
Road Impact on Plant Colonization in the Arid Timanfaya National Park

 , ,
, , 
Abstract
:1. Introduction
2. Results
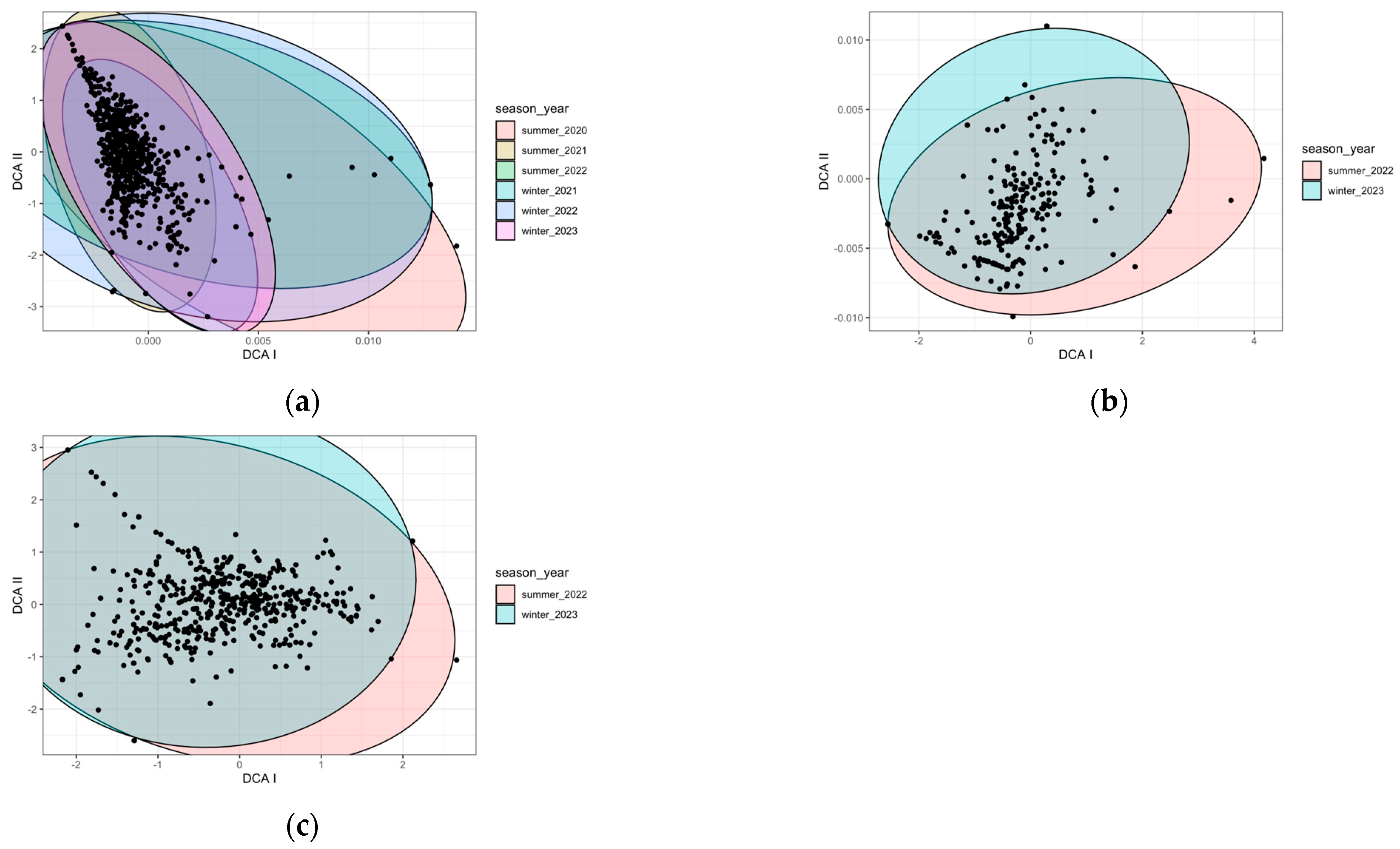
2.1. Species Composition
2.1.1. Zones
2.1.2. Position
2.2. Species Richness
Species Richness of Endemic, Native, Introduced, and Invasive Species
3. Discussion
3.1. Species Composition
3.2. Species Richness
4. Materials and Methods
4.1. Study Site
4.2. Design of the Experiment
4.3. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species [42,44,45,55,56,57] | Family [42,55] | Endemicity * [42] |
|---|---|---|
| Aeonium lancerottense | Crassulaceae | *NS |
| Agave americana | Agavaceae | II |
| Aichryson tortuosum | Crassulaceae | *NS |
| Aizoon canariense | Aizoaceae | NS |
| Ajuga iva | Lamiaceae | NP |
| Amaranthus viridis | Amaranthaceae | IS |
| Anacyclus radiatus ssp. coronatus | Asteraceae | NS |
| Andryala perezii | Asteraceae | *NS |
| Arenaria leptoclados | Caryophyllaceae | NP |
| Asparagus horridus | Convallariaceae | NS |
| Asphodelus tenuifolius | Asphodelaceae | NP |
| Asteriscus intermedius | Asteraceae | *NS |
| Astragalus solandri | Fabaceae | NO |
| Atractylis cancellata | Asteraceae | NP |
| Atriplex semibaccata | Chenopodiaceae | II |
| Avena canariensis | Poaceae | *NS |
| Brachypodium stacei | Poaceae | NP |
| Bromus madritensis | Poaceae | NO |
| Bromus rubens | Poaceae | NS |
| Bupleurum semicompositum | Apiaceae | NS |
| Calendula afin tripterocarpa | Asteraceae | NP |
| Campanula erinus | Campanulaceae | NO |
| Carrichtera annua | Brassicaceae | NP |
| Centaurea melitensis | Asteraceae | NO |
| Chamaesyce serpens | Euphorbiaceae | IP |
| Chenopodium murale | Chenopodiaceae | IP |
| Crassula tillaea | Crassulaceae | NO |
| Cuscuta approximata | Cuscutaceae | NP |
| Cyperus capitatus | Cyperaceae | NS |
| Dipcadi serotinum | Liliaceae | NP |
| Echium lancerottense | Boraginaceae | *NS |
| Emex spinosa | Polygonaceae | NP |
| Eragrostis barrelieri | Poaceae | NO |
| Erigeron floribundus | Asteraceae | IS |
| Erodium cicutarium | Geraniaceae | NO |
| Erodium hesperium | Geraniaceae | NO |
| Erodium neuradifolium | Geraniaceae | NP |
| Erodium salzmannii | Geraniaceae | NO |
| Erodium touchyanum | Geraniaceae | NO |
| Erucastrum canariense | Brassicaceae | *NS |
| Euphorbia peplus var. minima | Euphorbiaceae | NO |
| Euphorbia terracina | Euphorbiaceae | NP |
| Fagonia cretica | Zygophyllaceae | NP |
| Ficus carica | Moraceae | IP |
| Filago desertorum | Asteraceae | NP |
| Foeniculum vulgare ssp. piperitum | Apiaceae | NP |
| Forsskaolea angustifolia | Urticaceae | *NS |
| Frankenia capitata | Frankeniaceae | NS |
| Fumaria montana | Fumariaceae | NS |
| Galium spurium | Rubiaceae | NO |
| Geranium rotundifolium | Geraniaceae | NO |
| Glebionis coronaria | Asteraceae | IP |
| Hedypnois rhagadioloides | Asteraceae | NO |
| Helianthemum canariense | Cistaceae | NS |
| Heliotropium bacciferum | Boraginaceae | NS |
| Hirschfeldia incana | Brassicaceae | NO |
| Hordeum murinum ssp. glaucum | Poaceae | NO |
| Hyoscyamus albus | Solanaceae | NO |
| Hyparrhenia sinaica | Poaceae | NS |
| Ifloga spicata | Asteraceae | NO |
| Juncus acutus | Juncaceae | NP |
| Kickxia sagittata | Scrophulariaceae | NS |
| Kleinia neriifolia | Asteraceae | *NS |
| Lactuca serriola | Asteraceae | NO |
| Lamarckia aurea | Poaceae | NP |
| Lasiopogon muscoides | Asteraceae | NS |
| Launaea arborescens | Asteraceae | NS |
| Launaea nudicaulis | Asteraceae | NP |
| Lavandula pinnata | Lamiaceae | NS |
| Leontodon taraxacoides ssp. longirostris | Asteraceae | IP |
| Linum strictum | Linaceae | NP |
| Lotus glinoides | Fabaceae | NP |
| Lotus lancerottensis | Fabaceae | NS |
| Lycopersicon esculentum | Solanaceae | IS |
| Lysimachia arvensis | Primulaceae | NO |
| Malva parviflora | Malvaceae | NO |
| Medicago laciniata | Fabaceae | NS |
| Medicago littoralis | Fabaceae | NS |
| Medicago polymorpha | Fabaceae | NO |
| Melilotus sulcatus | Fabaceae | IS |
