
Effect of the Nonpathogenic Strain Fusarium oxysporum FO12 on Fe Acquisition in Rice (Oryza sativa L.) Plants

 ,
,  , ,
, ,  , ,
, , 
Abstract
:1. Introduction
2. Results
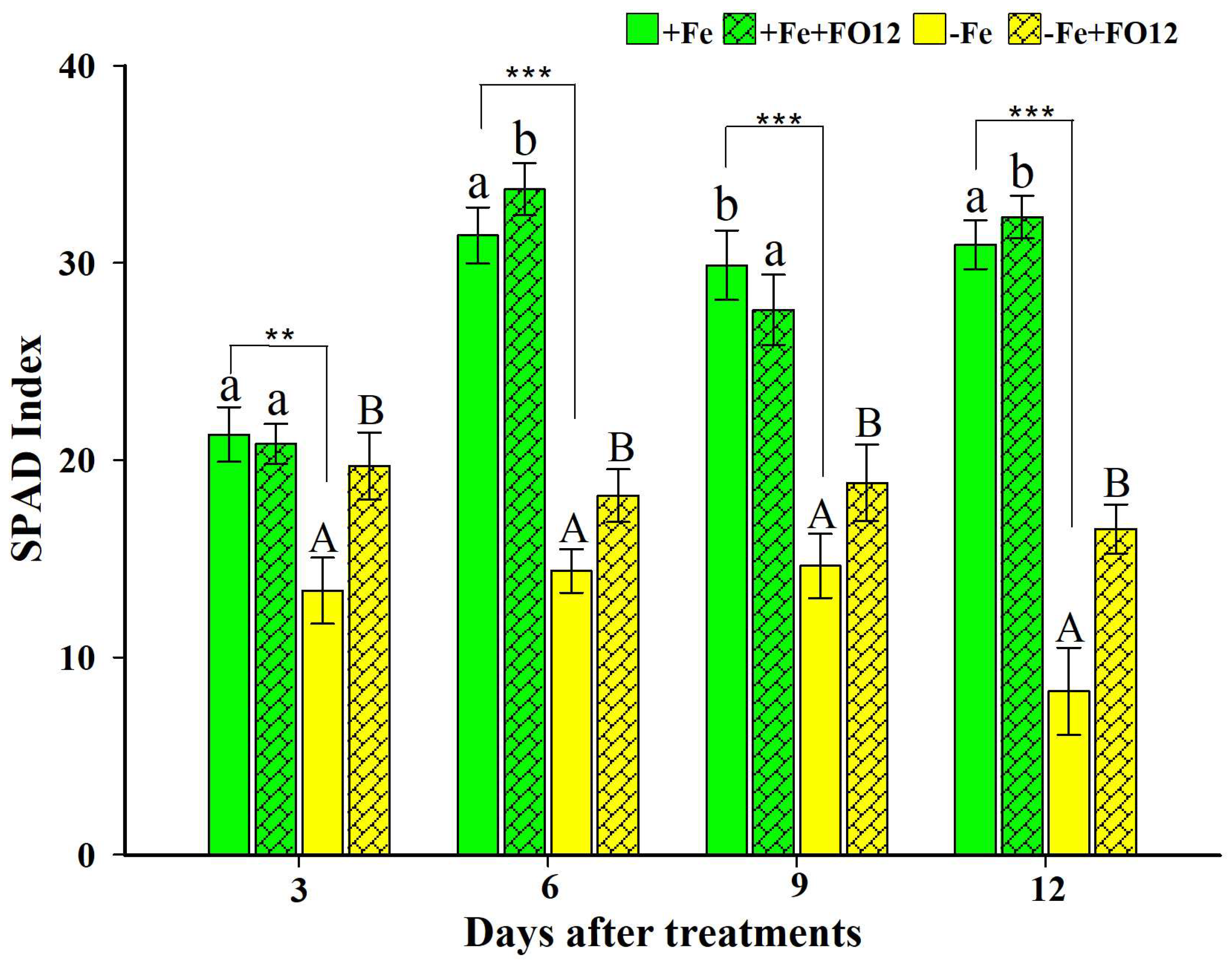
2.1. Effect of the Inoculation with FO12 on Fe Chlorosis
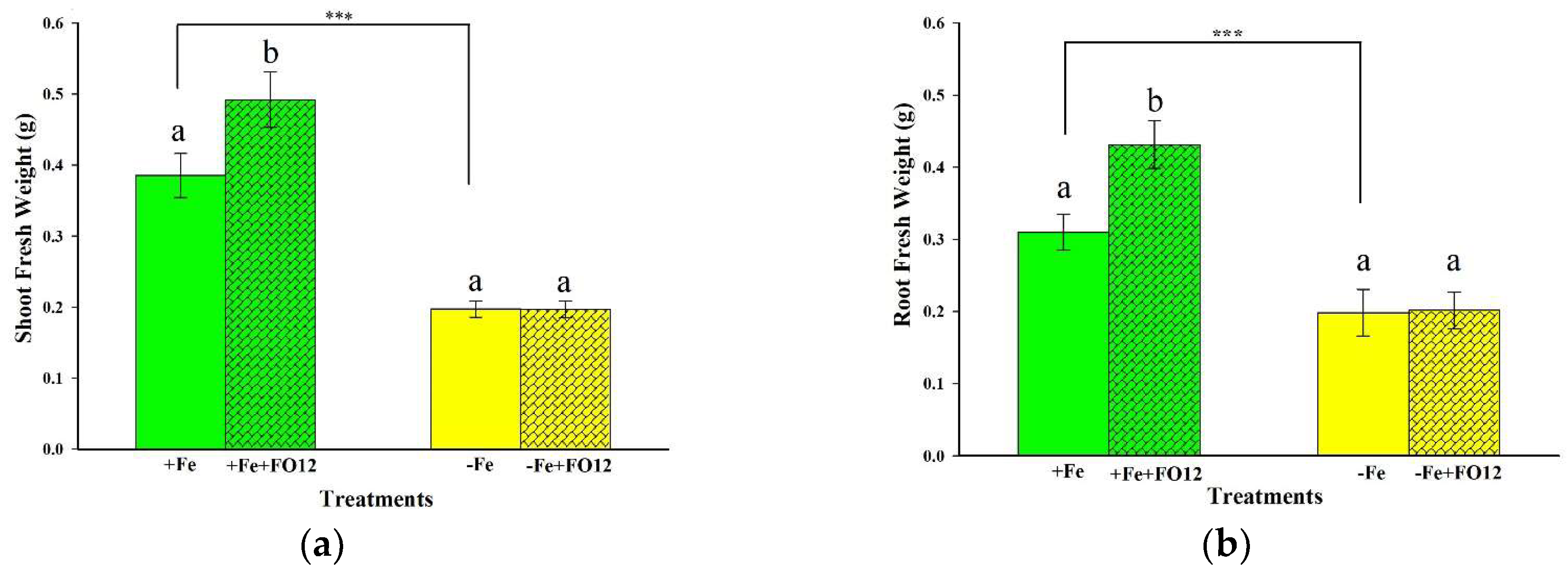
2.2. Effect of the Inoculation with FO12 on Growth Promotion
2.3. Effect of the Inoculation with FO12 on Phytosiderophores Production
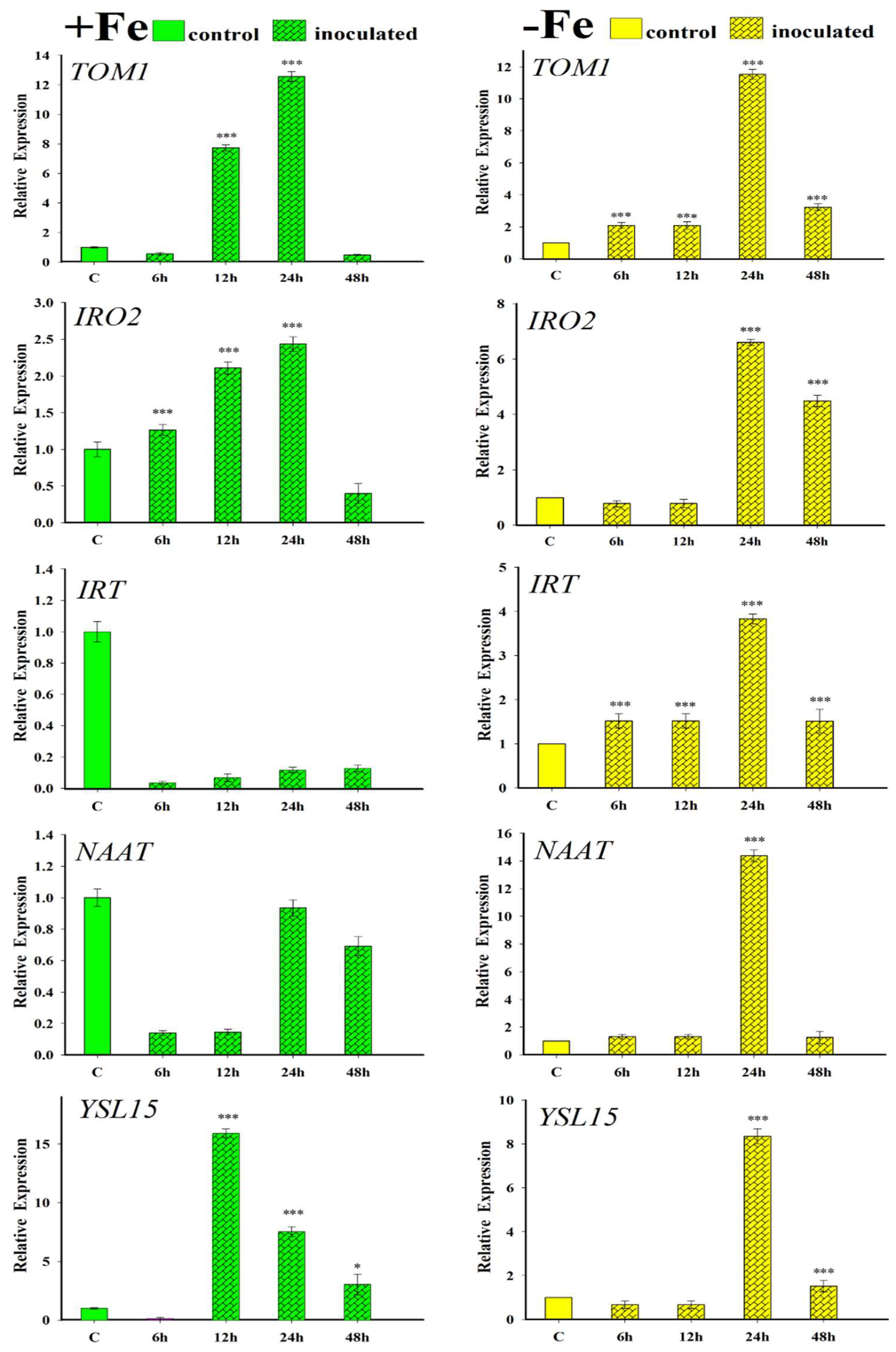
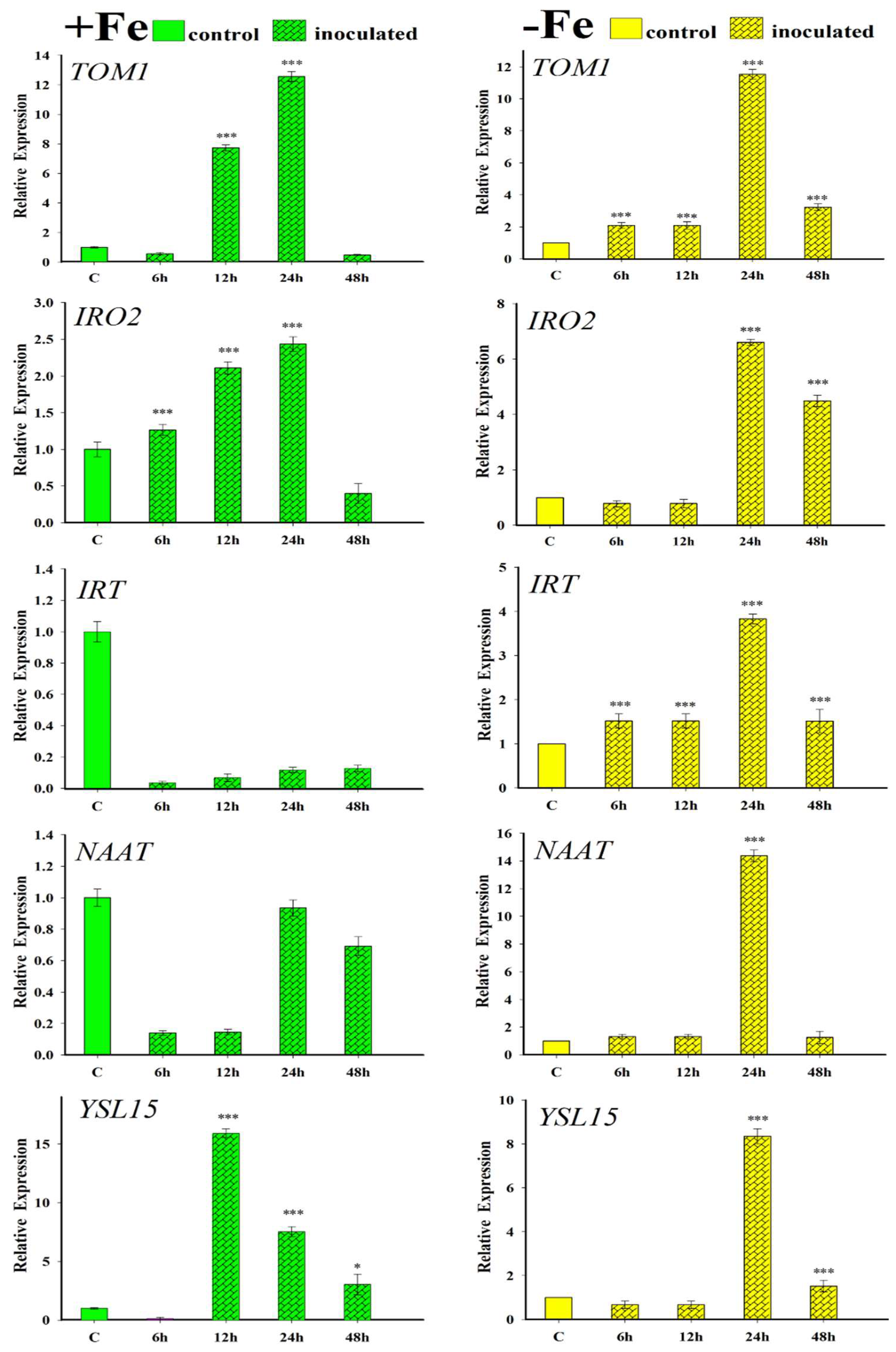
2.4. Effect of the Inoculation with FO12 on Expression of Fe-Related Genes
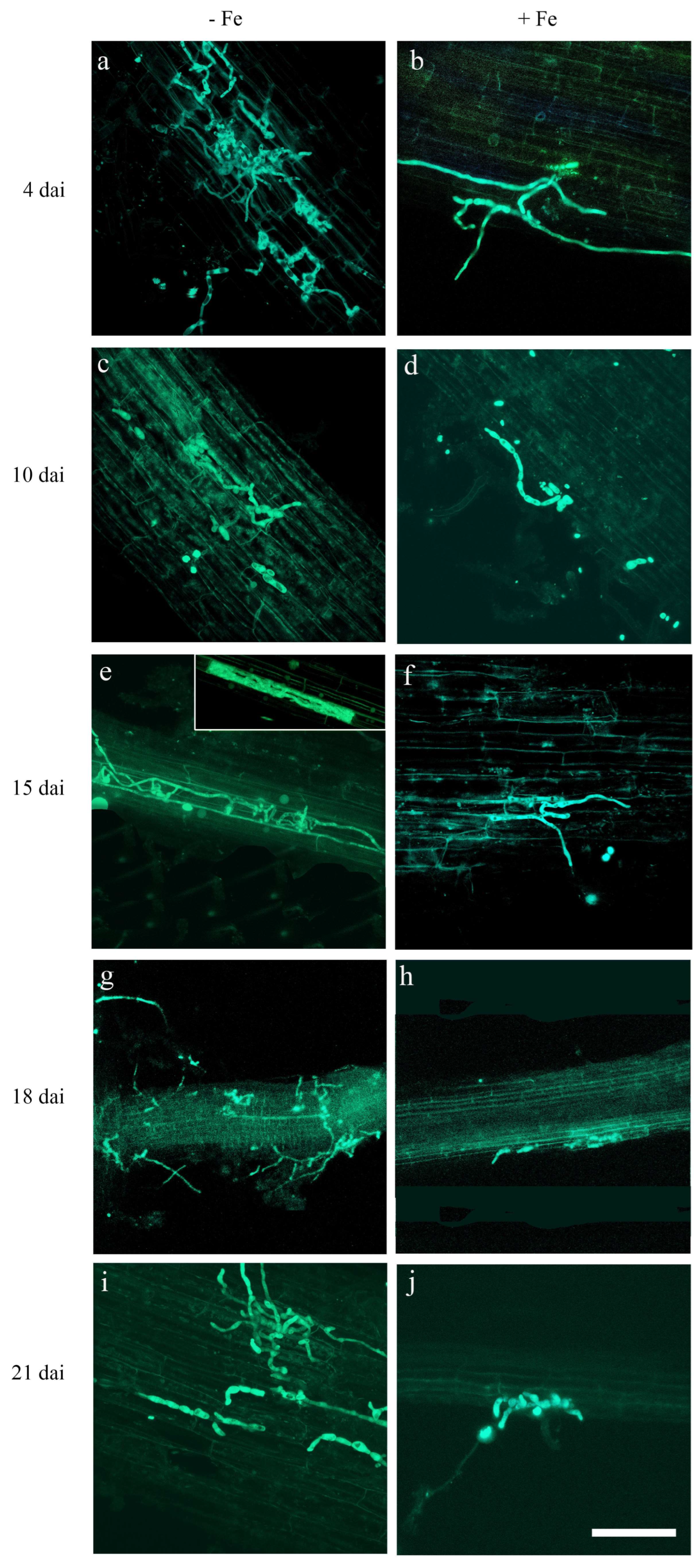
2.5. Iron Concentration in the Plant Substrate Affects FO12 Colonization in Rice Root Tissues
3. Discussion
4. Materials and Methods
4.1. Seed Germination and Plant Cultivation
4.2. Cultivation of Fungus and Inoculum Preparation
4.3. Plant Inoculation
4.4. Physiological and Morphological Assessments
4.5. Fusarium oxysporum Colonization Studies in Rice Roots
4.6. qRT-PCR Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations Department of Economic and Social Affairs Population Division. World Population Prospects; Summary of Results; United Nations: New York, NY, USA, 2022. [Google Scholar]
- Fukagawa, N.K.; Ziska, L.H. Rice: Importance for Global Nutrition. J. Nutr. Sci. Vitaminol. 2019, 65, S2–S3. [Google Scholar] [CrossRef] [PubMed]
- Horrigan, L.; Lawrence, R.S.; Walker, P. How sustainable agriculture can address the environmental and human health harms of industrial agriculture. Environ. Health Perspect. 2002, 110, 445–456. [Google Scholar] [CrossRef] [PubMed]
- Dubey, V. Impact of continuous use of chemical fertilizer. Int. J. Eng. Res. Dev. 2012, 3, 13–16. [Google Scholar]
- Savci, S. Investigation of effect of chemical fertilizers on environment. APCBEE Procedia 2012, 1, 287–292. [Google Scholar] [CrossRef]
- Bhardwaj, D.; Ansari, M.W.; Sahoo, R.K.; Tuteja, N. Biofertilizers function as key player in sustainable agriculture by improving soil fertility, plant tolerance and crop productivity. Microb. Cell Factories 2014, 13, 66. [Google Scholar] [CrossRef]
- FAO. The Future of Food and Agriculture—Alternative Pathways to 2050; FAO: Rome, Italy, 2018; (CC BY-NC-SA 3.0 IGO). [Google Scholar]
- Shen, J.; Li, C.; Mi, G.; Li, L.; Yuan, L.; Jiang, R.; Zhang, F. Maximizing root/rhizosphere efficiency to improve crop productivity and nutrient use efficiency in intensive agriculture of China. J. Exp. Bot. 2013, 64, 1181–1192. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.; Bakker, P.A. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef]
- Pathania, P. Role of plant growth—Promoting bacteria in sustainable agriculture. Biocatal. Agric. Biotechnol. 2020, 30, 101842. [Google Scholar] [CrossRef]
