

Yield and Composition of the Essential Oil of Clinopodium nepeta subsp. spruneri as Affected by Harvest Season and Cultivation Method, i.e., Outdoor, Greenhouse and In Vitro Culture



Abstract
:1. Introduction
2. Results and Discussion
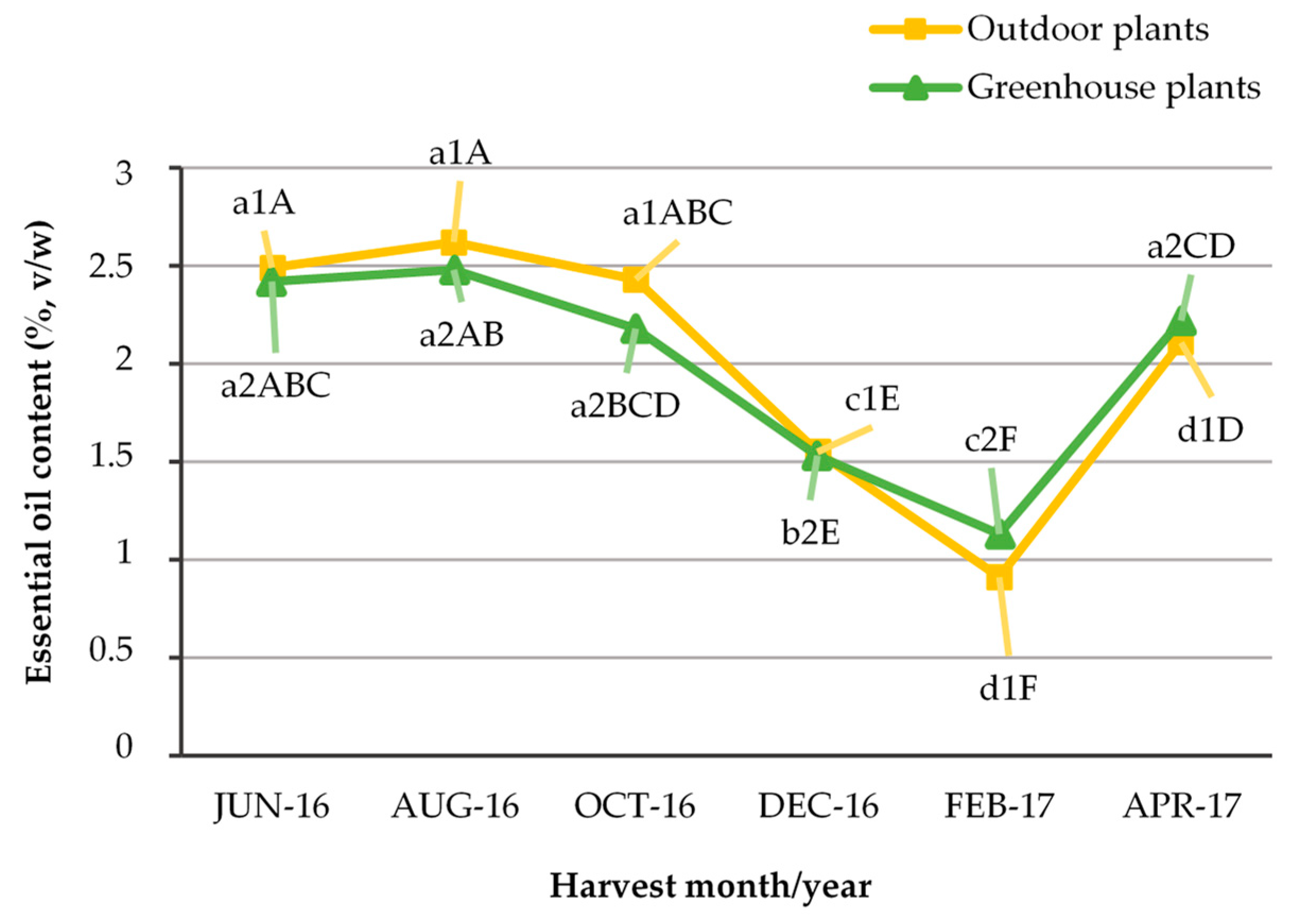
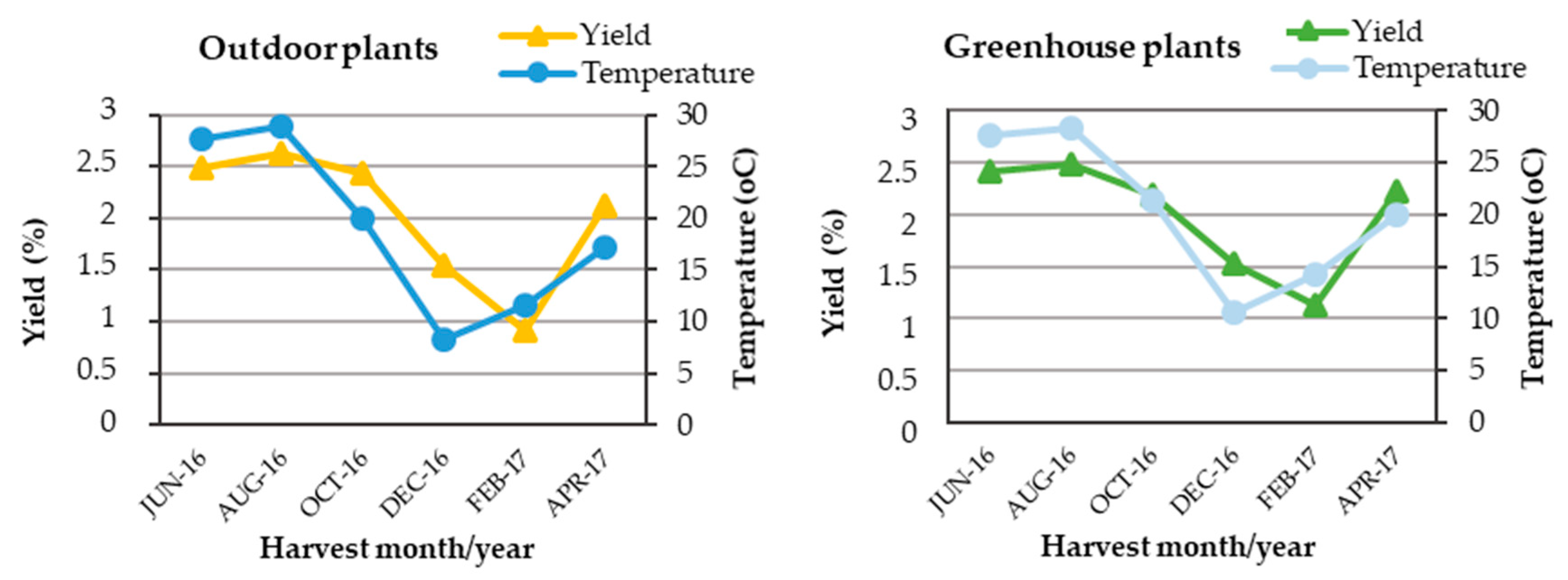
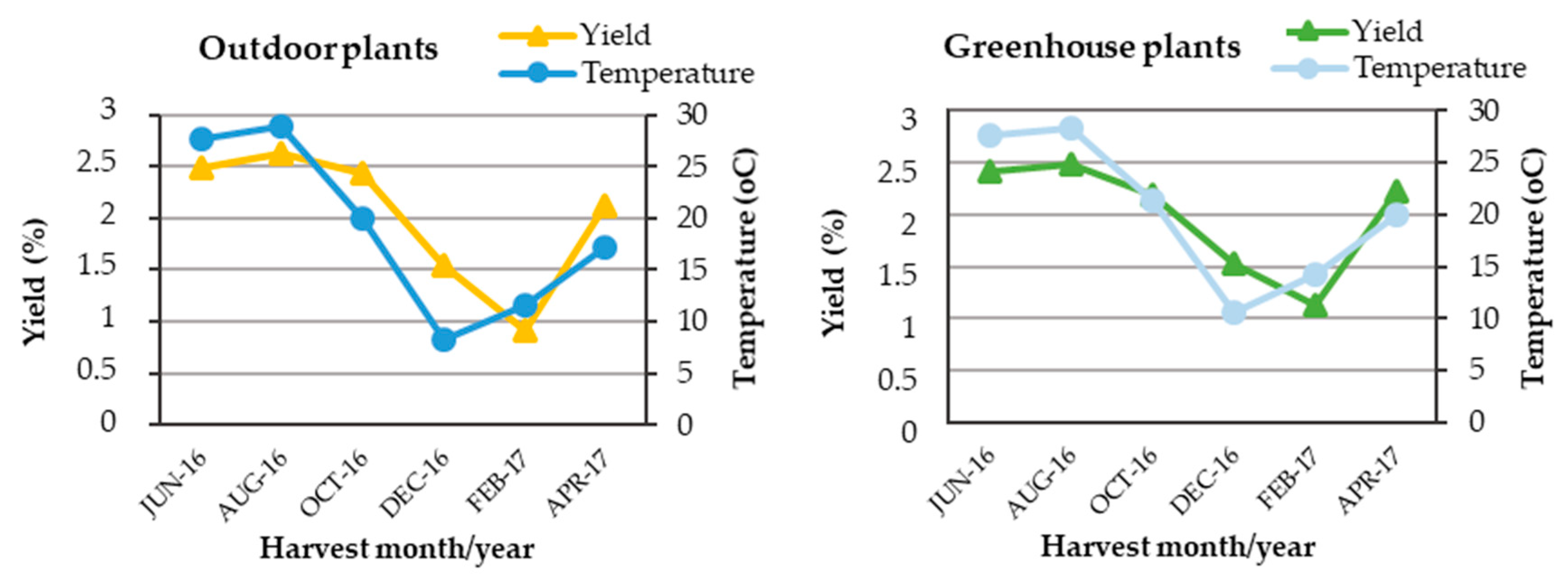
2.1. Average Percentage Content and Seasonal Variation of Essential Oil Extracted from Outdoor, Greenhouse and In Vitro Plants
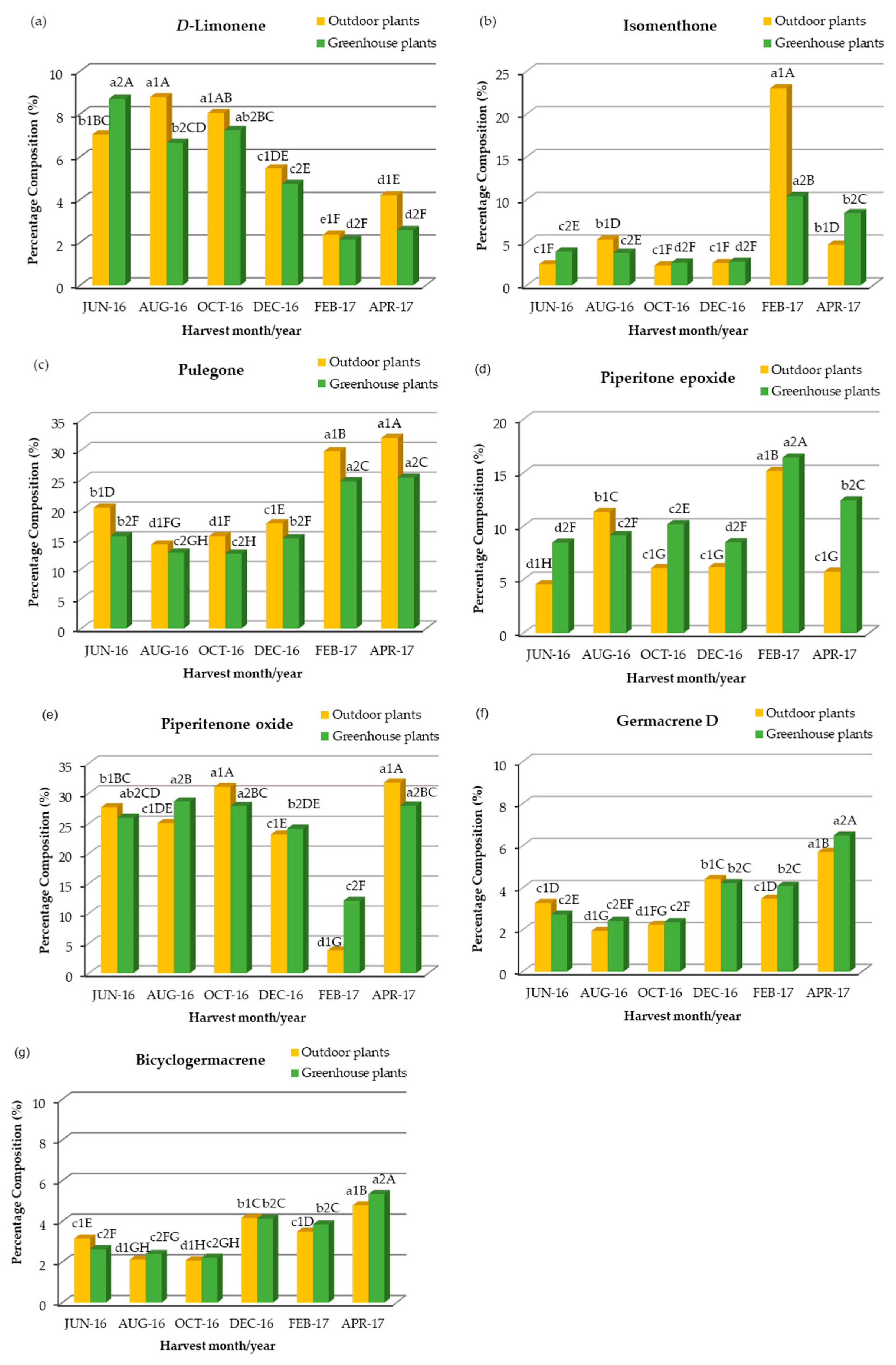
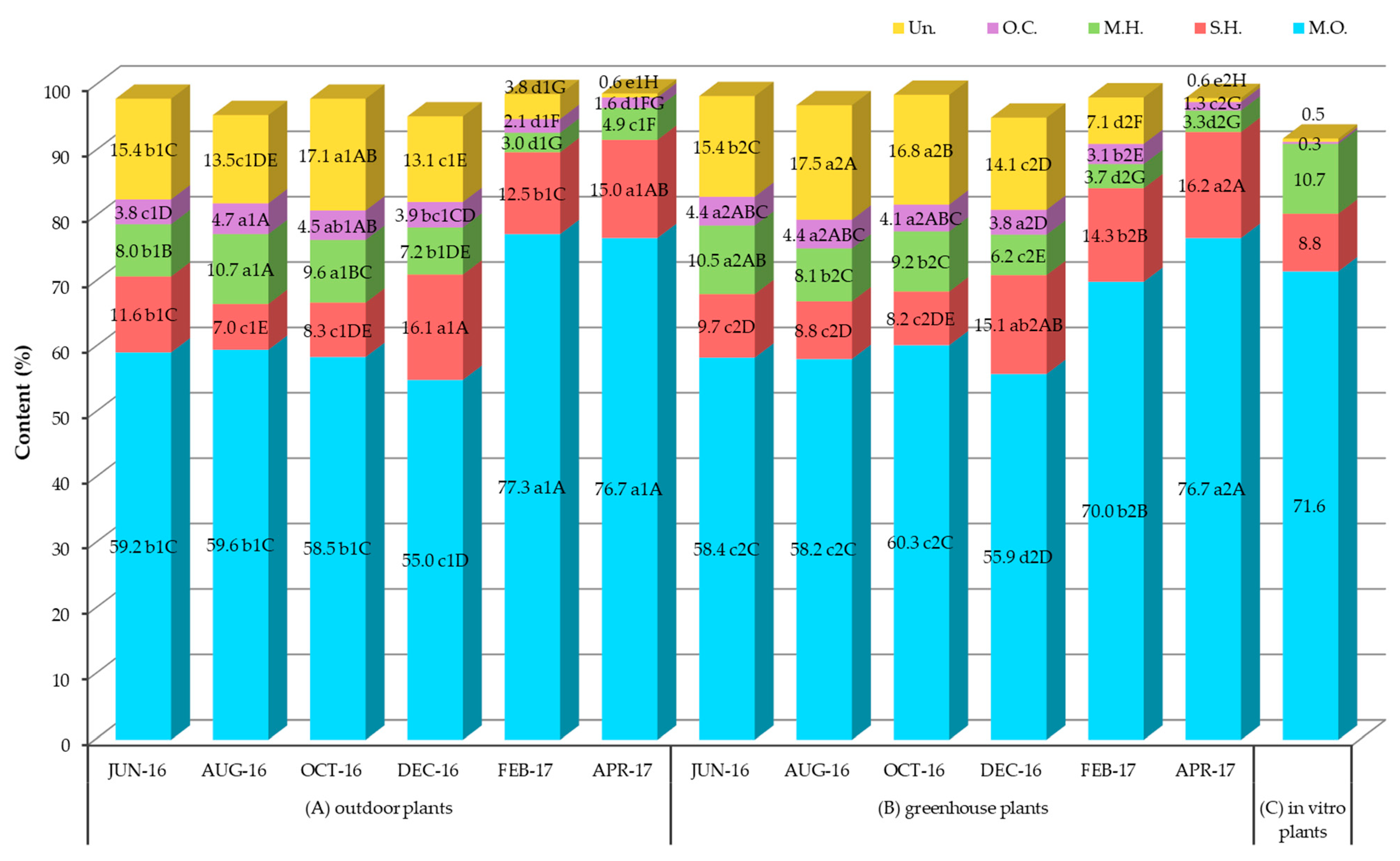
2.2. Chemical Composition, Seasonal Variation and Comparative Study of the Main Chemical Groups of Essential Oil Extracted from Outdoor, Greenhouse and In Vitro Plants
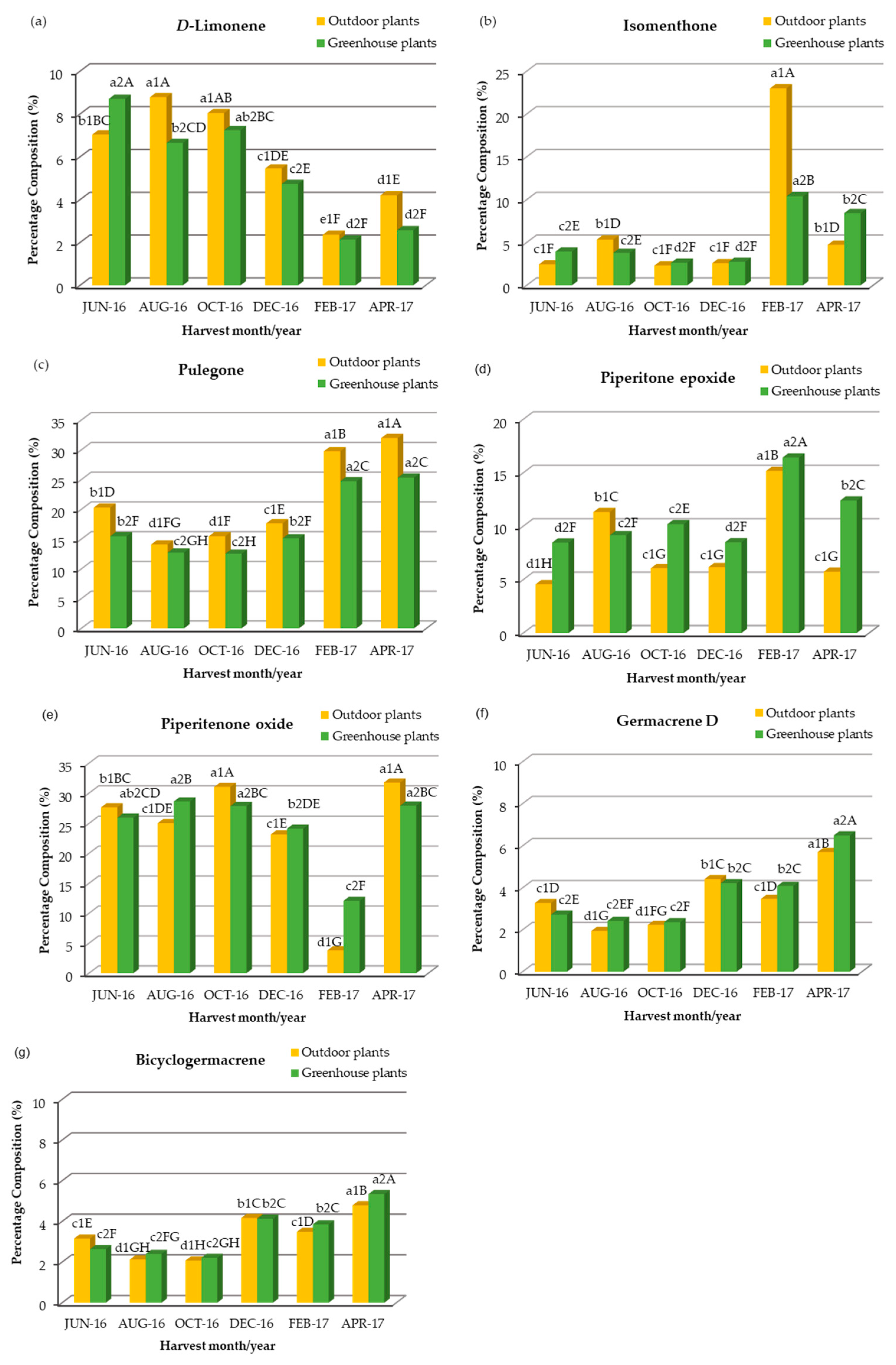
2.3. Chemical Composition, Seasonal Variation and Comparative Study of the Concentration and Seasonal Variation of the Main Compounds of Essential Oil Extracted from Outdoor, Greenhouse and In Vitro Plants
3. Materials and Methods
3.1. Plant Material
3.1.1. Outdoor and Greenhouse Plants (In Vivo Plants)
3.1.2. In Vitro Plants
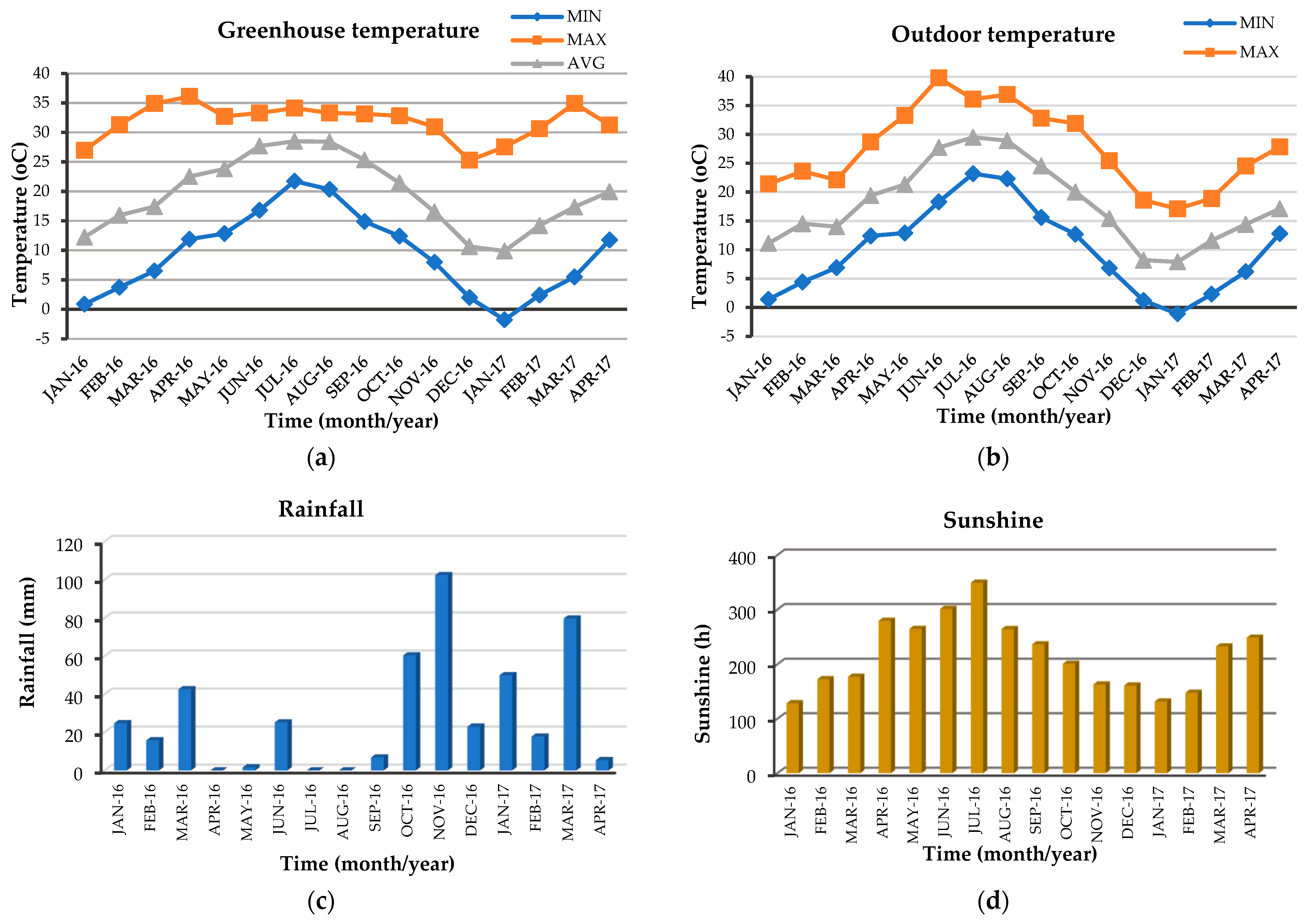
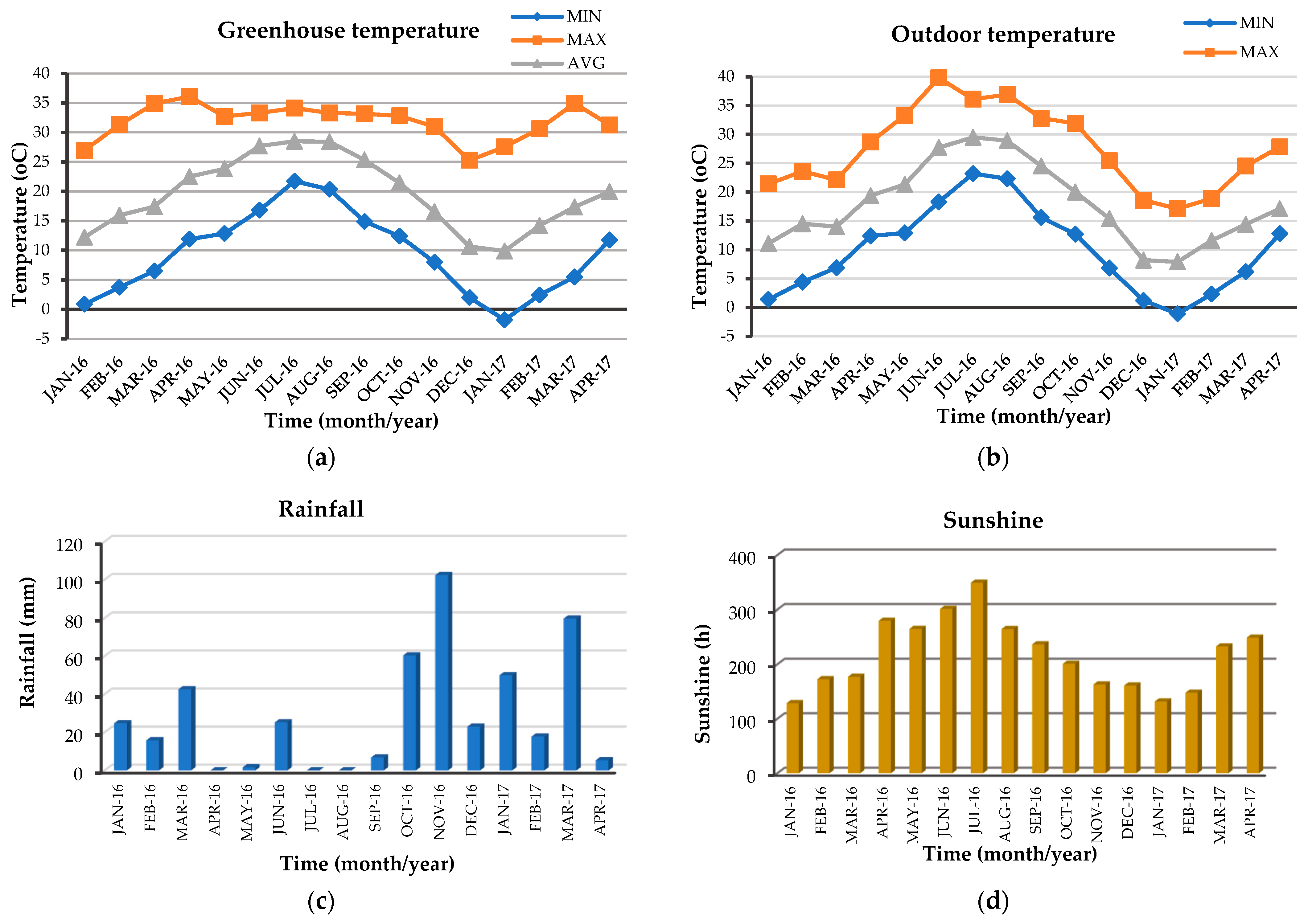
3.2. Meteorological Data
3.3. Harvesting, Drying and Preparation of Plant Samples
3.3.1. Outdoor and Greenhouse Plant Material
3.3.2. In Vitro Plant Material
3.4. Essential Oil Isolation
3.5. Gas Chromatography-Mass Spectrometry (GC/MS) Analysis
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Adams, M.; Berset, C.; Kessler, M.; Hamburger, M. Medicinal herbs for the treatment of rheumatic disorders—A survey of European herbals from the 16th and 17th century. J. Ethnopharmacol. 2009, 121, 343–359. [Google Scholar] [CrossRef] [PubMed]
- Nacer-bey, N.; Chabane, D.; Haouli, A.K.; Aribi, I. Traditional herbal medicine in Jijel region, Northeast of Algeria. Adv. Environ. Biol. 2015, 9, 54–61. [Google Scholar]
- Rashrash, M.; Schommer, J.C.; Brown, L.M. Prevalence and Predictors of Herbal Medicine Use Among Adults in the United States. J. Patient Exp. 2017, 4, 108–113. [Google Scholar] [CrossRef]
- Salim, M.A.; Ranjitkar, S.; Hart, R.; Khan, T.; Ali, S.; Kiran, C.; Parveen, A.; Batool, Z.; Bano, S.; Xu, J. Regional trade of medicinal plants has facilitated the retention of traditional knowledge: Case study in Gilgit-Baltistan Pakistan. J. Ethnobiol. Ethnomed. 2019, 15, 6. [Google Scholar] [CrossRef]
- Zhang, L.; Zhuang, H.; Zhang, Y.; Wang, L.; Zhang, Y.; Geng, Y.; Yi, G.; Pei, S.; Wang, Y. Plants for health: An ethnobotanical 25-year repeat survey of traditional medicine sold in a major marketplace in North-west Yunnan, China. J. Ethnopharmacol. 2018, 224, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Alotiby, A.A.; Al-Harbi, L.N. Prevalence of using herbs and natural products as a protective measure during the COVID-19 pandemic among the Saudi population: An online cross-sectional survey. Saudi Pharm. J. 2021, 29, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Ball, P.W.; Getliffe, F. Calamintha Miller. In Flora Europaea; Tutin, T., Heywood, V., Burges, N., Moore, D., Valentine, D., Walters, S., Webb, D., Eds.; Cambridge University Press: Cambridge, UK, 1968; Volume 3, pp. 166–167. [Google Scholar]
- Davis, P.H.; Leblebici, E. Flora of Turkey and East Aegean Islands; University Press: Edinburgh, UK, 1987; Volume 7, pp. 323–329. [Google Scholar]
- Davis, P.H.; Mill, R.R.; Tan, K. Flora of Turkey and East Aegean Islands; University Press: Edinburgh, UK, 1988; Volume 10, p. 207. [Google Scholar]
- Alan, S.; Ocak, A. Taxonomical and morphological studies on the genus Calamintha Miller (Lamiaceae) in Turkey. Biol. Divers. Conserv. 2009, 2, 125–143. [Google Scholar]
- Bown, D. The Herb Society of America Encyclopedia of Herbs and Their Uses; Dorling Kindersley: New York, NY, USA, 1995; Volume 97, p. 252. [Google Scholar]
- Chevallier, A. Encyclopedia of Medicinal Plants; Dorling Kindersley: London, UK, 2001; pp. 211–212. [Google Scholar]
- Brankovic, S.V.; Kitic, D.V.; Radenkovic, M.M.; Veljkovic, S.M.; Golubovic, T.D. Calcium blocking activity as a mechanism of the spasmolytic effect of the essential oil of Calamintha glandulosa Silic on the isolated rat ileum. Gen. Physiol. Biophys. 2009, 28, 174–178. [Google Scholar]
- Flamini, G.; Cioni, P.L.; Puleio, R.; Morelli, I.; Panizzi, L. Antimicrobial activity of the essential oil of Calamintha nepeta and its constituent pulegone against bacteria and fungi. Phyther. Res. 1999, 13, 349–351. [Google Scholar] [CrossRef]
