
The Developmental Mechanism of the Root System of Cultivated Terrestrial Watercress

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
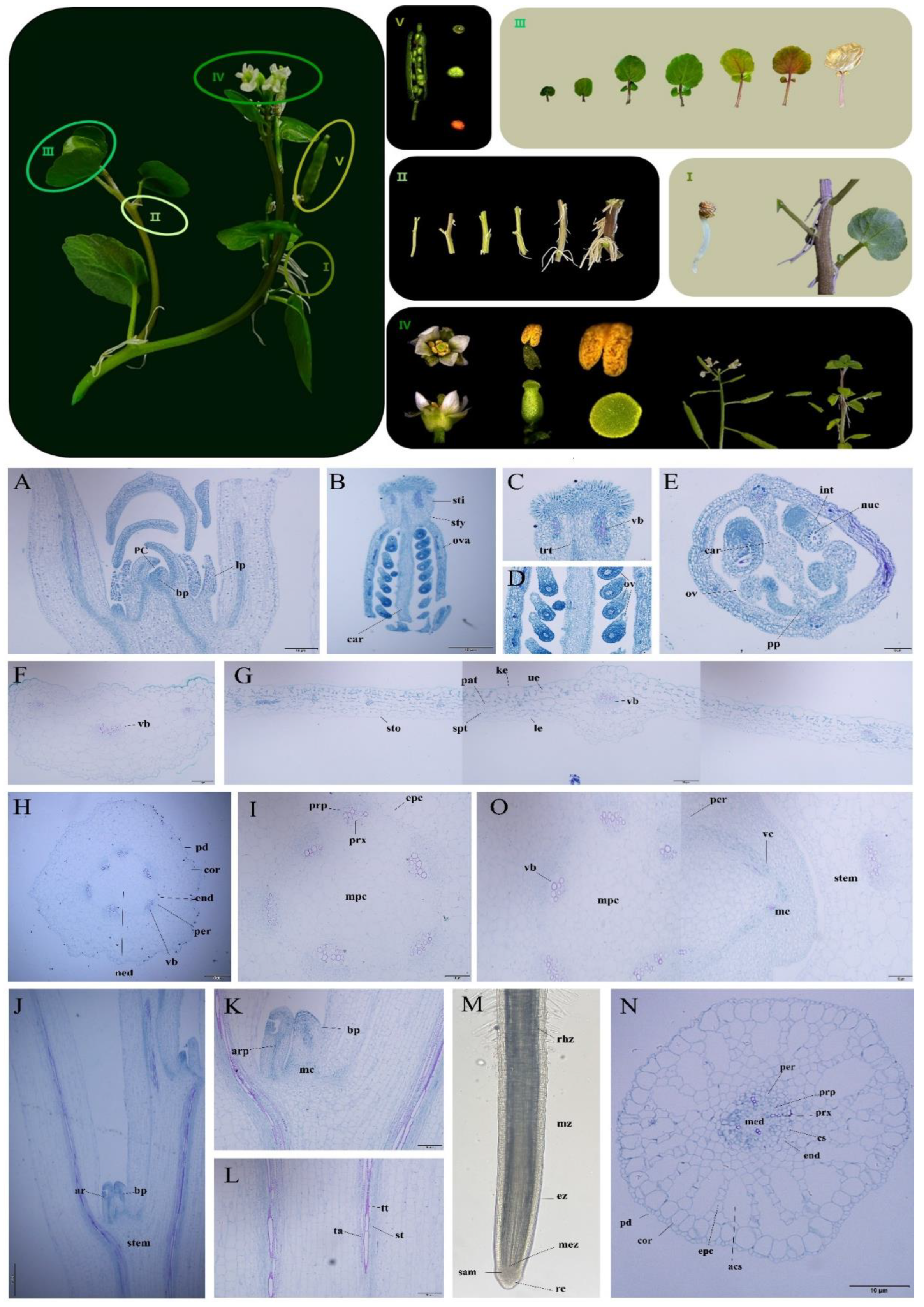
2.1. Botanical Characteristics and Tissue Slices of Watercress
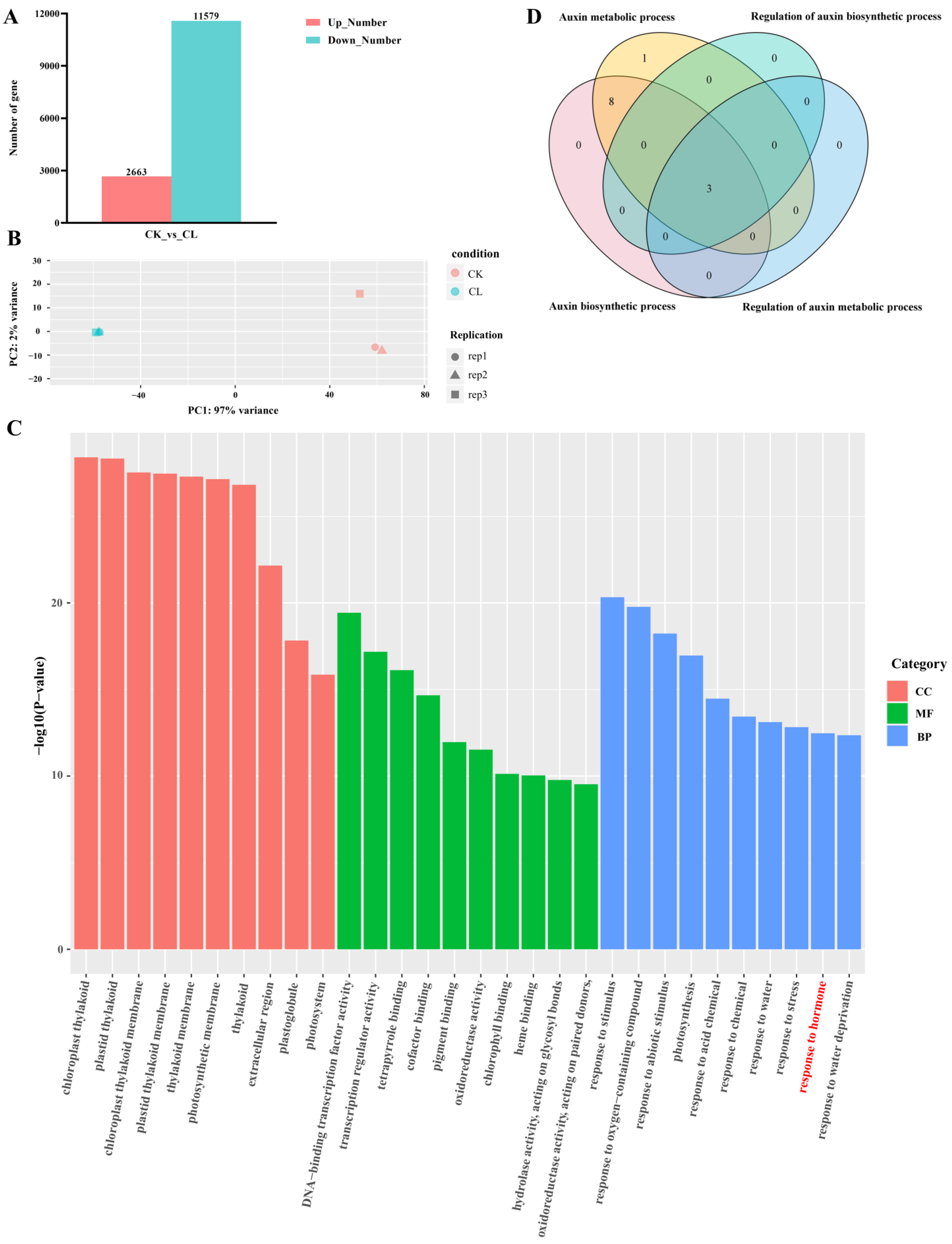
2.2. Transcriptome Analysis of Root Tissue in Watercress
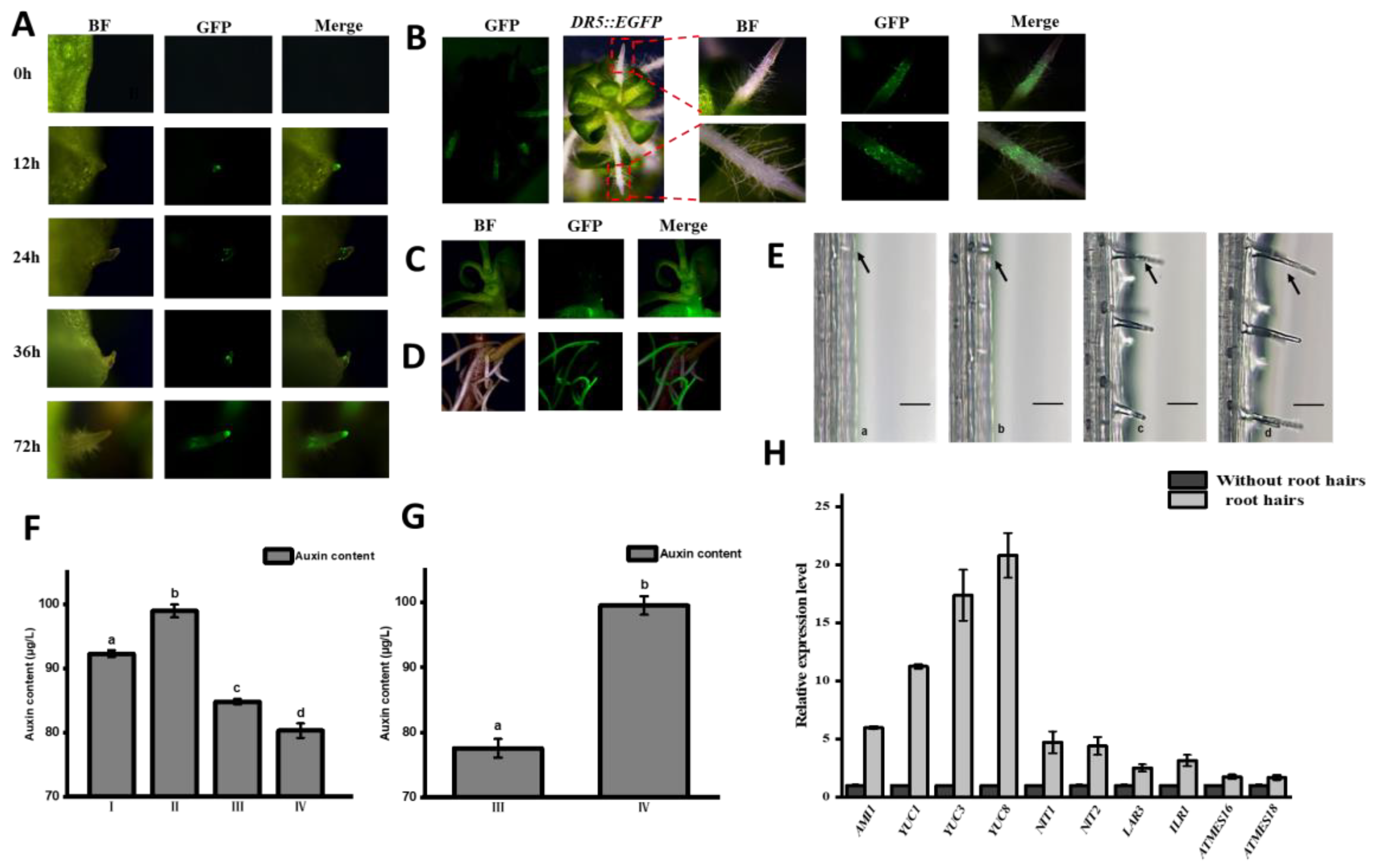
2.3. Dynamic Distribution of Auxin in Root Development of Watercress
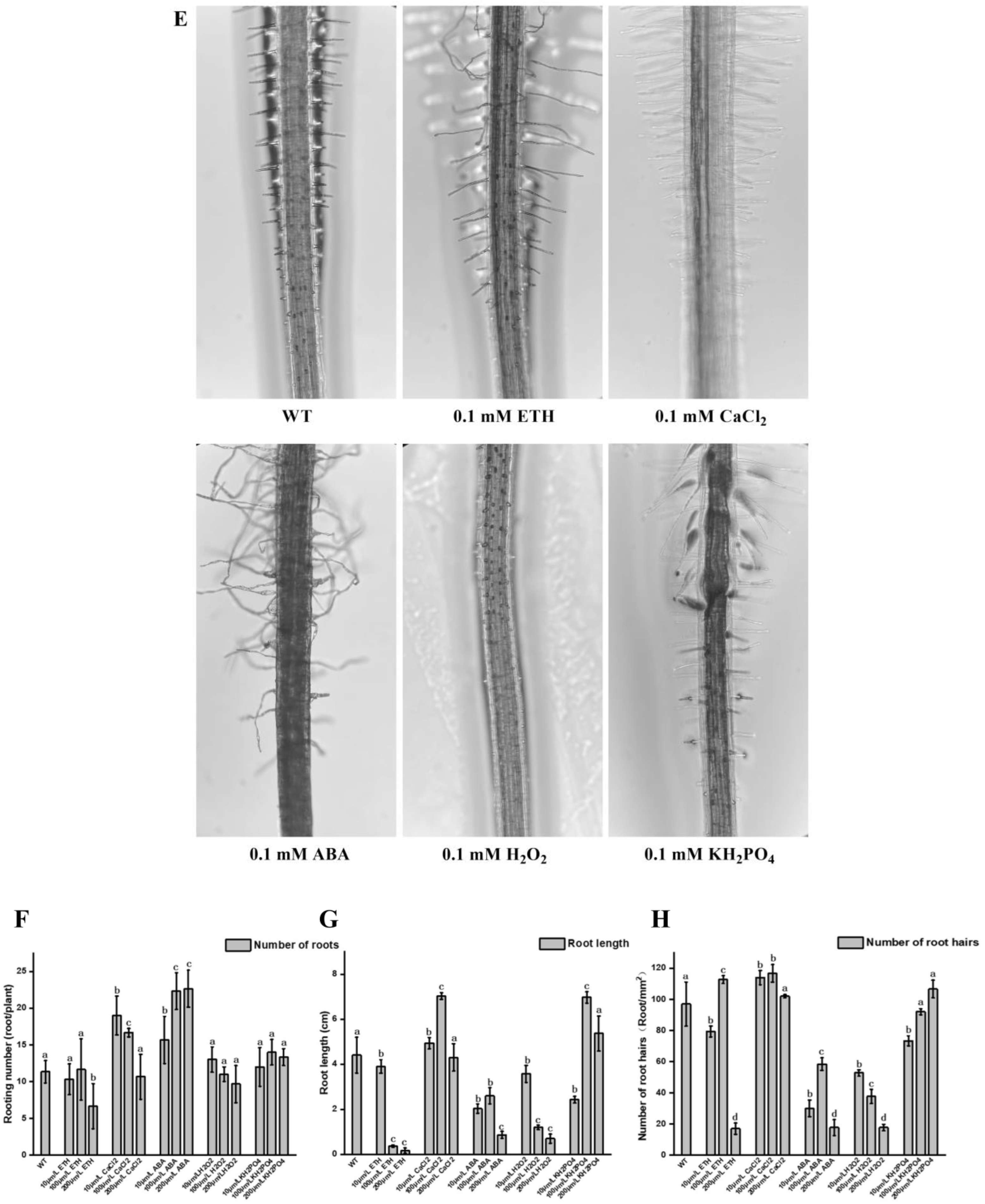
2.4. Other Important Factors Affecting the Occurrence of Root Hairs in Watercress
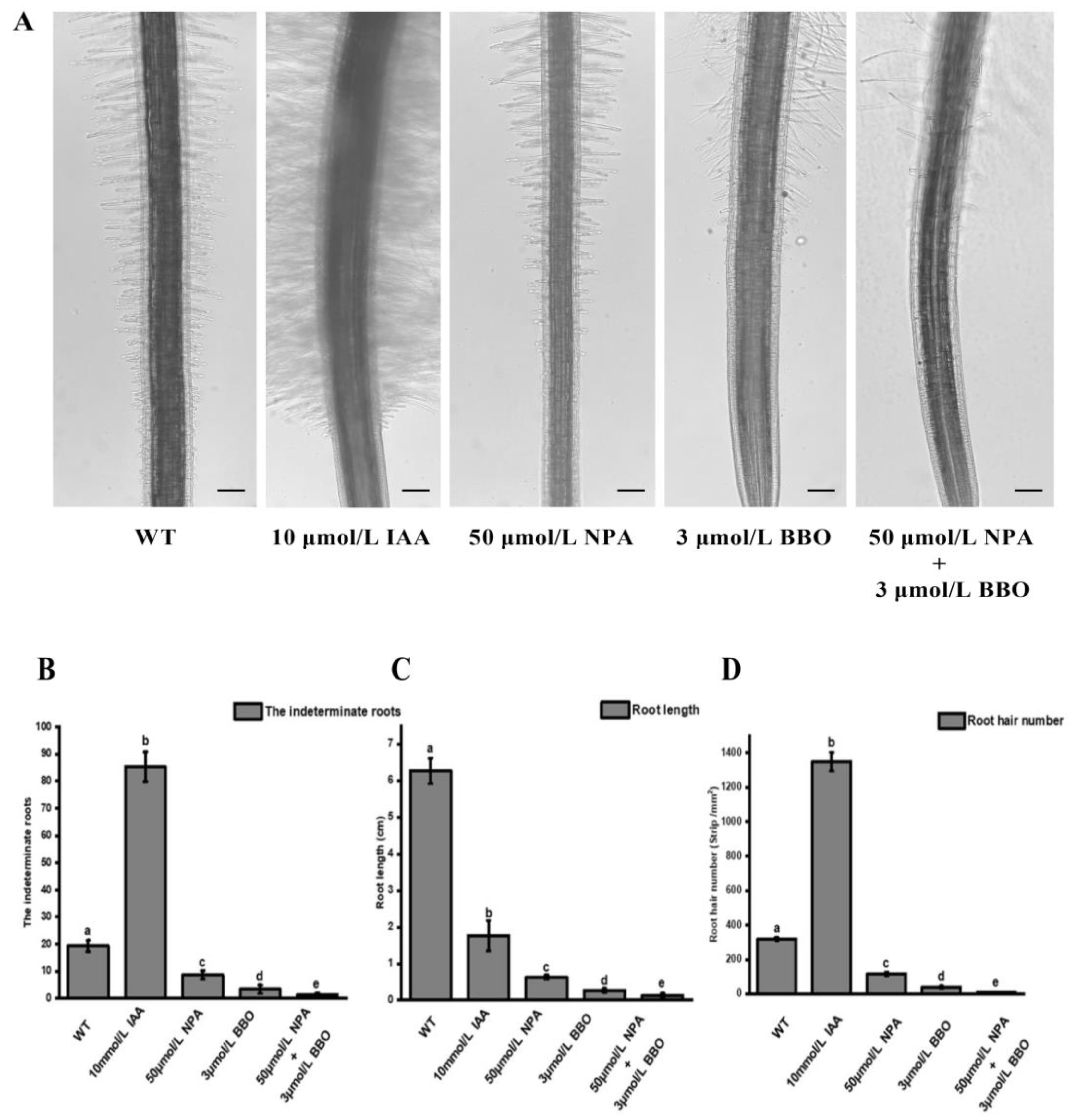
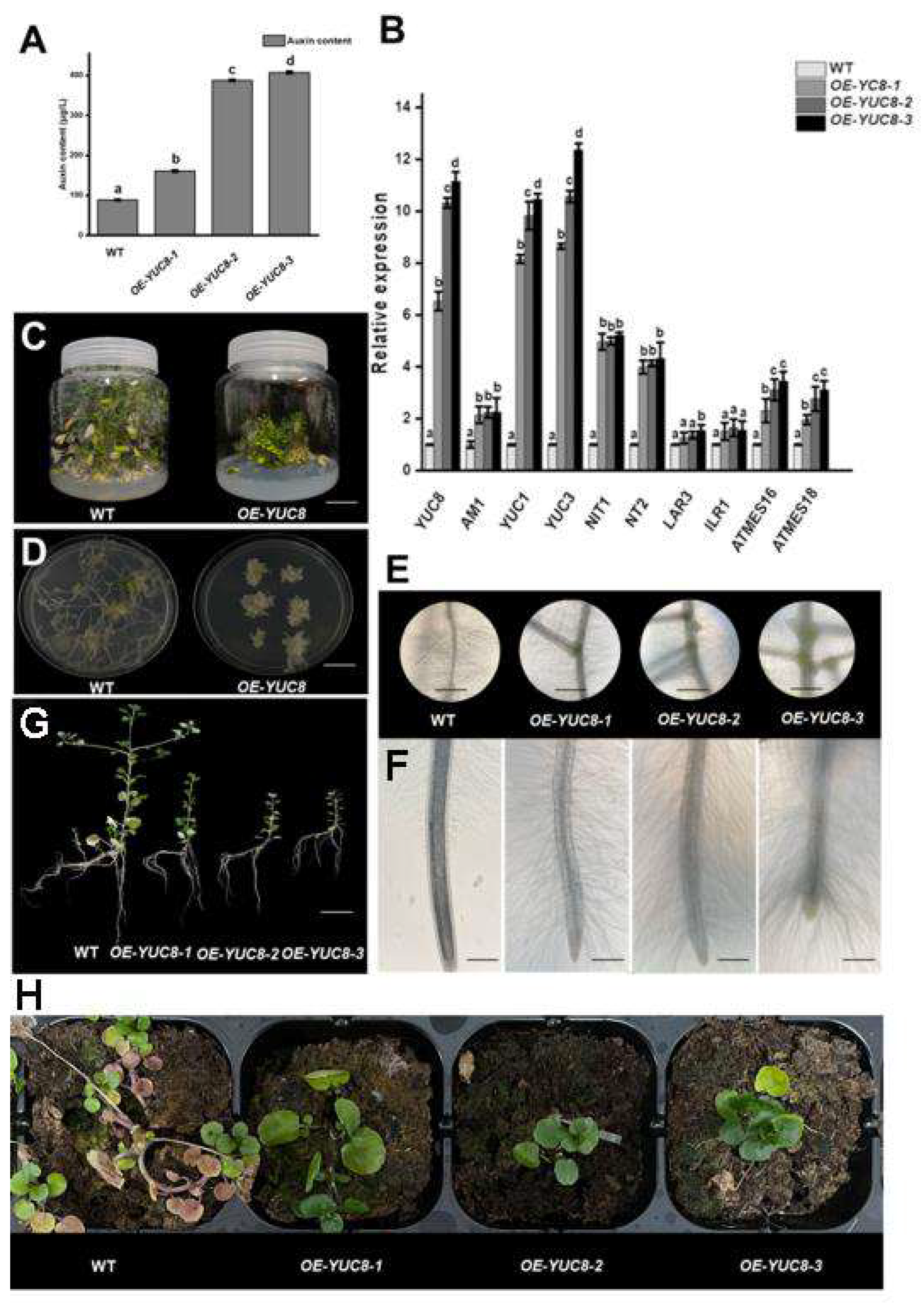
2.5. Regulation of NoYUC8 on Root Development in Watercress
3. Discussion
4. Materials and Methods
4.1. Plant Materials, Growth Conditions, and Treatments
4.2. Paraffin Sectioning of Watercress
4.3. Auxin Determination
4.4. Vectors and Identification of Transgenic Watercress
4.5. Measurement of Root Length and Root Hair Number in Watercress
4.6. Genetic Transformation of Watercress
4.7. Transcriptome Sequencing of Watercress Adventitious Roots at Different Stages
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Batianoff, G.N.; Butler, D.W. Assessment of invasive naturalized plants in south-east Queensland. Plant Prot. Q. 2002, 17, 27–34. [Google Scholar]
- Faizy, H.S.; Esmail, L.S.; Mahdi, H.S. Phytochemicals analysis in watercress (Nasturtium officinale) plant extracts. IOP Conf. Ser. Earth Environ. Sci. 2021, 761, 12042. [Google Scholar] [CrossRef]