- Zamioudis, C.; Hanson, J.; Pieterse, C.M.J. β-Glucosidase BGLU42 is a MYB72-dependent key regulator of rhizobacteria-induced systemic resistance and modulates iron deficiency responses in Arabidopsis roots. New Phytol. 2014, 204, 368–379. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H.A.; López-Bucio, J.S.; Méndez-Bravo, A.; Macías-Rodríguez, L.; Ramos-Vega, M.; Guevara-García, Á.A.; López-Bucio, J. Mitogen-Activated protein kinase 6 and ethylene and auxin signaling pathways are involved in Arabidopsis root-system architecture alterations by Trichoderma atroviride. Mol. Plant-Microbe Interact. 2015, 28, 701–710. [Google Scholar] [CrossRef]
- García-López, A.M.; Avilés, M.; Delgado, A. Effect of various microorganisms on phosphorus uptake from insoluble Ca-phosphates by cucumber plants. J. Plant Nutr. Soil Sci. 2016, 179, 454–465. [Google Scholar] [CrossRef]
- Garnica-Vergara, A.; Barrera-Ortiz, S.; Muñoz-Parra, E.; Raya-González, J.; Méndez-Bravo, A.; Macías-Rodríguez, L.; Ruiz-Herrera, L.F.; López-Bucio, J. The volatile 6-pentyl-2H-pyran-2-one from Trichoderma atroviride regulates Arabidopsis thaliana root morphogenesis via auxin signaling and ETHYLENE INSENSITIVE 2 functioning. New Phytol. 2016, 209, 1496–1512. [Google Scholar] [CrossRef]
- Verbon, E.H.; Trapet, P.L.; Stringlis, I.A.; Kruijs, S.; Bakker, P.A.; Pieterse, C.M.J. Iron and Immunity. Annu. Rev. Phytopathol. 2017, 55, 355–375. [Google Scholar] [CrossRef]
- Briat, J.F.; Dubos, C.; Gaymard, F. Iron nutrition, biomass production, and plant product quality. Trends Plant Sci. 2015, 20, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Vélez-Bermúdez, I.C.; Schmidt, W. Plant strategies to mine iron from alkaline substrates. Plant Soil 2022, 483, 1–25. [Google Scholar] [CrossRef]
- Loeppert, R.H. Reactions of iron and carbonates in calcareous soils. J. Plant Nutr. 1986, 9, 195–214. [Google Scholar] [CrossRef]
- Romera, F.J.; García, M.J.; Lucena, C.; Martínez-Medina, A.; Aparicio, M.A.; Ramos, J.; Alcántara, E.; Angulo, M.; Pérez-Vicente, R. Induced systemic resistance (ISR) and Fe deficiency responses in dicot plants. Front. Plant Sci. 2019, 10, 287. [Google Scholar] [CrossRef]
- Taalab, A.; Ageeb, G.; Siam, H.S.; Mahmoud, S.A. Some Characteristics of Calcareous soils. Middle East J. 2019, 8, 96–105. [Google Scholar]
- Marschner, P. Rhizosphere Biology. In Marschner’s Mineral Nutrition of Higher Plants; Elsevier: Amsterdam, The Netherlands, 2012; pp. 369–388. [Google Scholar]
- Hänsch, R.; Mendel, R.R. Physiological functions of mineral micronutrients (Cu, Zn, Mn, Fe, Ni, Mo, B, Cl). Curr. Opin. Plant Biol. 2009, 12, 259–266. [Google Scholar] [CrossRef]