- Monforte, M.T.; Tzakou, O.; Nostro, A.; Zimbalatti, V.; Galati, E.M. Chemical composition and biological activities of Calamintha officinalis Moench essential oil. J. Med. Food 2011, 14, 297–303. [Google Scholar] [CrossRef]
- Pardo-de-Santayana, M.; Tardío, J.; Blanco, E.; Carvalho, A.; Lastra, J.; San Miguel, E.; Morales, R. Traditional knowledge of wild edible plants used in the northwest of the Iberian Peninsula (Spain and Portugal): A comparative study. J. Ethnobiol. Ethnomed. 2007, 3, 27. [Google Scholar] [CrossRef] [PubMed]
- Sarac, N.; Ugur, A. The in vitro antimicrobial activities of the essential oils of some Lamiaceae species from Turkey. J. Med. Food 2009, 12, 902–907. [Google Scholar] [CrossRef] [PubMed]
- Verma, D.; Irchhaiya, M.; Singh, R.; Kailasiya, P.P.; Kanaujia, V. Studies on antiulcer activity of essential oil of Calamintha officinalis Moench. Int. J. Res. Pharm. Sci. 2011, 2, 2733–2736. [Google Scholar]
- Amira, S.; Dade, M.; Schinella, G.; Ríos, J.L. Anti-inflammatory, anti-oxidant, and apoptotic activities of four plant species used in folk medicine in the Mediterranean basin. Pak. J. Pharm. Sci. 2012, 25, 65–72. [Google Scholar] [PubMed]
- Ceker, S.; Agar, G.; Alpsoy, L.; Nardemir, G.; Kizil, H.E. Protective role of essential oils of Calamintha nepeta L. on oxidative and genotoxic damage caused by Alfatoxin B1 in vitro. Fresenius Environ. Bull. 2013, 22, 3258–3263. [Google Scholar]
- Sarikurkcu, C.; Ozer, M.S.; Tepe, B.; Dilek, E.; Ceylan, O. Phenolic composition, antioxidant and enzyme inhibitory activities of acetone, methanol and water extracts of Clinopodium vulgare L. subsp. vulgare L. Ind. Crops Prod. 2015, 76, 961–966. [Google Scholar] [CrossRef]
- Shams Moattar, F.; Sariri, R.; Giahi, M.; Yaghmaee, P. Essential oil composition and antioxidant activity of Calamintha officinalis Moench. J. Appl. Biotechnol. Rep. 2018, 5, 55–58. [Google Scholar] [CrossRef]
- Beddiar, H.; Boudiba, S.; Benahmed, M.; Tamfu, A.N.; Ceylan, Ö.; Hanini, K.; Küçükaydın, S.; Elomari, A.; Chawki, B.; Laouer, H.; et al. Chemical Composition, Anti-Quorum Sensing, Enzyme Inhibitory, and Antioxidant Properties of Phenolic Extracts of Clinopodium nepeta L. Kuntze. Plants 2021, 10, 1955. [Google Scholar] [CrossRef]
- Petrova, M.; Dimitrova, L.; Dimitrova, M.; Denev, P.; Teneva, D.; Georgieva, A.; Petkova-Kirova, P.; Lazarova, M.; Tasheva, K. Antitumor and Antioxidant Activities of In Vitro Cultivated and Wild-Growing Clinopodium vulgare L. Plants. Plants 2023, 12, 1591. [Google Scholar] [CrossRef]
- Rodenak-Kladniew, B.; Castro, M.A.; Gambaro, R.C.; Girotti, J.; Cisneros, J.S.; Viña, S.; Padula, G.; Crespo, R.; Castro, G.R.; Gehring, S.; et al. Cytotoxic Screening and Enhanced Anticancer Activity of Lippia alba and Clinopodium nepeta Essential Oils-Loaded Biocompatible Lipid Nanoparticles against Lung and Colon Cancer Cells. Pharmaceutics 2023, 15, 2045. [Google Scholar] [CrossRef]
- Pignatti, S. Flora D’Italia; Edagricole: Bologna, Italy, 1982; Volume 2, pp. 345–347. [Google Scholar]
- Filibeck, G.; Cornelini, P.; Petrella, P. Floristic analysis of a high-speed railway embankment in a Mediterranean landscape. Acta Bot. Croat. 2012, 71, 229–248. [Google Scholar] [CrossRef]
- Dimopoulos, P.; Raus, T.; Bergmeier, E.; Constantinidis, T.; Iatrou, G.; Kokkini, S.; Strid, A.; Tzanoudakis, D. Vascular Plants of Greece: An Annotated Checklist; Englera 31; Botanic Garden and Botanical Museum Berlin-Dahlem and Hellenic Botanical Society: Berlin, Germany, 2013; Volume 106, p. 174. [Google Scholar]
- Brussell, D. Medicinal Plants of Mt. Pelion, Greece. Econ. Bot. 2004, 58, S174–S202. [Google Scholar] [CrossRef]
- Baytop, T. Therapy with Medicinal Plants in Turkey Past and Present, 2nd ed.; Nobel Tip Basimevi: Istanbul, Turkey, 1999; p. 371. [Google Scholar]
- Marongiu, B.; Piras, A.; Porcedda, S.; Falconieri, D.; Maxia, A.; Gonçalves, M.J.; Cavaleiro, C.; Salgueiro, L. Chemical composition and biological assays of essential oils of Calamintha nepeta (L.) Savi subsp. nepeta (Lamiaceae). Nat. Prod. Res. 2010, 24, 1734–1742. [Google Scholar] [CrossRef]
- Demirci, B.; Temel, H.E.; Portakal, T.; Kırmızıbekmez, H.; Demirci, F.; Baser, K.H.C. Inhibitory effect of Calamintha nepeta subsp. glandulosa essential oil on lipoxygenase. Turk. J. Biochem. 2011, 36, 290–295. [Google Scholar]
- Božović, M.; Ragno, R. Calamintha nepeta (L.) Savi and its Main Essential Oil Constituent Pulegone: Biological Activities and Chemistry—A review. Molecules 2017, 22, 290. [Google Scholar] [CrossRef] [PubMed]