- Bayrami, A.; Ghorbani, E.; Pouran, S.R.; Habibi-Yangjeh, A.; Khataee, A.; Bayrami, M. Enriched zinc oxide nanoparticles by Nasturtium officinale leaf extract: Joint ultrasound-microwave-facilitated synthesis, characterization, and implementation for diabetes control and bacterial inhibition. Ultrason. Sonochem. 2019, 58, 104613. [Google Scholar] [CrossRef] [PubMed]
- Adlravan, E.; Jalilzadeh-Razin, S.; Nejati, K.; Karimi, M.A.; Mousazadeh, H.; Abbasi, A.; Dadashpour, M. Potential activity of free and PLGA/PEG nanoencapsulated Nasturtium officinale extract in inducing cytotoxicity and apoptosis in human lung carcinoma A549 cells. J. Drug Deliv. Sci. Technol. 2021, 61, 102256. [Google Scholar] [CrossRef]
- Ramezani, S.; Javadi, I.; Kokhdan, E.P.; Omidifar, N.; Nikbakht, J.; Sadeghi, H.; Doustimotlagh, A.H.; Danaei, N.; Abbasi, R.; Sadeghi, H. Protective and therapeutic effects of ethanolic extract of Nasturtium officinale (watercress) and vitamin E against bleomycin-induced pulmonary fibrosis in rats. Res. Pharm. Sci. 2021, 16, 94. [Google Scholar]
- Schulze, H.; Hornbacher, J.; Wasserfurth, P.; Reichel, T.; Günther, T.; Krings, U.; Krüger, K.; Hahn, A.; Papenbrock, J.; Schuchardt, J.P. Immunomodulating Effect of the Consumption of Watercress (Nasturtium officinale) on Exercise-Induced Inflammation in Humans. Foods 2021, 10, 1774. [Google Scholar] [CrossRef] [PubMed]
- Vissenberg, K.; Claeijs, N.; Balcerowicz, D.; Schoenaers, S. Hormonal regulation of root hair growth and responses to the environment in Arabidopsis. J. Exp. Bot. 2020, 71, 2412–2427. [Google Scholar] [CrossRef]
- Datta, S.; Kim, C.M.; Pernas, M.; Pire, N.D.; Proust, H.; Tam, T.; Vijayakumar, P.; Dolan, L. Root hairs: Development, growth and evolution at the plant-soil interface. Plant Soil 2011, 346, 1–14. [Google Scholar] [CrossRef]
