
Dense Cover, but Not Allelopathic Potential, of Naturalized Alien Cenchrus echinatus L. Threatens the Native Species in Urban Vegetation

 and
and
Abstract
:1. Introduction
2. Results
2.1. Floristic Composition
2.2. Effect of Cenchrus echinatus L. on Vegetation
2.3. Effect of C. echinatus on Soil Properties
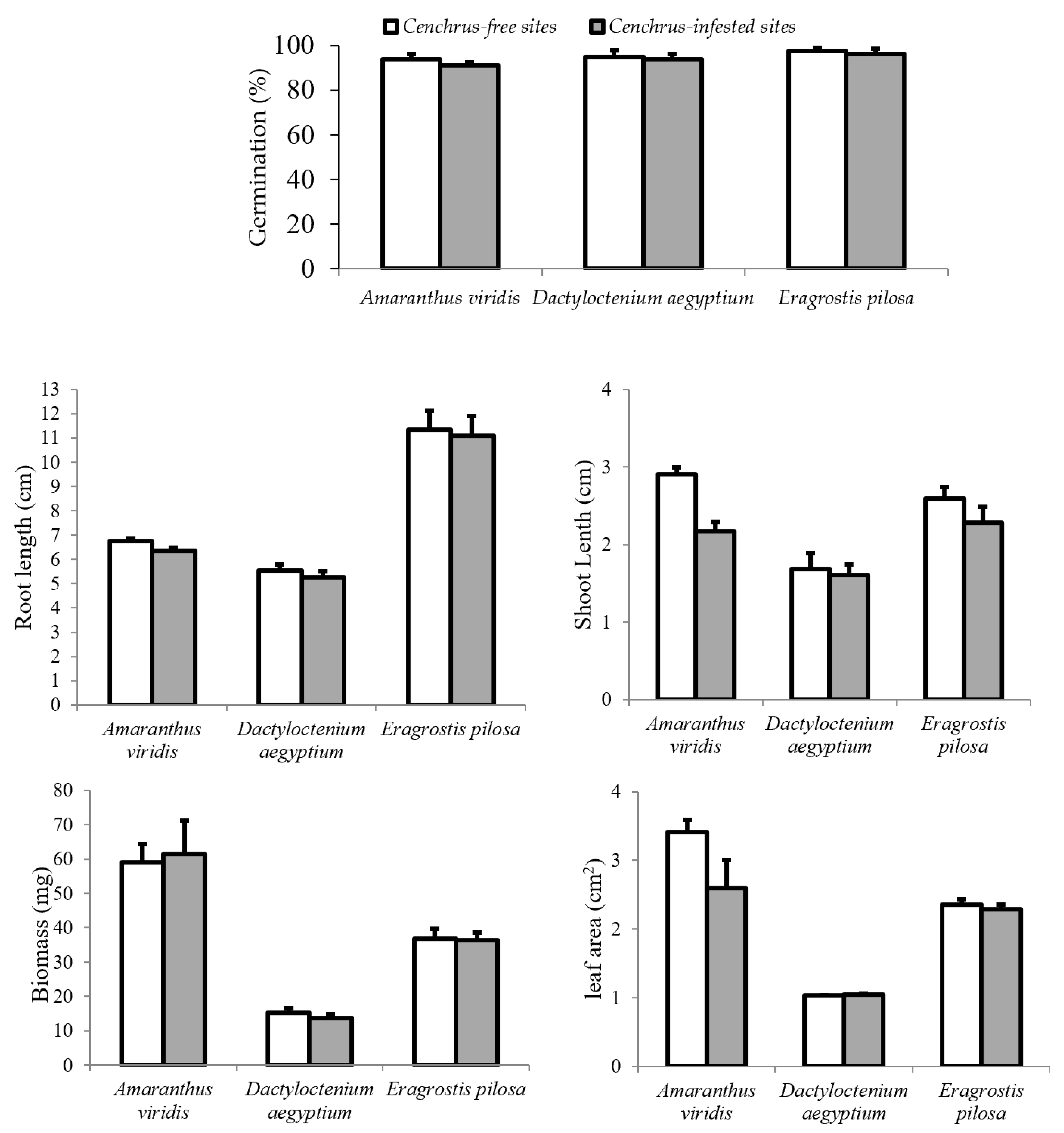
2.4. Allelopathic Potential of C. echinatus
3. Discussion
4. Materials and Methods
4.1. Vegetation Survey
4.2. Soil Seed Bank Sampling
4.3. Soil Analysis
4.4. Allelopathic Potential of C. echinatus
4.4.1. Soil Sampling and Preparation
4.4.2. The Target Species
4.4.3. Test for Allelopathy
4.5. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Castro-Díez, P.; Alonso, Á. Effects of non-native riparian plants in riparian and fluvial ecosystems: A review for the Iberian Peninsula. Limnetica 2017, 36, 525–541. [Google Scholar] [CrossRef]
- Gaertner, M.; Den Breeyen, A.; Hui, C.; Richardson, D.M. Impacts of alien plant invasions on species richness in Mediterranean-type ecosystems: A meta-analysis. Prog. Phys. Geogr. 2009, 33, 319–338. [Google Scholar] [CrossRef]
- Hassan, M.O.; Mohamed, H.Y. Allelopathic interference of the exotic naturalized Paspalum dilatatum Poir. threatens diversity of native plants in urban gardens. Flora 2020, 266, 151593. [Google Scholar] [CrossRef]
- Qu, T.; Du, X.; Peng, Y.; Guo, W.; Zhao, C.; Losapio, G. Invasive species allelopathy decreases plant growth and soil microbial activity. PLoS ONE 2021, 16, e0246685. [Google Scholar] [CrossRef] [PubMed]
- Vilà, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarošík, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pyšek, P. Ecological impacts of invasive alien plants: A meta-analysis of their effects on species, communities and ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef]
- El-Beheiry, M.; Hosni, H.; Sharaf El-din, A.; Shaltout, S.K.; Ahmed, D. Updating the checklist of the alien flora in Egypt. Taeckholmia 2020, 40, 41–56. [Google Scholar] [CrossRef]
- Pyšek, P.; Richardson, D.M. Invasive species, environmental change and management, and health. Annu. Rev. Environ. Resour. 2010, 35, 25–55. [Google Scholar] [CrossRef]
- Raizada, P.; Raghubanshi, A.S.; Singh, J.S. Impact of invasive alien plant species on soil processes: A review. Proc. Nat. Acad. Sci. India Sect. B 2008, 78, 288–298. [Google Scholar]
- Zhang, Z.; van Kleunen, M. Common alien plants are more competitive than rare natives but not than common natives. Ecol. Lett. 2019, 22, 1378–1386. [Google Scholar] [CrossRef]