- Kroh, G.E.; Pilon, M. Regulation of Iron Homeostasis and Use in Chloroplasts. Int. J. Mol. Sci. 2020, 21, 3395. [Google Scholar] [CrossRef]
- Brumbarova, T.; Bauer, P.; Ivanov, R. Molecular mechanisms governing Arabidopsis iron uptake. Trends Plant Sci. 2015, 20, 124–133. [Google Scholar] [CrossRef]
- Lucena, C.; Romera, F.J.; García, M.J.; Alcántara, E.; Pérez-Vicente, R. Ethylene participates in the regulation of Fe deficiency responses in Strategy I plants and in rice. Front. Plant Sci. 2015, 6, 1056. [Google Scholar] [CrossRef]
- Li, W.; Lan, P. The understanding of the plant iron deficiency responses in Strategy I plants and the role of ethylene in this process by omic approaches. Front. Plant Sci. 2017, 8, 40. [Google Scholar] [CrossRef]
- Kobayashi, T.; Nakanishi Itai, R.; Nishizawa, N.K. Iron deficiency responses in rice roots. Rice 2014, 7, 27. [Google Scholar] [CrossRef]
- Mori, S. Iron acquisition by plants. Curr. Opin. Plant Biol. 1999, 2, 250–253. [Google Scholar] [CrossRef]
- Kobayashi, T.; Nishizawa, N.K. Iron uptake, translocation, and regulation in higher plants. Annu. Rev. Plant Biol. 2012, 63, 131–152. [Google Scholar] [CrossRef]
- Inoue, H.; Takahashi, M.; Kobayashi, T.; Suzuki, M.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. Identification and localisation of the rice nicotianamine aminotransferase gene OsNAAT1 expression suggests the site of phytosiderophore synthesis in rice. Plant Mol. Biol. 2008, 66, 193–203. [Google Scholar] [CrossRef]
- Ishimaru, Y.; Suzuki, M.; Tsukamoto, T.; Suzuki, K.; Nakazono, M.; Kobayashi, T.; Wada, Y.; Watanabe, S.; Matsuhashi, S.; Takahashi, M.; et al. Rice plants take up iron as an Fe3+-phytosiderophore and as Fe2+. Plant J. 2006, 45, 335–346. [Google Scholar] [CrossRef]
- Ishimaru, Y.; Kakei, Y.; Shimo, H.; Bashir, K.; Sato, Y.; Sato, Y.; Uozumi, N.; Nakanishi, H.; Nishizawa, N.K. A Rice Phenolic Efflux Transporter Is Essential for Solubilizing Precipitated Apoplasmic Iron in the Plant Stele. J. Biol. Chem. 2011, 286, 24649–24655. [Google Scholar] [CrossRef]
- Ricachenevsky, F.K.; Sperotto, R.A. There and back again, or always there? The evolution of rice combined strategy for Fe uptake. Front. Media SA 2014, 5, 189. [Google Scholar] [CrossRef]
- Wairich, A.; De Oliveira, B.H.N.; Arend, E.B.; Duarte, G.L.; Ponte, L.R.; Sperotto, R.A.; Ricachenevsky, F.K.; Fett, J.P. The combined strategy for iron uptake is not exclusive to domesticated rice (Oryza sativa). Sci. Rep. 2019, 9, 16144. [Google Scholar] [CrossRef] [PubMed]
- Rai, S.; Singh, P.K.; Mankotia, S.; Swain, J.; Satbhai, S.B. Iron homeostasis in plants and its crosstalk with copper, zinc, and manganese. Plant Stress 2021, 1, 100008. [Google Scholar] [CrossRef]
