

MtCLE08, MtCLE16, and MtCLE18 Transcription Patterns and Their Possible Functions in the Embryogenic Calli of Medicago truncatula

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Search for CLE Peptides Related with MtWOX9-1
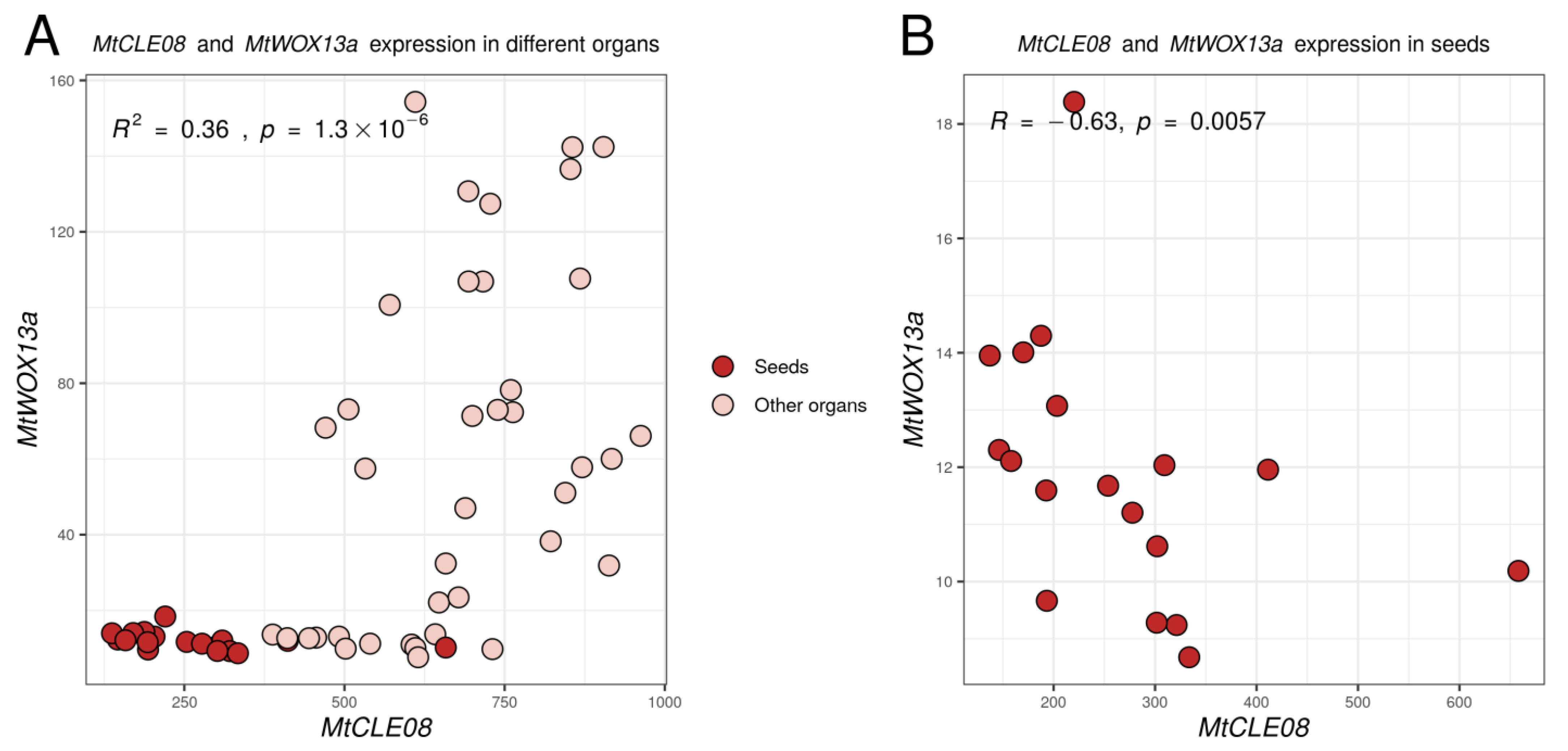
2.2. MtCLE08, 16 and 18 Expression Patterns in Different Organs
2.3. Evaluation of the Expression Dynamics of MtCLE08, 16, and 18 during Somatic Embryogenesis
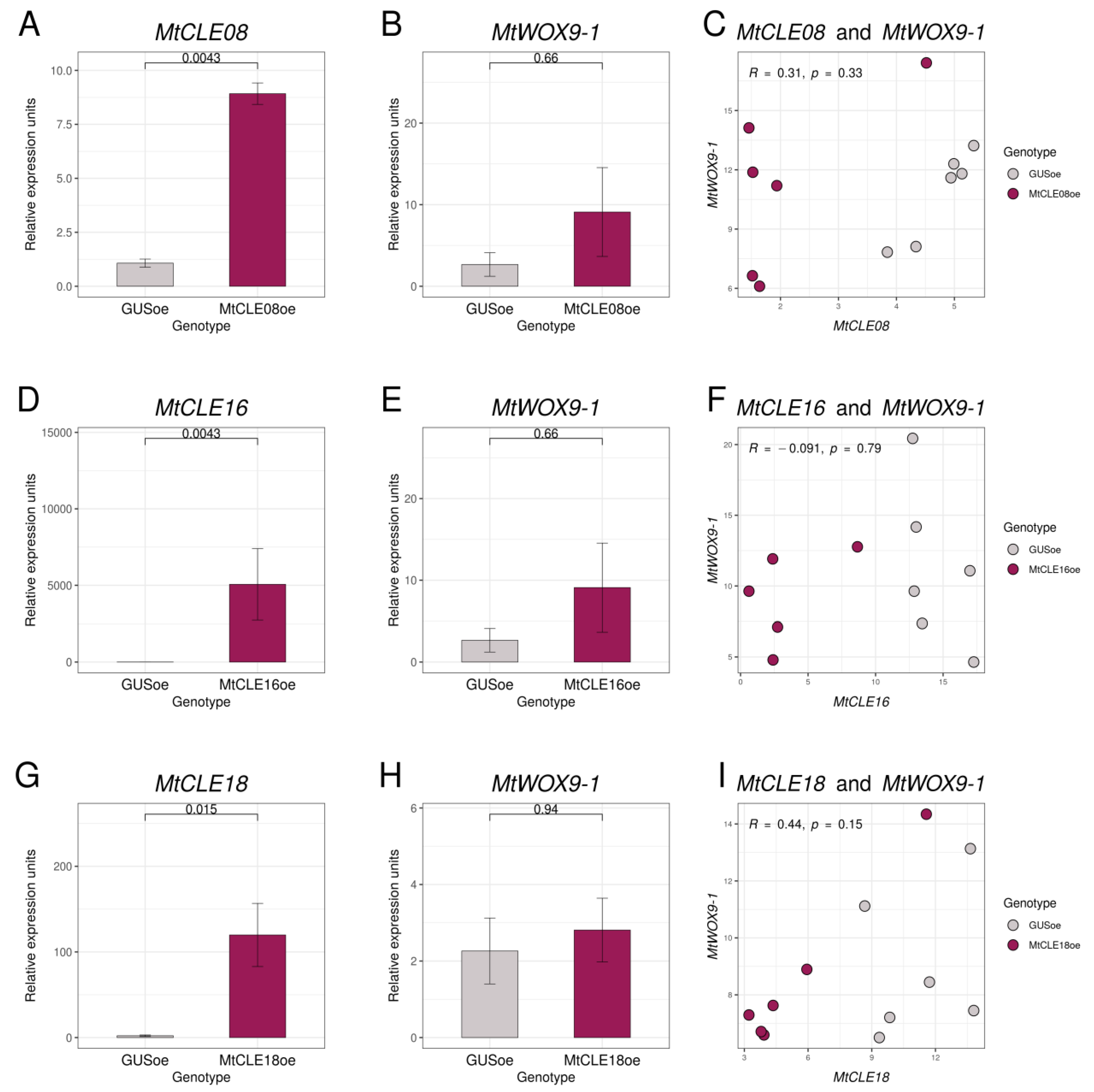
2.4. Evaluation of the Influence of MtCLE08, 16, and 18 Overexpression on MtWOX Expression Levels
2.5. Evaluation of the Influence of MtCLE08, 16, and 18 Overexpression on Somatic Embryogenesis
3. Discussion
4. Materials and Methods
4.1. Plants and Microorganisms
4.2. Molecular Cloning and qPCR Analysis
4.3. Software
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fletcher, J.C. Recent Advances in Arabidopsis CLE Peptide Signaling. Trends Plant Sci. 2020, 25, 1005–1016. [Google Scholar] [CrossRef]
- Willoughby, A.C.; Nimchuk, Z.L. WOX Going on: CLE Peptides in Plant Development. Curr. Opin. Plant Biol. 2021, 63, 102056. [Google Scholar] [CrossRef] [PubMed]
- Hastwell, A.H.; de Bang, T.C.; Gresshoff, P.M.; Ferguson, B.J. CLE Peptide-Encoding Gene Families in Medicago Truncatula and Lotus Japonicus, Compared with Those of Soybean, Common Bean and Arabidopsis. Sci. Rep. 2017, 7, 9384. [Google Scholar] [CrossRef]
- Song, X.-F.; Hou, X.-L.; Liu, C.-M. CLE Peptides: Critical Regulators for Stem Cell Maintenance in Plants. Planta 2021, 255, 5. [Google Scholar] [CrossRef]
- Leibfried, A.; To, J.P.C.; Busch, W.; Stehling, S.; Kehle, A.; Demar, M.; Kieber, J.J.; Lohmann, J.U. WUSCHEL Controls Meristem Function by Direct Regulation of Cytokinin-Inducible Response Regulators. Nature 2005, 438, 1172–1175. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Miotk, A.; Šutiković, Z.; Ermakova, O.; Wenzl, C.; Medzihradszky, A.; Gaillochet, C.; Forner, J.; Utan, G.; Brackmann, K.; et al. WUSCHEL Acts as an Auxin Response Rheostat to Maintain Apical Stem Cells in Arabidopsis. Nat. Commun. 2019, 10, 5093. [Google Scholar] [CrossRef] [PubMed]
