains were selected in media lacking both histidine and leucine and confirmed by PCR. Positive DNA-protein interactions were assessed by comparing differential growth in the presence of a competitive inhibitor of HIS3 enzyme (3-Amino-1,2,4-triazol, 3AT). Three biological replicates confirmed by two independent experiments are shown in results.
4.9. Statistics
To assess for any significant statistical differences between groups, the Tukey post hoc test in one-way ANOVA was applied between treatments and genotypes. Significant differences are shown as different letters in each box (at α = 0.05).
4.10. Assession Numbers
Accession numbers are as follows: GmMYB29A2, Glyma.02G005600; GmNAC42-1, Glyma.02G284300; GmHSF6-1, Gyma.03G135800; HIDH, Glyma.01G239600; I2′H, Glyma.15G156100.
5. Conclusions
The elicitor molecule WGE from the pathogen P. sojae upregulates 87 putative enzyme-coding genes spanning primary metabolism to glyceollin biosynthesis in soybean hairy roots. Overexpressing the transcription factor GmMYB29A2 upregulated genes for most, but not eight enzymes. The transcription factor GmNAC42-1 fails to upregulate most genes in the absence of WGE, coinciding with its recently discovered physical interaction with the negative regulators GmJAZ1-6/9. We discovered that the WGE-upregulated transcription factor GmHSF6-1 is also a direct positive regulator of several glyceollin biosynthesis genes. Our current model (Figure 6) suggests that multiple activators and removing GmNAC42-1′s suppression by GmJAZ1-6/9 will be needed to activate the transcription of all glyceollin biosynthesis genes.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants12030545/s1, Figure S1. Ontologies of genes co-regulated and exclusively regulated by WGE. (A) Percentage of ontologies of genes upregulated (top) and downregulated (bottom) by WGE and p35S::GmMYB29A2. (C) Percentage of ontologies of genes upregulated (top) and downregulated (bottom) by WGE and p35S::GmNAC42-1. (D) Percentage of ontologies of genes upregulated (top) and downregulated (bottom) by WGE only, p35S::GmMYB29A2 and p35S::GmNAC42-1. Ontology analysis was conducted using the SoyBase.org Gene model Data Mining and Analysis tool. Figure S2. Similarity and identity among GmMYB29A2 and VvMYB14 amino acid sequences. Table S1. Genes commonly upregulated by WGE and p35S-GmMYB29A2. Table S2. Genes commonly upregulated by WGE and p35S-GmNAC42-1. Table S3. Genes commonly upregulated by WGE, p35S-GmMYB29A2 and p35S-GmNAC42-1. Table S4. Genes commonly downregulated by WGE and p35S-GmMYB29A2. Table S5. Genes commonly downregulated by WGE and p35S-GmNAC42-1. Table S6. Genes commonly downregulated by WGE, p35S-GmMYB29A2 and p35S-GmNAC42-1. Table S7. Biosynthetic genelist. Table S8. Narrowing down glyceollin transcription factor gene candidates. Table S9. Primers list.
Author Contributions
N.K. conceived and designed the experiments. J.L., M.A.J., I.M. (Ivan Monsalvo) and M.L. performed the experiments. J.L., I.M. (Ivan Monsalvo), I.M. (Izabella Martirosyan) and D.W., N.K. analyzed the data. N.K., J.L. and I.M. (Ivan Monsalvo) wrote the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Natural Sciences and Engineering Research Council of Canada (NSERC), Funding Number RGPIN-2020-06111, and by the USDA National Institute of Food and Agriculture (NIFA; hatch, accession no. 1010200). Jie Lin was funded by the China Scholarship Council (CSC, 202107980003), Ivan Monsalvo and Dasol Wi by York University’s Dean’s Undergraduate Research Award (DURA).
Data Availability Statement
All data supporting the finding of this study are available within the paper, Supplementary Materials and the GEO under the accession number GSE221901.