- Romera, F.J.; Lucena, C.; García, M.J.; Alcántara, E.; Pérez-Vicente, R. Stress Signaling in Plants: Genomics and Proteomics Perspectives; Springer Science and Business Media LLC.: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Romera, F.J.; Alcantara, E.; De La Guardia, M.D. Ethylene production by Fe-deficient roots and its involvement in the regulation of Fe-deficiency stress responses by Strategy I plants. Ann. Bot. 1999, 83, 51–55. [Google Scholar] [CrossRef]
- Wu, J.; Wang, C.; Zheng, L.; Wang, L.; Chen, Y.; Whelan, J.; Shou, H. Ethylene is involved in the regulation of iron homeostasis by regulating the expression of iron-acquisition-related genes in Oryza sativa. J. Exp. Bot. 2011, 62, 667–674. [Google Scholar] [CrossRef]
- Itai, R.N.; Ogo, Y.; Kobayashi, T.; Nakanishi, H.; Nishizawa, N.K. Rice genes involved in phytosiderophore biosynthesis are synchronously regulated during the early stages of iron deficiency in roots. Rice 2013, 6, 16. [Google Scholar] [CrossRef]
- Suzuki, M.; Takahashi, M.; Tsukamoto, T.; Watanabe, S.; Matsuhashi, S.; Yazaki, J.; Kishimoto, N.; Kikuchi, S.; Nakanishi, H.; Mori, S. Biosynthesis and secretion of mugineic acid family phytosiderophores in zinc-deficient barley. Plant J. 2006, 48, 85–97. [Google Scholar] [CrossRef]
- Li, Y.; Wang, N.; Zhao, F.; Song, X.; Yin, Z.; Huang, R.; Zhang, C. Changes in the transcriptomic profiles of maize roots in response to iron-deficiency stress. Plant Mol. Biol. 2014, 85, 349–363. [Google Scholar] [CrossRef]
- Alabouvette, C.; Olivain, C. Modes of action of non-pathogenic strains of Fusarium oxysporum in controlling Fusarium wilts. Plant Prot. Sci. 2002, 38, 195–199. [Google Scholar] [CrossRef]
- Patil, S.; Sriram, S.; Savitha, M.J.; Arulmani, N. Induced systemic resistance in tomato by non-pathogenic Fusarium species for the management of Fusarium wilt. Arch. Phytopathol. Plant Prot. 2011, 44, 1621–1634. [Google Scholar] [CrossRef]
- Varo, A.; Raya-Ortega, M.C.; Trapero, A. Selection and evaluation of micro-organisms for biocontrol of Verticillium dahliae in olive. J. Appl. Microbiol. 2016, 121, 767–777. [Google Scholar] [CrossRef]
- Constantin, M.E.; Vlieger, B.V.; Takken, F.L.W.; Rep, M. Diminished pathogen and enhanced endophyte colonization upon coinoculation of endophytic and pathogenic Fusarium strains. Microorganisms 2020, 8, 544. [Google Scholar] [CrossRef]
- De Lamo, F.J.; Takken, F.L. Biocontrol by Fusarium oxysporum using endophyte-mediated resistance. Front. Plant Sci. 2020, 11, 37. [Google Scholar] [CrossRef]