- Perales, M.; Rodriguez, K.; Snipes, S.; Yadav, R.K.; Diaz-Mendoza, M.; Reddy, G.V. Threshold-Dependent Transcriptional Discrimination Underlies Stem Cell Homeostasis. Proc. Natl. Acad. Sci. USA 2016, 113, E6298–E6306. [Google Scholar] [CrossRef]
- Brand, U.; Fletcher, J.C.; Hobe, M.; Meyerowitz, E.M.; Simon, R. Dependence of Stem Cell Fate in Arabidopsis on a Feedback Loop Regulated by CLV3 Activity. Science 2000, 289, 617–619. [Google Scholar] [CrossRef] [PubMed]
- Poliushkevich, L.O.; Gancheva, M.S.; Dodueva, I.E.; Lutova, L.A. Receptors of CLE Peptides in Plants. Russ. J. Plant Physiol. 2020, 67, 1–16. [Google Scholar] [CrossRef]
- Berckmans, B.; Kirschner, G.; Gerlitz, N.; Stadler, R.; Simon, R. CLE40 Signaling Regulates Root Stem Cell Fate. Plant Physiology 2020, 182, 1776–1792. [Google Scholar] [CrossRef]
- Hirakawa, Y.; Kondo, Y.; Fukuda, H. TDIF Peptide Signaling Regulates Vascular Stem Cell Proliferation via the WOX4 Homeobox Gene in Arabidopsis. Plant Cell 2010, 22, 2618–2629. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Tucker, E.; Hermann, M.; Laux, T. A Molecular Framework for the Embryonic Initiation of Shoot Meristem Stem Cells. Dev. Cell 2017, 40, 264–277.e4. [Google Scholar] [CrossRef]
- Ren, S.-C.; Song, X.-F.; Chen, W.-Q.; Lu, R.; Lucas, W.J.; Liu, C.-M. CLE25 Peptide Regulates Phloem Initiation in Arabidopsis through a CLERK-CLV2 Receptor Complex. J. Integr. Plant Biol. 2019, 61, 1043–1061. [Google Scholar] [CrossRef] [PubMed]
- Fiume, E.; Fletcher, J.C. Regulation of Arabidopsis Embryo and Endosperm Development by the Polypeptide Signaling Molecule CLE8. Plant Cell 2012, 24, 1000–1012. [Google Scholar] [CrossRef] [PubMed]
