
Declining Radial Growth in Major Western Carpathian Tree Species: Insights from Three Decades of Temperate Forest Monitoring

Abstract
:1. Introduction
2. Materials and Methods
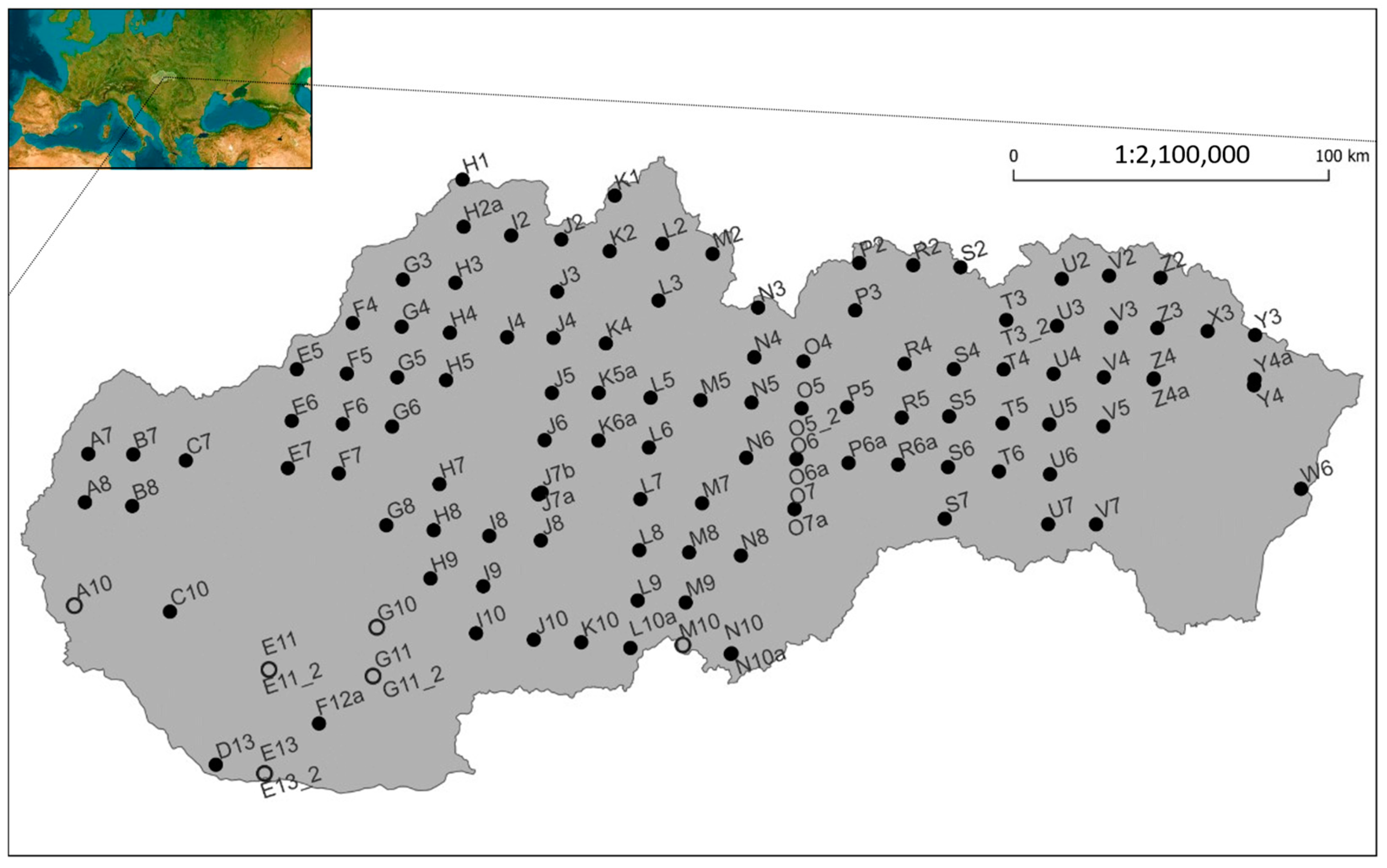
2.1. ICP Forests Level I Tree Growth Database
2.2. Detrending and Representativeness
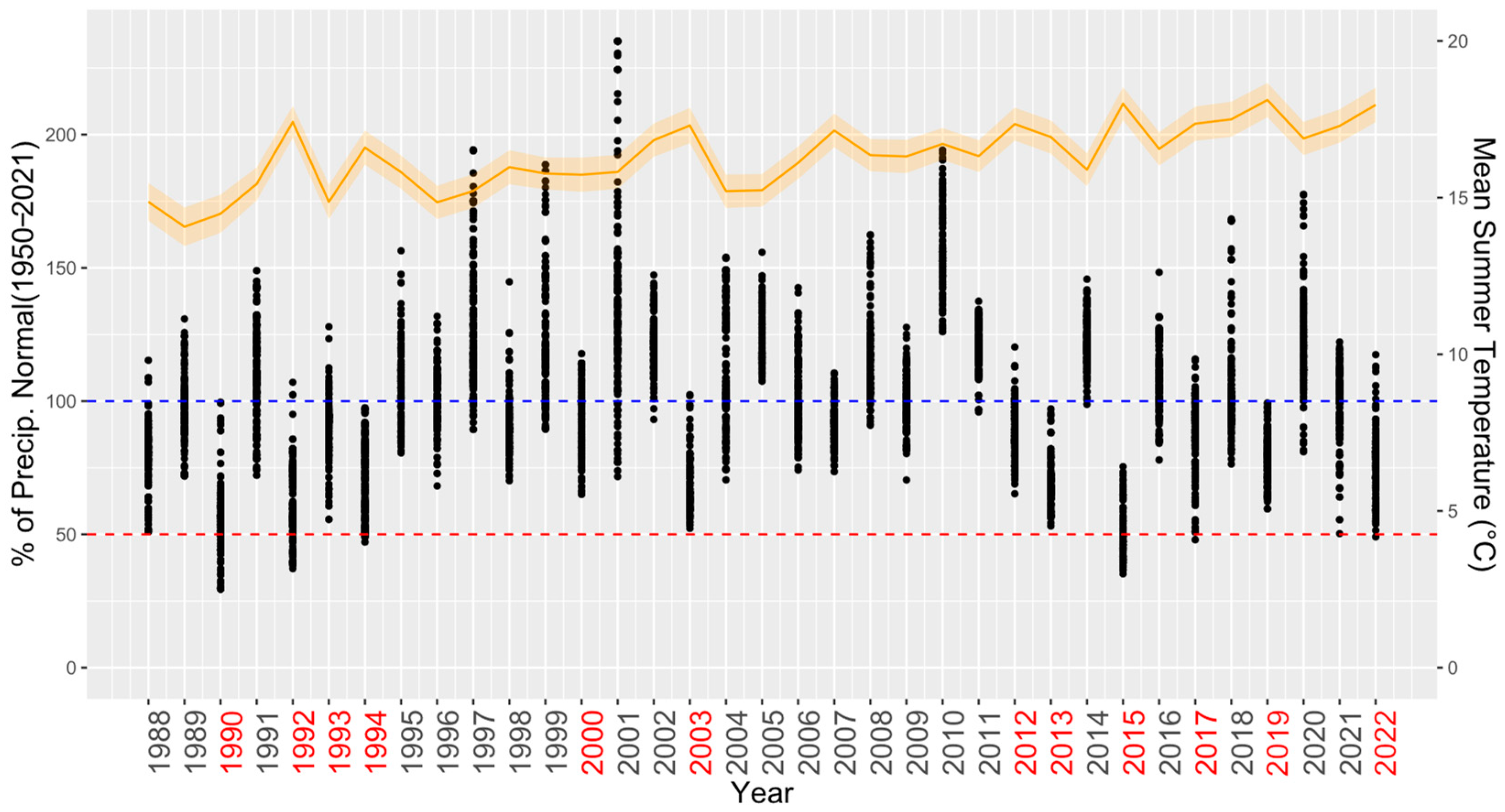
2.3. Site and Climatic Characteristics
2.4. Analysis
3. Results
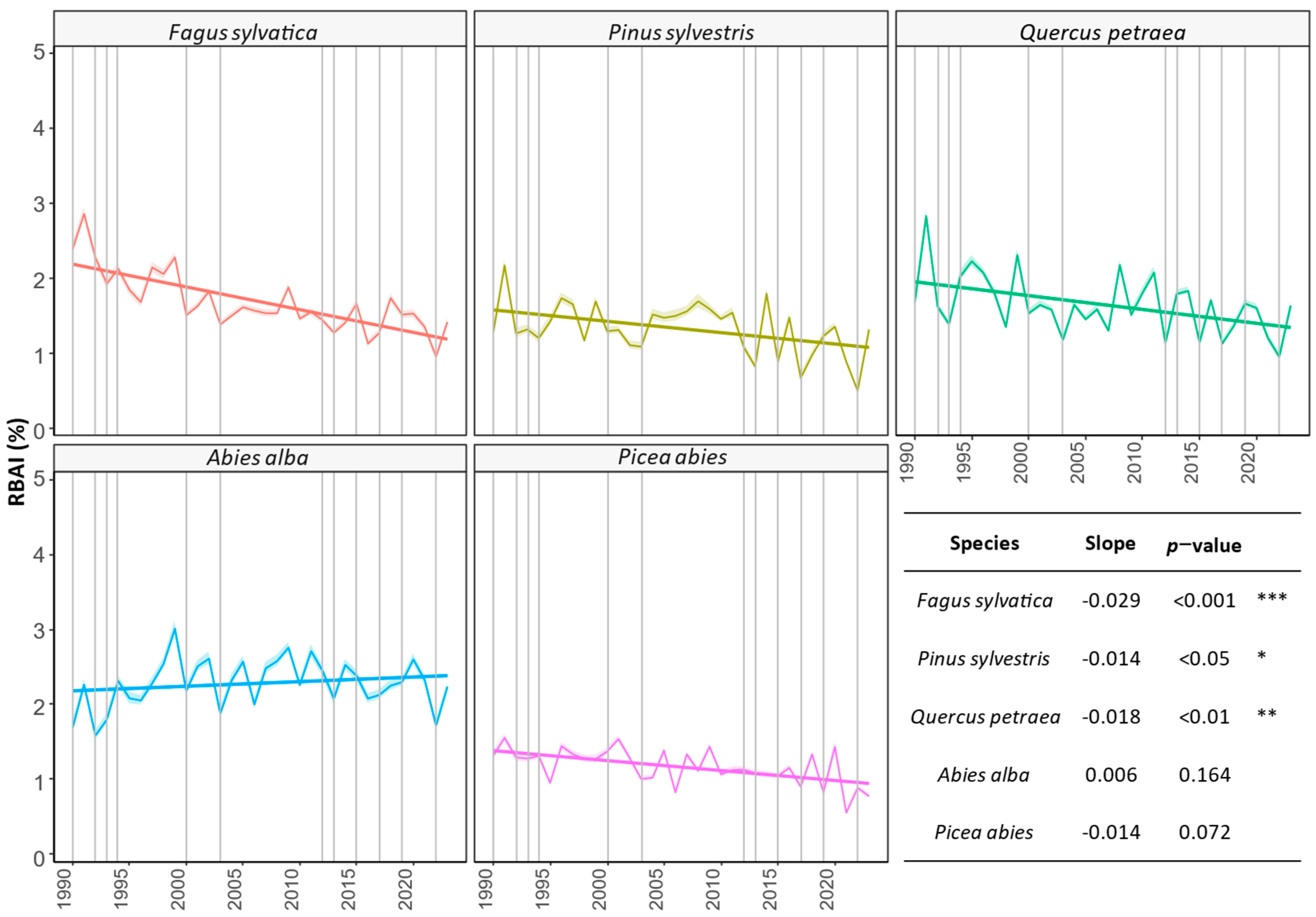
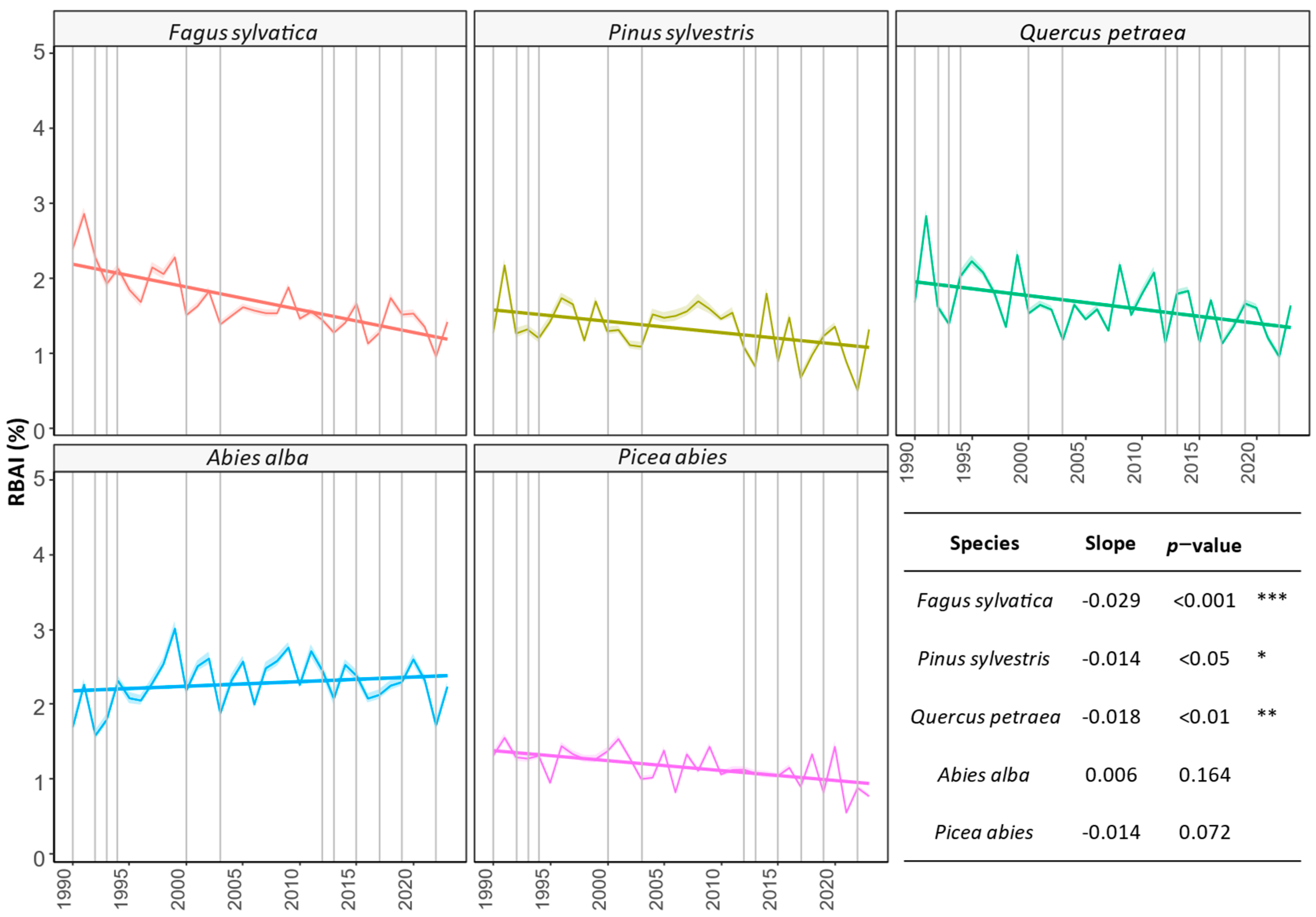
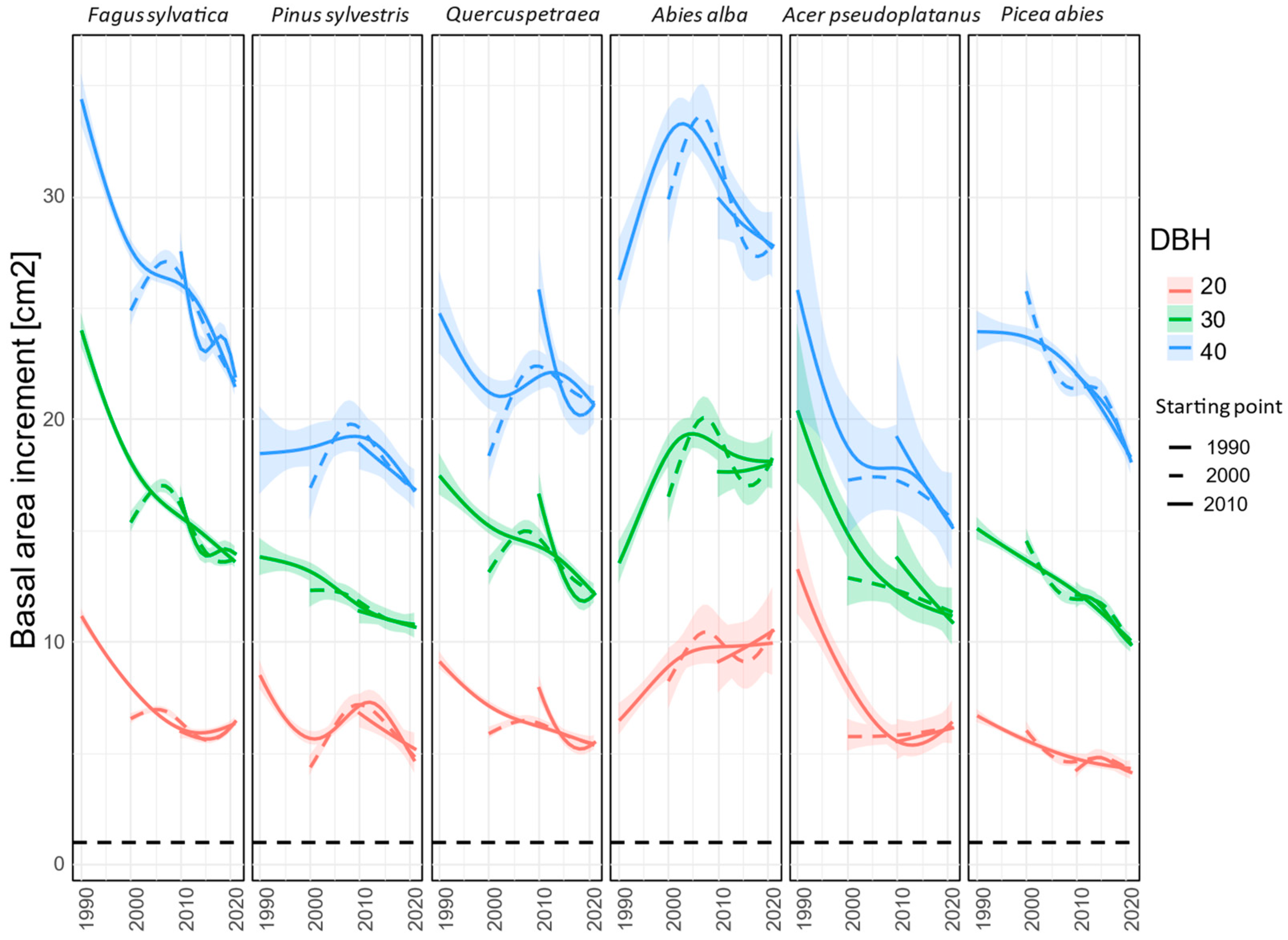
3.1. Comparing Three Decades of Species Growth (H1)
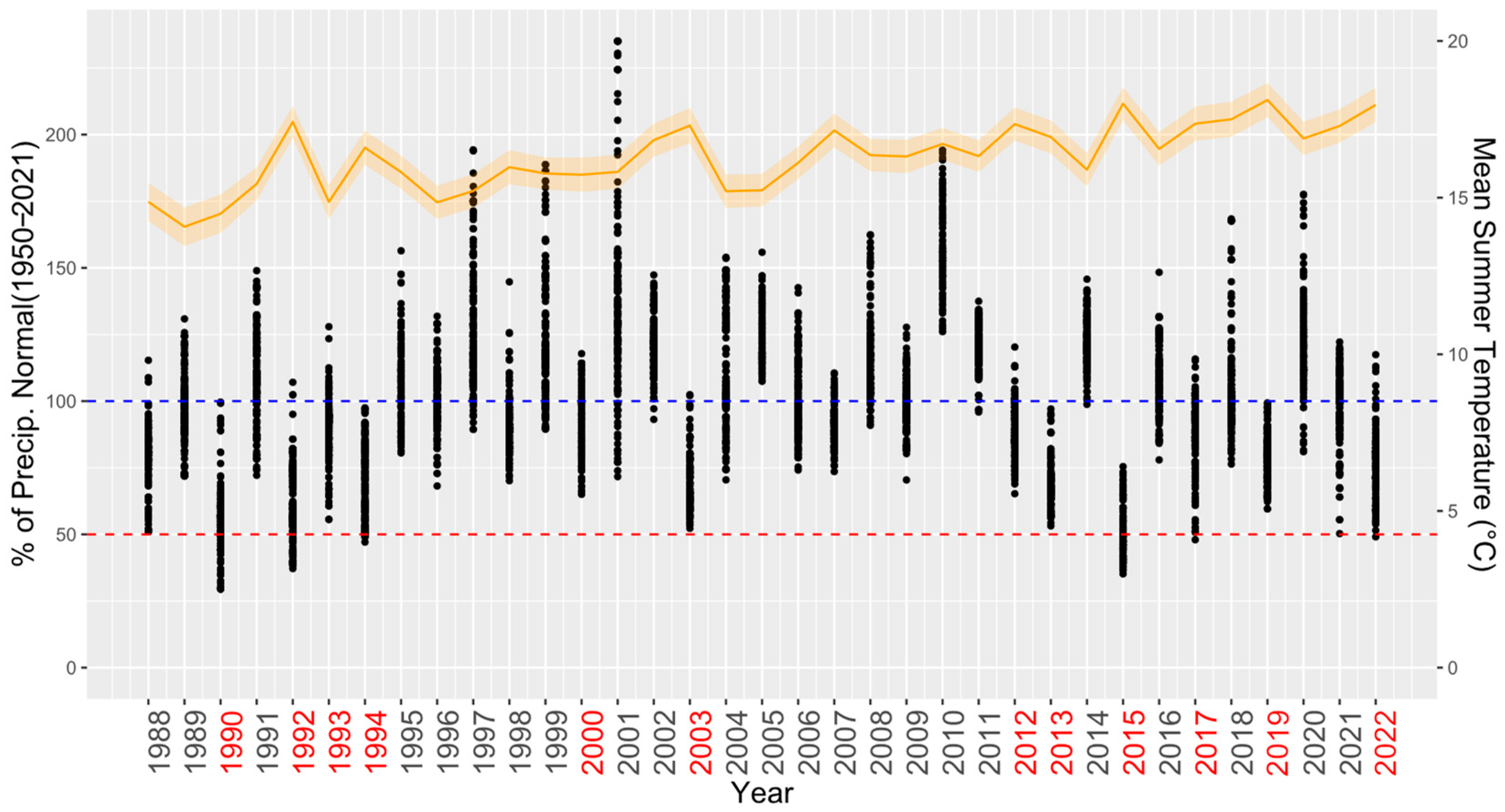
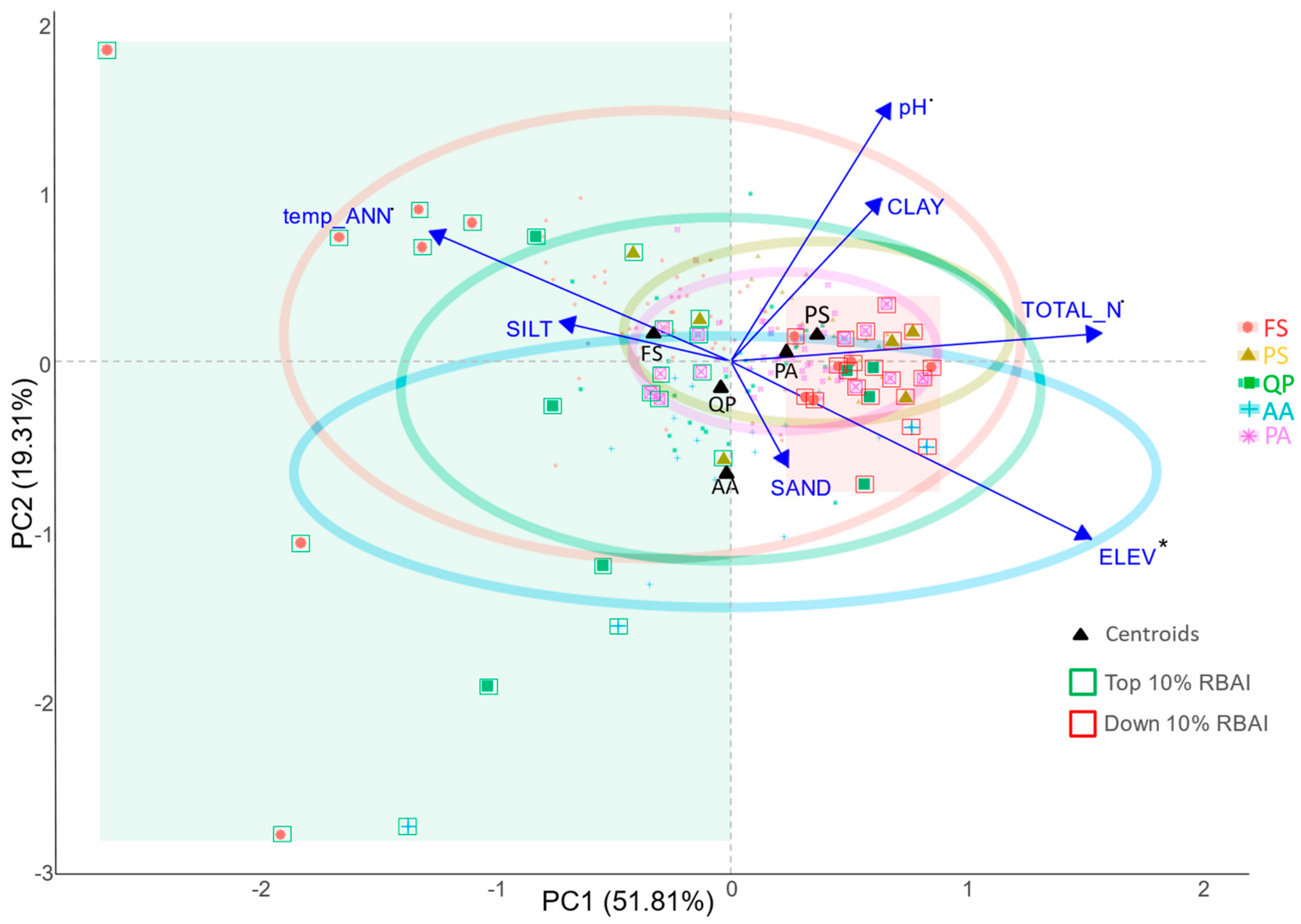
3.2. Comparing Radial Growth in Non-Favourable Climatic Conditions (H2) and Limiting Factors