Acknowledgments
We would like to thank Ryan Percifield and Niel Infante (West Virginia University) and James Denvir and Don Primerano of the Marshall University Genomics Core Facility (RRID:SCR_018885) for their assistance with RNA-seq. N.K. would like to thank Brad Lace for the generous donation.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Desmedt, W.; Kudjordjie, E.N.; Chavan, S.N.; Zhang, J.; Li, R.; Yang, B.; Nicolaisen, M.; Mori, M.; Peters, R.J.; Vanholme, B. Rice diterpenoid phytoalexins are involved in defence against parasitic nematodes and shape rhizosphere nematode communities. New Phytol. 2022, 235, 1231–1245. [Google Scholar] [CrossRef] [PubMed]
- Jahan, M.A.; Harris, B.; Lowery, M.; Infante, A.M.; Percifield, R.J.; Kovinich, N. Glyceollin Transcription Factor GmMYB29A2 Regulates Soybean Resistance to Phytophthora sojae. Plant Physiol. 2020, 183, 530–546. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.H.; Trotel-Aziz, P.; Clément, C.; Jeandet, P.; Baillieul, F.; Aziz, A. Camalexin accumulation as a component of plant immunity during interactions with pathogens and beneficial microbes. Planta. 2022, 255, 116. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, I.; Kissen, R.; Bones, A.M. Phytoalexins in defense against pathogens. Trends Plant Sci. 2012, 17, 73–90. [Google Scholar] [CrossRef]
- Fang, C.; Fernie, A.R.; Luo, J. Exploring the diversity of plant metabolism. Trends Plant Sci. 2019, 24, 83–98. [Google Scholar] [CrossRef]
- Großkinsky, D.K.; van der Graaff, E.; Roitsch, T. Phytoalexin transgenics in crop protection—Fairy tale with a happy end? Plant Sci. 2012, 195, 54–70. [Google Scholar] [CrossRef]
- Sharma, I.; Thakur, A.; Sharma, A.; Singh, N.; Kumar, R.; Sharma, A. Phytoalexins: Implications in Plant Defense and Human Health. In Plant Secondary Metabolites; Springer: Berlin/Heidelberg, Germany, 2022; pp. 329–353. [Google Scholar]
- Wang, C.; Wu, J.; Mei, X. Enhancement of taxol production and excretion in Taxus chinensis cell culture by fungal elicitation and medium renewal. Appl. Microbiol. Biotechnol. 2001, 55, 404–410. [Google Scholar] [CrossRef]
- Dittrich, H.; Kutchan, T.M. Molecular cloning, expression, and induction of berberine bridge enzyme, an enzyme essential to the formation of benzophenanthridine alkaloids in the response of plants to pathogenic attack. Proc. Natl. Acad. Sci. USA 1991, 88, 9969–9973. [Google Scholar] [CrossRef]
- Li, J.; Zhao, G.-Z.; Varma, A.; Qin, S.; Xiong, Z.; Huang, H.-Y.; Zhu, W.-Y.; Zhao, L.-X.; Xu, L.-H.; Zhang, S. An endophytic Pseudonocardia species induces the production of artemisinin in Artemisia annua. PLoS ONE. 2012, 7, e51410. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.-M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Ohanenye, I.C.; Okagu, I.U.; Ogunrinola, O.A.; Agboinghale, P.E.; Udenigwe, C.C. Soybean Glyceollins and Human Health. In Phytochemicals in Soybeans; CRC Press: Boca Raton, FL, USA, 2022; pp. 169–184. [Google Scholar]
- Pham, T.H.; Lecomte, S.; Efstathiou, T.; Ferriere, F.; Pakdel, F. An update on the effects of glyceollins on human health: Possible anticancer effects and underlying mechanisms. Nutrients. 2019, 11, 79. [Google Scholar] [CrossRef] [PubMed]
- Walker, R.R.; Patel, J.R.; Gupta, A.; Davidson, A.M.; Williams, C.C.; Payton-Stewart, F.; Boué, S.M.; Burow, M.E.; Khupse, R.; Tilghman, S.L. Glyceollins Trigger Anti-Proliferative Effects in Hormone-Dependent Aromatase-Inhibitor-Resistant Breast Cancer Cells through the Induction of Apoptosis. Int. J. Mol. Sci. 2022, 23, 2887. [Google Scholar] [CrossRef] [PubMed]