| Mercurialis annua | Euphorbiaceae | IP |
| Mesembryanthemum crystallinum | Aizoaceae | NO |
| Mesembryanthemum nodiflorum | Aizoaceae | NO |
| Micromeria mahanensis | Lamiaceae | NS |
| Misopates salvagense | Scrophulariaceae | NS |
| Nicotiana glauca | Solanaceae | II |
| Notoceras bicorne | Brassicaceae | NP |
| Oligomeris linifolia | Resedaceae | NS |
| Ononis hesperia | Fabaceae | NS |
| Ononis serrata | Fabaceae | NO |
| Ononis tournefortii | Fabaceae | NP |
| Ophioglossum polyphyllum | Ophioglossaceae | NS |
| Opuntia dillenii | Cactaceae | II |
| Papaver somniferum ssp. setigerum | Papaveraceae | NO |
| Parietaria debilis | Urticaceae | NP |
| Patellifolia patellaris | Chenopodiaceae | NS |
| Pelargonium capitatum | Geraniaceae | II |
| Phagnalon rupestre | Asteraceae | NP |
| Phagnalon saxatile | Asteraceae | NP |
| Phelipanche gratiosa | Orobanchaceae | *NS |
| Plantago afra | Plantaginaceae | NO |
| Plantago coronopus | Plantaginaceae | NP |
| Plantago lagopus | Plantaginaceae | NP |
| Plantago ovata | Plantaginaceae | NP |
| Polycarpaea divaricata | Caryophyllaceae | *NS |
| Polycarpaea robusta | Caryophyllaceae | *NS |
| Polycarpon tetraphyllum | Caryophyllaceae | NO |
| Portulaca canariensis | Portulacaceae | *NS |
| Reichardia tingitana | Asteraceae | NP |
| Reseda crystallina | Brassicaceae | *NS |
| Rostraria pumila | Poaceae | NP |
| Rumex lunaria | Polygonaceae | *NS |
| Rumex vesicarius | Polygonaceae | NP |
| Salvia aegyptiaca | Lamiaceae | NS |
| Schismus barbatus | Poaceae | NO |
| Scrophularia arguta | Scrophulariaceae | NS |
| Senecio flavus | Asteraceae | NP |
| Senecio leucanthemifolius | Asteraceae | NP |
| Senecio massaicus | Asteraceae | NP |
| Senecio vulgaris | Asteraceae | NO |
| Silene apetala | Caryophyllaceae | NO |
| Silene nocturna | Caryophyllaceae | NO |
| Sisymbrium erysimoides | Brassicaceae | NO |
| Sonchus asper | Asteraceae | NO |
| Sonchus bourgeaui | Asteraceae | NS |
| Sonchus oleraceus | Asteraceae | NO |
| Sonchus tenerrimus | Asteraceae | NO |
| Spergula fallax | Caryophyllaceae | NP |
| Spergularia fimbriata | Caryophyllaceae | NP |
| Stipa capensis | Poaceae | NP |
| Torilis elongata | Apiaceae | NO |
| Trachynia distachya | Poaceae | NO |
| Trigonella stellata | Fabaceae | NS |
| Trisetaria loeflingiana | Poaceae | *NS |
| Umbilicus gaditanus | Crassulaceae | NP |
| Urospermum picroides | Asteraceae | NO |
| Vicia benghalensis | Fabaceae | IP |
| Volutaria tubuliflora | Asteraceae | NO |
| Wahlenbergia lobelioides | Campanulaceae | NS |
| 2020 | 2021 | 2022 | 2023 | |||||
|---|---|---|---|---|---|---|---|---|
| Zones | Road Type | Position | Summer | Winter | Summer | Winter | Summer | Winter |
| Altos Timanfaya | Road | Border | 4 | 17 | 4 | 14 | 6 | 21 |
| Middle | 2 | 12 | 3 | 8 | 3 | 19 | ||
| Interior | 3 | 12 | 3 | 8 | 6 | 23 | ||
| Calderas Quemadas | Road | Border | 3 | 12 | 4 | 5 | 4 | 7 |
| Middle | 0 | 8 | 1 | 0 | 1 | 3 | ||
| Interior | 1 | 6 | 1 | 0 | 0 | 2 | ||
| Caldereta Riscada | Road | Border | 3 | 17 | 8 | 8 | 5 | 15 |
| Middle | 3 | 14 | 6 | 8 | 6 | 14 | ||
| Interior | 1 | 12 | 3 | 3 | 3 | 12 | ||
| Islote | Road | Border | 3 | 52 | 21 | 45 | 26 | 58 |
| Middle | 2 | 22 | 4 | 13 | 8 | 27 | ||
| Interior | 2 | 13 | 2 | 5 | 3 | 13 | ||
| Manto Virgen | Road | Border | 2 | 10 | 3 | 4 | 2 | 6 |
| Middle | 1 | 5 | 1 | 1 | 2 | 3 | ||
| Interior | 2 | 8 | 2 | 4 | 2 | 4 | ||
| Mazo | Trail | Border | 2 | 34 | 7 | 9 | 8 | 26 |
| Middle | 2 | 15 | 5 | 5 | 4 | 15 | ||
| Interior | 2 | 7 | 6 | 4 | 3 | 12 | ||
| Montaña Rajada | Road | Border | 2 | 16 | 5 | 7 | 5 | 14 |
| Middle | 1 | 8 | 2 | 2 | 3 | 9 | ||
| Interior | 1 | 12 | 3 | 2 | 1 | 7 | ||
| Taro | Road | Border | 2 | 53 | 25 | 42 | 18 | 65 |
| Middle | 2 | 20 | 5 | 10 | 6 | 29 | ||
| Interior | 1 | 19 | 3 | 6 | 6 | 22 | ||
| Tremesana | Trail | Border | 9 | 28 | 7 | 6 | 6 | 36 |
| Middle | 1 | 10 | 1 | 2 | 2 | 19 | ||
| Interior | 1 | 5 | 1 | 0 | 0 | 4 | ||
