
Multitemporal Land Use and Cover Analysis Coupled with Climatic Change Scenarios to Protect the Endangered Taxon Asphodelus bento-rainhae subsp. bento-rainhae

 ,
,  ,
,  ,
,  and
and
Abstract
1. Introduction
2. Results
2.1. Variable Importance and Model Accuracy
2.2. Current and Future Potential Distribution
2.3. Land Use and Land Cover (LULC) Change Analysis
3. Discussion
4. Materials and Methods
4.1. Study Area
4.2. Species Occurrence Data
4.3. Environmental Data
4.4. Land Use and Land Cover Data
4.5. Modeling
4.6. Land Use and Land Cover (LULC) Change Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Classification | Description | Study Area (%) | Occurrences (%) |
|---|---|---|---|---|
| AS | AS | Social Area | 2.3 | 0.0 |
| CM1 | CM.dy.cr(ha) | Chromi-Dystric Cambisols | 15.8 | 35.3 |
| CM2 | CM.dy.ha | Hapli-Dystric Cambisols | 5.0 | 0.0 |
| CM3 | CM.dy.ha(cr) | Hapli-Dystric Cambisols | 10.9 | 0.0 |
| FL | FL.dy | Distric Fluvisols | 1.3 | 0.0 |
| R | R | Rocky Outcrops | 1.4 | 0.0 |
| RG1 | RG.ai.dy | Dystri-Aric Regosols | 9.7 | 1.1 |
| RG2 | RG.lep.dy | Dystri-Epileptic Regosols | 27.7 | 56.8 |
| RG3 | RG.lep.dy(sk) | Dystri-Epileptic Regosols | 11.4 | 0.0 |
| UM | UM.lep.hu | Humic Epileptic Umbrisols | 14.5 | 6.8 |
| Area | LULC Classes | Description |
|---|---|---|
| SA/OA | Artificialized Territories | Urban fabric; industrial, commercial, and transport units; mine, dump, and construction sites; artificial non-agricultural vegetated areas. |
| SA | Agriculture | Arable land, lands under a rotation system used for annually harvested plants and fallow lands, which are rain-fed or irrigated and for permanent crops. Areas of annual crops associated with permanent crops on the same parcel, annual crops cultivated under trees, areas of annual crops, meadows and/or crops and pastures mixed with natural vegetation. |
| OA | Agriculture | Agriculture (SA) excluding permanent crops. |
| Vineyards | Permanent crops, lands not under a rotation system, with vineyards. | |
| Fruit orchards | Permanent crops, lands not under a rotation system, with fruit orchards, trees or shrubs of one or more species intended for fruit production. | |
| Olive groves | Permanent croplands not under a rotation system, with olive groves. | |
| SA/OA | Grasslands | Pasture lands that are permanently used (at least 5 years) for fodder production. Includes natural or sown herbaceous species, unimproved or lightly improved meadows and grazed or mechanically harvested meadows. |
| SA | Agroforestry areas | Landscapes in which crops and pastures are intimately mixed with Mediterranean forest (Quercus suber, Quercus rotundifolia and other oaks) and natural vegetation or natural areas. |
| Forests | All forest species resulting from natural regeneration, seeding or planting. | |
| OA | Cork oak | Quercus suber forest. |
| Other oaks | Quercus pyrenaica, Quercus robur, Quercus faginea forests, and other oaks but not including Quercus suber and Quercus rotundifolia. | |
| Chestnut | Castanea sativa forest. | |
| Eucalyptus | Eucalyptus spp. forest. | |
| Invasive species | Invasive forest (e.g., Acacia dealbata, Ailanthus altissima). | |
| Deciduous forests | Broadleaved forests (e.g., Salix spp., Populus spp., Platanus spp., Alnus glutinosa, and Juglans regia) for wood purposes. | |
| Pine woods | Pinus pinaster or Pinus pinaster and Pinus pinea mixed stands. | |
| SA/OA | Shrublands | Natural areas of spontaneous vegetation, sparsely with shrubs or very dense. |
| SA/OA | Sparsely vegetated areas | Natural areas with little or no vegetation, including recently burnt areas and bare rock. |