- Constantin, M.E.; de Lamo, F.J.; Vlieger, B.V.; Rep, M.; Takken, F.L.W. Endophyte-mediated resistance in tomato to Fusarium oxysporum is independent of ET, JA, and SA. Front. Plant Sci. 2019, 10, 979. [Google Scholar] [CrossRef] [PubMed]
- Veloso, J.; Díaz, J. Fusarium oxysporum Fo47 confers protection to pepper plants against Verticillium dahliae and Phytophthora capsici, and induces the expression of defence genes. Plant Pathol. 2012, 61, 281–288. [Google Scholar] [CrossRef]
- Aparicio, M.A.; Lucena, C.; García, M.J.; Ruiz-Castilla, F.J.; Jiménez-Adrián, P.; López-Berges, M.S.; Prieto, P.; Alcántara, E.; Pérez-Vicente, R.; Ramos, J.; et al. The nonpathogenic strain of Fusarium oxysporum FO12 induces Fe deficiency responses in cucumber (Cucumis sativus L.) plants. Planta 2023, 257, 50. [Google Scholar] [CrossRef] [PubMed]
- Mulero-Aparicio, A.; Agustí Brisach, C.; Varo, A.; López-Escudero, F.J.; Trapero, A. A non-pathogenic strain of Fusarium oxysporum as a potential biocontrol agent against Verticillium wilt of olive. Biol. Control 2019, 139, 104045. [Google Scholar] [CrossRef]
- Mulero-Aparicio, A.; Cernava, T.; Turrà, D.; Schaefer, A.; Di Pietro, A.; López-Escudero, F.J.; Trapero, A.; Berg, G. The Role of Volatile Organic Compounds and Rhizosphere Competence in Mode of Action of the Non-pathogenic Fusarium oxysporum FO12 Toward Verticillium Wilt. Front. Microbiol. 2019, 10, 1808. [Google Scholar] [CrossRef]
- Mulero-Aparicio, A.; Trapero, F.J.; López-Escudero, F.J. A nonpathogenic strain of Fusarium oxysporum and grape marc compost control Verticillium wilt of olive. Phytopathol. Mediterr. 2020, 59, 159–167. [Google Scholar] [CrossRef]
- Mulero-Aparicio, A.; Varo, A.; Agustí-Brisach, C.; López-Escudero, F.J.; Trapero, A. Biological control of Verticillium wilt of olive in the field. Crop Prot. 2020, 128, 104993. [Google Scholar] [CrossRef]
- Tottey, S.; Block, M.A.; Allen, M.; Westergren, T.; Albrieux, C.; Scheller, H.V.; Merchant, S.; Jensen, P.E. Arabidopsis CHL27, located in both envelope and thylakoid membranes, is required for the synthesis of protochlorophyllide. Proc. Natl. Acad. Sci. USA 2003, 100, 16119–16124. [Google Scholar] [CrossRef]
- Velazquez, E.; Peix, A.; Zurdo-Pineiro, J.L.; Palomo, J.L.; Mateos, P.F.; Rivas, R.; Munoz-Adelantodo, E.; Toro, N.; Garcia-Benavides, P.; Martinez-Molina, E. The coexistence of symbiosis and pathogenicity-determining genes in Rhizobium rhizogenes strains enables them to induce nodules and tumors or hairy roots in plants. Mol. Plant-Microbe Interact. 2005, 18, 1325–1332. [Google Scholar] [CrossRef] [PubMed]
- Berg, G. Plant–microbe interactions promoting plant growth and health: Perspectives for controlled use of microorganisms in agriculture. Appl. Microbiol. Biotechnol. 2009, 84, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.E.; Barea, J.M.; McNeill, A.M.; Prigent-Combaret, C. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil. 2009, 321, 305–339. [Google Scholar] [CrossRef]