- Tvorogova, V.E.; Fedorova, Y.A.; Potsenkovskaya, E.A.; Kudriashov, A.A.; Efremova, E.P.; Kvitkovskaya, V.A.; Wolabu, T.W.; Zhang, F.; Tadege, M.; Lutova, L.A. The WUSCHEL-Related Homeobox Transcription Factor MtWOX9-1 Stimulates Somatic Embryogenesis in Medicago Truncatula. Plant Cell Tissue Organ Cult. 2019, 138, 517–527. [Google Scholar] [CrossRef]
- Tvorogova, V.E.; Lebedeva, M.A.; Lutova, L.A. Expression of WOX and PIN Genes during Somatic and Zygotic Embryogenesis in Medicago Truncatula. Russ. J. Genet. 2015, 51, 1189–1198. [Google Scholar] [CrossRef]
- Tvorogova, V.E.; Krasnoperova, E.Y.; Kudriashov, A.A.; Kuznetsova, K.A.; Potsenkovskaya, E.A.; Fedorova, Y.A.; Lutova, L.A. Transcriptomic Analysis of Medicago Truncatula Calli with MtWOX9-1 Overexpression. Vavilov J. Genet. Breed. 2019, 23, 691–699. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Benedito, V.A.; Torres-Jerez, I.; Murray, J.D.; Andriankaja, A.; Allen, S.; Kakar, K.; Wandrey, M.; Verdier, J.; Zuber, H.; Ott, T.; et al. A Gene Expression Atlas of the Model Legume Medicago Truncatula. Plant J. 2008, 55, 504–513. [Google Scholar] [CrossRef] [PubMed]
- Lowe, K.; La Rota, M.; Hoerster, G.; Hastings, C.; Wang, N.; Chamberlin, M.; Wu, E.; Jones, T.; Gordon-Kamm, W. Rapid Genotype “Independent” Zea mays L. (Maize) Transformation via Direct Somatic Embryogenesis. Vitr. Cell Dev. Biol. Plant 2018, 54, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Kadri, A.; Grenier De March, G.; Guerineau, F.; Cosson, V.; Ratet, P. WUSCHEL Overexpression Promotes Callogenesis and Somatic Embryogenesis in Medicago Truncatula Gaertn. Plants 2021, 10, 715. [Google Scholar] [CrossRef] [PubMed]