| SA | Water bodies | Inland wetlands. |
References
- Grímsson, F.; Bouchal, J.M.; Xafis, A.; Zetter, R. Combined LM and SEM study of the middle Miocene (Sarmatian) palynoflora from the Lavanttal Basin, Austria: Part V. Magnoliophyta 3—Myrtales to Ericales. Grana 2020, 59, 127–193. [Google Scholar] [CrossRef] [PubMed]
- Barredo, J.I.; Caudullo, G.; Dosio, A. Mediterranean habitat loss under future climate conditions: Assessing impacts on the Natura 2000 protected area network. Appl. Geogr. 2016, 75, 83–92. [Google Scholar] [CrossRef]
- Peñuelas, J.; Sardans, J.; Filella, I.; Estiarte, M.; Llusià, J.; Ogaya, R.; Carnicer, J.; Bartrons, M.; Rivas-Ubach, A.; Grau, O.; et al. Impacts of Global Change on Mediterranean Forests and Their Services. Forests 2017, 8, 463. [Google Scholar] [CrossRef]
- Carrión, J.S.; Van Geel, B. Fine-resolution Upper Weichselian and Holocene palynological record from Navarrés (Valencia, Spain) and a discussion about factors of Mediterranean forest succession. Rev. Palaeobot. Palynol. 1999, 106, 209–236. [Google Scholar] [CrossRef]
- Rick, T.; Ontiveros, M.Á.C.; Jerardino, A.; Mariotti, A.; Méndez, C.; Williams, A.N. Human-environmental interactions in Mediterranean climate regions from the Pleistocene to the Anthropocene. Anthropocene 2020, 31, 100253. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Pinto da Silva, A.R. Asphodelus bento-rainhae P. Silva, sp. Nov. Agron. Lusit. 1956, 18, 20–21. [Google Scholar]
- Malmir, M.; Serrano, R.; Caniça, M.; Silva-Lima, B.; Silva, O. A Comprehensive Review on the Medicinal Plants from the Genus Asphodelus. Plants 2018, 7, 20. [Google Scholar] [CrossRef]
- Raunkiær, C. The Life Forms of Plants and Statistical Plant Geography Being the Collected Papers of C. Raunkiaer; Oxford at the Clarendon Press: Oxford, UK, 1934. [Google Scholar]
- Franco, J.A.; Afonso, M.L.R. Nova Flora de Portugal (Continente e Açores); Fascículo I (Alismatacea—Iridacea); Escolar Editora: Lisboa, Portugal, 1994; Volume III. [Google Scholar]
- Díaz-Lifante, Z.; Valdés, B. Revisión del género Asphodelus L. (Asphodelaceae) en el Mediterráneo Occidental. Boissiera 1996, 52, 1–189. [Google Scholar]
- ICNF. O Plano Setorial da Rede Natura 2000. Available online: https://www.icnf.pt/biodiversidade/natura2000/planosetorial (accessed on 11 April 2023).
- Travassos, J.S. Serra da Gardunha, que História? ADRACES: Fundão, Portugal, 1999. [Google Scholar]
- Delgado, F.M.G. Conservação e Valorização de Asphodelus Bento-rainhae P. Silva e Lavandula luisieri (Rozeira) Rivas-Martínez da Beira Interior. Ph.D. Thesis, Instituto Superior de Agronomia, Universidade Técnica de Lisboa, Lisboa, Portugal, 2010. [Google Scholar]
- Ribeiro, S.; Delgado, F.; Espírito-Santo, M. Comunidades de Asphodelus bento-rainhae P. Silva: Diversidade, ecologia e dinâmica serial. Silva Lusit. 2012, 20, 138–143. [Google Scholar]
- Pinto, M.J.; Silva, J.P. How much habitat connectivity affects the regional trend. In Proceedings of the V Congresso da Sociedade Portuguesa de Ecologia/1º Congresso Ibérico de Ecologia, Santiago de Compostela, Spain, 25–28 September2001. [Google Scholar]
- Cotrim, H.M.; Silva, J.P.; Fay, M.F.; Chase, M.W. Analysis of genetic diversity in Asphodelus bento-rainhae P. In Proceedings of the II Congresso Internacional Sobre a Situação da Rede Natura 2000 nos Países Mediterrâneos, Lisboa, Portugal, 6–8 December 2002. [Google Scholar]
- Díaz-Lífante, Z. Reproductive biology of Asphodelus aestivus (Asphodelaceae). Plant Syst. Evol. 1996, 200, 177–191. [Google Scholar] [CrossRef]