- Hua, T.; Tai, N.T.; Liem, P.T.; Ut, V.N.; Ako, H. Effects of different stocking densities on growth performance of asian swamp eel monopterus albus, water quality and plant growth of watercress Nasturtium officinale in an aquaponic recirculating system. Aquaculture 2019, 503, 96–104. [Google Scholar]
- Thang, N.Q.; Tho, N.T.; Phuong, N.T. Nitrate, nitrite, and lead contamination in leafy vegetables collected from local market sites of go vap district, Ho Chi Minh City. Vietnam. J. Chem. 2021, 59, 79–86. [Google Scholar]
- Tang, W.J.; Xiao, L.; Peng, X.; Liu, H.; Zhu, Y.; Zheng, Y. Effects of brassinolide on cadmium accumulation and growth of emerged accumulator plant Nasturtium officinale. Chem. Ecol. 2022, 38, 301–311. [Google Scholar] [CrossRef]
- Xue, C.; Li, W.; Shen, R.; Lan, P. Impacts of iron on phosphate starvation-induced root hair growth in Arabidopsis. Plant Cell Environ. 2023, 46, 215–238. [Google Scholar] [CrossRef] [PubMed]
- Biswas, M.S.; Fukaki, H.; Mori, I.C.; Nakahara, K.; Mano, J. Reactive oxygen species and reactive carbonyl species constitute a feed-forward loop in auxin signaling for lateral root formation. Plant J. 2019, 100, 536–548. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.E.; Marzol, E.; Estevez, J.M.; Muday, G.K. Ethylene signaling increases reactive oxygen species accumulation to drive root hair initiation in Arabidopsis. Development 2022, 149, dev200487. [Google Scholar] [CrossRef] [PubMed]