- Kyo, M.; Maida, K.; Nishioka, Y.; Matsui, K. Coexpression of WUSCHEL Related Homeobox (WOX) 2 with WOX8 or WOX9 Promotes Regeneration from Leaf Segments and Free Cells in Nicotiana tabacum L. Plant Biotechnol. 2018, 35, 23–30. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Iwase, A.; Rymen, B.; Lambolez, A.; Kojima, M.; Takebayashi, Y.; Heyman, J.; Watanabe, S.; Seo, M.; De Veylder, L.; et al. Wounding Triggers Callus Formation via Dynamic Hormonal and Transcriptional Changes1[OPEN]. Plant Physiol. 2017, 175, 1158–1174. [Google Scholar] [CrossRef]
- Tvorogova, V.E.; Krasnoperova, E.Y.; Potsenkovskaia, E.A.; Kudriashov, A.A.; Dodueva, I.E.; Lutova, L.A. What Does the WOX Say? Review of Regulators, Targets, Partners. Mol. Biol. 2021, 55, 311–337. [Google Scholar] [CrossRef]
- Kang, J.; Wang, X.; Ishida, T.; Grienenberger, E.; Zheng, Q.; Wang, J.; Zhang, Y.; Chen, W.; Chen, M.; Song, X.-F.; et al. A Group of CLE Peptides Regulates de Novo Shoot Regeneration in Arabidopsis Thaliana. New Phytologist 2022, 235, 2300–2312. [Google Scholar] [CrossRef]
- Chen, S.-K.; Kurdyukov, S.; Kereszt, A.; Wang, X.-D.; Gresshoff, P.M.; Rose, R.J. The Association of Homeobox Gene Expression with Stem Cell Formation and Morphogenesis in Cultured Medicago Truncatula. Planta 2009, 230, 827–840. [Google Scholar] [CrossRef]
- Potsenkovskaia, E.; Tvorogova, V.; Yakovleva, D.; Zlydneva, N.; Lutova, L. Novel NF-Y Genes Expressed during Somatic Embryogenesis in Medicago Truncatula. Plant Gene 2022, 31, 100364. [Google Scholar] [CrossRef]
- Curtis, M.D.; Grossniklaus, U. A Gateway Cloning Vector Set for High-Throughput Functional Analysis of Genes in Planta. Plant Physiol. 2003, 133, 462–469. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Okonechnikov, K.; Golosova, O.; Fursov, M. UGENE team Unipro UGENE: A Unified Bioinformatics Toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [PubMed]