- ADESGAR. Asphodelus Bento-Rainhae—Medidas de Conservação e Gestão; Project LIFE NAT/P/005229, Final Report; ADESGAR: Fundão, Portugal, 2004. [Google Scholar]
- Harper, J.L. Population Biology of Plants; Academic Press: Cambridge, MA, USA, 1977. [Google Scholar]
- Mayer, A.M.; Poljakoff-Mayber, A. The Germination of Seeds, 4th ed.; Pergamon Press: Oxford, New York, USA, 1989. [Google Scholar]
- Odoemena, C.S. Breaking of seed coat dormancy in a medicinal plant Tetrapleura tetraptera (Schum & Thonn). J. Agric. Sci. 1988, 111, 393–394. [Google Scholar] [CrossRef]
- Graaff, J.L.; Van Staden, J. The effect of different chemical and physical treatments on seed coat structure and seed germination of Sesbania species. Z. Für Pflanzenphysiol. 1983, 112, 221–230. [Google Scholar] [CrossRef]
- Malmir, M.; Serrano, R.; Lima, K.; Duarte, M.P.; Moreira da Silva, I.; Silva Lima, B.; Caniça, M.; Silva, O. Monographic Quality Parameters and Genotoxicity Assessment of Asphodelus bento-rainhae and Asphodelus macrocarpus Root Tubers as Herbal Medicines. Plants 2022, 11, 3173. [Google Scholar] [CrossRef]
- Malmir, M.; Lima, K.; Póvoas Camões, S.; Manageiro, V.; Duarte, M.P.; Paiva Miranda, J.; Serrano, R.; Moreira da Silva, I.; Silva Lima, B.; Caniça, M.; et al. Identification of Marker Compounds and In Vitro Toxicity Evaluation of Two Portuguese Asphodelus Leaf Extracts. Molecules 2023, 28, 2372. [Google Scholar] [CrossRef] [PubMed]
- Peksel, A. Evaluation of antioxidant and antifungal potential of Asphodelus aestivus Brot. growing in Turkey. J. Med. Plants Res. 2012, 6, 253–265. [Google Scholar] [CrossRef]
- Caldas, F.B.; Moreno Saiz, J.C. Asphodelus bento-rainhae. Available online: https://doi.org/10.2305/IUCN.UK.2011-1.RLTS.T162190A5555926.en (accessed on 11 April 2023).
- Carapeto, A.; Francisco, A.; Pereira, P.; Porto, M. Lista Vermelha da Flora Vascular de Portugal Continental; Sociedade Portuguesa de Botânica, Associação Portuguesa de Ciência da Vegetação—PHYTOS, Instituto da Conservação da Natureza e das Florestas, IP e Câmara Municipal de Lisboa: Lisboa, Portugal, 2020; Volume 7, p. 374. [Google Scholar]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Valavi, R.; Guillera-Arroita, G.; Lahoz-Monfort, J.J.; Elith, J. Predictive performance of presence-only species distribution models: A benchmark study with reproducible code. Ecol. Monogr. 2022, 92, e01486. [Google Scholar] [CrossRef]
- Almeida, A.M.; Martins, M.J.; Campagnolo, M.L.; Fernandez, P.; Albuquerque, T.; Gerassis, S.; Gonçalves, J.C.; Ribeiro, M.M. Prediction scenarios of past, present, and future environmental suitability for the Mediterranean species Arbutus unedo L. Sci. Rep. 2022, 12, 84. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Fernández, A.J.; Fernández-Ceular, Á.; Alcaraz-Segura, D.; Ballesteros, M.; Peñas, J. The Fate of Endemic Species Specialized in Island Habitat under Climate Change in a Mediterranean High Mountain. Plants 2022, 11, 3193. [Google Scholar] [CrossRef] [PubMed]
- Morales, N.S.; Fernández, I.C. Land-Cover Classification Using MaxEnt: Can We Trust in Model Quality Metrics for Estimating Classification Accuracy? Entropy 2020, 22, 342. [Google Scholar] [CrossRef]
- Esteves, M.L.E. Contribuição Para o Estudo da Ecologia e da Conservação de Asphodelus bento-rainhae P. Silva. Master’s Thesis, Universidade dos Açores, Castelo Branco, Portugal, 2005. [Google Scholar]
- Quinta-Nova, L.; Roque, N.; Ribeiro, S. Habitat suitability modelling of Asphodelus bento-rainhae P. Silva using spatial multicriteria analysis. In Proceedings of the Livro de atas do III Congresso de Ciência e Desenvolvimento dos Açores & 8.º Congresso de Gestão e Conservação da Natureza, Angra do Heroísmo, Portugal, 19–21 June 2015. [Google Scholar]