- Halpern, M.; Bar-Taly, A.; Ofeky, M.; Minzy, D.; Mullerx, T.; Yermiyahu, U. The Use of biostimulants for enhancing nutrient uptake. Adv. Agron. 2015, 130, 141–174. [Google Scholar] [CrossRef]
- Doni, F.; Fathurrahman, F.; Mispan, M.S.; Suhaimi, N.S.M.; Yusoff, W.M.W.; Uphoff, N. Transcriptomic profiling of rice seedlings inoculated with the symbiotic fungus Trichoderma asperellum SL2. J. Plant Growth Regul. 2019, 38, 1507–1515. [Google Scholar] [CrossRef]
- Pal, G.; Kumar, K.; Verma, A.; White, J.F.; Verma, S.K. Functional Roles of Seed-Inhabiting Endophytes of Rice. In Seed Endophytes: Biology and Biotechnology; Verma, S.K., White, J.J.F., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2019; pp. 213–236. [Google Scholar] [CrossRef]
- Hossain, G.M.A.; Ghazali, A.H.; Islam, T.; Mia, M.A.B. Enhanced Nutrient Accumulation in Non-Leguminous Crop Plants by the Application of Endophytic Bacteria Bacillus Species. In Bacilli in Agrobiotechnology: Plant Stress Tolerance, Bioremediation, and Bioprospecting; Islam, M.T., Rahman, M., Pandey, P., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2022; pp. 349–364. [Google Scholar] [CrossRef]
- Verma, N.; Narayan, O.P.; Prasad, D.; Jogawat, A.; Panwar, S.L.; Dua, M.; Johri, A.K. Functional characterization of a high-affinity iron transporter PiFTR) from the endophytic fungus Piriformospora indica and its role in plant growth and development. Environ. Microbiol. 2022, 24, 689–706. [Google Scholar] [CrossRef]
- Bakhshandeh, E.; Gholamhosseini, M.; Yaghoubian, Y.; Pirdashti, H. Plant growth promoting microorganisms can improve germination, seedling growth and potassium uptake of soybean under drought and salt stress. Plant Growth Regul. 2020, 90, 123–136. [Google Scholar] [CrossRef]
- Fontenelle, A.D.B.; Guzzo, S.D.; Lucon, C.M.M.; Harakava, R. Growth promotion and induction of resistance in tomato plant against Xanthomonas euvesicatoria and Alternaria solani by Trichoderma spp. Crop Prot. 2011, 30, 1492–1500. [Google Scholar] [CrossRef]
- Ipek, M.; Pirlak, L.; Esitken, A.; Figen Dönmez, M.; Turan, M.; Sahin, F. Plant growth-promoting rhizobacteria (Pgpr) increase yield, growth and nutrition of strawberry under high-calcareous soil conditions. J. Plant Nutr. 2014, 37, 990–1001. [Google Scholar] [CrossRef]
- Liu, J.; Tang, L.; Gao, H.; Zhang, M.; Guo, C. Enhancement of alfalfa yield and quality by plant growth-promoting rhizobacteria under saline-alkali conditions. J. Sci. Food Agric. 2019, 99, 281–289. [Google Scholar] [CrossRef]