- Bahmani, R.; Kim, D.G.; Modareszadeh, M.; Hwang, S. Cadmium enhances root hair elongation through reactive oxygen species in Arabidopsis. Environ. Exp. Bot. 2022, 196, 104813. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, F.; Ding, Y.; Du, H.; Zhang, Q.; Dang, X.; Cao, Y.; Dodd, I.C.; Xu, W. Abscisic acid mediates barley rhizosheath formation under mild soil drying by promoting root hair growth and auxin response. Plant Cell Environ. 2021, 44, 1935–1945. [Google Scholar] [CrossRef] [PubMed]
- Tanimoto, M.; Roberts, K.; Dolan, L. Ethylene is a positive regulator of root hair development in Arabidopsis thaliana. Plant J. 1995, 8, 943–948. [Google Scholar] [CrossRef]
- Zhu, C.; Gan, L.; Shen, Z.; Xia, K. Interactions between jasmonates and ethylene in the regulation of root hair development in Arabidopsis. J. Exp. Bot. 2006, 57, 1299–1308. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, T.; Wang, R.; Zhao, Y. Recent advances in auxin research in rice and their implications for crop improvement. J. Exp. Bot. 2017, 69, 255–263. [Google Scholar] [CrossRef]
- Wang, T.; Li, C.; Wu, Z.; Jia, Y.; Wang, H.; Sun, S.; Mao, C.; Wang, X. Abscisic Acid Regulates Auxin Homeostasis in Rice Root Tips to Promote Root Hair Elongation. Front. Plant Sci. 2017, 8, 1121. [Google Scholar] [CrossRef]
- Zhao, Y. Auxin Biosynthesis: A Simple Two-Step Pathway Converts Tryptophan to Indole-3-Acetic Acid in Plants. Mol. Plant. 2012, 5, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Christensen, S.K.; Fankhauser, C.; Cashman, J.R.; Cohen, J.D.; Weigel, D.; Chory, J. A Role for Flavin Monooxygenase-Like Enzymes in Auxin Biosynthesis. Science 2001, 291, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Mashiguchi, K.; Tanaka, K.; Sakai, T.; Sugawara, S.; Kawaide, H.; Natsume, M.; Hanada, A.; Yaeno, T.; Shirasu, K.; Yao, H. The main auxin biosynthesis pathway in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 18512–18517. [Google Scholar] [CrossRef] [PubMed]