- Meinshausen, M.; Nicholls, Z.R.J.; Lewis, J.; Gidden, M.J.; Vogel, E.; Freund, M.; Beyerle, U.; Gessner, C.; Nauels, A.; Bauer, N.; et al. The shared socio-economic pathway (SSP) greenhouse gas concentrations and their extensions to 2500. Geosci. Model Dev. 2020, 13, 3571–3605. [Google Scholar] [CrossRef]
- Van Meter, K.J.; Basu, N.B.; Veenstra, J.J.; Burras, C.L. The nitrogen legacy: Emerging evidence of nitrogen accumulation in anthropogenic landscapes. Environ. Res. Lett. 2016, 11, 035014. [Google Scholar] [CrossRef]
- Ferreira, M.R.; Almeida, A.M.; Quintela-Sabarís, C.; Roque, N.; Fernandez, P.; Ribeiro, M.M. The role of littoral cliffs in the niche delimitation on a microendemic plant facing climate change. PLoS ONE 2021, 16, e0258976. [Google Scholar] [CrossRef]
- Parducci, L.; Szmidt, A.E.; Ribeiro, M.M.; Drouzas, A.D. Taxonomic position and origin of the endemic sicilian fir Abies nebrodensis (Lojac.) Mattei based on allozyme analysis. For. Genet. 2001, 8, 119–127. [Google Scholar]
- Coelho, N.; Gonçalves, S.; Romano, A. Endemic Plant Species Conservation: Biotechnological Approaches. Plants 2020, 9, 345. [Google Scholar] [CrossRef]
- Draper, D.; Marques, I.; Graell, A.R.; Costa, F.; Martins-Loução, M.A. Conservação de Recursos Genéticos–O Banco de Sementes “António Luís Belo Correia”. In Curso Avançado Sobre “Métodos de Conservação a Longo Prazo de Recursos Fitogenéticos: Conservação Pelo Frio; Museu Nacional de História Natural: Lisboa, Portugal, 2004; pp. 3–7. [Google Scholar]
- ICNF. Paisagem Protegida Regional da Serra da Gardunha. Available online: https://www.icnf.pt/conservacao/paisagensprotegidas/ppregionaldaserradagardunha (accessed on 11 April 2023).
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- EC-Earth. EC-Earth-Consortium EC-Earth3-Veg Model Output Prepared for CMIP6 ScenarioMIP; Earth System Grid Federation 2019. Available online: https://doi.org/10.22033/ESGF/CMIP6.727 (accessed on 24 February 2023).
- DGADR. Sistema Nacional de Informação do Solo. Available online: https://snisolos.dgadr.gov.pt/ (accessed on 24 February 2023).
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species Distribution Models: Ecological Explanation and Prediction Across Space and Time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Leroy, B.; Meynard, C.N.; Bellard, C.; Courchamp, F. Virtualspecies, an R package to generate virtual species distributions. Ecography 2016, 39, 599–607. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Sequeira, A.C.; Nunes, L.; Caetano, M.; Marcelino, F.; Castro Rego, F. Mapa Agrícola e Florestal de Portugal Continental 1951–1980 (MAF1951-80); Instituto Superior de Agronomia: Lisboa, Portugal, 2022. [Google Scholar]
- DGT. Carta de Uso e Ocupação do Solo. Available online: https://snig.dgterritorio.gov.pt/rndg/srv/por/catalog.search#/search?resultType=details&sortBy=referenceDateOrd&anysnig=COS&fast=index&from=1&to=20 (accessed on 24 February 2023).
- DGT. Especificações Técnicas da Carta de Uso e Ocupação do Solo (COS) de Portugal Continental para 1995, 2007, 2010, 2015 e 2018; Direção-Geral do Território: Lisboa, Portugal, 2023. [Google Scholar]
- Phillips, S.J.; Dudík, M. Schapire RE: Maxent Software for Modeling Species Niches and Distributions (Version 3.4.1). Available online: https://www.scirp.org/(S(czeh2tfqw2orz553k1w0r45))/reference/referencespapers.aspx?referenceid=2490081 (accessed on 11 January 2023).