- El_Komy, M.H.; Hassouna, M.G.; Abou-Taleb, E.M.; Al-Sarar, A.S.; Abobakr, Y. A mixture of Azotobacter, Azospirillum, and Klebsiella strains improves root-rot disease complex management and promotes growth in sunflowers in calcareous soil. Eur. J. Plant Pathol. 2020, 156, 713–726. [Google Scholar] [CrossRef]
- Roesti, D.; Gaur, R.; Johri, B.; Imfeld, G.; Sharma, S.; Kawaljeet, K.; Aragno, M. Plant growth stage, fertiliser management and bio-inoculation of arbuscular mycorrhizal fungi and plant growth promoting rhizobacteria affect the rhizobacterial community structure in rain–Fed wheat fields. Soil Biol. Biochem. 2006, 38, 1111–1120. [Google Scholar] [CrossRef]
- Kavroulakis, N.; Ntougias, S.; Zervakis, G.I.; Ehaliotis, C.; Haralampidis, K.; Papadopoulou, K.K. Role of ethylene in the protection of tomato plants against soil-borne fungal pathogens conferred by an endophytic Fusarium solani strain. J. Exp. Bot. 2007, 58, 3853–3864. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.J.; Lucena, C.; Romera, F.J.; Alcantara, E.; Perez-Vicente, R. Ethylene and nitric oxide involvement in the up-regulation of key genes related to iron acquisition and homeostasis in Arabidopsis. J. Exp. Bot. 2010, 61, 3885–3899. [Google Scholar] [CrossRef]
- Garcia, M.J.; Romera, F.J.; Alcantara, E.; Perez-Vicente, R. A new model involving ethylene, nitric oxide and Fe to explain the regulation of Fe-acquisition genes in Strategy I plants. Plant Physiol. Biochem. 2011, 49, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Zamioudis, C.; Korteland, J.; Van Pelt, J.A.; van Hamersveld, M.; Dombrowski, N.; Bai, Y.; Hanson, J.; Van Verk, M.C.; Ling, H.Q.; Schulze-Lefert, P.; et al. Rhizobacterial volatiles and photosynthesis-related signals coordinate MYB72 expression in Arabidopsis roots during onset of induced systemic resistance and iron-deficiency responses. Plant J. 2015, 84, 309–322. [Google Scholar] [CrossRef]
- Zhou, C.; Guo, J.; Zhu, L.; Xiao, X.; Xie, Y.; Zhu, J.; Ma, Z.; Wang, J. Paenibacillus polymyxa BFKC01 enhances plant iron absorption via improved root systems and activated iron acquisition mechanisms. Plant Physiol. Biochem. 2016, 105, 162–173. [Google Scholar] [CrossRef]
- Martínez-Medina, A.; Van Wees, S.C.M.; Pieterse, C.M.J. Airborne signals from Trichoderma fungi stimulate iron uptake responses in roots resulting in priming of jasmonic acid-dependent defences in shoots of Arabidopsis thaliana and Solanum lycopersicum. Plant Cell Environ. 2017, 40, 2691–2705. [Google Scholar] [CrossRef]
- Guerra, C.; Anderson, P. The effect of iron and boron amendments on infection of bean by Fusarium solani. Phytopathology 1985, 75, 989–991. [Google Scholar] [CrossRef]