- Won, C.; Shen, X.; Mashiguchi, K.; Zheng, Z.; Dai, X.; Cheng, Y.; Kasahara, H.; Kamiya, Y.; Chory, J.; Zhao, Y. Conversion of tryptophan to indole-3-acetic acid by Tryptophan Aminotransferases of Arabidopsis and Yuccas in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 18518–18523. [Google Scholar] [CrossRef] [PubMed]
- Goh, T.; Kasahara, H.; Mimura, T.; Kamiya, Y.; Fukaki, H. Multiple AUX/IAA–ARF modules regulate lateral root formation: The role of Arabidopsis SHY2/IAA3-mediated auxin signalling. Phil. Trans. R. Soc. 2012, 367, 1461–1468. [Google Scholar] [CrossRef] [PubMed]
- Uehara, T.; Okushima, Y.; Mimura, T.; Tasaka, M.; Fukaki, H. Domain II Mutations in CRANE/IAA18 Suppress Lateral Root Formation and Affect Shoot Development in Arabidopsis thaliana. Plant Cell Physiol. 2008, 49, 1025–1038. [Google Scholar] [CrossRef] [PubMed]
- Fukaki, H.; Nakao, Y.; Okushima, Y.; Theologis, A.; Tasaka, M. Tissue-specific expression of stabilized SOLITARY-ROOT/IAA14 alters lateral root development in Arabidopsis. Plant J. 2005, 44, 382–395. [Google Scholar] [CrossRef] [PubMed]
- Tatematsu, K.; Kumagai, S.; Muto, H.; Sato, A.; Watahiki, M.K.; Harper, R.M.; Liscum, E.; Yamamoto, K.T. MASSUGU2 Encodes Aux/IAA19, an Auxin-Regulated Protein That Functions Together with the Transcriptional Activator NPH4/ARF7 to Regulate Differential Growth Responses of Hypocotyl and Formation of Lateral Roots in Arabidopsis thaliana. Plant Cell 2004, 16, 379–393. [Google Scholar] [CrossRef]
- Wan, Q.; Zhai, N.; Xie, D.; Liu, W.; Xu, L. WOX11: The founder of plant organ regeneration. Cell Regen. 2023, 12, 1. [Google Scholar] [CrossRef]