- Phillips, S.J.; Anderson, R.P.; DudÍk, M.; Schapire, R.E.; Blair, M.E. Opening the black box: An open-source release of Maxent. Ecography 2017, 40, 887–893. [Google Scholar] [CrossRef]
- Phillips, S.J. A Brief Tutorial on Maxent. Available online: http://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 11 January 2023).
- Phillips, S.J.; Dudík, M.; Schapire, R.E. A maximum entropy approach to species distribution modeling. In Proceedings of the Twenty-First International Conference on Machine Learning, Banff, AB, Canada, 4–8 July 2004; pp. 655–662. [Google Scholar]
- Radosavljevic, A.; Anderson, R.P. Making better Maxent models of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]









| Variable | Percent Contribution | Permutation Importance | Training Gain without | Training Gain with Only |
|---|---|---|---|---|
| BIO4 | 68.07 | 59.69 | 1.06 | 0.77 |
| BIO7 | 9.83 | 11.20 | 1.18 | 0.10 |
| Aspect | 7.20 | 1.84 | 1.15 | 0.28 |
| Soils | 5.90 | 0.00 | 1.19 | 0.43 |
| BIO2 | 4.96 | 22.46 | 1.16 | 0.44 |
| BIO15 | 2.76 | 4.08 | 1.18 | 0.35 |
| BIO12 | 1.28 | 0.73 | 1.19 | 0.53 |
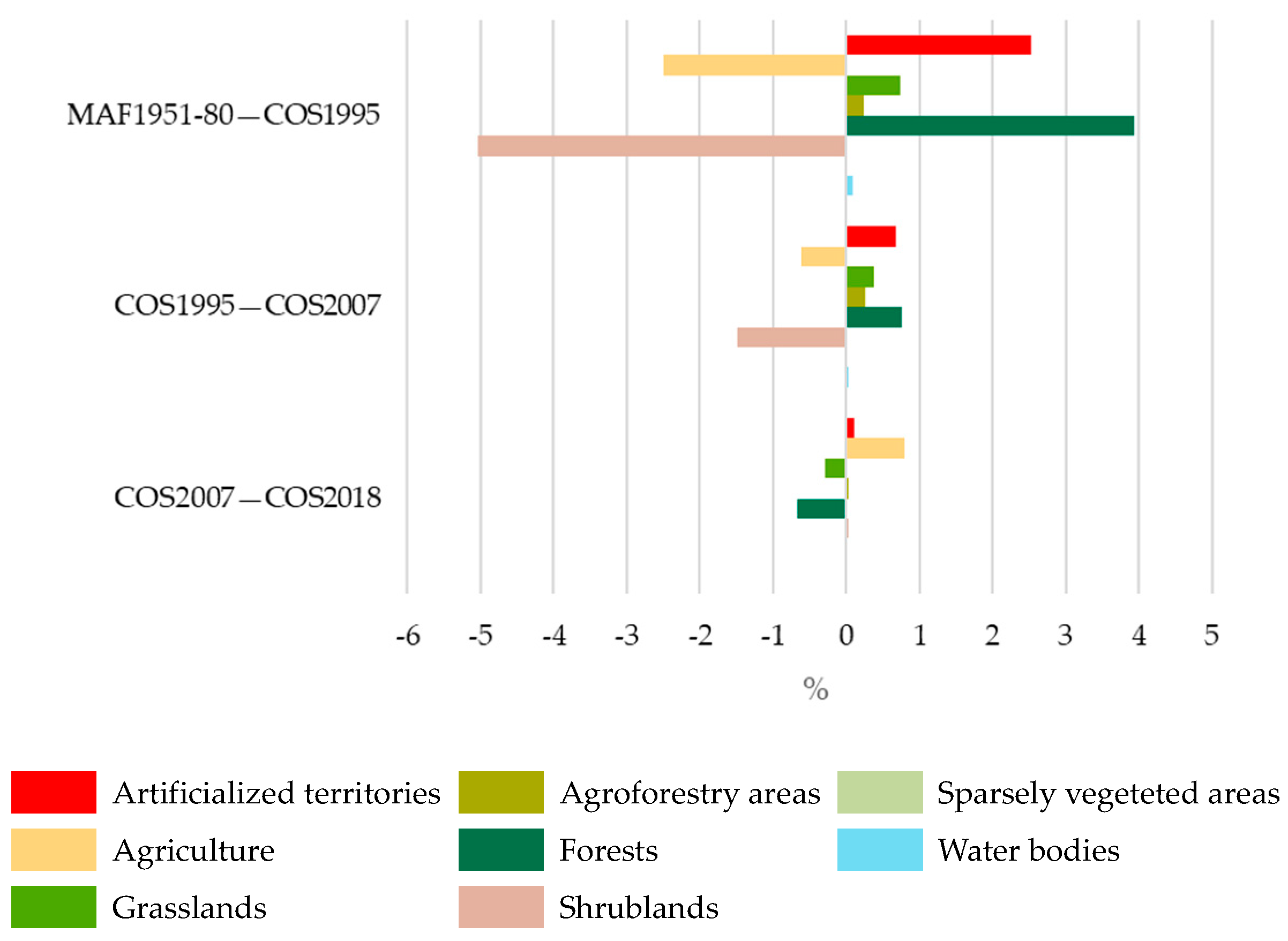
| LULC Class | MAF1951–1980 | COS1995 | COS2007 | COS2018 |
|---|---|---|---|---|
| Artificialized territories | 2.0 | 4.5 | 5.2 | 5.3 |
| Agriculture | 46.8 | 44.3 | 43.6 | 44.5 |
| Grasslands | 0.0 | 0.7 | 1.1 | 0.8 |
| Agroforestry areas | 0.4 | 0.6 | 0.9 | 0.9 |
| Forests | 39.7 | 43.7 | 44.5 | 43.8 |
| Shrublands | 11.1 | 6.1 | 4.6 | 4.6 |