- Debbabi, H.; Mokni, R.E.; Chaieb, I.; Nardoni, S.; Maggi, F.; Caprioli, G.; Hammami, S. Chemical Composition, Antifungal and Insecticidal Activities of the Essential Oils from Tunisian Clinopodium nepeta subsp. nepeta and Clinopodium nepeta subsp. glandulosum. Molecules 2020, 25, 2137–2149. [Google Scholar] [CrossRef]
- Lee, S.; Tsao, R.; Peterson, C.; Coats, J.R. Insecticidal activity of monoterpenoids to western corn rootworm (Coleoptera: Chrysomelidae), twospotted spider mite (Acari: Tetranychidae), and house fly (Diptera: Muscidae). J. Econ. Entomol. 1997, 90, 883–892. [Google Scholar] [CrossRef]
- Souleles, C.; Argyriadou, N.; Philianos, S. Constituents of the essential oil of Calamintha nepeta. J. Nat. Prod. 1987, 50, 510–511. [Google Scholar] [CrossRef]
- Kokkalou, E.; Stefanou, E. The volatile oil of Calamintha nepeta (L.) Savi ssp. glandulosa (Req.) Ball, endemic to Greece. Flavour Fragr. J. 1990, 5, 23–26. [Google Scholar] [CrossRef]
- Baldovini, N.; Ristorcelli, D.; Tomi, F.; Casanova, J. Intraspecific variability of the essential oil of Calamintha nepeta from Corsica (France). Flavour Fragr. J. 2000, 15, 50–54. [Google Scholar] [CrossRef]
- Couladis, M.; Tzakou, O. Essential oil of Calamintha nepeta subsp. glandulosa from Greece. J. Essent. Oil Res. 2001, 13, 11–12. [Google Scholar] [CrossRef]
- Kitic, D.; Stojanović, G.; Palic, R.; Randjelovic, V. Chemical composition and microbial activity of the essential oil of Calamintha nepeta (L.) Savi ssp. nepeta var. subisodonda (Borb.) Hayek from Serbia. J. Essent. Oil Res. 2005, 17, 701–703. [Google Scholar] [CrossRef]
- Cook, C.M.; Lanaras, T.; Kokkini, S. Essential oils of two Calamintha glandulosa (Req.) Bentham chemotypes in a wild population from Zakynthos, Greece. J. Essent. Oil Res. 2007, 19, 534–539. [Google Scholar] [CrossRef]
- Karousou, R.; Hanlidou, E.; Lazari, D. Essential-oil diversity of three Calamintha species from Greece. Chem. Biodivers. 2012, 9, 1364–1372. [Google Scholar] [CrossRef] [PubMed]
- Baser, K.H.C.; Kirimer, N.; Tümen, G. Pulegone-rich essential oils of Turkey. J. Essent. Oil Res. 1998, 10, 1–8. [Google Scholar] [CrossRef]
- Dhingra, A.K.; Chopra, B.; Bhardwaj, S.; Dhar, K.L. Synthesis and characterization of novel pulegone derivatives as substitutes of 4-(1,1 dimethylethyl) cyclohexan-1-ol acetate. J. Pharm. Res. 2011, 4, 19–21. [Google Scholar]
- Yang, L.; Kui-Shan Wen, K.-S.; Ruan, X.; Zhao, Y.-X.; Wei, F.; Wang, Q. Response of Plant Secondary Metabolites to Environmental Factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [PubMed]
- Jan, R.; Asaf, S.; Numan, M.; Lubna; Kim, K.-M. Plant Secondary Metabolite Biosynthesis and Transcriptional Regulation in Response to Biotic and Abiotic Stress Conditions. Agronomy 2021, 11, 968. [Google Scholar] [CrossRef]
- Pant, P.; Pandey, S.; Dall’Acqua, S. The Influence of Environmental Conditions on Secondary Metabolites in Medicinal Plants: A Literature Review. Chem. Biodivers. 2021, 18, e2100345. [Google Scholar] [CrossRef]
- Giuffrè, A.M.; Nobile, R. Citrus bergamia, Risso: The peel, the juice and the seed oil of the bergamot fruit of Reggio Calabria (South Italy). Emir. J. Food Agric. 2020, 32, 522–532. [Google Scholar] [CrossRef]
- Şanli, A.; Karadoğan, T. Geographical impact on essential oil composition of endemic Kundmannia anatolica Hub.-Mor. (Apiaceae). Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Gioffrè, G.; Ursino, D.; Labate, M.L.C.; Giuffrè, A.M. The peel essential oil composition of bergamot fruit (Citrus bergamia, Risso) of Reggio Calabria (Italy): A review. Emir. J. Food Agric. 2020, 32, 835–845. [Google Scholar] [CrossRef]
- de Araujo, J.; Silvestre, W.P.; Pauletti, G.F.; Muniz, L.A.R. Influence of the Absolute Pressure of the Extraction System on the Yield and Composition of Corymbia citriodora (Hook.) K.D. Hill and L.A.S. Johnson Leaf Essential Oil Extracted by Steam Distillation. ChemEngineering 2023, 7, 67. [Google Scholar] [CrossRef]
- Perry, N.B.; Anderson, R.E.; Brennan, N.J.; Douglas, M.H.; Heaney, A.J.; McGimpsey, J.A.; Smallfield, B.M. Essential oils from dalmatian sage (Salvia officinalis L.): Variations among individuals, plant parts, seasons, and sites. J. Agric. Food Chem. 1999, 47, 2048–2054. [Google Scholar] [CrossRef]
- Yavari, A.; Nazeri, V.; Sefidkon, F.; Hassani, M.E. Influence of some environmental factors on the essential oil variability of Thymus migricus. Nat. Prod. Commun. 2010, 5, 943–948. [Google Scholar] [CrossRef]
