
Cover Crop Yield, Nutrient Storage and Release under Different Cropping Technologies in the Sustainable Agrosystems


Abstract
:1. Introduction
2. Results and Discussion
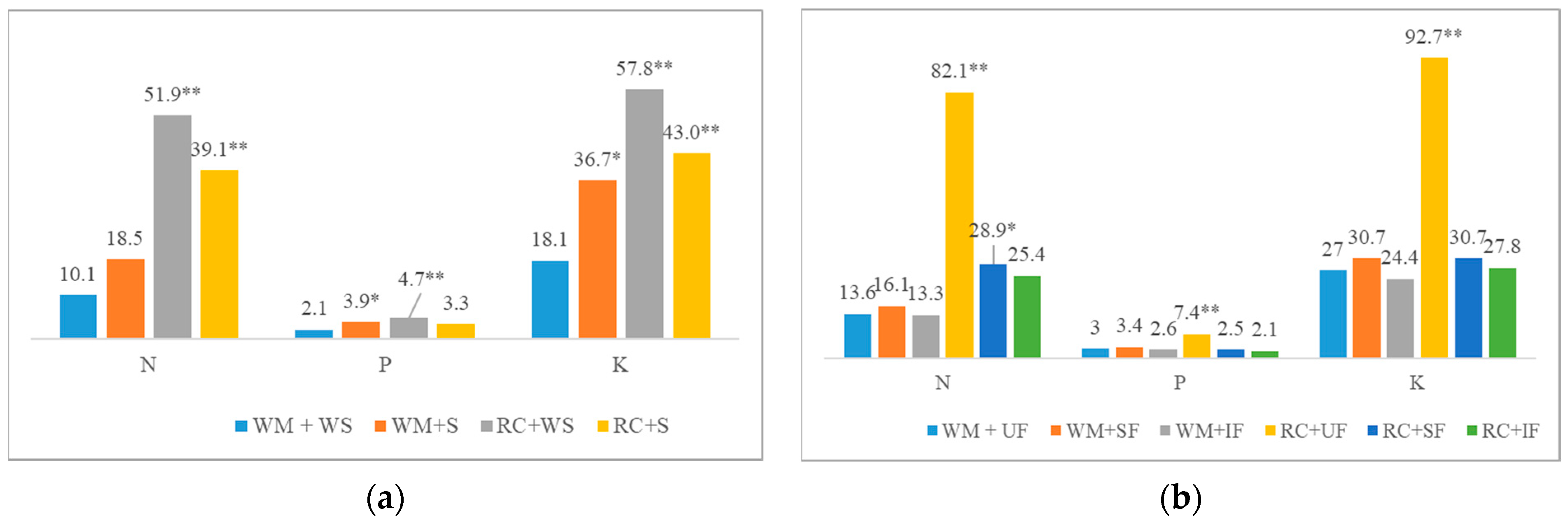
2.1. The Influence of Cover Crop Types, Their Sowing Methods, and the Fertilization of Main Cropss and the Use of Straw as Fertilizer on Cover Biomass and Accumulated Nutrient Content
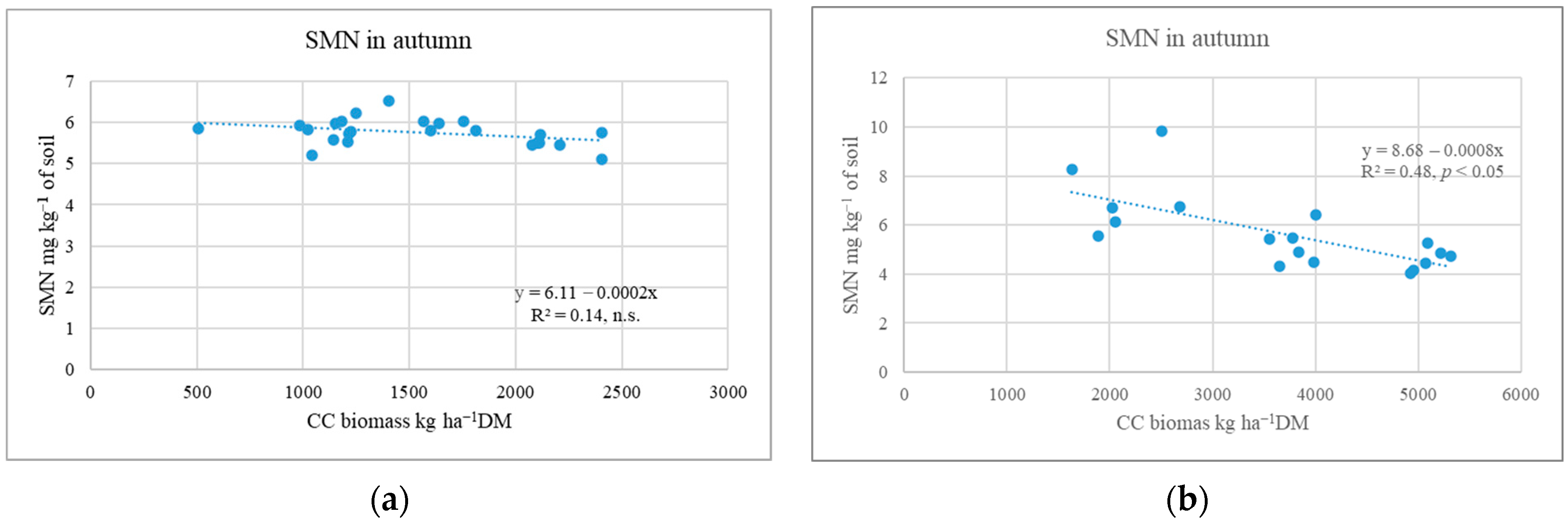
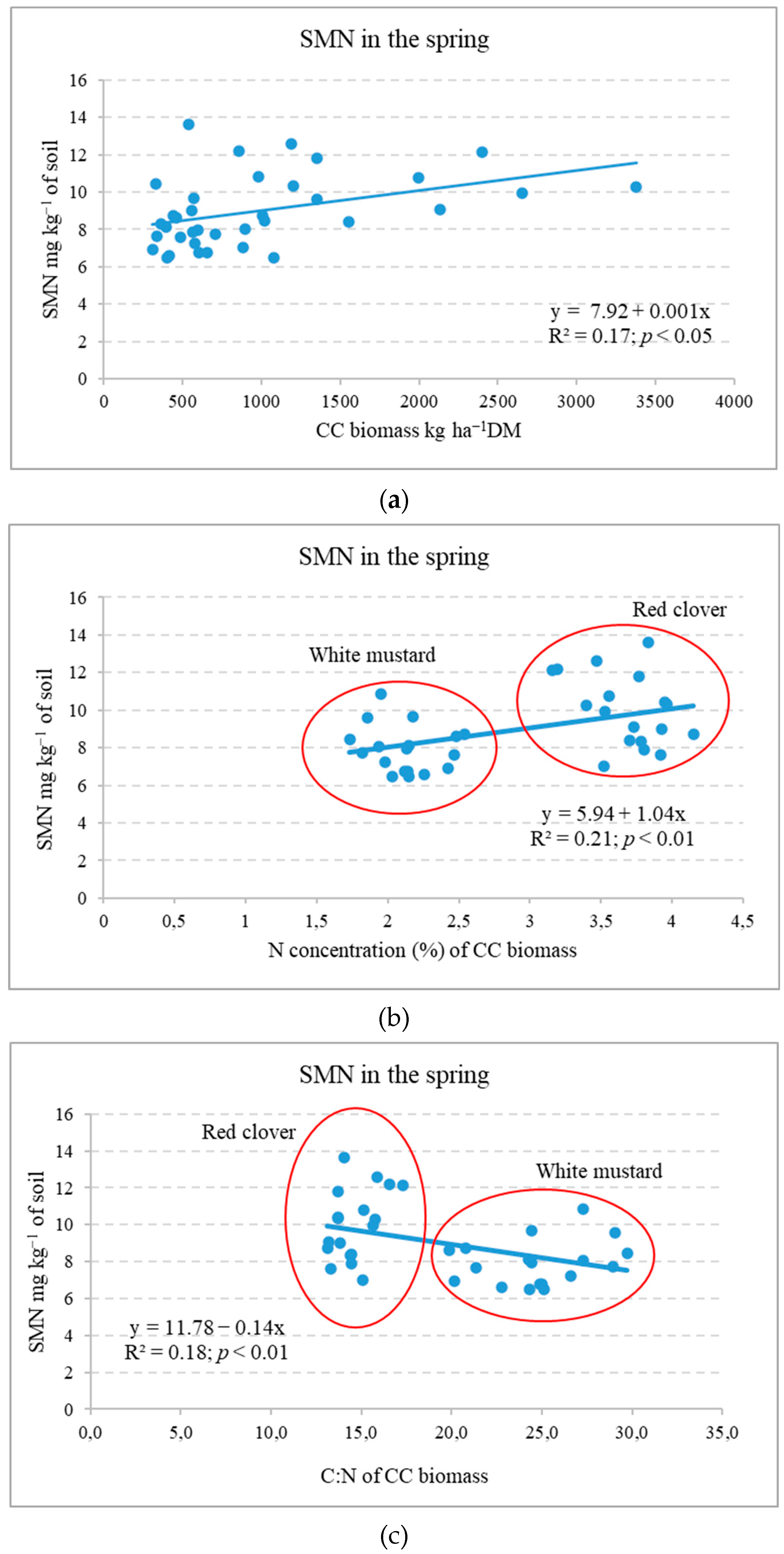
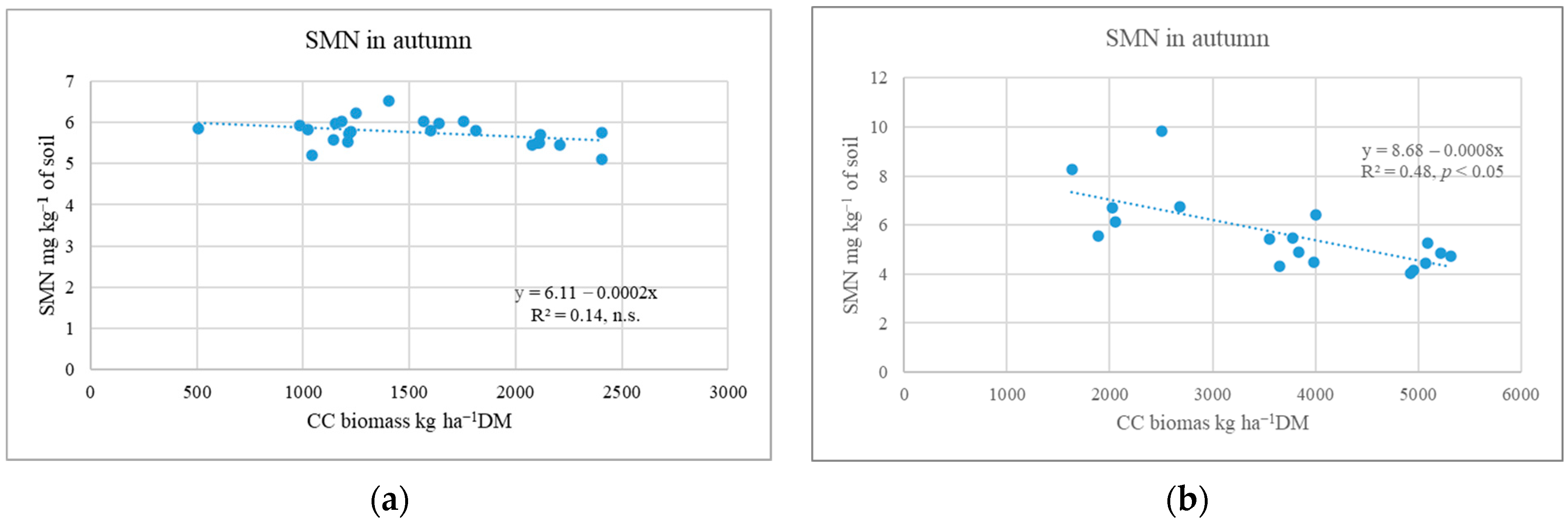
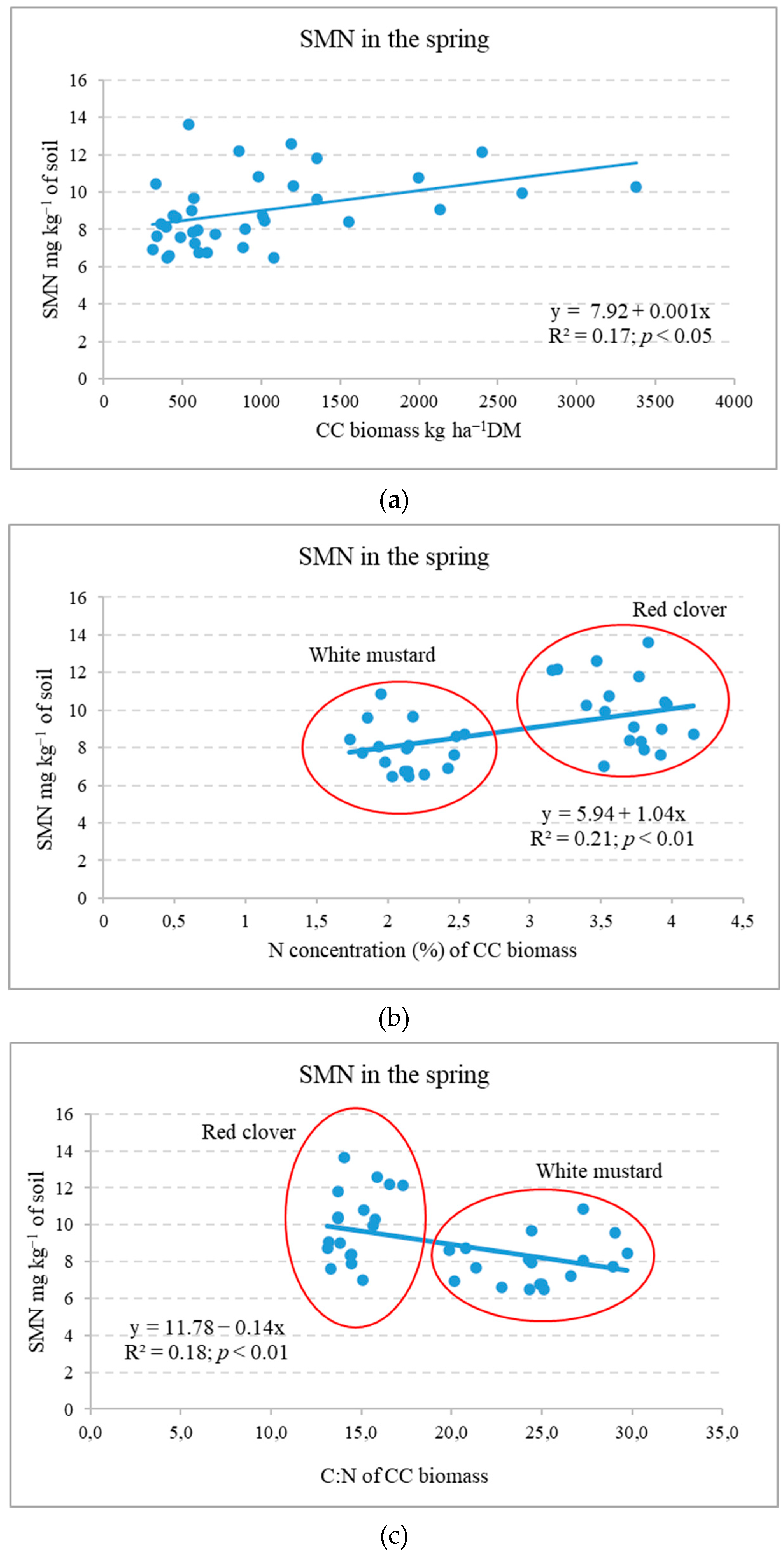
2.2. Influence of Cover Crops on Soil Mineral N Variation in Autumn and Spring
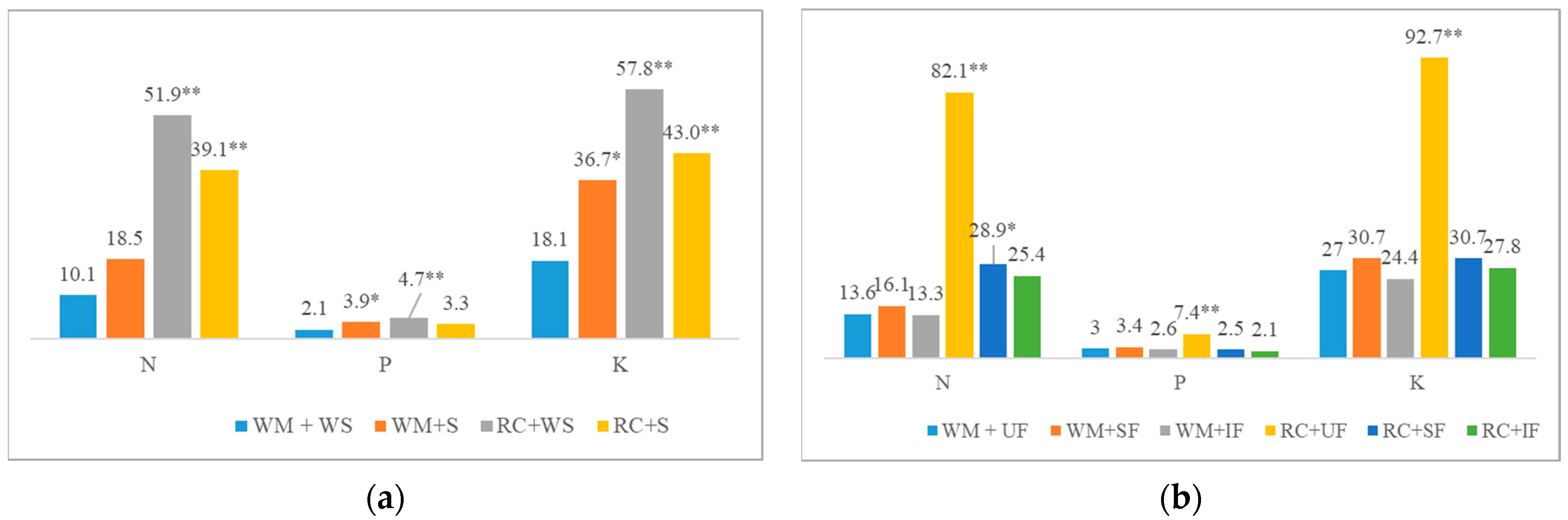
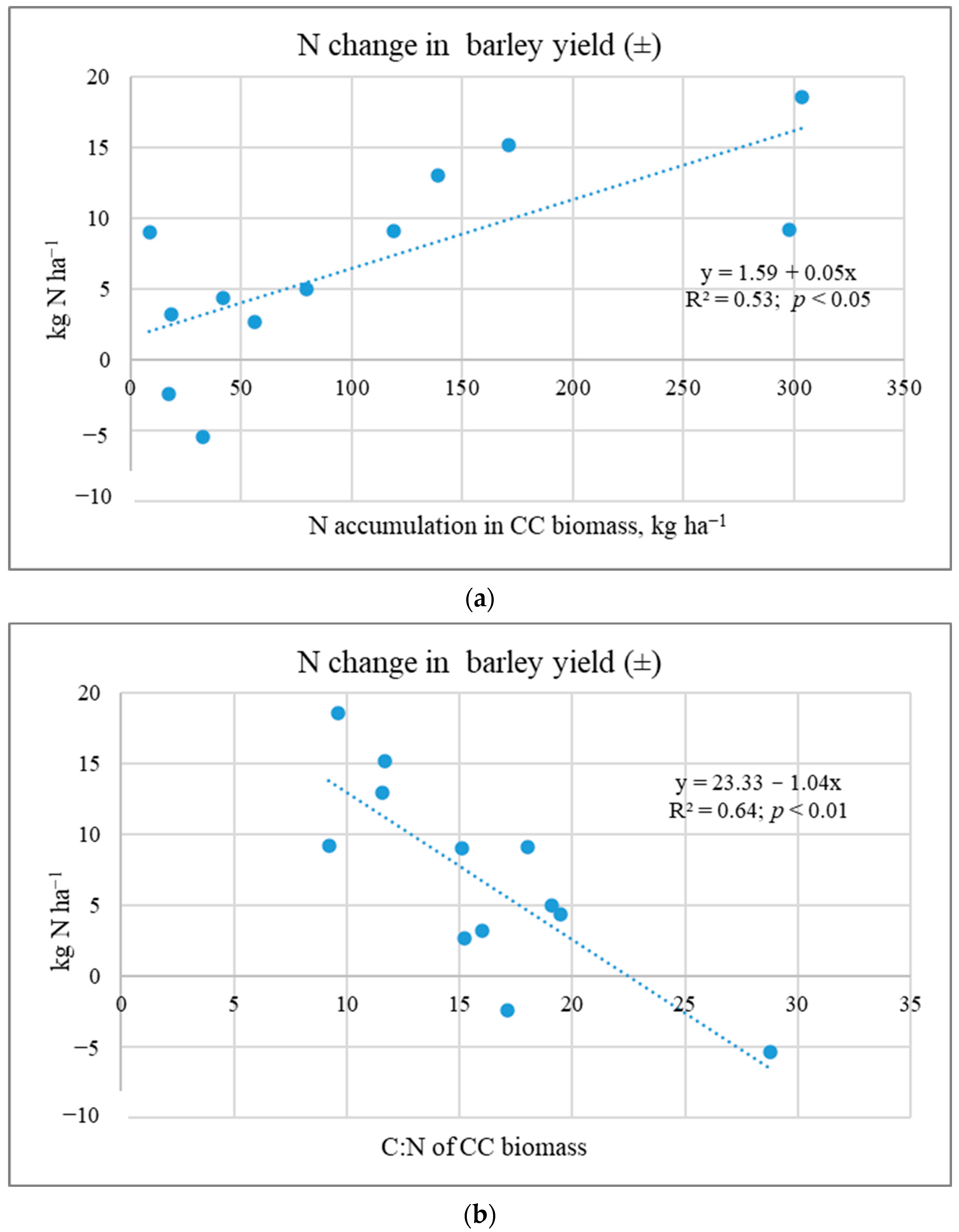
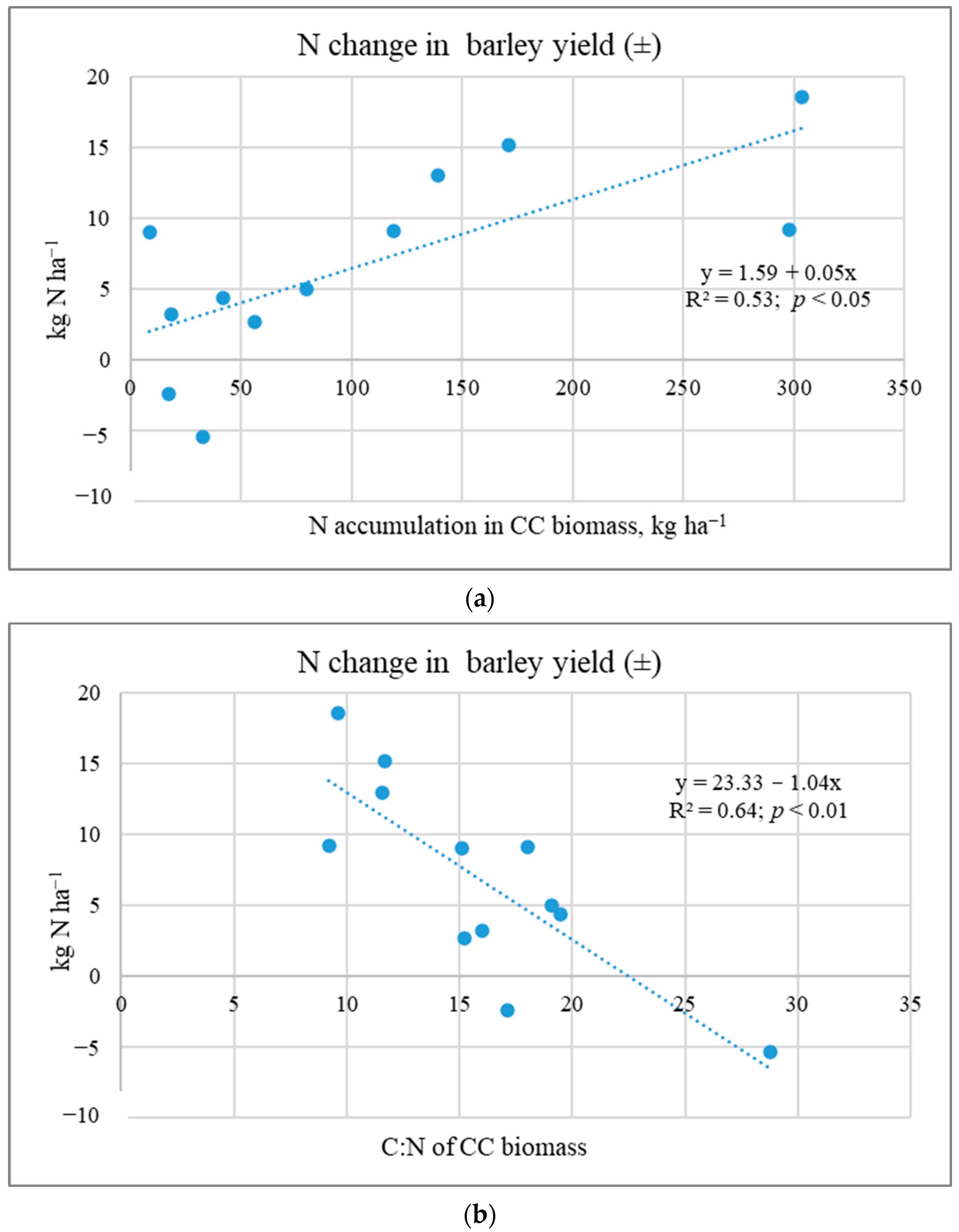
2.3. Nutrient Release from CC Shoot Biomass to Subsequent Crops
3. Materials and Methods
3.1. Experimental Sites
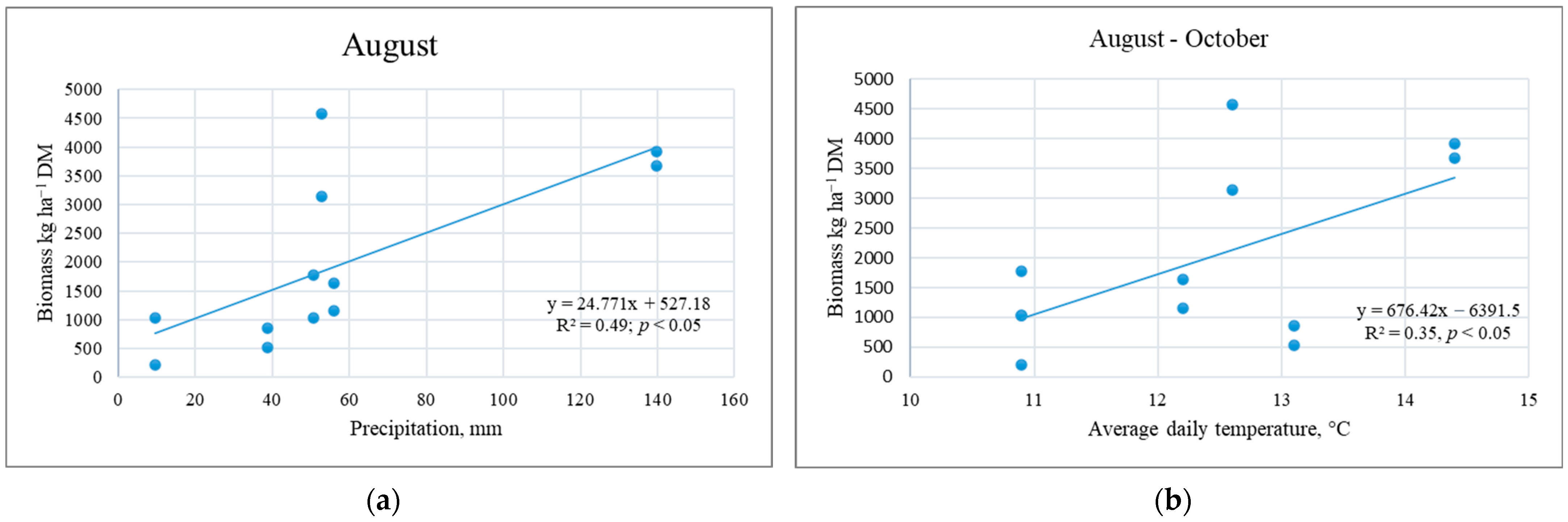
3.2. Meteorological Conditions
3.3. Experimental Designs and Details
3.4. Plant and Soil Analyses
3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ladha, J.K.; Tirol-Padre, A.; Reddy, C.K.; Cassman, K.G.; Verma, S.; Powlson, D.S.; Van Kessel, C.; De Richter, D.B.; Chakraborty, D.; Pathak, H. Global nitrogen budgets in cereals: A 50-year assessment for maize, rice, and wheat production systems. Sci. Rep. 2016, 6, 19355. [Google Scholar] [CrossRef]
- Kim, N.; Zabaloy, M.C.; Guan, K.; Villamil, M.B. Do cover crops benefit soil microbiome? A meta-analysis of current research. Soil Biol. Biochem. 2020, 142, 107701. [Google Scholar] [CrossRef]
- Faghih, H.; Behmanesh, J.; Rezaie, H.; Khalili, K. Climate and rainfed wheat yield. Theor. Appl. Climatol. 2021, 144, 13–24. [Google Scholar] [CrossRef]
- Klages, S.; Heidecke, C.; Osterburg, B. The impact of agricultural production and policy on water quality during the dry year 2018, a case study from Germany. Water 2020, 12, 1519. [Google Scholar] [CrossRef]
- Stancevičius, A.; Bogužas, V.; Trečiokas, K. The role of catch crops in Lithuanian agriculture. In Present and Future of Agricultural Science; Petraitis, V., Ed.; Akademija: Kėdainiai, Lithuania, 1996; pp. 108–115. [Google Scholar]
- Maikštėnienė, S. (Ed.) Sustainable Agriculture in Productive Soils; Lithuanian Institute of Agriculture: Kėdainiai, Lithuania, 2008; pp. 140–167. [Google Scholar]
- Blanco-Canqui, H.; Shaver, T.M.; Lindquist, J.L.; Shapiro, C.A.; Elmore, R.W.; Francis, C.A.; Hergert, G.W. Cover Crops and Ecosystem Services: Insights from Studies in Temperate Soils. Agron. Hortic. Fac. Publ. 2015, 107, 2449–2474. [Google Scholar] [CrossRef]
