

Construction of High-Density Genetic Map and QTL Mapping for Grain Shape in the Rice RIL Population


Abstract
1. Introduction
2. Results
2.1. Phenotypic Variation in the ZP37 X R8605 RIL Population
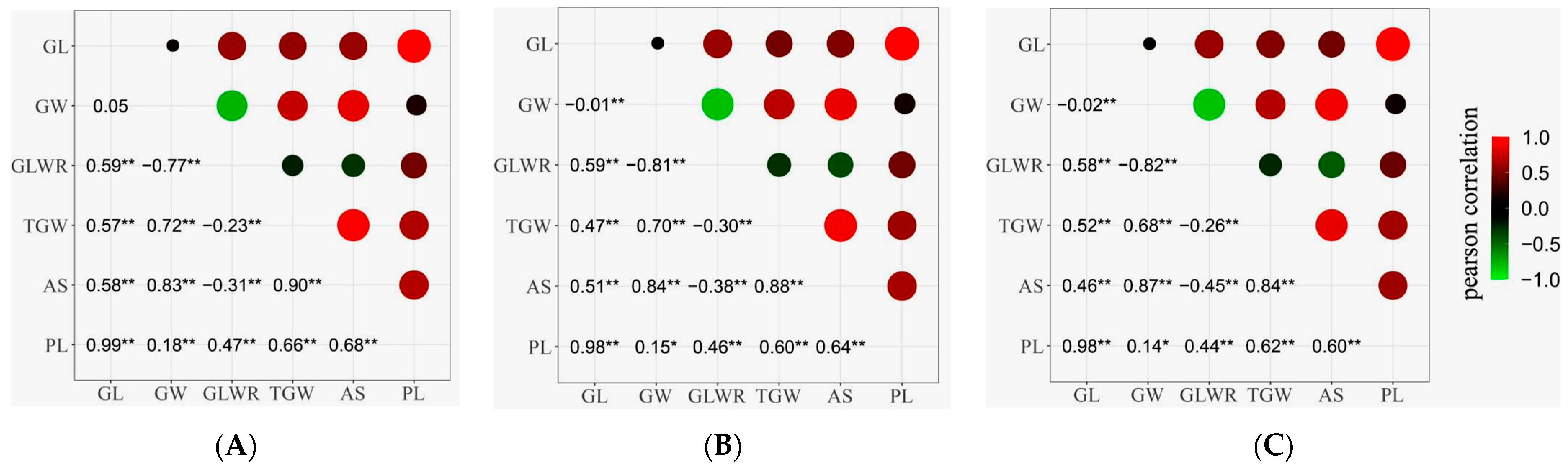
2.2. Correlation Analysis
2.3. Sequencing Data Analysis and Construction of the High-Density Genetic Map
2.4. QTL Mapping for Grain Shape Traits
2.5. Analysis of Loci with Stable Genetic Effects of Grain Shape
2.6. Analysis of Genetic Pleiotropy and Pyramiding Effects of Three Grain-Related Loci
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Phenotype Investigation and Data Analysis
4.3. Genotype Identification and High-Density Genetic Map Construction of the RIL Population
4.4. QTL Mapping
4.5. Analysis of Genetically Stable Loci and Pyramiding Effects
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ito, S. Contemporary global rice economies: Structural changes of rice production/consumption and trade. J. Nutr. Sci. Vitaminol. 2019, 65, S23–S25. [Google Scholar] [CrossRef] [PubMed]
- Roychowdhury, R.; Karmakar, J.; Karmakar, J.; Adak, M.K.; Dey, N. Physio-biochemical and microsatellite based profiling of lowland rice (Oryza sativa L.) landraces for osmotic stress tolerance. Am. J. Plant Sci. 2013, 4, 52–63. [Google Scholar] [CrossRef]
- Karmakar, J.; Roychowdhury, R.; Kar, R.K.; Deb, D.; Dey, N. Profiling of selected indigenous rice (Oryza sativa L.) landraces of Rarh Bengal in relation to osmotic stress tolerance. Physiol. Mol. Biol. Plants 2012, 18, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Anumalla, M.; Roychowdhury, R.; Geda, C.K.; Bharathkumar, S.; Goutam, K.D.; Mohandev, T.S.S. Mechanism of stress signal transduction and involvement of stress inducible transcription factors and genes in response to abiotic stresses in plant. Int. J. Rec. Sci. Res. 2016, 7, 12754–12771. [Google Scholar]
- Anumalla, M.; Roychowdhury, R.; Geda, C.K.; Mazid, M.; Rathoure, A.K. Utilization of plant genetic resources and diversity analysis tools for sustainable crop improvement with special emphasis on rice. Int. J. Adv. Res. 2015, 3, 1155–1175. [Google Scholar]
- Zuo, J.; Li, J. Molecular genetic dissection of quantitative trait loci regulating rice grain size. Annu. Rev. Genet. 2014, 48, 99–118. [Google Scholar] [CrossRef]
- Xing, Y.; Zhang, Q. Genetic and molecular bases of rice yield. Annu. Rev. Plant Biol. 2010, 61, 421–442. [Google Scholar] [CrossRef]
- Huang, R.; Jiang, L.; Zheng, J.; Wang, T.; Hong, Z. Genetic bases of rice grain shape: So many genes, so little known. Trends Plant Sci. 2012, 18, 218–226. [Google Scholar] [CrossRef]
- Che, R.; Tong, H.; Shi, B.; Liu, Y.; Fang, S.; Liu, D.; Xiao, Y.; Hu, B.; Liu, L.; Wang, H.; et al. Control of grain size and rice yield by GL2-mediated brassinosteroid responses. Nat. Plants 2015, 2, 15195. [Google Scholar] [CrossRef]
- Zhang, Y.D.; Zhu, Z.; Zhao, Q.Y.; Chen, T.; Yao, S.; Zhou, L.H.; Zhao, L.; Zhao, C.F.; Wang, C.L. Haplotypes of qGL3 and their roles in grain size regulation with GS3 alleles in rice. Genet. Mol. Res. 2016, 15, gmr.15017587. [Google Scholar] [CrossRef]