| Sparsely vegetated areas | 0.0 | 0.0 | 0.0 | 0.0 |
| Water bodies | 0.0 | 0.1 | 0.1 | 0.1 |
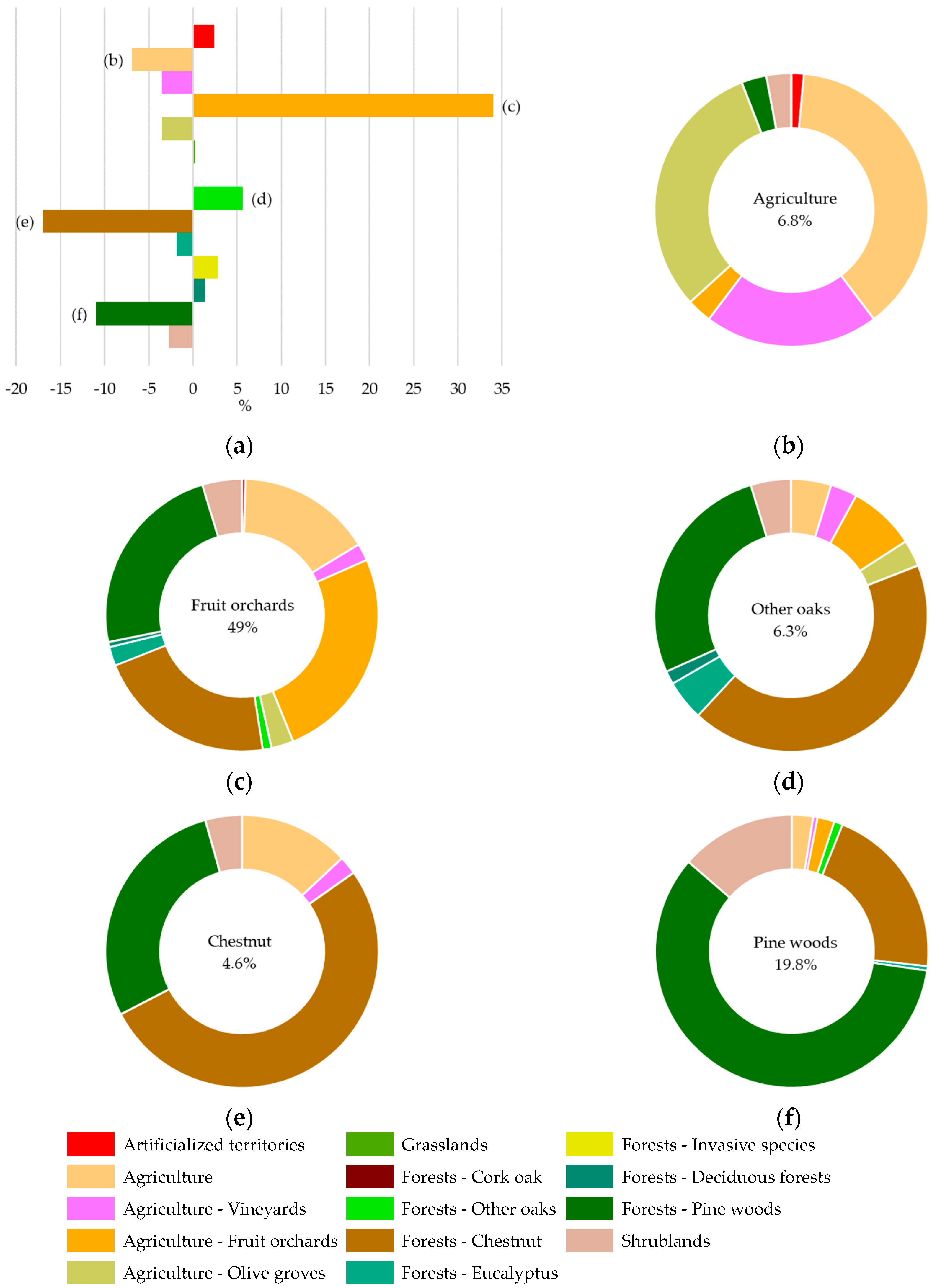
| LULC Class | MAF1951–1980 | COS1995 | COS2007 | COS2018 |
|---|---|---|---|---|
| Artificialized territories | 1.0 | 2.9 | 3.5 | 3.4 |
| Agriculture | 13.7 | 10.5 | 7.0 | 6.8 |
| Agriculture–Vineyards | 3.6 | 0.1 | 0.1 | 0.1 |
| Agriculture–Fruit orchards | 15.0 | 44.8 | 48.5 | 49.0 |
| Agriculture–Olive groves | 4.8 | 1.1 | 1.3 | 1.3 |
| Grasslands | 0.0 | 0.1 | 0.3 | 0.2 |
| Forests–Cork oak | 0.1 | 0.0 | 0.0 | 0.0 |
| Forests–Other oaks | 0.7 | 6.8 | 6.6 | 6.3 |
| Forests–Chestnut | 21.5 | 4.5 | 4.8 | 4.6 |
| Forests–Eucalyptus | 2.1 | 0.2 | 0.2 | 0.2 |
| Forests–Invasive species | 0.0 | 0.0 | 2.9 | 2.9 |
| Forests–Deciduous forests | 0.4 | 5.5 | 1.8 | 1.8 |
| Forests–Pine woods | 30.8 | 19.5 | 20.2 | 19.8 |
| Shrublands | 6.3 | 4.0 | 2.8 | 3.6 |
| Symbol | Variable | Study Area | Species Occurrences | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Min | Max | Mean | Std | Min | Max | Mean | Std | ||
| BIO1 | Annual mean temperature (°C) | 12.0 | 14.7 | 14.0 | 0.5 | 12.9 | 14.3 | 13.7 | 0.3 |
| BIO2 | Mean diurnal range (°C) | 8.0 | 10.1 | 9.5 | 0.4 | 8.7 | 9.7 | 9.3 | 0.2 |
| BIO3 | Isothermality (%) | 33.3 | 40.0 | 37.7 | 1.3 | 35.2 | 38.5 | 36.9 | 0.6 |
| BIO4 | Temperature seasonality (%) | 545.5 | 582.9 | 564.3 | 8.8 | 561.1 | 581.0 | 573.6 | 3.6 |
| BIO5 | Max. temperature of the warmest month (°C) | 26.2 | 29.4 | 28.6 | 0.5 | 27.5 | 29.0 | 28.4 | 0.3 |