- Scavo, A.; Fontanazza, S.; Restuccia, A.; Pesce, G.R.; Abbate, C.; Mauromicale, G. The role of cover crops in improving soil fertility and plant nutritional status in temperate climates. A review. Agron. Sustain. Dev. 2022, 42, 93. [Google Scholar] [CrossRef]
- Abdalla, M.; Hastings, A.; Cheng, K.; Yue, Q.; Chadwick, D.; Espenberg, M.; Truu, J.; Rees, R.M.; Smith, P. A critical review of the impacts of cover crops on nitrogen leaching, net greenhouse gas balance and crop productivity. Glob. Chang. Biol. 2019, 25, 2530–2543. [Google Scholar] [CrossRef]
- Thapa, R.; Mirsky, S.B.; Tully, K.L. Cover crops reduce nitrate leaching in agroecosystems: A global meta-analysis. J. Environ. Qual. 2018, 47, 1400–1411. [Google Scholar] [CrossRef]
- McClelland, S.C.; Paustian, K.; Schipanski, M.E. Management of cover crops in temperate climates influences soil organic carbon stocks: A meta-analysis. Ecol. Appl. 2021, 31, e02278. [Google Scholar] [CrossRef]
- Rumpel, C.; Amiraslani, F.; Chenu, C.; Garcia Cardenas, M.; Kaonga, M.; Koutika, L.S.; Ladha, J.; Madari, B.; Shirato, Y.; Smith, P.; et al. The 4p1000 initiative: Opportunities, limitations and challenges for implementing soil organic carbon sequestration as a sustainable development strategy. Ambio 2020, 49, 350–360. [Google Scholar] [CrossRef]
- Haruna, S.I.; Nkongolo, N.V.; Anderson, S.H.; Eivazi, F.; Zaibon, S. In situ infiltration as influenced by cover crop and tillage management. J. Soil Water Conserv. 2018, 73, 164–172. [Google Scholar] [CrossRef]
- Chalise, K.S.; Singh, S.; Wegner, B.R.; Kumar, S.; Pérez-Gutiérrez, J.D.; Osborne, S.L.; Nleya, T.; Guzman, J.; Rohila, J.S. Cover Crops and returning residue impact on soil organic carbon, bulk density, penetration resistance, water retention, infiltration, and soybean yield. Agron. J. 2019, 111, 99–108. [Google Scholar] [CrossRef]
- Blanco-Canqui, H.; Ruis, S.J. No-tillage and soil physical environment. Geoderma 2018, 326, 64–200. [Google Scholar] [CrossRef]
- Wagg, C.; Bender, S.F.; Widmer, F.; van der Heijden, M.G. Soil biodiversity and soil community composition determine ecosystem multifunctionality. Proc. Natl. Acad. Sci. USA 2014, 111, 5266–5270. [Google Scholar] [CrossRef]
- Luna, J.M.; Sullivan, D.; Garrett, A.M.; Xue, L. Cover crop nitrogen contribution to organic broccoli production. Renew. Agric. Food Syst. 2018, 35, 49–58. [Google Scholar] [CrossRef]
- Garba, I.I.; Bell, L.W.; Williams, A. Cover crop legacy impacts on soil water and nitrogen dynamics, and on subsequent crop yields in drylands: A meta-analysis. Agron. Sustain. Dev. 2022, 42, 34. [Google Scholar] [CrossRef]