- Yuan, L.; Li, J.M.; Yu, F.H.; Oduor, A.M.; van Kleunen, M. Allelopathic and competitive interactions between native and alien plants. Biol. Invasions 2021, 23, 3077–3090. [Google Scholar] [CrossRef]
- Van Kleunen, M.; Weber, E.; Fischer, M. A meta-analysis of trait differences between invasive and non-invasive plant species. Ecol. Lett. 2010, 13, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Pyšek, P.; Richardson, D.M. Traits associated with invasiveness in alien plants: Where do we stand? In Biological Invasisons; Nentwig, W., Ed.; Springer: Berlin/Heidelberg, Germany, 2007; Volume 193, pp. 97–125. [Google Scholar]
- Callaway, R.M.; Ridenour, W.M.; Laboski, T.; Weir, T.; Vivanco, J.M. Natural selection for resistance to the allelopathic effects of invasive plants. J. Ecol. 2005, 93, 576–583. [Google Scholar] [CrossRef]
- Callaway, R.M.; Ridenour, W.M. Novel weapons: Invasive success and the evolution of increased competitive ability. Front. Ecol. Environ. 2004, 2, 436–443. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P.; Rejmanek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and invasion of alien plants: Concepts and definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Gaertner, M.; Wilson, J.R.U.; Cadotte, M.W.; MacIvor, J.S.; Zenni, R.D.; Richardson, D.M. Non-native species in urban environments: Patterns, processes, impacts and challenges. Biol. Invasions 2017, 19, 3461–3469. [Google Scholar] [CrossRef]
- Williams, N.S.; Hahs, A.K.; Vesk, P.A. Urbanisation, plant traits and the composition of urban floras. Perspect. Plant Ecol. Evol. Syst. 2015, 17, 78–86. [Google Scholar] [CrossRef]
- Malavasi, M.; Carboni, M.; Cutini, M.; Carranza, M.L.; Acosta, A.T. Landscape fragmentation, land-use legacy and propagule pressure promote plant invasion on coastal dunes: A patch-based approach. Landsc. Ecol. 2014, 29, 1541–1550. [Google Scholar] [CrossRef]
- Deutschewitz, K.; Lausch, A.; Kühn, I.; Klotz, S. Native and alien plant species richness in relation to spatial heterogeneity on a regional scale in Germany. Glob. Ecol. Biogeogr. 2003, 12, 299–311. [Google Scholar] [CrossRef]
- Sukopp, H. Human-caused impact on preserved vegetation. Landsc. Urban Plan. 2004, 68, 347–355. [Google Scholar] [CrossRef]
- Štajerová, K.; Šmilauer, P.; Brůna, J.; Pyšek, P. Distribution of invasive plants in urban environment is strongly spatially structured. Landsc. Ecol. 2017, 32, 681–692. [Google Scholar] [CrossRef]