4. Discussion
4.1. Observed Growth Trends (H1)
4.1.1. Abies alba
4.1.2. Picea abies
4.1.3. Pinus sylvestris
4.1.4. Fagus sylvatica
4.1.5. Quercus petraea
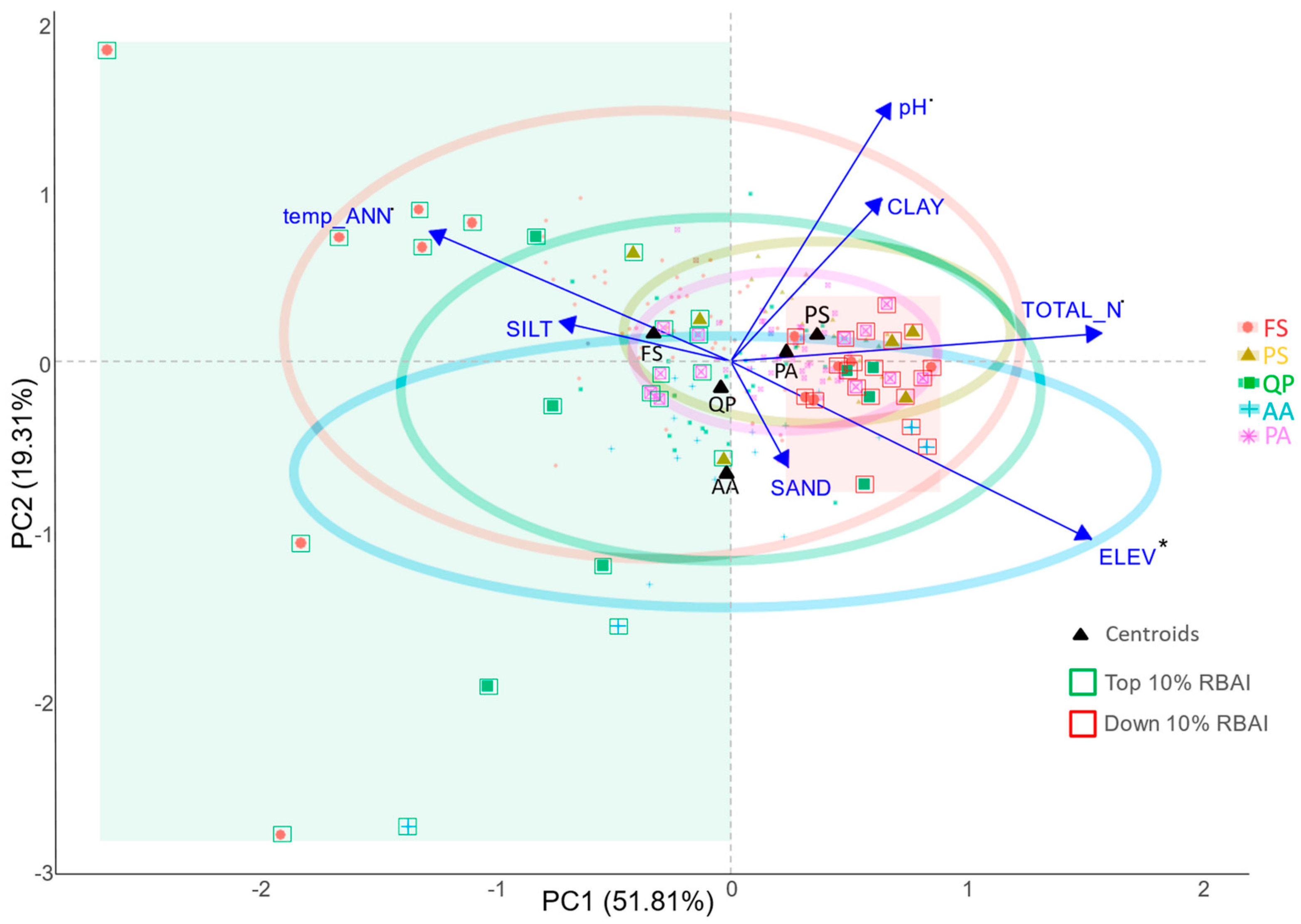
4.2. Adaptation Potential and Limiting Factors (H2)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bolte, A.; Ammer, C.; Löf, M.; Madsen, P.; Nabuurs, G.J.; Schall, P.; Spathelf, P.; Rock, J. Adaptive Forest Management in Central Europe: Climate Change Impacts, Strategies and Integrative Concept. Scand. J. For. Res. 2009, 24, 473–482. [Google Scholar] [CrossRef]
- Jaagus, J.; Aasa, A.; Aniskevich, S.; Boincean, B.; Bojariu, R.; Briede, A.; Danilovich, I.; Fernando, I.; Castro, D.; Dumitrescu, A.; et al. Long-term Changes in Drought Indices in Eastern and Central Europe. Int. J. Clim. 2022, 42, 225–249. [Google Scholar] [CrossRef]