- Sharma, P.; Singh, A.; Kahlon, C.S.; Brar, A.S.; Grover, K.K.; Dia, M.; Steiner, R.L. The role of cover crops towards sustainable soil health and agriculture—A review paper. Am. J. Plant Sci. 2018, 9, 1935–1951. [Google Scholar] [CrossRef]
- Lawson, A.; Cogger, C.; Bary, A.; Fortuna, A.M. Influence of seeding ratio, planting date, and termination date on Rye-Hairy Vetch cover crop mixture performance under organic management. PLoS ONE 2015, 10, e0129597. [Google Scholar] [CrossRef]
- Hansen, V.; Eriksen, J.; Jensen, L.S.; Thorup-Kristensen, K.; Magid, J. Towards integrated cover crop management: N, P and S release from aboveground and belowground residues. Agric. Ecosyst. Environ. 2021, 313, 107392. [Google Scholar] [CrossRef]
- Kanders, M.J.; Berendonk, C.; Fritz, C.; Watson, C.; Wichern, F. Catch crops store more nitrogen below-ground when considering Rhizodeposits. Plant Soil 2017, 417, 287–299. [Google Scholar] [CrossRef]
- Finney, D.M.; White, C.M.; Kaye, J.P. Biomass production and carbon/nitrogen ratio influence ecosystem services from cover crop mixtures. Agron. J. 2016, 108, 39–52. [Google Scholar] [CrossRef]
- Sievers, T.; Cook, R.L. Aboveground and root decomposition of cereal Rye and Hairy Vetch cover crops. Soil Sci. Soc. Am. J. 2018, 82, 147–155. [Google Scholar] [CrossRef]
- Constantin, J.; Mary, B.; Laurent, F.; Aubrion, G.; Fontaine, A.; Kerveillant, P.; Beaudoin, N. Effects of catch crops, no till and reduced nitrogen fertilization on nitrogen leaching and balance in three long-term experiments. Agric. Ecosyst. Environ. 2010, 135, 268–278. [Google Scholar] [CrossRef]
- Grosse, M.; Haase, T.; Heß, J. Tillage system comparison in organic farming: Effects on N mineralization, soil microbial biomass, and yield. Org. Farming 2019, 5, 23–36. [Google Scholar] [CrossRef]
- Arlauskienė, A.; Gecaitė, V.; Toleikienė, M.; Šarūnaitė, L.; Kadžiulienė, Ž. Soil nitrate nitrogen content and grain yields of organically grown cereals as affected by a strip tillage and forage legume intercropping. Plants 2021, 10, 1453. [Google Scholar] [CrossRef]
- Teixeira, E.I.; Johnstone, P.; Chakwizira, E.; de Ruiter, J.; Malcolm, B.; Shaw, N.; Zyskowski, R.; Khaembah, E.; Sharp, J.; Meenken, E.; et al. Sources of variability in the effectiveness of winter cover crops for mitigating N leaching. Agric. Ecosyst. Environ. 2016, 220, 226–235. [Google Scholar] [CrossRef]
- Norberg, L.; Aronsson, H. Effects of cover crops sown in autumn on N and P leaching. Soil Use Manag. 2020, 36, 200–211. [Google Scholar] [CrossRef]