- Fan, C.; Xing, Y.; Mao, H.; Lu, T.; Han, B.; Xu, C.; Li, X.; Zhang, Q. GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein. Theor. Appl. Genet. 2006, 112, 1164–1171. [Google Scholar] [CrossRef] [PubMed]
- Qi, P.; Lin, Y.S.; Song, X.J.; Shen, J.B.; Huang, W.; Shan, J.X.; Zhu, M.Z.; Jiang, L.; Gao, J.P.; Lin, H.X. The novel quantitative trait locus GL3.1 controls rice grain size and yield by regulating Cyclin-T1;3. Cell Res. 2012, 22, 1666–1680. [Google Scholar] [CrossRef]
- Wu, W.; Liu, X.; Wang, M.; Meyer, R.S.; Luo, X.; Ndjiondjop, M.N.; Tan, L.; Zhang, J.; Wu, J.; Cai, H.; et al. A single-nucleotide polymorphism causes smaller grain size and loss of seed shattering during African rice domestication. Nat. Plants 2017, 3, 17064. [Google Scholar] [CrossRef]
- Si, L.; Chen, J.; Huang, X.; Gong, H.; Luo, J.; Hou, Q.; Zhou, T.; Lu, T.; Zhu, J.; Shangguan, Y.; et al. OsSPL13 controls grain size in cultivated rice. Nat. Genet. 2016, 48, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Song, X.J.; Huang, W.; Shi, M.; Zhu, M.Z.; Lin, H.X. A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase. Nat. Genet. 2007, 39, 623–630. [Google Scholar] [CrossRef]
- Liu, J.; Chen, J.; Zheng, X.; Wu, F.; Lin, Q.; Heng, Y.; Tian, P.; Cheng, Z.; Yu, X.; Zhou, K.; et al. GW5 acts in the brassinosteroid signalling pathway to regulate grain width and weight in rice. Nat. Plants 2017, 3, 17043. [Google Scholar] [CrossRef]
- Wang, S.; Wu, K.; Yuan, Q.; Liu, X.; Liu, Z.; Lin, X.; Zeng, R.; Zhu, H.; Dong, G.; Qian, Q.; et al. Control of grain size, shape and quality by OsSPL16 in rice. Nat. Genet. 2012, 44, 950–954. [Google Scholar] [CrossRef]
- Xu, C.; Liu, Y.; Li, Y.; Xu, X.; Xu, C.; Li, X.; Xiao, J.; Zhang, Q. Differential expression of GS5 regulates grain size in rice. J. Exp. Bot. 2015, 66, 2611–2623. [Google Scholar] [CrossRef] [PubMed]
- Ishimaru, K.; Hirotsu, N.; Madoka, Y.; Murakami, N.; Hara, N.; Onodera, H.; Kashiwagi, T.; Ujiie, K.; Shimizu, B.; Onishi, A.; et al. Loss of function of the IAA-glucose hydrolase gene TGW6 enhances rice grain weight and increases yield. Nat. Genet. 2013, 45, 707–711. [Google Scholar] [CrossRef]
- Ruan, B.; Shang, L.; Zhang, B.; Hu, J.; Wang, Y.; Lin, H.; Zhang, A.; Liu, C.; Peng, Y.; Zhu, L.; et al. Natural variation in the promoter of TGW2 determines grain width and weight in rice. New Phytol. 2020, 227, 629–640. [Google Scholar] [CrossRef]
- Ying, J.Z.; Ma, M.; Bai, C.; Huang, X.; Liu, J.L.; Fan, Y.Y.; Song, X.J. TGW3, a major QTL that negatively modulates grain length and weight in rice. Mol. Plant 2018, 11, 750–753. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, S.; Liu, Q.; Wu, K.; Zhang, J.; Wang, S.; Wang, Y.; Chen, X.; Zhang, Y.; Gao, C.; et al. The OsSPL16-GW7 regulatory module determines grain shape and simultaneously improves rice yield and grain quality. Nat. Genet. 2015, 47, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xiong, G.; Hu, J.; Jiang, L.; Yu, H.; Xu, J.; Fang, Y.; Zeng, L.; Xu, E.; Xu, J.; et al. Copy number variation at the GL7 locus contributes to grain size diversity in rice. Nat. Genet. 2015, 47, 944–948. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Wang, Y.; Fang, Y.; Zeng, L.; Xu, J.; Yu, H.; Shi, Z.; Pan, J.; Zhang, D.; Kang, S.; et al. A rare allele of GS2 enhances grain size and grain yield in rice. Mol. Plant 2015, 8, 1455–1465. [Google Scholar] [CrossRef]