- Yang, E.; Yang, H.; Li, C.; Zheng, M.; Song, H.; Zou, X.; Chen, X.; Zhang, J. Genome-Wide Identification and Expression Analysis of the Aux/IAA Gene Family of the Drumstick Tree (Moringa oleifera Lam.) Reveals Regulatory Effects on Shoot Regeneration. Int. J. Mol. Sci. 2022, 23, 15729. [Google Scholar] [CrossRef]
- Zhang, S.; Yu, R.; Yu, D.; Chang, P.; Guo, S.; Yang, X.; Liu, X.; Xu, C.; Hu, Y. The calcium signaling module CaM–IQM destabilizes IAA–ARF interaction to regulate callus and lateral root formation. Proc. Natl. Acad. Sci. USA 2022, 119, e2202669119. [Google Scholar] [CrossRef] [PubMed]
- Dubrovsky, J.G.; Sauer, M.; Napsucialy-Mendivil, S.; Ivanchenko, M.G.; Friml, J.; Shishkova, S.; Celenza, J.; Benková, E. Auxin acts as a local morphogenetic trigger to specify lateral root founder cells. Proc. Natl. Acad. Sci. USA 2008, 105, 8790–8794. [Google Scholar] [CrossRef] [PubMed]
- Peculis, B.A.; Steitz, J.A. Sequence and structural elements critical for u8 snrnp function in xenopus oocytes are evolutionarily conserved. Genes Dev. 1994, 8, 2241–2255. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Dhonukshe, P.; Brewer, P.B.; Friml, J. Spatiotemporal asymmetric auxin distribution: A means to coordinate plant development. Cell. Mol. Life Sci. 2006, 63, 2738–2754. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.J.; Li, M.; Lv, M.; Hepworth, S.R.; Li, D.; Ma, C.; Li, J.; Wang, S. SAUR15 Promotes Lateral and Adventitious Root Development via Activating H+-ATPases and Auxin Biosynthesis. Plant Physiol. 2020, 184, 837–851. [Google Scholar] [CrossRef] [PubMed]