- Nevins, C.J.; Lacey, C.; Armstrong, S. The synchrony of cover crop decomposition, enzyme activity, and nitrogen availability in a corn agroecosystem in the Midwest United States. Soil Tillage Res. 2020, 197, 104518. [Google Scholar] [CrossRef]
- Arlauskienė, A.; Maikštėnienė, S. Tarpinių pasėlių įtaka azoto išsiplovimo mažinimui sunkaus priemolio rudžemyje (The use of catch crops for the reduction of nitrogen leaching in a heavy loam Cambisol). Horticulture 2008, 27, 139–153. [Google Scholar]
- Arlauskienė, A.; Maikštėnienė, S. Mineral nitrogen dynamics and nitrate leaching in a catch crop and straw management system. J. Food Agric. Environ. 2012, 10, 1188–1193. [Google Scholar]
- Arlauskienė, A.; Cesevičienė, J.; Velykis, A. Improving mineral nitrogen control by combining catch crops, fertilisation, and straw management in a clay loam soil. Acta Agric. Scand. Sect. B Soil Plant Sci. 2019, 69, 422–431. [Google Scholar] [CrossRef]
- Arlauskienė, A.; Cesevičienė, J.; Šlepetienė, A. Effect of catch crop, straw management and fertilisation on the productivity of field pea and winter wheat crop sequence. Zemdirb. Agric. 2020, 107, 217–226. [Google Scholar] [CrossRef]
- De Neergaard, A.; Hauggaard-Nielsen, H.; Stoumann Jensen, L.; Magid, J. Decomposition of White clover (Trifolium repens) and Ryegrass (Lolium perenne) components: C and N dynamics simulated with the DAISY soil organic matter submodel. Eur. J. Agron. 2002, 16, 43–55. [Google Scholar] [CrossRef]
- Lindedam, J.; Magid, J.; Poulsen, P.; Luxhøi, J. Tissue architecture and soil fertility controls on decomposer communities and decomposition of roots. Soil Biol. Biochem. 2009, 41, 1040–1049. [Google Scholar] [CrossRef]
- Janušauskaitė, D.; Arlauskienė, A.; Maikštėnienė, S. Soil mineral nitrogen and microbial parameters as influenced by catch crops and straw management. Zemdirb. Agric. 2013, 100, 9–18. [Google Scholar] [CrossRef]
- Blanco-Canqui, H. Cover crops and carbon sequestration: Lessons from U.S. studies. Soil Sci. Soc. Am. J. 2022, 86, 501–519. [Google Scholar] [CrossRef]
- Hallama, M.; Pekrun, C.; Lambers, H. Hidden miners—The roles of cover crops and soil microorganisms in phosphorus cycling through agroecosystems. Plant Soil 2019, 434, 7–45. [Google Scholar] [CrossRef]
- Chahal, I.; Van Eerd, L.L. Evaluation of commercial soil health tests using a medium-term cover crop experiment in a humid, temperate climate. Plant Soil 2018, 427, 351–367. [Google Scholar] [CrossRef]
- Ruark, M.D.; Chawner, M.M.; Ballweg, M.J.; Proost, R.T.; Arriaga, F.J.; Stute, J.K. Does cover crop Radish supply nitrogen to corn? Agron. J. 2018, 110, 1513–1522. [Google Scholar] [CrossRef]