- Blauhut, V.; Stoelzle, M.; Ahopelto, L.; Brunner, M.I.; Teutschbein, C.; Wendt, D.E.; Akstinas, V.; Bakke, S.J.; Barker, L.J.; Bartošová, L.; et al. Lessons from the 2018-2019 European Droughts: A Collective Need for Unifying Drought Risk Management. Nat. Hazard. Earth Syst. 2022, 22, 2201–2217. [Google Scholar] [CrossRef]
- Gera, M.; Damborská, I.; Lapin, M.; Melo, M. Climate Changes in Slovakia: Analysis of Past and Present Observations and Scenarios of Future Developments. In The Handbook of Environmental Chemistry; Springer: Cham, Switzerland, 2017; pp. 21–47. [Google Scholar] [CrossRef]
- Spinoni, J.; Vogt, J.V.; Naumann, G.; Barbosa, P.; Dosio, A. Will Drought Events Become More Frequent and Severe in Europe? Int. J. Clim. 2018, 38, 1718–1736. [Google Scholar] [CrossRef]
- Lapin, M.; Gera, M.; Hrvoľ, J.; Melo, M.; Tomlain, J. Possible Impacts of Climate Change on Hydrologic Cycle in Slovakia and Results of Observations in 1951–2007. Biologia 2009, 64, 454–459. [Google Scholar] [CrossRef]
- Zeleňáková, M.; Vido, J.; Portela, M.M.; Purcz, P.; Blištán, P.; Hlavatá, H.; Hluštík, P. Precipitation Trends over Slovakia in the Period 1981–2013. Water 2017, 9, 922. [Google Scholar] [CrossRef]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On Underestimation of Global Vulnerability to Tree Mortality and Forest Die-off from Hotter Drought in the Anthropocene. Ecosphere 2015, 6, 1–55. [Google Scholar] [CrossRef]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-Wide Reduction in Primary Productivity Caused by the Heat and Drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Schuldt, B.; Buras, A.; Arend, M.; Vitasse, Y.; Beierkuhnlein, C.; Damm, A.; Gharun, M.; Grams, T.E.E.; Hauck, M.; Hajek, P.; et al. A First Assessment of the Impact of the Extreme 2018 Summer Drought on Central European Forests. Basic Appl. Ecol. 2020, 45, 86–103. [Google Scholar] [CrossRef]
- Toreti, A.; Masante, D.; Acosta, N.; Bavera, D. Drought in Europe July 2022. GDO Analytic Report. 2022. Available online: https://reliefweb.int/report/world/gdo-analytical-report-drought-europe-july-2022 (accessed on 10 September 2023).