- Isaac, I.C.; Johnson, T.J.; Berhow, M.; Baldwin, E.L.; Karki, B.; Woyengo, T.; Gibbons, W.R. Evaluating the efficacy of fungal strains to stimulate glyceollin production in soybeans. Mycol. Prog. 2017, 16, 223–230. [Google Scholar] [CrossRef]
- Zavadil, A. Developing Microbial Based Process to Produce High Value Natural Antimicrobial (Glyceollin) in Soybeans and Pullulan from Sucrose; South Dakota State University: Brookings, SD, USA, 2020. [Google Scholar]
- Seo, J.Y.; Kim, B.R.; Oh, J.; Kim, J.-S. Soybean-Derived Phytoalexins Improve Cognitive Function through Activation of Nrf2/HO-1 Signaling Pathway. Int. J. Mol. Sci. 2018, 19, 268. [Google Scholar] [CrossRef]
- Vannozzi, A.; Wong, D.C.J.; Höll, J.; Hmmam, I.; Matus, J.T.; Bogs, J.; Ziegler, T.; Dry, I.; Barcaccia, G.; Lucchin, M. Combinatorial regulation of stilbene synthase genes by WRKY and MYB transcription factors in grapevine (Vitis vinifera L.). Plant Cell Physiol. 2018, 59, 1043–1059. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, B.; Song, Y.; Chen, Y.; Fu, J.; Liu, J.; Ma, T.; Xi, Z.; Liu, H. Genome-wide (ChIP-seq) identification of target genes regulated by WRKY33 during submergence stress in Arabidopsis. BMC Genom. Data 2021, 22, 16. [Google Scholar] [CrossRef]
- Zhang, M.; Chen, Y.; Nie, L.; Jin, X.; Liao, W.; Zhao, S.; Fu, C.; Yu, L. Transcriptome-wide identification and screening of WRKY factors involved in the regulation of taxol biosynthesis in Taxus chinensis. Sci. Rep. 2018, 8, 5197. [Google Scholar] [CrossRef]
- Ahmed, S.; Kovinich, N. Regulation of phytoalexin biosynthesis for agriculture and human health. Phytochem. Rev. 2020, 20, 483–505. [Google Scholar] [CrossRef]
- Fu, X.; Peng, B.; Hassani, D.; Xie, L.; Liu, H.; Li, Y.; Chen, T.; Liu, P.; Tang, Y.; Li, L. AaWRKY9 contributes to light- and jasmonate-mediated to regulate the biosynthesis of artemisinin in Artemisia annua. New Phytol. 2021, 231, 1858–1874. [Google Scholar] [CrossRef]
- Liu, S.; Kracher, B.; Ziegler, J.; Birkenbihl, R.P.; Somssich, I.E. Negative regulation of ABA signaling by WRKY33 is critical for Arabidopsis immunity towards Botrytis cinerea 2100. eLife. 2015, 4, e07295. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Qamar, S.A.; Chen, Z.; Mengiste, T. Arabidopsis WRKY33 transcription factor is required for resistance to necrotrophic fungal pathogens. Plant J. 2006, 48, 592–605. [Google Scholar] [CrossRef] [PubMed]
- Mao, G.; Meng, X.; Liu, Y.; Zheng, Z.; Chen, Z.; Zhang, S. Phosphorylation of a WRKY transcription factor by two pathogen-responsive MAPKs drives phytoalexin biosynthesis in Arabidopsis. Plant Cell. 2011, 23, 1639–1653. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wang, X.; He, Y.; Sang, T.; Wang, P.; Dai, S.; Zhang, S.; Meng, X. Differential phosphorylation of the transcription factor WRKY33 by the protein kinases CPK5/CPK6 and MPK3/MPK6 cooperatively regulates camalexin biosynthesis in Arabidopsis. Plant Cell. 2020, 32, 2621–2638. [Google Scholar] [CrossRef]
- Zhou, J.; Mu, Q.; Wang, X.; Zhang, J.; Yu, H.; Huang, T.; He, Y.; Dai, S.; Meng, X. Multilayered synergistic regulation of phytoalexin biosynthesis by ethylene, jasmonate, and MAPK signaling pathways in Arabidopsis. Plant Cell. 2022, 34, 3066–3087. [Google Scholar] [CrossRef]