| Valle Tranquilidad | Road | Border | 2 | 18 | 6 | 8 | 7 | 14 |
| Middle | 1 | 13 | 5 | 9 | 5 | 18 | ||
| Interior | 1 | 10 | 3 | 6 | 6 | 11 | ||
| 2022 | 2023 | |||
|---|---|---|---|---|
| Zone | Road Type | Position | Summer | Winter |
| Yaiza | Road | Border | 16 | 58 |
| Middle | 5 | 39 | ||
| Interior | 6 | 27 | ||
| Echadero Camellos | Road | Border | 16 | 50 |
| Middle | 1 | 16 | ||
| Interior | 2 | 8 | ||
| Taro 1 | Road | Border | 15 | 60 |
| Middle | 6 | 25 | ||
| Interior | 7 | 27 | ||
| Centro Visitantes | Road | Border | 20 | 78 |
| Middle | 5 | 55 | ||
| Interior | 8 | 46 |
| (a) | ||||
|---|---|---|---|---|
| Species Richness | ||||
| Zone | Time | Native/Non-Native | Endemicity | No. of Species |
| Altos Timanfaya | winter 2021 | Native | *NS | 3 |
| N | 14 | |||
| Non-native | I | 0 | ||
| II | 2 | |||
| winter 2023 | Native | *NS | 6 | |
| N | 21 | |||
| Non-native | I | 1 | ||
| II | 2 | |||
| Calderas Quemadas | winter 2021 | Native | *NS | 2 |
| N | 10 | |||
| Non-native | I | 0 | ||
| II | 1 | |||
| winter 2023 | Native | *NS | 2 | |
| N | 4 | |||
| Non-native | I | 0 | ||
| II | 1 | |||
| Caldereta Riscada | winter 2021 | Native | *NS | 3 |
| N | 14 | |||
| Non-native | I | 0 | ||
| II | 2 | |||
| winter 2023 | Native | *NS | 3 | |
| N | 14 | |||
| Non-native | I | 0 | ||
| II | 2 | |||
| Islote | winter 2021 | Native | *NS | 6 |
| N | 44 | |||
| Non-native | I | 2 | ||
| II | 3 | |||
| winter 2023 | Native | *NS | 6 | |
| N | 47 | |||
| Non-native | I | 4 | ||
| II | 3 | |||
| Manto Virgen | winter 2021 | Native | *NS | 3 |
| N | 9 | |||
| Non-native | I | 0 | ||
| II | 1 | |||
| winter 2023 | Native | *NS | 2 | |
| N | 5 | |||
| Non-native | I | 0 | ||
| II | 1 | |||
| Mazo | winter 2021 | Native | *NS | 5 |
| N | 27 | |||
| Non-native | I | 1 | ||
| II | 2 | |||
| winter 2023 | Native | *NS | 4 | |
| N | 20 | |||
| Non-native | I | 2 | ||
| II | 2 | |||
| Montaña Rajada | winter 2021 | Native | *NS | 4 |
| N | 12 | |||
| Non-native | I | 0 | ||
| II | 2 | |||
| winter 2023 | Native | *NS | 2 | |
| N | 13 | |||
| Non-native | I | 0 | ||
| II | 1 | |||
| Taro | winter 2021 | Native | *NS | 6 |
| N | 47 | |||
| Non-native | I | 2 | ||
| II | 2 | |||
| winter 2023 | Native | *NS | 8 | |
| N | 57 | |||
| Non-native | I | 5 | ||
| II | 2 | |||
| Tremesana | winter 2021 | Native | *NS | 4 |
| N | 23 | |||
| Non-native | I | 2 | ||
| II | 1 | |||
| winter 2023 | Native | *NS | 4 | |
| N | 27 | |||
| Non-native | I | 3 | ||
| II | 2 | |||
| Valle Tranquilidad | winter 2021 | Native | *NS | 5 |
| N | 13 | |||
| Non-native | I | 0 | ||
| II | 2 | |||
| winter 2023 | Native | *NS | 5 | |
| N | 15 | |||
| Non-native | I | 0 | ||
| II | 2 | |||
| (b) | ||||
| Species Richness | ||||
| Zone | Time | Native/Non-Native | Endemicity | No. of Species |
| Centro Visitantes | summer 2022 | Native | *NS | 5 |
| N | 15 | |||
| Non-native | I | 1 | ||
| II | 2 | |||
| winter 2023 | Native | *NS | 9 | |
| N | 74 | |||
| Non-native | I | 7 | ||
| II | 2 | |||
| Echadero Camellos | summer 2022 | Native | *NS | 5 |
| N | 6 | |||
| Non-native | I | 2 | ||
| II | 3 | |||
| winter 2023 | Native | *NS | 6 | |
| N | 37 | |||
| Non-native | I | 4 | ||
| II | 3 | |||
| Taro 1 | summer 2022 | Native | *NS | 3 |
| N | 10 | |||
| Non-native | I | 1 | ||
| II | 2 | |||
| winter 2023 | Native | *NS | 4 | |
| N | 52 | |||
| Non-native | I | 4 | ||
| II | 3 | |||
| Yaiza | summer 2022 | Native | *NS | 4 |
| N | 9 | |||
| Non-native | I | 0 | ||
| II | 5 | |||
| winter 2023 | Native | *NS | 6 | |
| N | 50 | |||
| Non-native | I | 3 | ||
| II | 5 | |||

References
- Bennett, A.F. Roads, roadsides and wildlife conservation: A review. In Nature Conservation; Saunders, D.A., Hobbs, R.J., Eds.; 2: The Role of Corridors; Surrey Beatty & Sons: Chipping Norton, UK, 1991; pp. 99–118. [Google Scholar]
- Arévalo, J.R.; Otto, R.; Escudero, C.; Fernández-Lugo, S.; Arteaga, M.; Delgado, J.; Fernández-Palacios, J.M. Do anthropogenic corridors homogenize plant communities at a local scale? A case studied in Tenerife (Canary Islands). Plant Ecol. 2010, 209, 23–35. [Google Scholar] [CrossRef]