- Thurm, E.A.; Hernández, L.; Baltensweiler, A.; Ayan, S.; Rasztovits, E.; Bielak, K.; Zlatanov, T.; Hladnik, D.; Balic, B.; Freudenschuss, A.; et al. Alternative Tree Species Under Climate Warming in Managed European Forests. For. Ecol. Manag. 2018, 430, 485–487. [Google Scholar] [CrossRef]
- Hinze, J.; Albrecht, A.; Michiels, H.G. Climate-Adapted Potential Vegetation—A European Multiclass Model Estimating the Future Potential of Natural Vegetation. Forests 2023, 14, 239. [Google Scholar] [CrossRef]
- Bose, A.K.; Scherrer, D.; Camarero, J.J.; Ziche, D.; Babst, F.; Bigler, C.; Bolte, A.; Dorado-Liñán, I.; Etzold, S.; Fonti, P.; et al. Climate Sensitivity and Drought Seasonality Determine Post-Drought Growth Recovery of Quercus Petraea and Quercus Robur in Europe. Sci. Total Environ. 2021, 784, 147222. [Google Scholar] [CrossRef] [PubMed]
- Zang, C.; Hartl-Meier, C.; Dittmar, C.; Rothe, A.; Menzel, A. Patterns of Drought Tolerance in Major European Temperate Forest Trees: Climatic Drivers and Levels of Variability. Glob. Chang. Biol. 2014, 20, 3767–3779. [Google Scholar] [CrossRef] [PubMed]
- Trotsiuk, V.; Babst, F.; Grossiord, C.; Gessler, A.; Forrester, D.I.; Buchmann, N.; Schaub, M.; Eugster, W. Tree Growth in Switzerland Is Increasingly Constrained by Rising Evaporative Demand. J. Ecol. 2021, 109, 2981–2990. [Google Scholar] [CrossRef]
- Bertini, G.; Amoriello, T.; Fabbio, G.; Piovosi, M. Forest Growth and Climate Change: Evidences from the ICP-Forests Intensive Monitoring in Italy. iForest 2011, 4, 262. [Google Scholar] [CrossRef]
- Charru, M.; Seynave, I.; Hervé, J.C.; Bertrand, R.; Bontemps, J.D. Recent Growth Changes in Western European Forests Are Driven by Climate Warming and Structured across Tree Species Climatic Habitats. Ann. Sci. 2017, 74, 33. [Google Scholar] [CrossRef]
- Hilmers, T.; Avdagić, A.; Bartkowicz, L.; Bielak, K.; Binder, F.; Bončina, A.; Dobor, L.; Forrester, D.I.; Hobi, M.L.; Ibrahimspahi, A.; et al. The Productivity of Mixed Mountain Forests Comprised of Fagus sylvatica, Picea abies, and Abies alba across Europe. Forestry 2019, 92, 512–522. [Google Scholar] [CrossRef]
- Kruhlov, I.; Thom, D.; Chaskovskyy, O.; Keeton, W.S.; Scheller, R.M. Future Forest Landscapes of the Carpathians: Vegetation and Carbon Dynamics under Climate Change. Reg. Environ. Chang. 2018, 18, 1555–1567. [Google Scholar] [CrossRef]
- Škvarenina, J.; Križová, E.; Tomlain, J. Impact of the Climate Change on the Water Balance of Altitudinal Vegetation Stages in Slovakia. Ekológia 2004, 23, 13–29. [Google Scholar]
- Morin, X.; Thuiller, W. Comparing Niche- and Process-Based Models to Reduce Prediction Uncertainty in Species Range Shifts under Climate Change. Ecology 2009, 90, 1301–1313. [Google Scholar] [CrossRef]
- Thuiller, W. Patterns and Uncertainties of Species’ Range Shifts under Climate Change. Glob. Chang. Biol. 2004, 10, 2020–2027. [Google Scholar] [CrossRef]
- Seidl, R.; Schelhaas, M.J.; Lindner, M.; Lexer, M.J. Modelling Bark Beetle Disturbances in a Large Scale Forest Scenario Model to Assess Climate Change Impacts and Evaluate Adaptive Management Strategies. Reg. Environ. Chang. 2009, 9, 101–119. [Google Scholar] [CrossRef]
- Itter, M.S.; Finley, A.O.; D’Amato, A.W.; Foster, J.R.; Bradford, J.B. Variable Effects of Climate on Forest Growth in Relation to Climate Extremes, Disturbance, and Forest Dynamics. Ecol. Appl. 2017, 27, 1082–1095. [Google Scholar] [CrossRef] [PubMed]
- Bosela, M.; Lukac, M.; Castagneri, D.; Sedmák, R.; Biber, P.; Carrer, M.; Konôpka, B.; Nola, P.; Nagel, T.A.; Popa, I.; et al. Contrasting Effects of Environmental Change on the Radial Growth of Co-Occurring Beech and Fir Trees across Europe. Sci. Total Environ. 2018, 615, 1460–1469. [Google Scholar] [CrossRef]
- Giebink, C.L.; DeRose, R.J.; Castle, M.; Shaw, J.D.; Evans, M.E.K. Climatic Sensitivities Derived from Tree Rings Improve Predictions of the Forest Vegetation Simulator Growth and Yield Model. For. Ecol. Manag. 2022, 517, 120256. [Google Scholar] [CrossRef]
- Trouvé, R.; Bontemps, J.D.; Collet, C.; Seynave, I.; Lebourgeois, F. Radial Growth Resilience of Sessile Oak after Drought Is Affected by Site Water Status, Stand Density, and Social Status. Trees Struct. Funct. 2017, 31, 517–529. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G.; Uhl, E. Resistance of European Tree Species to Drought Stress in Mixed versus Pure Forests: Evidence of Stress Release by Inter-specific Facilitation. Plant Biol. 2013, 15, 483–495. [Google Scholar] [CrossRef]
- Lorenz, M. International Co-Operative Programme on Assessment and Monitoring of Air Pollution Effects on Forests-ICP Forests-. Water Air Soil Pollut. 1995, 85, 1221–1226. [Google Scholar] [CrossRef]
- Dobbertin, M.; Neumann, M. 2016 Manual on Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests; Thünen Institute of Forest Ecosystems: Eberswalde, Germany, 2016; 17p, Available online: http://www.icp-forests.org/manual.htm (accessed on 5 January 2023).
- Šebeň, V. Národná inventarizácia a monitoring lesov SR 2015-2016. In Informácie, Metódy, Výsledky; Národné Lesnícke Centrum—Lesnícky Výskumný Ústav: Zvolen, Slovakia, 2017; p. 255. ISBN 978-80-8093-234. [Google Scholar]
- Hiederer, R.; Michéli, E.; Durrant, T. Evaluation of BioSoil Demonstration Project; Soil Data Analysis; Joint Research Centre—European Commission: Geel, Belgium, 2011. [Google Scholar] [CrossRef]
- Cornes, R.C.; van der Schrier, G.; van den Besselaar, E.J.M.; Jones, P.D. An Ensemble Version of the E-OBS Temperature and Precipitation Data Sets. J. Geophys. Res. Atmos. 2018, 123, 9391–9409. [Google Scholar] [CrossRef]
- Kolář, T.; Čermák, P.; Trnka, M.; Žid, T.; Rybníček, M. Temporal Changes in the Climate Sensitivity of Norway Spruce and European Beech along an Elevation Gradient in Central Europe. Agric. For. Meteorol. 2017, 239, 24–33. [Google Scholar] [CrossRef]
- Levanič, T.; Ugarković, D.; Seletković, I.; Ognjenović, M.; Marušić, M.; Bogdanić, R.; Potočić, N. Radial Increment of Beech (Fagus sylvatica L.) is Under a Strong Impact of Climate in the Continental Biogeographical Region of Croatia. Plants 2023, 12, 2427. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2023. [Google Scholar]
- Wood, S.N. Thin-plate regression splines. J. R. Stat. Soc. 2003, 65, 95–114. [Google Scholar] [CrossRef]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. Vegan: Community Ecology Package. 2022. Available online: https://github.com/vegandevs/vegan (accessed on 30 October 2023).