- Fernández-Sestelo, M.; Carrillo, J.M. Environmental effects on yield and composition of essential oil in wild populations of spike lavender (Lavandula latifolia Medik.). Agriculture 2020, 10, 626. [Google Scholar] [CrossRef]
- Slavkovska, V.; Zlatković, B.; Bräuchler, C.; Stojanović, D.; Tzakou, O.; Couladis, M. Variations of essential oil characteristics of Clinopodium pulegium (Lamiaceae) depending on phenological stage. Bot. Serbica 2013, 37, 97–104. [Google Scholar]
- Matkowski, A. Plant in vitro culture for the production of antioxidants—A review. Biotechnol. Adv. 2008, 26, 548–560. [Google Scholar] [CrossRef]
- Silva, S.; Sato, A.; Lage, C.L.S.; San Gil, R.A.S.; de Azevedo, D.; Esquibel, M.A. Essential oil composition of Mellisa officinalis L. in vitro produced under the influence of growth regulators. J. Braz. Chem. Soc. 2005, 16, 1387–1390. [Google Scholar] [CrossRef]
- Affonso, V.R.; Bizzo, H.R.; Lima, S.S.; Esquibel, M.A.; Sato, A. Solid phase microextraction (SPME) analysis of volatile compounds produced by in vitro shoots of Lantana camara L. under the influence of auxins and cytokinins. J. Braz. Chem. Soc. 2007, 18, 1504–1508. [Google Scholar] [CrossRef]
- Pistelli, L.; Noccioli, C.; D’Angiolillo, F.; Pistelli, L. Composition of volatile in micropropagated and field grown aromatic plants from Tuscany Islands. Acta Biochim. Pol. 2013, 60, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, A.M.; Goleniowski, M.E.; Aimar, M.L.; Diaz, M.S.; Demmel, G.I.; Decarlini, M.F.; Cantero, J.J. Profile Characterization of Volatile Organic Compounds on in vitro Propagated Plants of Clinopodium odorum and it’s Comparison with the Wild Plant. SAJ Pharm. Pharmacol. 2018, 5, 103. [Google Scholar]
- Lila, M.A. Valuable Secondary Products from In Vitro Culture. In Plant Development and Biotechnology; Trigiano, R.N., Gray, D., Eds.; CRC Press: Boca Raton, FL, USA, 2005; pp. 285–289. [Google Scholar]
- Gounaris, Y. Biotechnology for the production of essential oils, flavours and volatile isolates. A review. Flavour Fragr. J. 2010, 25, 367–386. [Google Scholar] [CrossRef]
- Collin, H.A. Secondary product formation in plant tissue cultures. Plant Growth Regul. 2001, 34, 119–134. [Google Scholar] [CrossRef]
- Amaral, C.L.F.; Silva, A.B. Melhoramento biotecnológico de plantas medicinais. Biotecnol. Ciência Desenvolv. 2003, 30, 55–59. [Google Scholar]
- Wawrosch, C.; Zotchev, S.B. Production of bioactive plant secondary metabolites through in vitro technologies—Status and outlook. Appl. Microbiol. Biotechnol. 2021, 105, 6649–6668. [Google Scholar] [CrossRef]
- Affonso, V.R.; Bizzo, H.R.; Lage, C.L.S.; Sato, A. Influence of growth regulators in biomass production and volatile profile of in vitro plantlets of Thymus vulgaris (L.). J. Agric. Food Chem. 2009, 57, 6392–6395. [Google Scholar] [CrossRef]
- Vlachou, G.; Papafotiou, M.; Bertsouklis, K.F. In vitro propagation of Calamintha nepeta. Acta Hortic. 2016, 1113, 189–194. [Google Scholar] [CrossRef]
- Vlachou, G.; Papafotiou, M.; Bertsouklis, K. Studies on seed germination and micropropagation of Clinopodium nepeta: A medicinal and aromatic plant. Hort. Sci. 2019, 54, 1558–1564. [Google Scholar] [CrossRef]
- Şarer, E.; Solakel Pançalı, S. Composition of the essential oil from Calamintha nepeta (L.) Savi ssp. glandulosa (Req.) PW Ball. Flavour Fragr. J. 1998, 13, 31–32. [Google Scholar] [CrossRef]
- Bellomaria, B.; Valentini, G. Composizione dell’olio essenziale di Calamintha nepeta subsp. glandulosa. Plant Biosyst. 1985, 119, 237–245. [Google Scholar] [CrossRef]
- De Pooter, H.L.; de Buyck, L.F.; Schamp, N.M. The volatiles of Calamintha nepeta subsp. glandusola. Phytochemistry 1986, 25, 691–694. [Google Scholar] [CrossRef]
- Akgül, A.; De Pooter, H.L.; De Buyck, L.F. The essential oils of Calamintha nepeta subsp. glandulosa and Ziziphora clinopodioides from Turkey. J. Essent. Oil Res. 1991, 3, 7–10. [Google Scholar] [CrossRef]
- Panizzi, L.; Flamini, G.; Cioni, P.L.; Morelli, I. Composition and antimicrobial properties of essential oils of four Mediterranean Lamiaceae. J. Ethnopharmacol. 1993, 39, 167–170. [Google Scholar] [CrossRef]
- Mastelić, J.; Miloš, M.; Kuštrak, D.; Radonić, A. The essential oil and glycosidically bound volatile compounds of Calamintha nepeta (L.) Savi. Croat. Chem. Acta 1998, 71, 147–154. [Google Scholar]