- Spellerberg, I.F. 1998. Ecological effects of roads and traffic: A literature review. Glob. Ecol. Biogeogr. Let. 1998, 7, 317–333. [Google Scholar]
- Trombulak, S.C.; Frissell, C.A. Review of Ecological Effects of Roads on Terrestrial and Aquatic Communities. Conserv. Biol. 2001, 14, 18–30. [Google Scholar] [CrossRef]
- Song, I.J.; Hong, S.K.; Hyun, O.; Byun, B.; Gin, Y. The pattern of landscape patches and invasion of naturalized plants in developed areas of urban Seoul. Landsc. Urban Plan. 2005, 70, 205–219. [Google Scholar] [CrossRef]
- Jaarsma, R.; Willems, G. Reducing habitat fragmentation by minor rural roads through traffic calming. Landsc. Urban Plan. 2002, 58, 125–135. [Google Scholar] [CrossRef]
- Serrano, M.; Sanz, L.; Puig, J.; Pons, J. Landscape fragmentation caused by the transport network in Navarra (Spain): Two-scale analysis and landscape integration assessment. Landsc. Urban Plan. 2002, 58, 113–123. [Google Scholar] [CrossRef]
- Andrews, A. Fragmentation of habitat by roads and utility corridors: A review. Aust. Zool. 1990, 26, 130–141. [Google Scholar] [CrossRef]
- Marcantonio, M.; Rocchini, D.; Geri, F.; Bacaro, G.; Amici, V. Biodiversity, Roads and Landscape Fragmentation: Two Mediterranean Cases. Appl. Geogr. 2013, in press. [Google Scholar] [CrossRef]
- Fernández-Palacios, J.M.; Schrader, J.; de Nascimento, L.; Irl, S.; Sánchez-Pinto, L.; Otto, R. Are plant communities on the Canary Islands resistant to plant invasion? Divers. Distrib. 2022, 29, 51–60. [Google Scholar] [CrossRef]
- Iseli, E.; Chisholm, C.; Lenoir, J.; Haider, S.; Seipel, T.; Barros, A.; Hargreaves, A.; Kardol, P.; Lembrechts, J.; Mcdougall, K.; et al. Rapid upwards spread of non-native plants in mountains across continents. Nat. Ecol. Evol. 2023, 7, 405–413. [Google Scholar] [CrossRef]
- Mcdougall, K.; Lembrechts, J.; Rew, L.; Haider, S.; Cavieres, L.; Kueffer, C.; Milbau, A.; Naylor, B.; Nuñez, M.; Pauchard, A.; et al. Running off the road: Roadside non-native plants invading mountain vegetation. Biol. Invasions 2018, 20, 3461–3473. [Google Scholar] [CrossRef]
- Seipel, T.; Kueffer, C.; Rew, L.; Daehler, C.C.; Pauchard, A.; Naylor, B.J.; Alexander, J.M.; Parks, C.G.; Edwards, P.J.; Arevalo Sierra, J.R.; et al. Processes at multiple scales affect nonnative plant species richness and similarity in mountains around the world. Glob. Ecol. Biogeogr. 2012, 21, 236–246. [Google Scholar] [CrossRef]
- Pauchard, A.; Alaback, P. Influence of Elevation, Land Use, and Landscape Context on Patterns of Alien Plant Invasions along Roadsides in Protected Areas of South-Central Chile. Conserv. Biol. 2004, 18, 238–248. [Google Scholar] [CrossRef]
- Forman, R.T.T.; Alexander, L.E. Roads and Their Major Ecological Effects. Annu. Rev. Ecol. Syst. 1998, 29, 207–231. [Google Scholar] [CrossRef]
- Ballantyne, M.; Pickering, C.M. The impacts of trail infrastructure on vegetation and soils: Current literature and future directions. J Env. Manag. 2015, 164, 53–64. [Google Scholar] [CrossRef]
- Liedtke, R.; Barros, A.; Essl, F.; Lembrechts, J.J.; Wedegärtner, R.E.M.; Pauchard, A.; Dullinger, S. Hiking trails as conduits for the spread of non-native species in mountain areas. Biol. Invasions 2020, 22, 1121–1134. [Google Scholar] [CrossRef]
- Turner, S.; Esler, K.; Kalwij, J. Road verges facilitate exotic species expansion into undisturbed natural montane grasslands. Appl. Veg. Sci. 2021, 24, e12615. [Google Scholar] [CrossRef]
- Barni, P.; Fearnside, P.; De Alencastro Graça, P.M. Desmatamento no sul do Estado de Roraima: Padrões de distribuição em função de Projetos de Assentamento do INCRA e da distância das principais rodovias (BR-174 e BR-210). Acta Amaz. 2012, 42, 195–204. [Google Scholar] [CrossRef]