- Dalle Fratte, M.; Bolpagni, R.; Brusa, G.; Caccianiga, M.; Pierce, S.; Zanzottera, M.; Cerabolini, B.E. Alien plant species invade by occupying similar functional spaces to native species. Flora 2019, 257, 151419. [Google Scholar] [CrossRef]
- Lazzaro, L.; Bolpagni, R.; Buffa, G.; Gentili, R.; Lonati, M.; Stinca, A.; Acosta, A.T.R.; Adorni, M.; Aleffi, M.; Allegrezza, M.; et al. Impact of invasive alien plants on native plant communities and Natura 2000 habitats: State of the art, gap analysis and perspectives in Italy. J. Environ. Manag. 2020, 274, 111140. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.O. Leaf litter of Bombax ceiba L. threatens plant cover and floristic diversity in a new urban ecosystem. Flora 2018, 242, 22–30. [Google Scholar] [CrossRef]
- Hassan, M.O.; Hassan, Y.M. Effect of human activities on floristic composition and diversity of desert and urban vegetation in a new urbanized desert ecosystem. Heliyon 2019, 5, e02283. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.O.; Mohamed, H.Y.; Aboellil, A.H. Allelopathic potential of Ficus retusa L. leaf litter on understory vegetation in urban gardens. Acta Bot. Croat. 2021, 80, 131–139. [Google Scholar] [CrossRef]
- Shehata, H.S. Ecology and allelopathic control of the invasive species Cenchrus echinatus L. in some of newly reclaimed areas in Nile Delta, Egypt. Life Sci. J. 2014, 11, 246–260. [Google Scholar]
- Shaltout, K.H.; Hosni, H.A.; El-Kady, H.F.; El-Beheiry, M.A.; Shaltout, S.K. Composition and pattern of alien species in the Egyptian flora. Flora Morphol. Distrib. Funct. Ecol. Plants 2016, 222, 104–110. [Google Scholar] [CrossRef]
- Nascimento, E.A.; Terrones, M.G.; Morais, S.A.; Chang, R.; Andrade, G.A.; Santos, D.Q.; Pereira, B.H. Allelopathic activity of Cenchrus echinatus L. extracts on weeds and crops. Allelopath. J. 2009, 24, 363–372. [Google Scholar]
- Ma, W.; Miao, S.; Tao, W.; Xu, Z.; Wang, H.; Chen, J.; Huang, L. Study on the Allelopathic Effects of Alien Invasive Species Cenchrus echinatus on Seed Germination and Seedling Growth of Solanaceae Crops. Agric. Sci. Technol. 2014, 15, 885–889. [Google Scholar]
- Favaretto, A.; Scheffer-Basso, S.M.; Perez, N.B. Allelopathy in Poaceae species present in Brazil. A review. Agron. Sustain. Dev. 2018, 38, 22. [Google Scholar] [CrossRef]
- Da Silva Maia, S.; Ribeiro-Rocha, P.R.; Santiago-Castro, T.; Costa da Silva, I.K.E.; Ferreira-Barreto, G.; Torres de Souza, L.; Murga-Orrillo, H.; Abanto-Rodríguez, C. Interference of Cenchrus echinatus and Rottboellia exaltata in growth of cowpea crop. Bioagro 2021, 33, 21–28. [Google Scholar] [CrossRef]