- Patel, S.; Sawyer, J.E.; Lundvall, J.P. Can management practices enhance corn productivity in a Rye cover crop system? Agron. J. 2019, 111, 3161–3171. [Google Scholar] [CrossRef]
- Gieske, M.F.; Ackroyd, V.; Baas, D.G.; Mutch, D.R.; Wyse, D.L.; Durgan, B.R. Brassica cover crop effects on nitrogen availability and oat and corn yield. Agron. J. 2016, 108, 151–161. [Google Scholar] [CrossRef]
- Gaskin, J.W.; Cabrera, M.L.; Kissel, D.E.; Hitchcock, R. Using the cover crop N calculator for adaptive nitrogen fertilizer management: A proof of concept. Renew. Agric. Food Syst. 2020, 35, 550–560. [Google Scholar] [CrossRef]
- Saez-Plaza, P.; Navas, M.J.; Wybraniec, S.; Michałowski, T.; Asuero, A.G. An overview of the Kjeldahl method of nitrogen determination. Part II. Sample preparation, working scale, instrumental finish, and quality control. Crit. Rev. Anal. Chem. 2013, 43, 224–272. [Google Scholar] [CrossRef]
- Raudonius, S. Application of statistics in plant and crop research: Important issues. Zemdirb. Agric. 2017, 104, 377–382. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment (E) | Main Crop (MC) | Cover Crop (CC) | Biomass | N | P | K |
|---|---|---|---|---|---|---|
| kg ha−1 DM | ||||||
| Ia | WW | WM | 3141 ± 183 | 72.8 ± 6.4 | 3.7 ± 0.3 | 65.3 ± 3.9 |
| RC | 6050 ± 185 ** | 184.9 ± 12.3 ** | 31.4 ± 0.7 ** | 199.0 ± 5.8 ** | ||
| SB | WM | 4580 ± 131 ** | 111.7 ± 2.6 ** | 6.4 ± 0.7 | 101.0 ± 7.6 ** | |
| RC | 6590 ± 151 ** | 250.8 ± 13.7 ** | 30.3 ± 0.6 ** | 276.1 ± 5.8 ** | ||
| Ib | WW | WM | 1032 ± 190 ** | 16.4 ± 4.4 ** | 2.8 ± 0.5 | 21.2 ± 3.7 ** |
| RC | 348 ± 66 ** | 9.3 ± 1.9 ** | 0.8 ± 0.1 | 7.6 ± 1.5 ** | ||
| SB | WM | 210 ± 21 ** | 5.8 ± 0.2 ** | 0.5 ± 0.1 * | 3.8 ± 0.5 ** | |
| RC | 220 ± 18 ** | 6.3 ± 0.6 ** | 0.4 * ± 0.0 | 7.2 ± 0.7 ** | ||
| Ic | WW | WM | 1030 ± 188 ** | 20.2 ± 4.1 ** | 2.0 ± 0.3 | 21.3 ± 3.8 ** |
| RC | 3020 ± 740 | 86.2 ± 22.2 | 11.4 ± 3.3 ** | 82.5 ± 19.5 | ||
| SB | WM | 1782 ± 70 ** | 46.9 ± 2.7 | 3.6 ± 0.1 | 48.7 ± 2.7 | |
| RC | 3275 ± 57 | 117.2 ± 2.3 ** | 10.5 ± 0.2 ** | 98.6 ± 1.6 ** | ||
| Probability (p) level of the factors and their interaction | ||||||
| E (2) | <0.001 | <0.001 | <0.001 | <0.001 | ||
| MC (1) | <0.05 | <0.001 | ns | <0.001 | ||
| CC (1) | <0.001 | <0.001 | <0.001 | <0.001 | ||
| ExMC (2) | <0.01 | 0.001 | ns | <0.001 | ||
| ExCC (2) | <0.001 | <0.001 | <0.001 | <0.001 | ||
| MCxCC (1) | ns | ns | ns | ns | ||
| ExMCxCC (2) | ns | ns | ns | <0.05 | ||
| Experiment (E) | Straw Use (SU) | Cover Crops and Sowing Methods (CC) | Biomass | N | P | K |