- Kollarou, V.; Lantitsou, K.; Athanasopoulou, A.; Kollaros, G. Impact of roads on ecological conditions. In Proceedings of the 4th International Conference on Environmental Management, Engineering, Planning and Economics (CEMEPE) and SECOTOX Conference, Mykonos Island, Greece, 24–28 June 2013; ISBN 978-960-6865-68-8. [Google Scholar]
- Forman, R.T.T.; Sperling, D.; Bissonette, J.A.; Clevenger, A.P.; Cutshall, C.D.; Dale, V.H.; Fahrig, L.; France, R.; Goldman, C.R.; Heanue, K.; et al. Road Ecology: Science and Solutions; Island Press: Washington, DC, USA, 2002. [Google Scholar]
- Delgado, J.; Arroyo, N.; Arévalo, J.R.; Fernández-Palacios, J.M. Edge effects of roads on temperature, light, canopy cover, and canopy height in laurel and pine forests (Tenerife, Canary Islands). Landsc. Urban Plan. 2007, 81, 328–340. [Google Scholar] [CrossRef]
- Haider, S.; Kueffer, C.; Bruelheide, H.; Seipel, T.; Alexander, J.M.; Rew, L.J.; Arévalo, J.R.; Cavieres, L.A.; McDougall, K.L.; Milbau, A.; et al. Mountain roads and non-native species modify elevational patterns of plant diversity. Glob. Ecol. Biogeogr. 2018, 27, 667–678. [Google Scholar] [CrossRef]
- Bacaro, G.; Maccherini, S.; Chiarucci, A.; Jentsch, A.; Rocchini, D.; Torri, D.; Gioria, M.; Tordoni, E.; Martellos, S.; Altobelli, A.; et al. Distributional patterns of endemic, native and alien species along a roadside elevation gradient in Tenerife, Canary Islands. Community Ecol. 2015, 16, 223–234. [Google Scholar] [CrossRef]
- Tarvirdizadeh, H.; Nikooy, M.; Pourbabaei, H.; Naghdi, R. Effects of road construction on biodiversity and composition of herbaceous species cover, Asalem forest, Northern Iran. For. Ideas 2014, 20, 157–169. [Google Scholar]
- Cavieres, L.A.; Quiroz, C.L.; Molina-Montenegro, M.A.; Muñoz, A.A.; Pauchard, A. Nurse effect of the native cushion plant Azorella monantha on the invasive non-native Taraxacum officinale in the high-Andes of central chile. Perspect. Plant Ecol. Evol. Syst. 2005, 7, 217–226. [Google Scholar] [CrossRef]
- Whittaker, R.J. Island Biogeography: Ecology, Evolution, and Conservation; Oxford University Press: Oxford, UK, 1998. [Google Scholar]
- Fernández-Palacios, J.; Arévalo, J.R.; Delgado, J.; Otto, R. Canarias: Ecología, Medio Ambiente y Desarrollo, 1st ed.; Gobierno de Canarias: Consejería de Política Territorial y Medio Ambiente; Centro de la Cultura Popular de Canarias, La Laguna: Santa Cruz de Tenerife, Spain, 2004. [Google Scholar]
- Fernández-Palacios, J.; Martín Esquivel, J. Las islas como experimento de laboratorio. In Naturaleza de las Islas Canarias. Ecología y Conservación; Chapter: 1; Fernández-Palacios, J., Martín, J.L., Eds.; Turquesa: San Cristóbal de La Laguna, Spain, 2001. [Google Scholar] [CrossRef]
- Arteaga, M.A.; Delgado, J.D.; Otto, R.; Fernández-Palacios, J.M.; Arévalo, J.R. How do alien plants distribute along roads on oceanic islands? A case study in Tenerife, Canary Islands. Biol. Invasions 2009, 11, 1071–1086. [Google Scholar] [CrossRef]
- Pauchard, A.; Kueffer, C.; Dietz, H.; Daehler, C.; Alexander, J.; Edwards, P.; Arévalo, J.R.; Cavieres, L.; Guisan, A.; Haider, S.; et al. Ain’t no mountain high enough: Plant invasions reaching new elevations. Front. Ecol. Environ. 2009, 7, 479–486. [Google Scholar] [CrossRef]
- Arévalo, J.R.; Delgado, J.; Fernández-Palacios, J. Changes in plant species composition and litter production in response to roads and trails in the laurel forest of Tenerife (Canary Islands). Plant Biosyst. 2008, 142, 614–622. [Google Scholar] [CrossRef]
- Flory, L.; Clay, K. Effects of roads and forest successional age on experimental plant invasions. Biol. Conserv. 2009, 142, 2531–2537. [Google Scholar] [CrossRef]
- Cohen, S.; Groner, E.; Peeters, A.; Segoli, M. The Impact of Roads on the Redistribution of Plants and Associated Arthropods in a Hyper-Arid Ecosystem. J. Insect. Sci. 2021, 21, 4, PMCID:PMC8289131. [Google Scholar] [CrossRef] [PubMed]