- Li, Y.; Liu, K.; Tong, G.; Xi, C.; Liu, J.; Zhao, H.; Wang, Y.; Ren, D.; Han, S.J. MPK3/MPK6-mediated phosphorylation of ERF72 positively regulates resistance to Botrytis cinerea through directly and indirectly activating the transcription of camalexin biosynthesis enzymes. J. Exp. Bot. 2022, 73, 413–428. [Google Scholar] [CrossRef]
- Jiang, Y.; Yu, D.J. The WRKY57 transcription factor affects the expression of jasmonate ZIM-domain genes transcriptionally to compromise Botrytis cinerea resistance. Plant Physiol. 2016, 171, 2771–2782. [Google Scholar] [CrossRef]
- Makhazen, D.; Veremeichik, G.; Shkryl, Y.; Tchernoded, G.; Grigorchuk, V.; Bulgakov, V. Inhibition of the JAZ1 gene causes activation of camalexin biosynthesis in Arabidopsis callus cultures. J. Biotechnol. 2021, 342, 102–113. [Google Scholar] [CrossRef]
- Makhazen, D.S.; Veremeichik, G.; Shkryl, Y.; Grigorchuk, V.; Tchernoded, G.; Degtyarenko, A.; Bulgakov, V. RNA inhibition of the JAZ9 gene increases the production of resveratrol in grape cell cultures. Plant Cell Tissue Organ Cult. 2021, 147, 611–618. [Google Scholar] [CrossRef]
- Jahan, M.A.; Harris, B.; Lowery, M.; Coburn, K.; Infante, A.M.; Percifield, R.J.; Ammer, A.G.; Kovinich, N. The NAC family transcription factor GmNAC42-1 regulates biosynthesis of the anticancer and neuroprotective glyceollins in soybean. BMC Genom. 2019, 20, 149. [Google Scholar] [CrossRef]
- Saga, H.; Ogawa, T.; Kai, K.; Suzuki, H.; Ogata, Y.; Sakurai, N.; Shibata, D.; Ohta, D. Identification and characterization of ANAC042, a transcription factor family gene involved in the regulation of camalexin biosynthesis in Arabidopsis. Mol. Plant Microbe. 2012, 25, 684–696. [Google Scholar] [CrossRef] [PubMed]
- Höll, J.; Vannozzi, A.; Czemmel, S.; D’Onofrio, C.; Walker, A.R.; Rausch, T.; Lucchin, M.; Boss, P.K.; Dry, I.B.; Bogs, J. The R2R3-MYB transcription factors MYB14 and MYB15 regulate stilbene biosynthesis in Vitis vinifera. Plant Cell 2013, 25, 4135–4149. [Google Scholar] [CrossRef] [PubMed]
- Orduña, L.; Li, M.; Navarro-Payá, D.; Zhang, C.; Santiago, A.; Romero, P.; Ramšak, Ž.; Magon, G.; Höll, J.; Merz, P. Direct regulation of shikimate, early phenylpropanoid, and stilbenoid pathways by Subgroup 2 R2R3-MYBs in grapevine. Plant J. 2022, 110, 529–547. [Google Scholar] [CrossRef]
- Jiang, J.; Xi, H.; Dai, Z.; Lecourieux, F.; Yuan, L.; Liu, X.; Patra, B.; Wei, Y.; Li, S.; Wang, L.J. VvWRKY8 represses stilbene synthase genes through direct interaction with VvMYB14 to control resveratrol biosynthesis in grapevine. J. Exp. Bot. 2019, 70, 715–729. [Google Scholar] [CrossRef] [PubMed]
- Sulieman, S.; Abdelrahman, M.; Tran, L.-S.P. Carbon metabolic adjustment in soybean nodules in response to phosphate limitation: A metabolite perspective. Environ. Exp. Bot. 2022, 196, 104810. [Google Scholar] [CrossRef]
- Wang, X.; Zhu, W.; Hashiguchi, A.; Nishimura, M.; Tian, J.; Komatsu, S. Metabolic profiles of flooding-tolerant mechanism in early-stage soybean responding to initial stress. Plant Mol. Biol. 2017, 94, 669–685. [Google Scholar] [CrossRef]
- Copley, T.R.; Aliferis, K.A.; Kliebenstein, D.J.; Jabaji, S.H. An integrated RNAseq-1H NMR metabolomics approach to understand soybean primary metabolism regulation in response to Rhizoctonia foliar blight disease. BMC Plant Biol. 2017, 17, 84. [Google Scholar] [CrossRef]