- Ogawa, D.; Kamura, T.; Suzuki, T.; Higashiyama, T.; Yamasaki, M.; Mori, H.; Inukai, Y.; Wu, J.; Kitano, H.; Sakakibara, H.; et al. Rare allele of a previously unidentified histone H4 acetyltransferase enhances grain weight, yield, and plant biomass in rice. Proc. Natl. Acad. Sci. USA 2015, 112, 76–81. [Google Scholar]
- Zhao, D.S.; Li, Q.F.; Zhang, C.Q.; Zhang, C.; Yang, Q.Q.; Pan, L.X.; Ren, X.Y.; Lu, J.; Gu, M.H.; Liu, Q.Q. GS9 acts as a transcriptional activator to regulate rice grain shape and appearance quality. Nat. Commun. 2018, 9, 1240. [Google Scholar] [CrossRef]
- Roychowdhury, R.; Das, S.P.; Gupta, A.; Parihar, P.; Chandrasekhar, K.; Sarker, U.; Kumar, A.; Ramrao, D.P.; Sudhakar, C. Multi-omics pipeline and omics-integration approach to decipher plant’s abiotic stress tolerance responses. Genes 2023, 14, 1281. [Google Scholar] [CrossRef]
- Wu, L.; Cui, Y.; Xu, Z.J.; Xu, Q. Identification of multiple grain shape-related loci in rice using bulked segregant analysis with high-throughput sequencing. Front. Plant Sci. 2020, 11, 303. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, J.; Wang, H.; LI, D.X.; Yang, G.L.; Huang, C.H.; Zhou, D.H.; Guo, T.; Chen, Z.Q.; Wang, H. QTL mapping for grain size related traits based on a high-density map in rice. Sci. Agric. Sin. 2020, 53, 225–238. (In Chinese) [Google Scholar]
- Zhang, Y.D.; Liang, W.H.; He, L.; Zhao, C.F.; Zhu, Z.; Chen, T.; Zhao, Q.Y.; Zhao, L.; Yao, S.; Zhou, L.H.; et al. Construction of high-density genetic map and QTL analysis of grain shape in rice RIL population. Sci. Agric. Sin. 2021, 54, 5163–5176. (In Chinese) [Google Scholar]
- Chen, L.; Gao, W.; Chen, S.; Wang, L.; Zou, J.; Liu, Y.; Wang, H.; Chen, Z.; Guo, T. High-resolution QTL mapping for grain appearance traits and co-localization of chalkiness-associated differentially expressed candidate genes in rice. Rice 2016, 9, 48. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Guo, Z.; Luo, L.; Gao, Q.; Xiao, W.; Wang, J.; Wang, H.; Chen, Z.; Guo, T. Identification of QTL and candidate genes involved in early seedling growth in rice via high-density genetic mapping and RNA-seq. Crop J. 2021, 9, 360–371. [Google Scholar] [CrossRef]
- Jin, S.; Xu, L.; Yang, Q.; Zhang, M.; Wang, S.; Wang, R.; Tao, T.; Hong, L.; Guo, Q.; Jia, S.; et al. High-resolution quantitative trait locus mapping for rice grain quality traits using genotyping by sequencing. Front. Plant Sci. 2023, 13, 1050882. [Google Scholar] [CrossRef]
- Yang, J.; Wei, J.; Xu, J.; Xiong, Y.; Deng, G.; Liu, J.; Fahad, S.; Wang, H. Mapping QTLs for anaerobic tolerance at germination and bud stages using new high density genetic map of rice. Front. Plant Sci. 2022, 13, 985080. [Google Scholar] [CrossRef]
- Heang, D.; Sassa, H. An atypical bHLH protein encoded by POSITIVE REGULATOR OF GRAIN LENGTH 2 is involved in controlling grain length and weight of rice through interaction with a typical bHLH protein APG. Breed. Sci. 2012, 62, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Qi, Z.; Xiong, L. Characterization of a purine permease family gene OsPUP7 involved in growth and development control in rice. J. Integr. Plant Biol. 2013, 55, 1119–1135. [Google Scholar] [CrossRef]