- Tielbörger, K.; Prasse, R. Do seeds sense each other? Testing for density-dependent germination in desert perennial plants. Oikos 2009, 118, 792–800. [Google Scholar] [CrossRef]
- Grundy, A.C.; Mead, A.; Burston, S. Modelling the germination response of weed seeds to burial depth: Interactions with seed density, weight and shape. J. Appl. Ecol. 2003, 40, 757–770. [Google Scholar] [CrossRef]
- Hassan, M.O.; Gomaa, N.H.; Fahmy, G.M.; González, L.; Hammouda, O.; Atteya, A.M. Interactions between Sonchus oleraceus L. and some weeds in agroecosystems in Egypt. Ann. Agric. Sci. 2014, 59, 221–228. [Google Scholar] [CrossRef]
- Foy, C.L. Inderjit Understanding the role of allelopathy in weed interference and declining plant diversity. Weed Technol. 2001, 15, 873–878. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Allelopathy and allelochemicals of Imperata cylindrica as an invasive plant species. Plants 2022, 11, 2551. [Google Scholar] [CrossRef]
- Wade, G.L. Grass competition and establishment of native species from forest soil seed banks. Landsc. Urban Plan. 1989, 17, 135–149. [Google Scholar] [CrossRef]
- Anju, M.V.; Warrier, R.R.; Kunhikannan, C. Significance of Soil Seed Bank in Forest Vegetation—A Review. Seeds 2022, 1, 181–197. [Google Scholar] [CrossRef]
- Morgan, J.B.; Connolly, E.L. Plant-Soil Interactions: Nutrient Uptake. Nat. Educ. Knowl. 2013, 4, 2. [Google Scholar]
- Furey, G.N.; Tilman, D. Plant biodiversity and the regeneration of soil fertility. Proc. Natl. Acad. Sci. USA 2021, 118, e2111321118. [Google Scholar] [CrossRef]
- Tilman, D.; Wedin, D.; Knops, J. Productivity and sustainability influenced by biodiversity in grassland ecosystems. Nature 1996, 379, 718–720. [Google Scholar] [CrossRef]
- Tecco, P.A.; Díaz, S.; Cabido, M.; Urcelay, C. Functional traits of alien plants across contrasting climatic and land-use regimes: Do aliens join the locals or try harder than them? J. Ecol. 2010, 98, 17–27. [Google Scholar] [CrossRef]
- Vilà, M.; Weiner, J. Are invasive plant species better competitors than native plant species? Evidence from pair-wise experiments. Oikos 2004, 105, 229–238. [Google Scholar] [CrossRef]
- Boulos, L. Azollaceae–Oxalidace. In Flora of Egypt; Al Hadara Publishing: Cairo, Egypt, 1999; Volume 1. [Google Scholar]
- Boulos, L. Geraniaceae–Boraginaceae. In Flora of Egypt; Al Hadara Publishing: Cairo, Egypt, 2000; Volume 2. [Google Scholar]