- Kitic, D.; Jovanovic, T.; Ristic, M.; Palic, R.; Stojanovic, G. Chemical composition and antimicrobial activity of the essential oil of Calamintha nepeta (L.) Savi ssp. glandulosa (Req.) PW Ball from Montenegro. J. Essent. Oil Res. 2002, 14, 150–152. [Google Scholar] [CrossRef]
- Hodaj-Çeliku, E.; Tsiftsoglou, O.; Shuka, L.; Abazi, S.; Hadjipavlou-Litina, D.; Lazari, D. Antioxidant activity and chemical composition of essential oils of some aromatic and medicinal plants from Albania. Nat. Prod. Commun. 2017, 12, 785–790. [Google Scholar] [CrossRef] [PubMed]
- Arantes, S.M.; Piçarra, A.; Guerreiro, M.; Salvador, C.; Candeias, F.; Caldeira, A.T.; Martins, M.R. Toxicological and pharmacological properties of essential oils of Calamintha nepeta, Origanum virens and Thymus mastichina of Alentejo (Portugal). Food Chem. Toxicol. 2019, 133, 110747. [Google Scholar] [CrossRef]
- Negro, C.; Notarnicola, S.; De Bellis, L.; Miceli, A. Intraspecific variability of the essential oil of Calamintha nepeta subsp. nepeta from Southern Italy (Apulia). Nat. Prod. Res. 2013, 27, 331–339. [Google Scholar] [CrossRef]
- Kofidis, G.; Bosabalidis, A.; Kokkini, S. Seasonal variation of essential oils in a linalool-rich chemotype of Mentha spicata grown wild in Greece. J. Essent. Oil Res. 2004, 16, 469–472. [Google Scholar] [CrossRef]
- Hussain, A.I.; Anwar, F.; Sherazi, S.T.H.; Przybylski, R. Chemical composition, antioxidant and antimicrobial activities of basil (Ocimum basilicum) essential oils depends on seasonal variations. Food Chem. 2008, 108, 986–995. [Google Scholar] [CrossRef] [PubMed]
- Khalid, A.K.; El-Gohary, A.E. Effect of seasonal variations on essential oil production and composition of Plectranthus amboinicus (Lour.) grow in Egypt. Int. Food Res. J. 2014, 21, 1859–1862. [Google Scholar]
- Lakušić, B.; Ristić, M.; Slavkovska, V.; Lakušić, D. Seasonal variations in the composition of the essential oils of rosemary (Rosmarinus officinalis, Lamiaceae). Nat. Prod. Commun. 2013, 8, 131–134. [Google Scholar] [CrossRef] [PubMed]
- Melito, S.; Petretto, G.L.; Chahine, S.; Pintore, G.; Chessa, M. Seasonal variation of essential oil in Rosmarinus officinalis leaves in Sardinia. Nat. Prod. Commun. 2019, 14, 1–7. [Google Scholar] [CrossRef]
- Guedes, A.P.; Amorim, L.R.; Vicente, A.; Fernandes-Ferreira, M. Variation of the essential oil content and composition in leaves from cultivated plants of Hypericum androsaemum L. Phytochem. Anal. Int. J. Plant Chem. Biochem. Tech. 2004, 15, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Lakušić, B.; Ristić, M.; Slavkovska, V.; Stojanović, D.; Lakušić, D. Variations in essential oil yields and compositions of Salvia officinalis (Lamiaceae) at different developmental stages. Bot. Serbica 2013, 37, 127–139. [Google Scholar]
- Chericoni, S.; Flamini, G.; Campeol, E.; Cioni, P.L.; Morelli, I. GC–MS analyses of the essential oil from the aerial parts of Artemisia verlotiorum: Variability during the year. Biochem. Syst. Ecol. 2004, 32, 423–429. [Google Scholar] [CrossRef]
- Palá-Paúl, J.; Pérez-Alonso, M.J.; Velasco-Negueruela, A.; Palá-Paúl, R.; Sanz, J.; Conejero, F. Seasonal variation in chemical constituents of Santolina rosmarinifolia L. ssp. rosmarinifolia. Biochem. Syst. Ecol. 2001, 29, 663–672. [Google Scholar] [CrossRef]
- Karousou, R.; Kokkini, S.; Bessière, J.M.; Vokou, D. Calamintha cretica (Lamiaceae), a Cretan endemic: Distribution and essential oil composition. Nord. J. Bot. 1996, 16, 247–252. [Google Scholar] [CrossRef]
- Juliani, H.R., Jr.; Koroch, A.R.; Juliani, H.R.; Trippi, V.S.; Zygadlo, J.A. Intraespecific variation in leaf oils of Lippia junelliana (mold.) tronc. Biochem. Syst. Ecol. 2002, 30, 163–170. [Google Scholar] [CrossRef]
- Aziz, E.E.; Badawy, E.M.; Zheljazkov, V.D.; Nicola, S.M.; Fouad, H. Yield and chemical composition of essential oil of Achillea millefolium L. as affected by harvest time. Egypt. J. Chem. 2019, 62, 533–540. [Google Scholar] [CrossRef]
- Gazim, Z.C.; Amorim, A.C.L.; Hovell, A.M.C.; Rezende, C.M.; Nascimento, I.A.; Ferreira, G.A.; Cortez, D.A.G. Seasonal variation, chemical composition, and analgesic and antimicrobial activities of the essential oil from leaves of Tetradenia riparia (Hochst.) Codd in Southern Brazil. Molecules 2010, 15, 5509–5524. [Google Scholar] [CrossRef] [PubMed]
- Hirata, T.; Murakami, S.; Ogihara, K.; Suga, T. Volatile monoterpenoid constituents of the plantlets of Mentha spicata produced by shoot tip culture. Phytochemistry 1990, 29, 493–495. [Google Scholar] [CrossRef]