- Organismo Autónomo Parques Nacionales. Seguimiento de Datos Socioeconómicos en las Áreas de Influencia Socioeconómica. Parque Nacional de Timanfaya. Informe de Resultados Para el Periodo 2017–2019; Ministerio para la Transición Ecológica y el Reto Demográfico: Madrid, Spain, 2019; p. 29.
- Scott, M.; Helfman, G. Native Invasions, Homogenization, and the Mismeasure of Integrity of Fish Assemblages. Fisheries 2001, 26, 6–15. [Google Scholar] [CrossRef]
- Zamani, M.; Nikooy, M.; Tavankar, F. Effects of forest roads on species composition of trees regeneration in selection cutting management of high forests. Forestist 2020, 70, 151–159. [Google Scholar] [CrossRef]
- Otto, R.; Arteaga, M.A.; Delgado, J.; Fernández-Palacios, J.M.; Arévalo, J.R. Road edge effect and elevation patterns of native and alien plants on an oceanic island (Tenerife, Canary Islands). Folia Geobot. 2014, 49, 65–82. [Google Scholar] [CrossRef]
- Vakhlamova, T.; Rusterholz, H.P.; Kanibolotskaya, Y.; Baur, B. Effects of road type and urbanization on the diversity and abundance of alien species in roadside verges in Western Siberia. Plant Ecol. 2016, 217, 241–252. [Google Scholar] [CrossRef]
- Sandoya, V.; Pauchard, A.; Cavieres, L. Natives and non-natives plants show different responses to elevation and disturbance on the tropical high Andes of Ecuador. Ecol. Evol. 2017, 7, 7909–7919. [Google Scholar] [CrossRef] [PubMed]
- Morente-Lopez, J.; Arjona, Y.; Salas-Pascual, M.; Reyes-Betancort, J.; Arco-Aguilar, M.; Emerson, B.; García-Gallo, A.; Jay-García, L.; Naranjo-Cigala, A.; Patiño, J. Biogeographic origins and drivers of alien plant invasions in the Canary Islands. J. Biogeogr. 2023, 50, 576–590. [Google Scholar] [CrossRef]
- Arechavaleta, M.; Rodríguez, S.; Zurita, N.; García, A. Lista de Especies Silvestres de Canarias. Hongos, Plantas y Animales Terrestres; Gobierno de Canarias: Santa Cruz de Tenerife, Spain, 2010; pp. 122–172.
- González-Carracedo, M.A.; Hernández-Ferrer, M.N.; Cabrera-Pérez, R.M.; Bernardos, M.; Pérez-Pérez, J.A. Phylogeographic analysis points toward invasion of the Timanfaya National Park (Lanzarote; Canary Islands) by a translocated native plant (Rumex lunaria). Biol. Invasions. In prep.
- Centellas, A. Guía de la Flora del Parque Nacional de Timanfaya; Organismo Autónomo de Parques Nacionales: Madrid, Spain, 2009; 226p.
- Cruz-Trujillo, G.M. Flora Vascular del Parque Nacional de Timanfaya (Lanzarote, Islas Canarias); Serie Técnica; Ministerio de Medio Ambiente, Organismo Autónomo Parques Nacionales, Naturaleza y Parques Nacionales: Madrid, Spain, 2005; p. 207.
- Wildpret-De-La-Torre, W.; Beltrán-Tejera, E.; González-Mancebo, J.M.; Centellas-Bodas, A. Pelargonium capitatum y Rumex lunaria, dos plantas invasoras en el Parque Nacional de Timanfaya (Lanzarote, Islas Canarias). Consideraciones ecológicas y fitosociológicas. Anuario Inst. Est. Canar. 1995, 39, 9–16. [Google Scholar]
- Instituto Geológico y Minero de España (IGME); Cartográfica de Canarias (GRAFCAN). Descripción de las Unidades Geológicas de Lanzarote; Mapa Geológico de Canarias; Gobierno de Canarias, CARTOGRAF, FEDER, Programa MAC 2007–2013, 1992–1994; Instituto Geológico y Minero de España: Madrid, Spain, 2015. [Google Scholar]
- Mayer, P.; Luque, A.; García-Hernández, F. Atlas Climático Interactivo de Canarias de alta resolución espacial. In Fuentes de Datos, Metodología y Resultados; Grupo de Geografía Física y Medio Ambiente de la ULPGC, Grafcan, SA y Consejería de Transición Ecológica Lucha contra el Cambio Climático y Planificación Territorial del Gobierno autónomo de Canarias: Barcelona, Spain, 2021. [Google Scholar]
- Marzol, M.V. Las Direcciones de los Vientos Dominantes en Canarias; Centro Meteorológico de Canarias, Instituto Nacional de Meteorología: Madrid, Spain, 1985; p. 29. [Google Scholar]
- Van der Maarel, E. Transformation of Cover-Abundance Values in Phytosociology and its Effects on Community Similarity. Plant Ecol. 1979, 39, 97–114. [Google Scholar] [CrossRef]
- R Core Team. R: A language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 23 May 2023).