- Boulos, L. Verbenaceae–Compositae. In Flora of Egypt; Al Hadara Publishing: Cairo, Egypt, 2002; Volume 3. [Google Scholar]
- Boulos, L. Monocotyledons: Alismataceae–Orchidaceae. In Flora of Egypt; Al Hadara Publishing: Cairo, Egypt, 2005; Volume 4. [Google Scholar]
- Pielou, E.C. Ecological Diversity; Wiley: London, UK, 1975. [Google Scholar]
- Zhang, J.T. Quantitative Methods in Vegetation Ecology; China Science and Technology Press: Beijing, China, 1995. [Google Scholar]
- Gutterman, Y. Seed Germination in Desert Plants, 1st ed.; Springer: Berlin/Heidelberg, Germany, 1993. [Google Scholar]
- Bouyoucos, G.J. Hydrometer method improved for making particle size analysis of soils. Agron. J. 1962, 54, 464–465. [Google Scholar] [CrossRef]
- Jackson, M.L. Soil Chemical Analysis-Advanced Course; Washington Department of Soil Sciences: Washington, DC, USA, 1967. [Google Scholar]
- Walkley, A.; Black, I.A. An examination of Degtjareff method for determining soil organic matter and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Allen, S.E. Chemical Analysis of Ecological Materials, 2nd ed.; Blackwell Scientific Publishers: Oxford, UK, 1989. [Google Scholar]


{kind=link}
{kind=link}
| Species | Family | Cenchrus-Free Sites | Cenchrus-Infested Sites |
|---|---|---|---|
| Amaranthus viridis L. † | Amaranthaceae | 39.82 ± 4.23 | 15.10 ** ± 4.41 |
| Apium leptophyllym Pers. † | Apiaceae | 15.60 ± 3.48 | 3.9 ** ± 0.45 |
| Cynodon dactylon Pers. †† | Poaceae | 142.21 ± 13.25 | 97.38 ** ± 9.46 |
| Dactyloctenium aegyptium (L.) Willd. † | Poaceae | 3.30 ± 0.64 | 0.0 ** ± 0.0 |
| Dichanthium annulatum (Forssk.) Stapf †† | Poaceae | 19.28 ± 4.48 | 8.50 ± 3.86 |
| Digitaria sanguinalis (L.) Scop. † | Poaceae | 16.38 ± 3.18 | 11.90 ± 3.27 |
| Eragrostis pilosa (L.) P.Beauv. † | Poaceae | 49.74 ± 8.42 | 14.81 ** ± 3.50 |
| Euphorbia peplus L. † | Euphorbiaceae | 4.35 ± 0.60 | 0.35 ** ± 0.26 |
| Oxalis corniculata L. †† | Oxalidaceae | 17.11 ± 6.79 | 9.44 ± 2.33 |
| Plantago lagopus L. † | Plantaginaceae | 17.00 ± 2.54 | 3.63 ** ± 2.63 |
| Diversity Indices | Cover of C. echinatus |
|---|---|
| Species richness | −0.54 ** |
| Shannon index | −0.70 ** |
| Evenness | −0.35 * |
| Simpson index | −0.71 ** |
| Species | Life Span | Cenchrus-Free Sites | Cenchrus-Infested Sites |
|---|---|---|---|
| Amaranthus viridis L. | Annual | - | 9.72 ** ± 1.01 b |
| Cenchrus echinatus L. | Annual | - | 2.73 ** ± 0.33 a |
| Dactyloctenium aegyptium (L.) Willd. | Annual | - | 14.80 ** ± 0.90 c |