| BIO6 | Min. temperature of the coldest month (°C) | 2.3 | 4.0 | 3.5 | 0.3 | 2.9 | 3.7 | 3.3 | 0.1 |
| BIO7 | Temperature annual range (°C) | 23.8 | 25.6 | 25.1 | 0.3 | 24.6 | 25.4 | 25.1 | 0.2 |
| BIO8 | Mean temperature of the wettest quarter (°C) | 6.4 | 9.1 | 8.3 | 0.5 | 7.2 | 8.6 | 7.9 | 0.3 |
| BIO9 | Mean temperature of the driest quarter (°C) | 19.6 | 21.8 | 21.3 | 0.4 | 20.5 | 21.5 | 21.2 | 0.2 |
| BIO10 | Mean temperature of the warmest quarter (°C) | 19.7 | 21.8 | 21.3 | 0.3 | 20.5 | 21.5 | 21.2 | 0.2 |
| BIO11 | Mean temperature of the coldest quarter (°C) | 5.6 | 8.4 | 7.6 | 0.5 | 6.5 | 7.9 | 7.3 | 0.3 |
| BIO12 | Annual precipitation (mm) | 977.2 | 1263.2 | 1058.9 | 51.8 | 1019.7 | 1176.5 | 1103.4 | 32.5 |
| BIO13 | Precipitation of the wettest month (mm) | 145.2 | 197.5 | 159.7 | 8.5 | 153.9 | 179.9 | 166.1 | 5.2 |
| BIO14 | Precipitation of the driest month (mm) | 9.0 | 13.7 | 10.9 | 1.0 | 10.0 | 13.0 | 12.0 | 0.6 |
| BIO15 | Precipitation seasonality (%) | 57.8 | 59.2 | 58.4 | 0.2 | 57.9 | 58.7 | 58.2 | 0.1 |
| BIO16 | Precipitation of the wettest quarter (mm) | 408.2 | 540.0 | 446.9 | 23.0 | 430.1 | 499.5 | 466.3 | 13.6 |
| BIO17 | Precipitation of the driest quarter (mm) | 52.0 | 80.5 | 59.6 | 5.0 | 55.2 | 71.6 | 64.0 | 3.2 |
| BIO18 | Precipitation of the warmest quarter (mm) | 52.2 | 81.8 | 60.9 | 5.5 | 55.2 | 73.6 | 64.3 | 3.4 |
| BIO19 | Precipitation of the coldest quarter (mm) | 397.2 | 522.6 | 437.9 | 22.4 | 421.8 | 487.7 | 457.2 | 13.7 |