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.; Wagner, H. Vegan: Community Ecology Package; R Package (Version 2.5–7). 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 23 May 2023).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Halekoh, U.; Højsgaard, S. A kenward-roger approximation and parametric bootstrap methods for tests in linear mixed models—The R package pbkrtest. J. Stat. Softw. 2014, 59, 1–30. Available online: http://www.jstatsoft.org/v59/i09/ (accessed on 25 July 2023). [CrossRef]
- Gobierno de Canarias. Banco de Datos de Biodiversidad de Canarias. Available online: https://www.biodiversidadcanarias.es/biota/ (accessed on 30 April 2023).
- Gil, J.; Peña, M. Guía Visual de la Flora Vascular de la Reserva de la Biosfera de Lanzarote; Oficina de la Reserva de la Biosfera del Cabildo de Lanzarote: Madrid, Spain, 2018; p. 583. [Google Scholar]
- Reyes-Betancort, J.A. Las hijas de Lanzarote. Un paseo por su flora endémica. In Lanzarote: Naturaleza Entre Volcanes; Afonso-Carrillo, J., Ed.; Actas X Semana Científica Telesforo Bravo, Instituto de Estudios Hispánicos de Canarias: Puerto de la Cruz, Spain, 2015; pp. 61–85. ISBN 978-84-608-1557-0. [Google Scholar]







| Model | AIC | p | |
|---|---|---|---|
| (a) | Log(richness) = position + zone + season | 3634.177 | 0.000999 *** |
| Log(richness) = position × zone × season | 3538.346 | 0.000999 *** | |
| Log(richness) = zone | 4533.766 | 0.000999 *** | |
| Log(richness) = position | 4554.275 | 0.000999 *** | |
| Log(richness) = season | 3942.127 | 0.000999 *** | |
| (b) | Log(richness) = position + zone + season | 533.1070 | 0.000999 *** |
| Log(richness) = position × zone × season | 541.8778 | 0.000999 *** | |
| Log(richness) = zone | 1065.6377 | 0.016983 * | |
| Log(richness) = position | 905.8038 | 0.000999 *** | |
| Log(richness) = season | 848.4138 | 0.000999 *** | |
| (c) | Log(richness) = position + zone + season | 2149.202 | 0.000999 *** |
| Log(richness) = position × zone × season | 1778.268 | 0.000999 *** | |
| Log(richness) = zone | 3108.800 | 0.000999 *** | |
| Log(richness) = position | 3024.786 | 0.000999 *** | |
| Log(richness) = season | 2515.595 | 0.000999 *** |
| Species Richness | ||||
|---|---|---|---|---|
| Season | ||||
| Zones | Position | Summer | Winter | |
| (a) | Altos Timanfaya | Border | 2.1 ± 1.0 aA | 6.1 ± 3.8 aB |
| Middle | 1.3 ± 0.5 aA | 5.4 ± 4.0 aB | ||
| Interior | 1.5 ± 0.8 aA | 5.3 ± 4.4 aB | ||
| Calderas Quemadas | Border | 1.8 ± 0.8 aA | 2.8 ± 1.5 aB | |
| Middle | 1.0 ± 0.0 aA | 2.3 ± 1.3 bA | ||
| Interior | 1.0 ± 0.0 aA | 1.7 ± 0.8 bA | ||
| Caldereta Riscada | Border | 2.9 ± 1.0 aA | 6.9 ± 2.5 aB | |
| Middle | 2.0 ± 1.3 aA | 5.3 ± 2.2 abB | ||
| Interior | 1.3 ± 0.5 aA | 4.4 ± 2.3 bA | ||
| Islote | Border | 6.2 ± 2.1 aA | 28.5 ± 6.1 aB | |
| Middle | 0.9 ± 1.5 bA | 14.1 ± 10.2 bB | ||
| Interior | 1.0 ± 2.2 bA | 11.3 ± 8.3 bB | ||
| Manto Virgen | Border | 5.3 ± 2.3 aA | 18.6 ± 5.6 aA | |
| Middle | 0.5 ± 0.5 aA | 5.0 ± 3.2 aA | ||
| Interior | 0.8 ± 0.9 aA | 3.2 ± 1.5 aA | ||
| Mazo | Border | 5.0 ± 3.2 aA | 15.5 ± 4.7 aB | |
| Middle | 2.0 ± 1.0 aA | 5.5 ± 3.7 bB | ||
| Interior | 1.4 ± 0.5 aA | 3.1 ± 2.0 bA | ||
| Montaña Rajada | Border | 1.6 ± 0.9 aA | 2.8 ± 1.4 aB | |
| Middle | 1.3 ± 0.6 aA | 1.9 ± 1.6 aA | ||
| Interior | 1.0 ± 0.0 aA | 2.3 ± 2.0 aA | ||
| Taro | Border | 2.0 ± 1.4 aA | 7.8 ± 4.9 aB | |
| Middle | 1.8 ± 1.2 aA | 3.4 ± 3.0 bB | ||