- Masucci, J.D.; Schiefelbein, J.W. The rhd6 mutation of arabidopsis thaliana alters root-hair initiation through an auxin- and ethylene-associated process. Plant Physiol. 1995, 106, 1335–1346. [Google Scholar] [CrossRef] [PubMed]
- Wada, T.; Tachibana, T.; Shimura, Y.; Okada, K. Epidermal cell differentiation in Arabidopsis determined by a Myb homolog, CPC. Science 1997, 277, 1113–1116. [Google Scholar] [CrossRef] [PubMed]
- Shibata, M.; Sugimoto, K. A gene regulatory network for root hair development. J. Plant Res. 2019, 132, 301–309. [Google Scholar] [CrossRef]
- Baluška, F.; Salaj, J.; Mathur, J.; Braun, M.; Jasper, F.; Samaj, J.; Chua, N.H.; Barlow, P.W.; Volkmann, D. Root Hair Formation: F-Actin-Dependent Tip Growth Is Initiated by Local Assembly of Profilin-Supported F-Actin Meshworks Accumulated within Expansin-Enriched Bulges. Dev. Biol. 2000, 227, 618–632. [Google Scholar] [CrossRef]
- Badri, D.V.; Vivanco, J.M. Regulation and function of root exudates. Plant Cell Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef]
- Yi, K.; Menand, B.; Bell, E.; Dolan, L. A basic helix-loop-helix transcription factor controls cell growth and size in root hairs. Nat. Genet. 2010, 42, 264–267. [Google Scholar] [CrossRef]
- Bruex, A.; Kainkaryam, R.M.; Wieckowski, Y.; Kang, Y.H.; Bernhardt, C.; Xia, Y.; Zheng, X.; Wang, J.Y.; Lee, M.M.; Benfey, P.; et al. A Gene Regulatory Network for Root Epidermis Cell Differentiation in Arabidopsis. PLoS Genet. 2012, 8, e1002446. [Google Scholar] [CrossRef]
- Desbrosses, G.; Josefsson, C.; Rigas, S.; Hatzopoulos, P.; Dolan, L. AKT1 and TRH1 are required during root hair elongation in Arabidopsis. J. Exp. Bot. 2003, 54, 781–788. [Google Scholar] [CrossRef]