| E | Elevation (m) | 425.1 | 1133.1 | 585.5 | 119.2 | 505.4 | 848.6 | 669.6 | 62.7 |
| S | Slope (%) | 0.3 | 57.5 | 15.9 | 10.9 | 5.0 | 45.2 | 27.6 | 8.3 |
| A | Aspect (°) | 0.0 | 360.0 | 188.3 | 121.4 | 0.1 | 359.6 | 219.5 | 151.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almeida, A.M.; Delgado, F.; Roque, N.; Ribeiro, M.M.; Fernandez, P. Multitemporal Land Use and Cover Analysis Coupled with Climatic Change Scenarios to Protect the Endangered Taxon Asphodelus bento-rainhae subsp. bento-rainhae. Plants 2023, 12, 2914. https://doi.org/10.3390/plants12162914
Almeida AM, Delgado F, Roque N, Ribeiro MM, Fernandez P. Multitemporal Land Use and Cover Analysis Coupled with Climatic Change Scenarios to Protect the Endangered Taxon Asphodelus bento-rainhae subsp. bento-rainhae. Plants. 2023; 12(16):2914. https://doi.org/10.3390/plants12162914
Chicago/Turabian StyleAlmeida, Alice Maria, Fernanda Delgado, Natália Roque, Maria Margarida Ribeiro, and Paulo Fernandez. 2023. "Multitemporal Land Use and Cover Analysis Coupled with Climatic Change Scenarios to Protect the Endangered Taxon Asphodelus bento-rainhae subsp. bento-rainhae" Plants 12, no. 16: 2914. https://doi.org/10.3390/plants12162914
APA StyleAlmeida, A. M., Delgado, F., Roque, N., Ribeiro, M. M., & Fernandez, P. (2023). Multitemporal Land Use and Cover Analysis Coupled with Climatic Change Scenarios to Protect the Endangered Taxon Asphodelus bento-rainhae subsp. bento-rainhae. Plants, 12(16), 2914. https://doi.org/10.3390/plants12162914