- Zhao, M.; Running, S.W. Drought-Induced Reduction in Global Terrestrial Net Primary Production from 2000 through 2009. Science 2010, 329, 940–943. [Google Scholar] [CrossRef]
- Senf, C.; Buras, A.; Zang, C.S.; Rammig, A.; Seidl, R. Excess Forest Mortality Is Consistently Linked to Drought across Europe. Nat. Commun. 2020, 11, 6200. [Google Scholar] [CrossRef] [PubMed]
- Babst, F.; Bouriaud, O.; Alexander, R.; Trouet, V.; Frank, D. Toward Consistent Measurements of Carbon Accumulation: A Multi-Site Assessment of Biomass and Basal area Increment Across Europe. Dendrochronologia 2014, 32, 153–161. [Google Scholar] [CrossRef]
- Díaz, S.; Hector, A.; Wardle, D.A. Biodiversity in Forest Carbon Sequestration Initiatives: Not just a side benefit. Curr Opin. Environ. Sustain. 2009, 1, 55–60. [Google Scholar] [CrossRef]
- Sabatini, F.M.; de Andrade, R.B.; Paillet, Y.; Ódor, P.; Bouget, C.; Campagnaro, T.; Gosselin, F.; Janssen, P.; Mattioli, W.; Nascimbene, J.; et al. Trade-offs Between Carbon Stocks and Biodiversity in European Temperate Forests. Glob. Chang. Biol. 2019, 25, 536–548. [Google Scholar] [CrossRef] [PubMed]
- Cailleret, M.; Jansen, S.; Robert, E.M.R.; Desoto, L.; Aakala, T.; Antos, J.A.; Beikircher, B.; Bigler, C.; Bugmann, H.; Caccianiga, M.; et al. A Synthesis of Radial Growth Patterns Preceding Tree Mortality. Glob. Chang. Biol. 2017, 23, 1675–1690. [Google Scholar] [CrossRef]
- Corcuera, L.; Camarero, J.J.; Gil-Pelegrín, E. Effects of a Severe Drought on Quercus ilex Radial Growth and Xylem Anatomy. Trees 2004, 18, 83–92. [Google Scholar] [CrossRef]
- Vitasse, Y.; Bottero, A.; Rebetez, M.; Conedera, M.; Augustin, S.; Brang, P.; Tinner, W. What Is the Potential of Silver Fir to Thrive under Warmer and Drier Climate? Eur. J. Res. 2019, 138, 547–560. [Google Scholar] [CrossRef]
- Bottero, A.; Forrester, D.I.; Cailleret, M.; Kohnle, U.; Gessler, A.; Michel, D.; Bose, A.K.; Bauhus, J.; Bugmann, H.; Cuntz, M.; et al. Growth Resistance and Resilience of Mixed Silver Fir and Norway Spruce Forests in Central Europe: Contrasting Responses to Mild and Severe Droughts. Glob. Chang. Biol. 2021, 27, 4403–4419. [Google Scholar] [CrossRef]
- Bošeľa, M.; Petráš, R.; Sitková, Z.; Priwitzer, T.; Pajtík, J.; Hlavatá, H.; Sedmák, R.; Tobin, B. Possible Causes of the Recent Rapid Increase in the Radial Increment of Silver Fir in the Western Carpathians. Environ. Pollut. 2014, 184, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Obladen, N.; Dechering, P.; Skiadaresis, G.; Tegel, W.; Keßler, J.; Höllerl, S.; Kaps, S.; Hertel, M.; Dulamsuren, C.; Seifert, T.; et al. Tree Mortality of European Beech and Norway Spruce Induced by 2018–2019 Hot Droughts in Central Germany. Agric. For. Meteorol. 2021, 307, 108482. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gazol, A. Will Silver Fir Be Under Higher Risk Due to Drought? A Comment on Walder et al. (2021). For. Ecol. Manag. 2022, 503, 119826. [Google Scholar] [CrossRef]
- Gomez-Gallego, M.; Galiano, L.; Martínez-Vilalta, J.; Stenlid, J.; Capador-Barreto, H.D.; Elfstrand, M.; Camarero, J.J.; Oliva, J. Interaction of Drought- and Pathogen-Induced Mortality in Norway Spruce and Scots Pine. Plant Cell Environ. 2022, 45, 2292–2305. [Google Scholar] [CrossRef]
- Haberstroh, S.; Werner, C.; Grün, M.; Grün, G.; Kreuzwieser, J.; Seifert, T.; Schindler, D.; Christen, A. Central European 2018 Hot Drought Shifts Scots Pine Forest to Its Tipping Point. Plant Biol. 2018, 24, 1186–1197. [Google Scholar] [CrossRef] [PubMed]
- Sidor, C.G.; Camarero, J.; Popa, I.; Badea, O.; Apostol, E.N.; Vlad, R. Forest Vulnerability to Extreme Climatic Events in Romanian Scots Pine Forests. Sci. Total Environ. 2019, 678, 721–727. [Google Scholar] [CrossRef]
- Leuschner, C. Drought Response of European Beech (Fagus sylvatica L.)—A Review. Perspect. Plant Ecol. Evol. Syst. 2020, 47, 125576. [Google Scholar] [CrossRef]
- Scharnweber, T.; Manthey, M.; Criegee, C.; Bauwe, A.; Schröder, C.; Wilmking, M. Drought Matters–Declining Precipitation Influences Growth of Fagus Sylvatica L. and Quercus Robur L. in North-Eastern Germany. For. Ecol. Manag. 2011, 262, 947–961. [Google Scholar] [CrossRef]
- Rohner, B.; Kumar, S.; Liechti, K.; Gessler, A.; Ferretti, M. Tree Vitality Indicators Revealed a Rapid Response of Beech Forests to the 2018 Drought. Ecol. Indic. 2021, 120, 106903. [Google Scholar] [CrossRef]
- Martinez del Castillo, E.; Zang, C.S.; Buras, A.; Hacket-Pain, A.; Esper, J.; Serrano-Notivoli, R.; Hartl, C.; Weigel, R.; Klesse, S.; de Dios, V.R.; et al. Climate-Change-Driven Growth Decline of European Beech Forests. Commun. Biol. 2022, 5, 163. [Google Scholar] [CrossRef]
- Daněk, P.; Šamonil, P.; Vrška, T. Four Decades of the Coexistence of Beech and Spruce in a Central European Old-Growth Forest. Which Succeeds on What Soils and Why? Plant Soil 2019, 437, 257–272. [Google Scholar] [CrossRef]