- Einhardt, A.M.; Oliveira, L.M.; Ferreira, S.; Araújo, W.L.; Medeiros, D.B.; Fernie, A.R.; Rodrigues, F.Á. Defense responses and oxidative metabolism of glyphosate-resistant soybean plants infected by Phakopsora pachyrhizi modulated by glyphosate and nickel. Physiol. Mol. Plant Pathol. 2022, 118, 101817. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Valliyodan, B.; Zhang, J.; Lenoble, M.E.; Yu, O.; Rogers, E.E.; Nguyen, H.T.; Sharp, R.E. Regulation of growth response to water stress in the soybean primary root. I. Proteomic analysis reveals region-specific regulation of phenylpropanoid metabolism and control of free iron in the elongation zone. Plant Cell Environ. 2010, 33, 223–243. [Google Scholar] [CrossRef]
- Gu, M.; Yang, J.; Tian, X.; Fang, W.; Xu, J.; Yin, Y. Enhanced total flavonoid accumulation and alleviated growth inhibition of germinating soybeans by GABA under UV-B stress. RSC Adv. 2022, 12, 6619–6630. [Google Scholar] [CrossRef]
- Yin, Y.; Tian, X.; He, X.; Yang, J.; Yang, Z.; Fang, W.J. Exogenous melatonin stimulated isoflavone biosynthesis in NaCl-stressed germinating soybean (Glycine max L.). Plant Physiol. Biochem. 2022, 185, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Malarz, J.; Michalska, K.; Yudina, Y.V.; Stojakowska, A. Hairy root cultures as a source of polyphenolic antioxidants: Flavonoids, stilbenoids and hydrolyzable tannins. Plants. 2022, 11, 1950. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.-S. Current perspectives on the beneficial effects of soybean isoflavones and their metabolites on plants. Antioxidants. 2022, 10, 1064. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, K.; Akashi, T.; Aoki, T.J. Molecular characterization of soybean pterocarpan 2-dimethylallyltransferase in glyceollin biosynthesis: Local gene and whole-genome duplications of prenyltransferase genes led to the structural diversity of soybean prenylated isoflavonoids. Plant Cell Physiol. 2016, 57, 2497–2509. [Google Scholar] [CrossRef]
- Exposito-Rodriguez, M.; Laissue, P.P.; López-Calcagno, P.E.; Mullineaux, P.M.; Raines, C.A.; Simkin, A.J. Development of pGEMINI, a plant gateway destination vector allowing the simultaneous integration of two cDNA via a single LR-clonase reaction. Plants. 2017, 6, 55. [Google Scholar] [CrossRef]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.X.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.J.; Thelen, J.J.; Cheng, J.L.; et al. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef]
- Anguraj Vadivel, A.K.; Krysiak, K.; Tian, G.; Dhaubhadel, S. Genome-wide identification and localization of chalcone synthase family in soybean (Glycine max [L] Merr). BMC Plant Biol. 2018, 18, 325. [Google Scholar] [CrossRef]
- Dastmalchi, M.; Bernards, M.; Dhaubhadel, S. Twin anchors of the soybean isoflavonoid metabolon: Evidence for tethering of the complex to the endoplasmic reticulum by IFS and C4H. Plant J. 2016, 85, 689–706. [Google Scholar] [CrossRef]
- Sukumaran, A.; McDowell, T.; Chen, L.; Renaud, J.; Dhaubhadel, S. Isoflavonoid-specific prenyltransferase gene family in soybean: Gm PT 01, a pterocarpan 2-dimethylallyltransferase involved in glyceollin biosynthesis. Plant J. 2018, 96, 966–981. [Google Scholar] [CrossRef]
- Kumar, M.; Busch, W.; Birke, H.; Kemmerling, B.; Nürnberger, T.; Schöffl, F. Heat shock factors HsfB1 and HsfB2b are involved in the regulation of Pdf1. 2 expression and pathogen resistance in Arabidopsis. Mol. Plant 2009, 2, 152–165. [Google Scholar] [CrossRef]