|---|---|---|---|---|---|---|
| kg ha−1 DM | ||||||
| IIa | WS | WMps | 1150 ± 68 | 38.9 ± 2.4 | 6.7 ± 0.5 | 34.8 ± 2.1 |
| WMus | 1040 ± 272 | 38.7 ± 10.1 | 6.5 ± 1.6 | 28.8 ± 7.7 | ||
| RCus | 2100 ± 171 ** | 60.4 ± 3.5 ** | 6.9 ± 0.5 | 50.0 ± 4.2 * | ||
| S | WMps | 1640 ± 58 * | 53.1 ± 1.7 * | 8.4 ± 0.3 | 51.3 ± 1.9 * | |
| WMus | 1140 ± 79 | 34.0 ± 2.3 | 4.7 ± 0.3 * | 26.0 ± 1.7 | ||
| RCus | 2210 ± 98 ** | 62.7 ± 2.3 ** | 7.3 ± 0.6 | 44.8 ± 1.8 | ||
| IIb | WS | WMps | 3680 ± 83 ** | 103.7 ± 0.3 ** | 15.4 ± 0.5 ** | 149.8 ± 4.7 ** |
| WMus | 5070 ± 87 ** | 152.6 ± 3.5 ** | 19.2 ± 0.3 ** | 168.8 ± 1.2 ** | ||
| NLps | 1990 ± 50 ** | 58.3 ± 1.7 ** | 6.6 ± 0.4 | 49.8 ± 1.3 * | ||
| S | WMps | 3920 ± 72 ** | 152.5 ± 3.4 ** | 16.2 ± 0.4 ** | 142.7 ± 3.3 ** | |
| WMus | 5119 ± 105 ** | 201.8 ± 5.8 ** | 20.2 ± 0.4 ** | 189.0 ± 6.7 ** | ||
| NLps | 2270 ± 324 ** | 66.6 ± 9.2 ** | 7.2 ± 1.1 | 50.4 ± 8.4 * | ||
| Probability (p) level of the factors and their interaction | ||||||
| E (1) | <0.001 | <0.001 | <0.001 | <0.001 | ||
| SU (1) | <0.05 | <0.001 | ns | ns | ||
| CC (2) | <0.001 | <0.001 | <0.001 | <0.001 | ||
| ExSU (1) | ns | <0.001 | ns | ns | ||
| ExCC (2) | <0.001 | <0.001 | <0.001 | <0.001 | ||
| SUxCC (2) | ns | <0.01 | ns | ns | ||
| ExSUxCC (2) | ns | <0.01 | ns | <0.01 | ||
| Cover Crops (CCs) | Straw Use (SU) | Fertilization (F) | Biomass kg ha−1 DM | N | P | K |
|---|---|---|---|---|---|---|
| % DM | ||||||
| WM | WS | UF | 524 ± 63 | 2.04 ± 0.04 | 0.45 ± 0.01 | 3.91 ± 0.06 |
| SF | 460 ± 103 | 2.27 ± 0.09 | 0.48 ± 0.03 | 3.81 ± 0.17 | ||
| IF | 397 ± 36 | 2.37 ± 0.11 | 0.48 ± 0.00 | 4.12 ± 0.03 | ||
| S | UF | 858 ± 144 | 1.95 ± 0.13 | 0.44 ± 0.02 | 3.96 ± 016 | |
| SF | 1107 ± 133 * | 1.98 ± 0.09 | 0.42 ± 0.01 | 3.95 ± 0.10 | ||
| IF | 773 ± 122 | 2.17 ± 0.21 | 0.43 ± 0.02 | 4.18 ± 0.12 | ||
| RC | WS | UF | 2445 ± 527 ** | 3.42 ± 0.16 ** | 0.32 ± 0.04 ** | 3.90 ± 0.26 |
| SF | 1131 ± 146 * | 3.48 ± 0.17 ** | 0.31 ± 0.01 ** | 3.75 ± 0.07 | ||
| IF | 873 ± 193 | 3.77 ± 0.13 ** | 0.31 ± 0.00 ** | 4.14 ± 0.11 | ||
| S | UF | 2263 ± 200 ** | 3.61 ± 0.06 ** | 0.30 ± 0.00 ** | 3.99 ± 0.17 | |
| SF | 458 ± 67 | 3.93 ± 0.01 ** | 0.34 ± 0.00 ** | 4.06 ± 0.13 | ||
| IF | 455 ± 60 | 3.91 ± 0.12 ** | 0.32 ± 0.01 ** | 4.21 ± 0.08 | ||
| Probability (p) level of the factors and their interaction | ||||||
| CC (1) | <0.001 | <0.001 | <0.001 | ns | ||
| SU (1) | ns | ns | ns | ns | ||
| F (2) | <0.001 | <0.01 | ns | <0.05 | ||
| CCxSU (1) | 0.001 | <0.01 | <0.05 | ns | ||
| CCxF (2) | <0.001 | ns | ns | ns | ||
| SUxF (2) | ns | ns | ns | ns | ||