| Euphorbia peplus L. | Annual | - | 9.64 ** ± 0.98 b |
| Soil Properties | Cenchrus-Free Sites | Cenchrus-Infested Sites |
|---|---|---|
| Sand (%) | 48.39 ± 2.15 | 49.00 ± 1.60 |
| Silt (%) | 32.62 ±1.58 | 32.02 ± 1.50 |
| Clay (%) | 18.99 ± 0.93 | 18.78 ± 0.70 |
| pH | 7.98 ± 0.03 | 7.92 ± 0.03 |
| EC (µS cm−1) | 377.54 ± 15.50 | 377.08 ± 10.50 |
| CaCO3 (%) | 2.47 ± 0.09 | 2.56 ± 0.07 |
| OC (%) | 1.97 ± 0.07 | 2.05 ± 0.05 |
| Available nutrients (mg kg−1 soil) | ||
| N | 67.2 ± 1.50 | 51.12 ** ± 3.90 |
| P | 12.55 ± 0.42 | 10.50 ** ± 0.45 |
| K | 411.02 ± 5.70 | 361.15 ** ± 6.20 |
| Zn | 1.60 ± 0.13 | 6.10 ** ± 0.31 |
| Cu | 2.28 ± 0.13 | 2.45 ± 0.14 |
| Climatic Parameters | Jan. | Feb. | Mar. | Apr. | May. | Jun. | Jul. | Aug. | Sep. | Oct. | Nov. | Dec. |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Maximum temperature (°C) | 29 | 28 | 36 | 41 | 44 | 42 | 43 | 44 | 40 | 36 | 35 | 24 |
| Minimum temperature (°C) | 3 | 2 | 5 | 5 | 16 | 16 | 20 | 21 | 18 | 14 | 11 | 3 |
| Relative humidity (%) | 52.25 | 52.19 | 49.5 | 36.75 | 27.31 | 33.38 | 32.62 | 33.19 | 43.75 | 48.38 | 52.31 | 62.19 |
| Precipitation (mm) | 0 | 0 | 10.54 | 0.30 | 0 | 0 | 0 | 0 | 0 | 0.30 | 4.50 | 9.3 |
| Wind speed (km h−1) | 31.36 | 35.96 | 35.64 | 33.62 | 33.48 | 25.45 | 24.70 | 28.73 | 34.34 | 27.32 | 31.42 | 35.57 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassan, M.O.; Mohamed, H.Y.; Okla, M.K.; Kiani, B.H.; Amro, A. Dense Cover, but Not Allelopathic Potential, of Naturalized Alien Cenchrus echinatus L. Threatens the Native Species in Urban Vegetation. Plants 2023, 12, 3736. https://doi.org/10.3390/plants12213736
Hassan MO, Mohamed HY, Okla MK, Kiani BH, Amro A. Dense Cover, but Not Allelopathic Potential, of Naturalized Alien Cenchrus echinatus L. Threatens the Native Species in Urban Vegetation. Plants. 2023; 12(21):3736. https://doi.org/10.3390/plants12213736
Chicago/Turabian StyleHassan, Mahmoud O., Howida Y. Mohamed, Mohammad K. Okla, Bushra Hafeez Kiani, and Ahmed Amro. 2023. "Dense Cover, but Not Allelopathic Potential, of Naturalized Alien Cenchrus echinatus L. Threatens the Native Species in Urban Vegetation" Plants 12, no. 21: 3736. https://doi.org/10.3390/plants12213736
APA StyleHassan, M. O., Mohamed, H. Y., Okla, M. K., Kiani, B. H., & Amro, A. (2023). Dense Cover, but Not Allelopathic Potential, of Naturalized Alien Cenchrus echinatus L. Threatens the Native Species in Urban Vegetation. Plants, 12(21), 3736. https://doi.org/10.3390/plants12213736