- Pecrix, Y.; Staton, S.E.; Sallet, E.; Lelandais-Brière, C.; Moreau, S.; Carrère, S.; Blein, T.; Jardinaud, M.-F.; Latrasse, D.; Zouine, M.; et al. Whole-Genome Landscape of Medicago Truncatula Symbiotic Genes. Nat. Plants 2018, 4, 1017. [Google Scholar] [CrossRef]
- Young, N.D.; Debellé, F.; Oldroyd, G.E.D.; Geurts, R.; Cannon, S.B.; Udvardi, M.K.; Benedito, V.A.; Mayer, K.F.X.; Gouzy, J.; Schoof, H.; et al. The Medicago Genome Provides Insight into the Evolution of Rhizobial Symbioses. Nature 2011, 480, 520–524. [Google Scholar] [CrossRef]
- Tang, H.; Krishnakumar, V.; Bidwell, S.; Rosen, B.; Chan, A.; Zhou, S.; Gentzbittel, L.; Childs, K.L.; Yandell, M.; Gundlach, H.; et al. An Improved Genome Release (Version Mt4.0) for the Model Legume Medicago Truncatula. BMC Genom. 2014, 15, 312. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kudriashov, A.A.; Zlydneva, N.S.; Efremova, E.P.; Tvorogova, V.E.; Lutova, L.A. MtCLE08, MtCLE16, and MtCLE18 Transcription Patterns and Their Possible Functions in the Embryogenic Calli of Medicago truncatula. Plants 2023, 12, 435. https://doi.org/10.3390/plants12030435
Kudriashov AA, Zlydneva NS, Efremova EP, Tvorogova VE, Lutova LA. MtCLE08, MtCLE16, and MtCLE18 Transcription Patterns and Their Possible Functions in the Embryogenic Calli of Medicago truncatula. Plants. 2023; 12(3):435. https://doi.org/10.3390/plants12030435
Chicago/Turabian StyleKudriashov, Andrei A., Natalia S. Zlydneva, Elena P. Efremova, Varvara E. Tvorogova, and Ludmila A. Lutova. 2023. "MtCLE08, MtCLE16, and MtCLE18 Transcription Patterns and Their Possible Functions in the Embryogenic Calli of Medicago truncatula" Plants 12, no. 3: 435. https://doi.org/10.3390/plants12030435
APA StyleKudriashov, A. A., Zlydneva, N. S., Efremova, E. P., Tvorogova, V. E., & Lutova, L. A. (2023). MtCLE08, MtCLE16, and MtCLE18 Transcription Patterns and Their Possible Functions in the Embryogenic Calli of Medicago truncatula. Plants, 12(3), 435. https://doi.org/10.3390/plants12030435