- Monshausen, G.B.; Messerli, M.A.; Gilroy, S. Imaging of the Yellow Cameleon 3.6 Indicator Reveals That Elevations in Cytosolic Ca2+ Follow Oscillating Increases in Growth in Root Hairs of Arabidopsis. Plant Physiol. 2008, 147, 1690–1698. [Google Scholar] [CrossRef]
- Macpherson, N.; Takeda, S.; Shang, Z.; Dark, A.; Mortimer, J.C.; Brownlee, C.; Dolan, L.; Davies, J.M. NADPH oxidase involvement in cellular integrity. Planta 2008, 227, 1415–1418. [Google Scholar] [CrossRef]
- Qin, H.; Wang, J.; Zhou, J.; Qiao, J.; Li, Y.; Quan, R.; Huang, R. Abscisic acid promotes auxin biosynthesis to inhibit primary root elongation in rice. Plant Physiol. 2022, 191, 1953–1967. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ran, J.; Ding, Q.; Wang, G.; Shen, Y.; Gao, Z.; Gao, Y.; Ma, X.; Hou, X. The Developmental Mechanism of the Root System of Cultivated Terrestrial Watercress. Plants 2023, 12, 3523. https://doi.org/10.3390/plants12203523
Ran J, Ding Q, Wang G, Shen Y, Gao Z, Gao Y, Ma X, Hou X. The Developmental Mechanism of the Root System of Cultivated Terrestrial Watercress. Plants. 2023; 12(20):3523. https://doi.org/10.3390/plants12203523
Chicago/Turabian StyleRan, Jiajun, Qiang Ding, Guangpeng Wang, Yunlou Shen, Zhanyuan Gao, Yue Gao, Xiaoqing Ma, and Xilin Hou. 2023. "The Developmental Mechanism of the Root System of Cultivated Terrestrial Watercress" Plants 12, no. 20: 3523. https://doi.org/10.3390/plants12203523
APA StyleRan, J., Ding, Q., Wang, G., Shen, Y., Gao, Z., Gao, Y., Ma, X., & Hou, X. (2023). The Developmental Mechanism of the Root System of Cultivated Terrestrial Watercress. Plants, 12(20), 3523. https://doi.org/10.3390/plants12203523