- Nishizawa-Yokoi, A.; Nosaka, R.; Hayashi, H.; Tainaka, H.; Maruta, T.; Tamoi, M.; Ikeda, M.; Ohme-Takagi, M.; Yoshimura, K.; Yabuta, Y. HsfA1d and HsfA1e involved in the transcriptional regulation of HsfA2 function as key regulators for the Hsf signaling network in response to environmental stress. Plant Cell Physiol. 2011, 52, 933–945. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Q.; Li, N.; Dong, L.; Zhang, D.; Fan, S.; Jiang, L.; Wang, X.; Xu, P.; Zhang, S. Overexpression of Soybean Isoflavone Reductase (GmIFR) Enhances Resistance to Phytophthora sojae in Soybean. Front. Plant Sci. 2015, 6, 1024. [Google Scholar] [CrossRef] [PubMed]
- Luniwal, A.; Khupse, R.; Reese, M.; Liu, J.; El-Dakdouki, M.; Malik, N.; Fang, L.; Erhardt, P. Multigram synthesis of glyceollin I. Org. Process Res. Dev. 2011, 15, 1149–1162. [Google Scholar] [CrossRef]
- Paddon, C.J.; Westfall, P.J.; Pitera, D.J.; Benjamin, K.; Fisher, K.; McPhee, D.; Leavell, M.; Tai, A.; Main, A.; Eng, D. High-level semi-synthetic production of the potent antimalarial artemisinin. Nature 2013, 496, 528. [Google Scholar] [CrossRef] [PubMed]
- Barrales-Cureño, H.J.; Ramos Valdivia, A.C.; Soto Hernández, M. Increased Production of Taxoids in Suspension Cultures of Taxus globosa after Elicitation. Future Pharmacol. 2022, 2, 45–54. [Google Scholar] [CrossRef]
- Ro, D.-K.; Paradise, E.M.; Ouellet, M.; Fisher, K.J.; Newman, K.L.; Ndungu, J.M.; Ho, K.A.; Eachus, R.A.; Ham, T.S.; Kirby, J. Production of the antimalarial drug precursor artemisinic acid in engineered yeast. Nature. 2006, 440, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Jahan, M.A.; Monsalvo, I.; Ly, M.; Kovinich, N. GmJAZ1 Proteins Bind GmNAC42s and Suppress the Activation of Glyceollin Phytoalexin Biosynthesis in Soybean. Frontiers in Plant Science. 2023. Submitted. [Google Scholar]
- Anguraj Vadivel, A.K.; McDowell, T.; Renaud, J.B.; Dhaubhadel, S. A combinatorial action of GmMYB176 and GmbZIP5 controls isoflavonoid biosynthesis in soybean (Glycine max). Commun. Biol. 2021, 4, 356. [Google Scholar] [CrossRef]
- Bian, S.; Li, R.; Xia, S.; Liu, Y.; Jin, D.; Xie, X.; Dhaubhadel, S.; Zhai, L.; Wang, J.; Li, X. Soybean CCA1-like MYB transcription factor GmMYB133 modulates isoflavonoid biosynthesis. Biochem. Biophys. Res. Commun. 2018, 507, 324–329. [Google Scholar] [CrossRef]
- Wunderlich, M.; Groß-Hardt, R.; Schöffl, F. Heat shock factor HSFB2a involved in gametophyte development of Arabidopsis thaliana and its expression is controlled by a heat-inducible long non-coding antisense RNA. Plant Mol. Biol. 2014, 85, 541–550. [Google Scholar] [CrossRef]
- Kanchiswamy, C.N.; Muroi, A.; Maffei, M.E.; Yoshioka, H.; Sawasaki, T.; Arimura, G.-i. Ca2+-dependent protein kinases and their substrate HsfB2a are differently involved in the heat response signaling pathway in Arabidopsis. Plant Biotechnol. 2010, 27, 469–473. [Google Scholar] [CrossRef]
- Kanchiswamy, C.N.; Takahashi, H.; Quadro, S.; Maffei, M.E.; Bossi, S.; Bertea, C.; Zebelo, S.A.; Muroi, A.; Ishihama, N.; Yoshioka, H. Regulation of Arabidopsis defense responses against Spodoptera littoralisby CPK-mediated calcium signaling. BMC Plant Biol. 2010, 10, 97. [Google Scholar] [CrossRef] [PubMed]
- Farrell, K.; Jahan, M.A.; Kovinich, N. Distinct mechanisms of biotic and chemical elicitors enable additive elicitation of the anticancer phytoalexin glyceollin I. Molecules. 2017, 22, 1261. [Google Scholar] [CrossRef] [PubMed]
- Welle, R.; Grisebach, H. Induction of phytoalexin synthesis in soybean: Enzymatic cyclization of prenylated pterocarpans to glyceollin isomers. Arch. Biochem. Biophys. 1988, 263, 191–198. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).