- Navarro-Velasco, G.Y.; Prados-Rosales, R.C.; Ortiz-Urquiza, A.; Quesada-Moraga, E.; Di Pietro, A. Galleria mellonella as model host for the trans-kingdom pathogen Fusarium oxysporum. Fungal Genet. Biol. 2011, 48, 1124–1129. [Google Scholar] [CrossRef]
- Inal, A.; Gunes, A.; Zhang, F.; Cakmak, I. Peanut/maize intercropping induced changes in rhizosphere and nutrient concentrations in shoots. Plant Physiol. Biochem. 2007, 45, 350–356. [Google Scholar] [CrossRef]
- Reichman, S.M.; Parker, D.R. Critical evaluation of three indirect assays for quantifying phytosiderophores released by the roots of Poaceae. Eur. J. Soil Sci. 2007, 58, 844–853. [Google Scholar] [CrossRef]
- Prieto, P.; Moore, G.; Shaw, P. Fluorescence in situ hybridization on vibratome sections of plant tissues. Nat. Protoc. 2007, 2, 1831–1838. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence 5-3 |
|---|---|
| OsNAAT1 | Forward: TAAGAG GATAATTGATTTGCTTAC |
| Reverse: CTG ATCATTCCAATCCTAGTACAAT | |
| OsYSL15 | Forward: AACATAAGGGGGACTG GTAC |
| Reverse: TGATTACCGCAATGATGCTTAG | |
| OsIRO2 | Forward: CTCCCATCGTTTCGGCTACCT |
| Reverse: GCTGGGCACTCCTCGTTGATC | |
| OsTOM1 | Forward: GCCCAAGAACGCCAAAATGA |
| Reverse: GGCTTGAAGGTCAACGCAAG | |
| OsIRT1 | Forward: CGTC TTCTTCTTCTCCACCACGAC |
| Reverse: GCAGCTGATGATCGAGTCTG ACC | |
| OsActin | Forward: TGCTATGTACGTCGC CATCCAG |
| Reverse: AATGAGTAACCACGCTCCGTCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Núñez-Cano, J.; Romera, F.J.; Prieto, P.; García, M.J.; Sevillano-Caño, J.; Agustí-Brisach, C.; Pérez-Vicente, R.; Ramos, J.; Lucena, C. Effect of the Nonpathogenic Strain Fusarium oxysporum FO12 on Fe Acquisition in Rice (Oryza sativa L.) Plants. Plants 2023, 12, 3145. https://doi.org/10.3390/plants12173145
Núñez-Cano J, Romera FJ, Prieto P, García MJ, Sevillano-Caño J, Agustí-Brisach C, Pérez-Vicente R, Ramos J, Lucena C. Effect of the Nonpathogenic Strain Fusarium oxysporum FO12 on Fe Acquisition in Rice (Oryza sativa L.) Plants. Plants. 2023; 12(17):3145. https://doi.org/10.3390/plants12173145
Chicago/Turabian StyleNúñez-Cano, Jorge, Francisco J. Romera, Pilar Prieto, María J. García, Jesús Sevillano-Caño, Carlos Agustí-Brisach, Rafael Pérez-Vicente, José Ramos, and Carlos Lucena. 2023. "Effect of the Nonpathogenic Strain Fusarium oxysporum FO12 on Fe Acquisition in Rice (Oryza sativa L.) Plants" Plants 12, no. 17: 3145. https://doi.org/10.3390/plants12173145
APA StyleNúñez-Cano, J., Romera, F. J., Prieto, P., García, M. J., Sevillano-Caño, J., Agustí-Brisach, C., Pérez-Vicente, R., Ramos, J., & Lucena, C. (2023). Effect of the Nonpathogenic Strain Fusarium oxysporum FO12 on Fe Acquisition in Rice (Oryza sativa L.) Plants. Plants, 12(17), 3145. https://doi.org/10.3390/plants12173145