- Arikat, N.A.; Jawad, F.M.; Karam, N.S.; Shibli, R.A. Micropropagation and accumulation of essential oils in wild sage (Salvia fruticosa Mill.). Sci. Hortic. 2004, 100, 193–202. [Google Scholar] [CrossRef]
- Amaral-Baroli, A.; Lago, J.H.G.; de Almeida, C.V.; de Almeida, M.; Scotti, M.T.; Leone, G.F.; Soares, M.G.; Cavalari, A.A.; Sartorelli, P. Variability in essential oil composition produced by micropropagated (in vitro), acclimated (ex vitro) and in-field plants of Ocimum basilicum (Lamiaceae). Ind. Crops Prod. 2016, 86, 180–185. [Google Scholar] [CrossRef]
- Thiem, B.; Kikowska, M.; Kurowska, A.; Kalemba, D. Essential oil composition of the different parts and in vitro shoot culture of Eryngium planum L. Molecules 2011, 16, 7115–7124. [Google Scholar] [CrossRef]
- Nogueira, J.M.F.; Romano, A. Essential oils from micropropagated plants of Lavandula viridis. Phytochem. Anal. Int. J. Plant Chem. Biochem. Tech. 2002, 13, 4–7. [Google Scholar] [CrossRef] [PubMed]
- Manan, A.A.; Taha, R.M.; Mubarak, E.E.; Elias, H. In vitro flowering, glandular trichomes ultrastructure, and essential oil accumulation in micropropagated Ocimum basilicum L. Vitr. Cell. Dev. Biol.-Plant. 2016, 52, 303–314. [Google Scholar] [CrossRef]
- Shen, X.; Kane, M.E.; Chen, J. Effects of genotype, explant source, and plant growth regulators on indirect shoot organogenesis in Dieffenbachia cultivars. Vitr. Cell. Dev. Biol.-Plant 2008, 44, 282–288. [Google Scholar] [CrossRef]
- Amoo, S.; Adeyemi, A.; Staden, J. Shoot proliferation and rooting treatments influence secondary metabolite production and antioxidant activity in tissue culture-derived Aloe arborescens grown ex vitro. Plant Growth Regul. 2013, 70, 115–122. [Google Scholar] [CrossRef]
- Ćavar, S.; Vidic, D.; Maksimović, M. Volatile constituents, phenolic compounds, and antioxidant activity of Calamintha glandulosa (Req.) Bentham. J. Sci. Food Agric. 2012, 93, 1758–1764. [Google Scholar] [CrossRef] [PubMed]
- Öztürk, G.; Yilmaz, G.; Gülnur, E.K.Ş.İ.; Demitri, B. Chemical composition and antibacterial activity of Clinopodium nepeta subsp. glandulosum (Req.) Govaerts essential oil. Nat. Vol. Essent. Oils 2021, 8, 75–80. [Google Scholar] [CrossRef]
- Alan, S.; Kürkçüoglu, M.; Hüsnü Can Baser, K. Composition of Essential Oils of Calamintha nepeta (L.) Savi subsp. nepeta and Calamintha nepeta (L.) Savi subsp. glandulosa (Req.) P.W. Ball. Asian J. Chem. 2011, 23, 2357–2360. [Google Scholar]
- De Pooter, H.L.; Goetghebeur, P.; Schamp, N. Variability in composition of the essential oil of Calamintha nepeta. Phytochemistry 1987, 26, 3355–3356. [Google Scholar] [CrossRef]
- Ristorcelli, D.; Tomi, F.; Casanova, J. Essential oils of Calamintha nepeta subsp. nepeta and subsp. glandulosa from Corsica (France). J. Essent. Oil Res. 1996, 8, 363–366. [Google Scholar] [CrossRef]
- Kizil, S.; Ipek, A.; Arslan, N.; Khawar, K.M. Effect of different developing stages on some agronomical characteristics and essential oil composition of oregano (Oreganum onites). N. Zeal. J. Crop Hortic. Sci. 2008, 36, 71–76. [Google Scholar] [CrossRef]
- Lukas, B.; Schmiderer, C.; Novak, J. Essential oil diversity of European Origanum vulgare L. (Lamiaceae). Phytochemistry 2015, 119, 32–40. [Google Scholar] [CrossRef]
- Yurteri, E.; Seyis, F.; Küplemez, H. Investigation of the essential oil components of Calamintha nepeta (L.) savi. subsp. glandulosa PLANT. In Proceedings of the International Baku Scientific Reserch Congress, Baku, Azerbaijan, 15–16 October 2021; pp. 192–194. [Google Scholar]
- Hussain, A.I.; Anwar, F.; Nigam, P.S.; Ashraf, M.; Gilani, A.H. Seasonal variation in content, chemical composition and antimicrobial and cytotoxic activities of essential oils from four Mentha species. J. Sci. Food Agric. 2010, 90, 1827–1836. [Google Scholar] [CrossRef]
- Pacifico, S.; Galasso, S.; Piccolella, S.; Kretschmer, N.; Pan, S.P.; Marciano, S.; Bauer, R.; Monaco, P. Seasonal variation in phenolic composition and antioxidant and anti-inflammatory activities of Calamintha nepeta (L.). Savi. Food Res. Int. 2015, 69, 121–132. [Google Scholar] [CrossRef]
- Villa-Ruano, N.; Pacheco-Hernández, Y.; Cruz-Durán, R.; Lozoya-Gloria, E. Volatiles and seasonal variation of the essential oil composition from the leaves of Clinopodium macrostemum var. laevigatum and its biological activities. Ind. Crops Prod. 2015, 77, 741–747. [Google Scholar] [CrossRef]