- Slanař, J.; Vacek, Z.; Vacek, S.; Bulušek, D.; Cukor, J.; Štefančík, I.; Bílek, L.; Krl, J. Long-Term Transformation of Submontane Spruce-Beech Forests in the Jizerské Hory Mts.: Dynamics of Natural Regeneration. Cent. Eur. For. J. 2017, 63, 212–224. [Google Scholar] [CrossRef]
- Kunz, J.; Löffler, G.; Bauhus, J. Minor European Broadleaved Tree Species Are More Drought-Tolerant than Fagus sylvatica but Not More Tolerant Than Quercus petraea. For. Ecol. Manag. 2018, 414, 15–27. [Google Scholar] [CrossRef]
- Hlásny, T.; Barcza, Z.; Fabrika, M.; Balázs, B.; Churkina, G.; Pajtík, J.; Sedmák, R.; Turčáni, M. Climate Change Impacts on Growth and Carbon Balance of Forests in Central Europe. Clim. Res. 2011, 47, 219–236. [Google Scholar] [CrossRef]
- Fuchs, S.; Schuldt, B.; Leuschner, C. Identification of Drought-Tolerant Tree Species through Climate Sensitivity Analysis of Radial Growth in Central European Mixed Broadleaf Forests. For. Ecol. Manag. 2021, 494, 119287. [Google Scholar] [CrossRef]
- Bosela, M.; Tumajer, J.; Cienciala, E.; Dobor, L.; Kulla, L.; Marčiš, P.; Popa, I.; Sedmák, R.; Sedmáková, D.; Sitko, R.; et al. Climate Warming Induced Synchronous Growth Decline in Norway Spruce Populations across Biogeographical Gradients since 2000. Sci. Total Environ. 2021, 752, 14194. [Google Scholar] [CrossRef] [PubMed]
- Ponocná, T.; Spyt, B.; Kaczka, R.; Büntgen, U.; Treml, V. Growth Trends and Climate Responses of Norway Spruce along Elevational Gradients in East-Central Europe. Trees Struct. Funct. 2016, 30, 1633–1646. [Google Scholar] [CrossRef]
- King, G.M.; Gugerli, F.; Fonti, P.; Frank, D.C. Tree Growth Response along an Elevational Gradient: Climate or Genetics? Oecologia 2013, 173, 1587–1600. [Google Scholar] [CrossRef]
- Csilléry, K.; Ovaskainen, O.; Sperisen, C.; Buchmann, N.; Widmer, A.; Gugerli, F. Adaptation to Local Climate in Multi-Trait Space: Evidence from Silver Fir (Abies alba Mill.) Populations Across a Heterogeneous Environment. Heredity 2020, 124, 77–92. [Google Scholar] [CrossRef]
- Dinca, L.; Marin, M.; Radu, V.; Murariu, G.; Drasovean, R.; Cretu, R.; Georgescu, L.; Timiș-gânsac, V. Which are the Best Site and Stand Conditions for Silver Fir (Abies Alba Mill.) Located in the Carpathian Mountains? Diversity 2022, 14, 547. [Google Scholar] [CrossRef]
- Vospernik, S.; Nothdurft, A. Can Trees at High Elevations Compensate for Growth Reductions at Low Elevations Due to Climate Warming? Can. J. For. Res. 2018, 48, 650–662. [Google Scholar] [CrossRef]
- Gessler, A.; Bächli, L.; Rouholahnejad Freund, E.; Treydte, K.; Schaub, M.; Haeni, M.; Weiler, M.; Seeger, S.; Marshall, J.; Hug, C.; et al. Drought Reduces Water Uptake in Beech from the Drying Topsoil, but No Compensatory Uptake Occurs from Deeper Soil Layers. New Phytol. 2022, 233, 194–206. [Google Scholar] [CrossRef] [PubMed]
- Bosela, M.; Kulla, L.; Roessiger, J.; Šebeň, V.; Dobor, L.; Büntgen, U.; Lukac, M. Long-Term Effects of Environmental Change and Species Diversity on Tree Radial Growth in a Mixed European Forest. For. Ecol. Manag. 2019, 446, 293–303. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Species Proportion [% ± SD] (NFI) | Number of Plots (ICP Forests I) | Number of Individuals (ICP Forests I) | Number of Increment Records (ICP Forests I) |
|---|---|---|---|---|
| Fagus sylvatica | 30.1 ± 2.4 | 67 | 2251 | 53,255 |
| Picea abies | 18.7 ± 2.0 | 55 | 2236 | 43,438 |
| Quercus petraea | 7.2 ± 1.3 | 34 | 651 | 14,703 |
| Pinus sylvestris | 4.8 ± 1.1 | 27 | 643 | 14,117 |
| Abies alba | 2.8 ± 0.9 | 19 | 351 | 7485 |
| Σ | 63.6 | 115 orig. plots | 6132 | 132,998 |
| Site Characteristics | R2 | p-Value | |
|---|---|---|---|
| ELEVATION | 3.45 | 0.028 | * |
| ASPECT | 0.75 | 0.493 | |
| SLOPE | 0.22 | 0.808 | |
| TREE RICHNESS | 1.53 | 0.205 | |
| pH (H2O) | 2.79 | 0.058 | . |
| CLAY | 1.34 | 0.279 | |
| SILT | 0.58 | 0.564 | |
| SAND | 0.45 | 0.657 | |
| TOTAL_N | 2.51 | 0.082 | . |
| ORGANIC_C | 1.84 | 0.151 | |
| C_N_ratio | 0.23 | 0.801 | |
| precip_ANN | 2.47 | 0.095 | . |
| temp_ANN | 2.21 | 0.120 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rybar, J.; Sitková, Z.; Marcis, P.; Pavlenda, P.; Pajtík, J. Declining Radial Growth in Major Western Carpathian Tree Species: Insights from Three Decades of Temperate Forest Monitoring. Plants 2023, 12, 4081. https://doi.org/10.3390/plants12244081
Rybar J, Sitková Z, Marcis P, Pavlenda P, Pajtík J. Declining Radial Growth in Major Western Carpathian Tree Species: Insights from Three Decades of Temperate Forest Monitoring. Plants. 2023; 12(24):4081. https://doi.org/10.3390/plants12244081
Chicago/Turabian StyleRybar, Jergus, Zuzana Sitková, Peter Marcis, Pavel Pavlenda, and Jozef Pajtík. 2023. "Declining Radial Growth in Major Western Carpathian Tree Species: Insights from Three Decades of Temperate Forest Monitoring" Plants 12, no. 24: 4081. https://doi.org/10.3390/plants12244081
APA StyleRybar, J., Sitková, Z., Marcis, P., Pavlenda, P., & Pajtík, J. (2023). Declining Radial Growth in Major Western Carpathian Tree Species: Insights from Three Decades of Temperate Forest Monitoring. Plants, 12(24), 4081. https://doi.org/10.3390/plants12244081