| Interior | 1.8 ± 1.5 aA | 2.6 ± 2.7 bB | ||
| Tremesana | Border | 1.4 ± 0.6 aA | 3.6 ± 2.7 aB | |
| Middle | 1.2 ± 0.4 aA | 2.2 ± 1.4 bB | ||
| Interior | 1.1 ± 0.4 aA | 2.9 ± 2.1 bA | ||
| Valle Tranquilidad | Border | 5.5 ± 4.9 aA | 16.1 ± 13.5 aB | |
| Middle | 2.0 ± 1.3 aA | 6.5 ± 4.7 aB | ||
| Interior | 1.8 ± 0.9 aA | 4.5 ± 3.4 aB | ||
| (b) | Yaiza | Border | 3.5 ± 2.4 aA | 14.8 ± 7.1 aB |
| Middle | 0.9 ± 1.1 bA | 8.8 ± 9.3 bB | ||
| Interior | 1.0 ± 1.3 bA | 3.8 ± 6.2 bB | ||
| Echadero Camellos | Border | 5.3 ± 2.3 aA | 18.6 ± 5.6 aB | |
| Middle | 0.5 ± 0.5 bA | 5.0 ± 3.2 bB | ||
| Interior | 0.8 ± 0.9 bA | 3.2 ± 1.5 bB | ||
| Taro 1 | Border | 7.8 ± 1.3 aA | 29.8 ± 3.6 aB | |
| Middle | 2.0 ± 1.6 bA | 9.8 ± 4.3 bB | ||
| Interior | 2.4 ± 2.1 bA | 8.8 ± 8.2 bB | ||
| Centro Visitantes | Border | 6.2 ± 2.1 aA | 28.5 ± 6.1 aB | |
| Middle | 0.9 ± 1.5 bA | 14.1 ± 10.2 bB | ||
| Interior | 1.0 ± 2.2 bA | 11.3 ± 8.3 bB | ||
| Zone | No. Transects | Elevation (m a.s.l.) | Mean Slope (°Sex.) | Exposure | Mean Annual Temperature (°C) [48] | Mean Annual Precipitation (mm) [48] | Road Type | No. Years Sampled |
|---|---|---|---|---|---|---|---|---|
| Altos Timanfaya | 7 | 430.4 ± 13.7 | 11.9 ± 5.8 | N | 18.3 ± 0.1 | 174.0 ± 2.8 | Road | 3 |
| Calderas Quemadas | 6 | 293.2 ± 20.6 | 11.0 ± 4.9 | SE | 18.9 ± 0.1 | 157.1 ± 2.4 | Road | 3 |
| Caldereta Riscada | 4 | 356.3 ± 25.6 | 11.4 ± 5.3 | SO | 18.7 ± 0.1 | 159.8 ± 2.3 | Road | 3 |
| Islote | 8 | 317.2 ± 11.5 | 10.4 ± 4.4 | N | 18.9 ± 0.0 | 156.9 ± 1.7 | Road | 3 |
| Manto Virgen | 6 | 288.2 ± 18.0 | 9.2 ± 4.5 | N | 19.0 ± 0.1 | 155.5 ± 1.3 | Road | 3 |
| Mazo | 8 | 263.8 ± 25.9 | 2.7 ± 1.6 | SO | 19.0 ± 0.1 | 158.8 ± 4.2 | Trail | 3 |
| Montaña Rajada | 8 | 278.6 ± 33.0 | 13.2 ± 9.5 | N | 19.0 ± 0.1 | 157.4 ± 4.0 | Road | 3 |
| Taro | 6 | 300.9 ± 4.3 | 3.2 ± 2.0 | N | 18.9 ± 0.0 | 158.5 ± 1.7 | Road | 3 |
| Tremesana | 8 | 196.0 ± 10.3 | 4.3 ± 2.4 | E | 19.4 ± 0.1 | 148.8 ± 0.6 | Trail | 3 |
| Valle Tranquilidad | 6 | 341.6 ± 26.4 | 9.3 ± 3.7 | N | 18.7 ± 0.1 | 163.0 ± 3.2 | Road | 3 |
| Yaiza | 11 | 225.7 ± 25.0 | 4.0 ± 1.6 | E | 19.3 ± 0.1 | 154.0 ± 2.2 | Road | 1 |
| Echadero Camellos | 8 | 333.2 ± 20.7 | 5.4 ± 2.9 | E | 18.8 ± 0.1 | 166.6 ± 1.6 | Road | 1 |
| Taro 1 | 5 | 307.8 ± 17.5 | 6.5 ± 7.3 | N | 18.8 ± 0.1 | 161.8 ± 1.8 | Road | 1 |
| Centro Visitantes | 10 | 294.9 ± 12.4 | 3.1 ± 2.6 | N | 18.7 ± 0.0 | 167.3 ± 2.4 | Road | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernardos, M.; Cornejo, N.S.; Torres Hassan, A.D.; Cabrera, R.; Arévalo, J.R. Road Impact on Plant Colonization in the Arid Timanfaya National Park. Plants 2023, 12, 3568. https://doi.org/10.3390/plants12203568
Bernardos M, Cornejo NS, Torres Hassan AD, Cabrera R, Arévalo JR. Road Impact on Plant Colonization in the Arid Timanfaya National Park. Plants. 2023; 12(20):3568. https://doi.org/10.3390/plants12203568
Chicago/Turabian StyleBernardos, María, Natalia Sierra Cornejo, Antonio Daniel Torres Hassan, Raimundo Cabrera, and José Ramón Arévalo. 2023. "Road Impact on Plant Colonization in the Arid Timanfaya National Park" Plants 12, no. 20: 3568. https://doi.org/10.3390/plants12203568
APA StyleBernardos, M., Cornejo, N. S., Torres Hassan, A. D., Cabrera, R., & Arévalo, J. R. (2023). Road Impact on Plant Colonization in the Arid Timanfaya National Park. Plants, 12(20), 3568. https://doi.org/10.3390/plants12203568