- Yesil Celiktas, O.; Hames Kocabas, E.E.; Bedir, E.; Vardar Sukan, F.; Ozek, T.; Baser, K.H.C. Antimicrobial activities of methanol extracts and essential oils of Rosmarinus officinalis, depending on location and seasonal variations. Food Chem. 2007, 100, 553–559. [Google Scholar] [CrossRef]
- Atti-Santos, A.C.; Pansera, M.R.; Paroul, N.; Atti-Serafini, L.; Moyna, P. Seasonal variation of essential oil yield and composition of Thymus vulgaris L. (Lamiaceae) from South Brazil. J. Essent. Oil Res. 2004, 16, 294–295. [Google Scholar] [CrossRef]
- McGimpsey, J.A.; Douglas, M.H.; Van Klink, J.W.; Beauregard, D.A.; Perry, N.B. Seasonal variation in essential oil yield and composition from naturalized Thymus vulgaris L. in New Zealand. Flavour Fragr. J. 1994, 9, 347–352. [Google Scholar] [CrossRef]
- Chebel, A.V.; Koroch, A.R.; Juliani, J.R.; Juliani, H.R.; Trippi, V.S. Micropropagation of Minthostachys mollis (H.B.K.) Grieseb. and essential oil composition of clonally propagated plants. Vitr. Cell. Dev. Biol.-Plant 1998, 34, 249. [Google Scholar]
- Fortunato, M.I.; Avato, P. Plant development and synthesis of essential oils in micropropagated and mycorrhiza inoculated plants of Origanum vulgare L. ssp. hirtum (Link) Ietswaart. Plant Cell Tiss. Org. Cult. 2008, 93, 139–149. [Google Scholar] [CrossRef]
- Kuźma, Ł.; Kalemba, D.; Różalski, M.; Różalska, B.; Więckowska-Szakiel, M.; Krajewska, U.; Wysokińska, H. Chemical composition and biological activities of essential oil from Salvia sclarea plants regenerated in vitro. Molecules 2009, 14, 1438–1447. [Google Scholar] [CrossRef]
- Skała, E.; Kalemba, D.; Wajs, A.; Różalski, M.; Krajewska, U.; Różalska, B.; Więckowska-Szakiel, M.; Wysokińska, H. In vitro propagation and chemical and biological studies of the essential oil of Salvia przewalskii Maxim. Z. Für Naturforschung 2007, 62, 839–848. [Google Scholar] [CrossRef]
- Argyropoulou, C.; Daferera, D.; Tarantilis, P.A.; Fasseas, C.; Polissiou, M. Chemical composition of the essential oil from leaves of Lippia citriodora H.B.K. at two developmental stages. Biochem. Syst. Ecol. 2007, 35, 831–837. [Google Scholar] [CrossRef]
- Prins, C.L.; Vieira, I.J.C.; Freitas, S.P. Growth regulators and essential oil production. Braz. J. Plant Physiol. 2010, 22, 91–102. [Google Scholar] [CrossRef]
- Ioannidis, D.; Bonner, L.; Johnson, C.B. UV-B is Required for Normal Development of Oil Glands in Ocimum basilicum L. (Sweet Basil). Ann. Bot. 2002, 90, 453–460. [Google Scholar] [CrossRef]
- Schreiner, M.; Mewis, I.; Huyskens-Keil, S.; Jansen, M.A.K.; Zrenner, R.; Winkler, J.B.; O’Brien, N.; Krumbein, A. UV-B-Induced Secondary Plant Metabolites—Potential Benefits for Plant and Human Health. Crit. Rev. Plant Sci. 2012, 31, 229–240. [Google Scholar] [CrossRef]
- Pandey, A.; Agrawal, M.; Agrawal, S.B. Ultraviolet-B and Heavy Metal-Induced Regulation of Secondary Metabolites in Medicinal Plants: A Review. Metabolites 2023, 13, 341. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- Lucero, M.; Estell, R.; Tellez, M.; Fredricksona, E.D. Retention Index Calculator Simplifies Identification of Plant Volatile Organic Compounds. Phytochem. Anal. 2009, 20, 378–384. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| June | August | October | December | February | April | June | August | October | December | February | April | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| R.I.exp | Compounds | Outdoor Plants | Greenhouse Plants | In Vitro Plants | |||||||||||
| 1. | 938 | α-Pinene | 0.2 | 0.6 | 0.6 | 0.6 | 0.2 | 0.2 | 0.6 | 0.4 | 0.7 | 0.5 | 0.5 | 0.2 | 0.2 |
| 2. | 981 | β-Pinene | 0.4 | 0.6 | 0.6 | 0.6 | 0.3 | 0.4 | 0.7 | 0.5 | 0.7 | 0.5 | 0.8 | 0.4 | 0.2 |
| 3. | 990 | β-Myrcene | 0.3 | 0.5 | 0.3 | 0.4 | 0.1 | 0.1 | 0.4 | 0.4 | 0.5 | 0.4 | 0.3 | 0.1 | 0.3 |
| 4. | 994 | 3-Octanol | 2.5 | 2.9 | 3.0 | 2.7 | 1.0 | 1.2 | 2.9 | 2.6 | 2.6 | 2.5 | 1.8 | 1.0 | 0.2 |
| 5. | 1032 | D-Limonene | 7.1 | 8.8 | 8.0 | 5.5 | 2.4 | 4.2 | 8.7 | 6.7 | 7.2 | 4.7 | 2.1 | 2.6 | 10.0 |
| 6. | 1062 | γ-Terpinene | nd | 0.2 | 0.1 | 0.1 | nd | nd | 0.1 | 0.1 | 0.1 | 0.1 | nd | nd | - |
| 7. | 1095 | 3-Nonanol | 0.2 | 0.6 | 0.3 | 0.2 | nd | nd | 0.3 | 0.4 | 0.3 | 0.2 | 0.3 | 0.2 | - |
| 8. | 1099 | trans-Sabinene hydrate | 0.1 | nd | 0.1 | 0.1 | nd | nd | 0.1 | nd | nd | nd | nd | nd | - |
| 9. | 1102 | n-Nonanal | 0.1 | 0.4 | 0.1 | 0.1 | 1.0 | 0.3 | 0.2 | 0.2 | 0.1 | 0.1 | nd | nd | 0.1 |
| 10. | 1156 | Menthone | - | - | - | - | - | - | - | - | - | - | - | - | 0.2 |
| 11. | 1165 | Isomenthone | 2.5 | 5.3 | 2.3 | 2.6 | 23.0 | 4.7 | 4.0 | 3.8 | 2.6 | 2.7 | 10.4 | 8.4 | 4.6 |