| CCxSUxF (2) | ns | ns | ns | ns | ||
| Experiment (E) | Main Crop (MC) | Cover Crop (CC) | N | P | K |
|---|---|---|---|---|---|
| kg ha−1 | |||||
| Ia | WW | WCC | 74.4 ± 0.9 a | 17.2 ± 0.3 a | 81.5 ± 1.0 a |
| WM | 79.4 ± 1.4 b | 19.2 ± 0.3 b | 80.8 ± 1.0 a | ||
| RC | 83.6 ± 1.6 c | 19.0 ± 0.5 b | 89.3 ± 1.0 b | ||
| SB | WCC | 96.4 ± 1.1 d | 20.9 ± 0.4 c | 86.0 ± 0.3 ab | |
| WM | 105.5 ± 0.8 e | 22.0 ± 0.1 d | 90.3 ± 0.2 b | ||
| RC | 115.0 ± 0.5 f | 22.8 ± 0.1 d | 94.4 ± 0.3 c | ||
| Ib | WW | WCC | 102.5 ± 0.9 b | 21.1 ± 1.3 b | 97.5 ± 0.4 ab |
| WM | 97.1 ± 1.5 ab | 19.4 ± 0.4 a | 94.7 ± 0.6 a | ||
| RC | 100.1 ± 1.2 b | 20.4 ± 0.1 a | 99.0 ± 0.5 b | ||
| SB | WCC | 95.4 ± 0.8 a | 19.3 ± 0.1 a | 94.8 ± 1.0 a | |
| WM | 104.4 ± 0.1 b | 21.4 ± 0.1 b | 97.3 ± 0.2 ab | ||
| RC | 98.6 ± 1.9 ab | 19.9 ± 0.9 a | 97.3 ± 0.2 ab | ||
| Ic | WW | WCC | 108.0 ± 1.0 ab | 25.5 ± 0.3 b | 98.0 ± 0.5 a |
| WM | 112.4 ± 1.3 b | 26.5 ± 0.4 b | 99.5 ± 0.4 b | ||
| RC | 121.0 ± 0.9 c | 27.7 ± 0.4 c | 102.6 ± 1.0 c | ||
| SB | WCC | 101.5 ± 0.7 a | 23.5 ± 0.7 a | 98.3 ± 0.5 ab | |
| WM | 103.7 ± 3.6 a | 24.1 ± 0.9 a | 98.3 ± 1.0 ab | ||
| RC | 116.2 ± 0.2 b | 25.7 ± 0.3 b | 104.3 ± 0.5 c | ||
| Probability (p) level of the factors and their interaction | |||||
| E (2) | <0.001 | <0.001 | <0.001 | ||
| MC (1) | <0.001 | ns | <0.001 | ||
| CC (2) | <0.001 | <0.001 | <0.001 | ||
| ExMC (2) | <0.001 | <0.001 | <0.001 | ||
| ExCC (4) | <0.001 | <0.05 | <0.001 | ||
| MCxCC (2) | <0.001 | ns | <0.001 | ||
| ExMCxCC (4) | <0.001 | <0.05 | <0.001 | ||
| Experiment (E) | Straw Use (SU) | Cover Crop (CC) and Sowing Methods | N | P | K |
|---|---|---|---|---|---|
| kg ha−1 | |||||
| IIa | WS | WCC | 74.2 ± 0.9 a | 11.7 ± 0.4 b | 54.5 ± 0.9 d |
| WMps | 86.1 ± 1.1 c | 12.9 ± 0.2 d | 59.7 ± 0.9 e | ||
| WMus | 84.5 ± 4.2 c | 12.7 ± 0.7 cd | 54.2 ± 0.3 cd | ||
| RCus | 77.6 ± 3.4 a | 12.0 ± 0.6 bcd | 65.2 ± 1.1 f | ||
| S | WCC | 76.0 ± 5.2 a | 11.4 ± 0.3 b | 43.5 ± 0.4 a | |
| WMps | 78.2 ± 1.4 a | 11.5 ± 0.5 b | 52.2 ± 0.9 c | ||
| WMus | 73.9 ± 1.0 a | 10.2 ± 0.5 a | 47.6 ± 1.3 b | ||
| RCus | 75.9 ± 1.8 a | 11.6 ± 0.4 b | 45.7 ± 0.5 b | ||
| IIb | WS | WCC | 92.7 ± 1.7 ab | 15.8 ± 0.2 a | 76.9 ± 1.2 a |
| WMps | 100.4 ± 2.9 cde | 16.9 ± 0.5 bc | 93.8 ± 1.1 cd | ||
| WMus | 103.0 ± 2.6 e | 17.0 ± 0.3 c | 88.7 ± 0.4 bcd | ||
| RCus | 98.7 ± 1.8 cde | 16.5 ± 0.4 abc | 93.9 ± 3.5 cd | ||
| S | WCC | 91.0 ± 0.9 a | 16.2 ± 0.3 abc | 83.0 ± 2.6 ab | |
| WMps | 98.2 ± 0.9 cde | 16.4 ± 0.3 abc | 93.0 ± 3.7 cd | ||
| WMus | 98.9 ± 5.3 cde | 16.9 ± 0.6 bc | 84.7 ± 0.9 b | ||