| 12. | 1178 | Isopulegone * | 0.6 | 0.3 | 0.3 | 1.1 | 0.9 | 0.4 | 0.3 | 0.2 | nd | 1.1 | 1.1 | 0.4 | 0.1 |
| 13. | 1182 | Unknown (M1) ** | 0.5 | 0.8 | 0.4 | 0.3 | 0.7 | nd | 0.4 | 0.6 | 0.8 | 0.2 | nd | nd | - |
| 14. | 1205 | Unknown (M2) ** | 0.3 | 0.9 | 0.5 | 0.5 | 1.3 | nd | 0.7 | 0.8 | 0.9 | 0.7 | 0.7 | nd | - |
| 15. | 1225 | 2-Ethenyl-1-methoxy-3-methylbenzene | 1.0 | 0.8 | 1.1 | 0.9 | 0.1 | 0.1 | 1.0 | 1.2 | 1.1 | 1.0 | 1.0 | 0.1 | - |
| 16. | 1240 | Pulegone | 20.3 | 14.1 | 15.5 | 17.6 | 29.8 | 32.0 | 15.5 | 12.7 | 12.5 | 15.1 | 24.7 | 25.3 | 15.0 |
| 17. | 1256 | Piperitone epoxide * | 4.6 | 11.3 | 6.1 | 6.2 | 15.2 | 5.7 | 8.5 | 9.2 | 10.2 | 8.5 | 16.4 | 12.4 | 15.5 |
| 18. | 1275 | Isopiperitenone | 0.9 | 0.9 | 1.0 | 0.6 | 0.5 | 0.5 | 1.0 | 1.0 | 0.9 | 0.8 | 0.6 | 0.5 | 0.3 |
| 19. | 1293 | Unknown (M3) ** | 8.8 | 7.2 | 9.9 | 7.1 | 1.3 | 0.2 | 8.6 | 9.7 | 9.1 | 7.7 | 4.0 | 0.2 | 0.2 |
| 20. | 1298 | Thymol | 0.6 | 1.0 | 0.7 | 0.8 | 1.0 | 0.3 | 0.8 | 0.9 | 1.0 | 0.9 | 1.2 | 0.3 | 0.2 |
| 21. | 1301 | 2-Hydroxypiperitone | 0.3 | 0.9 | 0.4 | 0.5 | 1.5 | nd | 0.8 | 0.9 | 1.0 | 0.8 | 1.8 | nd | |
| 22. | 1309 | Carvacrol | 0.5 | 0.1 | 0.1 | 0.4 | 0.3 | nd | 0.3 | 0.1 | 2.1 | 0.6 | 0.5 | 0.1 | 0.5 |
| 23. | 1343 | Piperitenone | 1.2 | 0.7 | 1.0 | 1.9 | 1.1 | 1.3 | 1.2 | 0.8 | 2.1 | 1.3 | 1.2 | 1.4 | 1.0 |
| 24. | 1366 | Piperitenone oxide | 27.6 | 25.0 | 31.0 | 23.2 | 3.8 | 31.8 | 25.9 | 28.6 | 27.9 | 24.1 | 12.1 | 27.9 | 34.2 |
| 25. | 1383 | α-Copaene | 0.2 | 0.2 | 0.3 | 0.5 | 0.1 | 0.3 | 0.3 | 0.3 | 0.3 | 0.4 | 0.2 | 0.2 | - |
| 26. | 1392 | β-Bourbonene | 0.7 | 0.3 | 0.8 | 0.3 | 0.2 | 0.2 | 0.6 | 0.8 | 0.7 | 0.8 | 0.2 | nd | - |
| 27. | 1397 | β-Elemene | 0.3 | 0.2 | 0.2 | 0.4 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.4 | 0.3 | 0.2 | 0.1 |
| 28. | 1407 | Unknown (M4) ** | 5.8 | 4.6 | 6.3 | 5.2 | 0.5 | 0.4 | 5.7 | 6.4 | 6.0 | 5.5 | 2.4 | 0.4 | 0.3 |
| 29. | 1428 | β-Caryuphyllene | 2.6 | 1.6 | 1.8 | 3.9 | 3.8 | 3.5 | 2.0 | 1.7 | 1.5 | 3.2 | 3.7 | 3.6 | 0.5 |
| 30. | 1436 | β-Copaene | 0.2 | 0.1 | 0.2 | 0.4 | 0.2 | nd | 0.2 | 0.1 | 0.1 | 0.3 | 0.3 | nd | - |
| 31. | 1447 | Aromadendrene | 0.2 | 0.1 | 0.1 | 0.3 | 0.1 | nd | 0.1 | 0.2 | 0.2 | 0.4 | 0.3 | nd | - |
| 32. | 1461 | α-Caryoplyllene | 0.1 | 0.1 | 0.2 | 0.4 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.3 | 0.3 | 0.2 | - |
| 33. | 1470 | cis-Muurola-4(14)5-diene | 0.2 | 0.1 | 0.1 | 0.3 | 0.2 | 0.1 | 0.2 | 0.1 | 0.1 | 0.2 | 0.2 | 0.1 | 0.1 |
| 34. | 1483 | γ-Muurolene | 0.2 | 0.1 | 0.1 | 0.3 | 0.2 | nd | 0.2 | 0.1 | 0.1 | 0.2 | 0.2 | nd | - |
| 35. | 1489 | Germacrene D | 3.3 | 1.9 | 2.2 | 4.4 | 3.5 | 5.7 | 2.7 | 2.4 | 2.4 | 4.2 | 4.1 | 6.5 | 2.9 |
| 36. | 1504 | Bicyclogermacrene | 3.2 | 2.1 | 2.1 | 4.2 | 3.5 | 4.8 | 2.6 | 2.4 | 2.2 | 4.1 | 3.9 | 5.3 | 5.2 |
| 37. | 1520 | γ-Cadinene | 0.2 | 0.1 | 0.1 | 0.3 | 0.1 | nd | 0.1 | 0.1 | 0.1 | 0.2 | 0.2 | nd | - |
| 38. | 1528 | δ-Cadinene | 0.2 | 0.1 | 0.1 | 0.3 | 0.2 | nd | 0.2 | 0.2 | 0.1 | 0.3 | 0.3 | 0.1 | - |
| 39. | 1544 | α-Cadinene | tr | nd | nd | 0.1 | nd | nd | 0.1 | tr | nd | 0.1 | 0.1 | nd | - |
| 40. | 1591 | Spathulenol | 0.1 | tr | 0.2 | 0.1 | nd | nd | 0.1 | 0.1 | nd | 0.1 | nd | nd | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vlachou, G.; Papafotiou, M.; Daferera, D.J.; Tarantilis, P.A. Yield and Composition of the Essential Oil of Clinopodium nepeta subsp. spruneri as Affected by Harvest Season and Cultivation Method, i.e., Outdoor, Greenhouse and In Vitro Culture. Plants 2023, 12, 4098. https://doi.org/10.3390/plants12244098
Vlachou G, Papafotiou M, Daferera DJ, Tarantilis PA. Yield and Composition of the Essential Oil of Clinopodium nepeta subsp. spruneri as Affected by Harvest Season and Cultivation Method, i.e., Outdoor, Greenhouse and In Vitro Culture. Plants. 2023; 12(24):4098. https://doi.org/10.3390/plants12244098
Chicago/Turabian StyleVlachou, Georgia, Maria Papafotiou, Dimitra J. Daferera, and Petros A. Tarantilis. 2023. "Yield and Composition of the Essential Oil of Clinopodium nepeta subsp. spruneri as Affected by Harvest Season and Cultivation Method, i.e., Outdoor, Greenhouse and In Vitro Culture" Plants 12, no. 24: 4098. https://doi.org/10.3390/plants12244098
APA StyleVlachou, G., Papafotiou, M., Daferera, D. J., & Tarantilis, P. A. (2023). Yield and Composition of the Essential Oil of Clinopodium nepeta subsp. spruneri as Affected by Harvest Season and Cultivation Method, i.e., Outdoor, Greenhouse and In Vitro Culture. Plants, 12(24), 4098. https://doi.org/10.3390/plants12244098