| RCus | 96.4 ± 1.1 bc | 16.3 ± 0.2 abc | 93.9 ± 2.2 d | ||
| Probability (p) level of the factors and their interaction | |||||
| E (1) | <0.001 | <0.001 | <0.001 | ||
| SU (1) | <0.05 | <0.01 | <0.001 | ||
| CC (3) | <0.01 | ns | <0.001 | ||
| ExSU (1) | ns | <0.05 | <0.001 | ||
| ExCC (3) | ns | ns | <0.05 | ||
| SUxCC (3) | ns | ns | <0.05 | ||
| ExSUxCC (3) | ns | ns | <0.01 | ||
| Month | 2001 | 2002 | 2003 | 2004 | 2006 | 2013 | SCN | |
|---|---|---|---|---|---|---|---|---|
| April | T, °C | 7.6 | 6.9 | 4.2 | 6.4 | 6.7 | 4.6 | 6.2 |
| P, mm | 36.4 | 28.4 | 37.1 | 8.3 | 23.3 | 36.9 | 37.4 | |
| May | T, °C | 10.5 | 14.6 | 12.3 | 10.3 | 12.2 | 16.2 | 12.3 |
| P, mm | 35.4 | 10.9 | 56.1 | 25.2 | 32.5 | 73.4 | 45.6 | |
| Junie | T, °C | 13.1 | 16.4 | 14.1 | 13.8 | 16.3 | 18.6 | 15.6 |
| P, mm | 147.9 | 81.1 | 57.2 | 64.0 | 6.8 | 44.4 | 59.4 | |
| July | T, °C | 19.6 | 19.8 | 19.7 | 16.1 | 20.9 | 18.9 | 17.2 |
| P, mm | 172.5 | 41.5 | 45.1 | 68.0 | 28.6 | 53.0 | 69.2 | |
| Main period | T, °C | 12.7 | 14.4 | 12.6 | 11.7 | 14.0 | 14.6 | 12.8 |
| P, mm | 392.2 | 161.9 | 195.5 | 165.5 | 91.2 | 207.7 | 211.6 | |
| August | T, °C | 16.2 | 18.1 | 16.5 | 17.4 | 18.7 | 18.6 | 17.1 |
| P, mm | 52.8 | 9.6 | 50.6 | 56.0 | 139.8 | 38.8 | 67.9 | |
| September | T, °C | 11.1 | 12,4 | 12.2 | 12.3 | 15.0 | 12.4 | 12.0 |
| P, mm | 59.8 | 27.5 | 29.5 | 69.8 | 48.2 | 81.1 | 57.9 | |
| October | T, °C | 9.9 | 2.4 | 6.1 | 4.4 | 9.4 | 6.3 | 6.3 |
| Until 15 | P, mm | 22.7 | 37.5 | 37.2 | 9.0 | 48.5 | 0.2 | 45.5 |
| Post-harvest period | T, °C | 12.4 | 11.0 | 11.6 | 11.4 | 14.4 | 12.4 | 11.8 |
| P, mm | 135.3 | 74.6 | 117.3 | 134.8 | 236.5 | 120.1 | 161.3 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arlauskienė, A.; Šarūnaitė, L. Cover Crop Yield, Nutrient Storage and Release under Different Cropping Technologies in the Sustainable Agrosystems. Plants 2023, 12, 2966. https://doi.org/10.3390/plants12162966
Arlauskienė A, Šarūnaitė L. Cover Crop Yield, Nutrient Storage and Release under Different Cropping Technologies in the Sustainable Agrosystems. Plants. 2023; 12(16):2966. https://doi.org/10.3390/plants12162966
Chicago/Turabian StyleArlauskienė, Aušra, and Lina Šarūnaitė. 2023. "Cover Crop Yield, Nutrient Storage and Release under Different Cropping Technologies in the Sustainable Agrosystems" Plants 12, no. 16: 2966. https://doi.org/10.3390/plants12162966
APA StyleArlauskienė, A., & Šarūnaitė, L. (2023). Cover Crop Yield, Nutrient Storage and Release under Different Cropping Technologies in the Sustainable Agrosystems. Plants, 12(16), 2966. https://doi.org/10.3390/plants12162966