- Sun, S.; Wang, L.; Mao, H.; Shao, L.; Li, X.; Xiao, J.; Ouyang, Y.; Zhang, Q. A G-protein pathway determines grain size in rice. Nat. Commun. 2018, 9, 851. [Google Scholar] [CrossRef] [PubMed]
- Golicz, A.A.; Bayer, P.E.; Edwards, D. Skim-based genotyping by sequencing. Methods Mol. Biol. 2015, 1245, 257–270. [Google Scholar] [PubMed]
- Yu, H.; Xie, W.; Li, J.; Zhou, F.; Zhang, Q. A whole-genome SNP array (RICE6K) for genomic breeding in rice. Plant Biotechnol. J. 2014, 12, 28–37. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, Y.; Wei, X.; Liang, W.; Zhao, C.; Zhou, L.; Yao, S.; Wang, C.; Zhang, Y. Mapping of QTLs for chlorophyll content in flag leaves of rice on high-density bin map. Sci. Agric. Sin. 2022, 55, 825–836. (In Chinese) [Google Scholar]
- Yano, M.; Harushima, Y.; Nagamura, Y.; Kurata, N.; Minobe, Y.; Sasaki, T. Identification of quantitative trait loci controlling heading date in rice using a high-density linkage map. Theor. Appl. Genet. 1997, 95, 1025–1032. [Google Scholar] [CrossRef]
- Lafitte, H.R.; Price, A.H.; Courtois, B. Yield response to water deficit in an upland rice mapping population: Associations among traits and genetic markers. Theor. Appl. Genet. 2004, 109, 1237–1246. [Google Scholar] [CrossRef]
- Kumar, A.; Dixit, S.; Ram, T.; Yadaw, R.B.; Mishra, K.K.; Mandal, N.P. Breeding high-yielding drought-tolerant rice: Genetic variations and conventional and molecular approaches. J. Exp. Bot. 2014, 65, 6265–6278. [Google Scholar] [CrossRef]
- Reumers, J.; De Rijk, P.; Zhao, H.; Liekens, A.; Smeets, D.; Cleary, J.; Van Loo, P.; Van Den Bossche, M.; Catthoor, K.; Sabbe, B.; et al. Optimized filtering reduces the error rate in detecting genomic variants by short-read sequencing. Nat. Biotechnol. 2011, 30, 61–68. [Google Scholar] [CrossRef]
- Huang, X.; Feng, Q.; Qian, Q.; Zhao, Q.; Wang, L.; Wang, A.; Guan, J.; Fan, D.; Weng, Q.; Huang, T. High-throughput genotyping by whole-genome resequencing. Genome Res. 2009, 19, 1068–1076. [Google Scholar] [CrossRef]
- McCouch, S.R.; Kochert, G.; Yu, Z.H.; Wang, Z.Y.; Khush, G.S.; Coffman, W.R.; Tanksley, S.D. Molecular mapping of rice chromosomes. Theor. Appl. Genet. 1988, 76, 815–829. [Google Scholar] [CrossRef]
- Xueqin, G.; Fengzhen, C.; Fei, G.; Ling, L.; Ke, L.; Lijin, Y.; Cong, H.; Fan, Y.; Wanliang, L.; Chunhua, P. CNSA: A data repository for archiving omics data. Database 2020, 2020, baaa055. [Google Scholar]
- Chen, F.Z.; You, L.J.; Yang, F.; Wang, L.N.; Wei, X.F. CNGBdb: China National GeneBank DataBase. Hereditas 2020, 42, 799–809. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Trait a | Environment b | Parents c | RIL Population | |||||
|---|---|---|---|---|---|---|---|---|
| ZP37 | R8605 | Mean | Range | Skewness | Kurtosis | CV d (%) | ||
| GL (mm) | 2019 | 8.56 ± 0.21 | 9.27 ± 0.10 ** | 8.91 | 7.96–10.23 | 0.42 | −0.06 | 4.78 |
| 2020 | 8.68 ± 0.22 | 9.49 ± 0.12 ** | 9.02 | 7.90–10.29 | 0.08 | 0.25 | 4.65 | |
| 2022 | 8.80 ± 0.09 | 9.54 ± 0.22 ** | 9.43 | 8.48–10.70 | 0.30 | 0.13 | 4.37 | |
| GW (mm) | 2019 | 2.40 ± 0.06 | 2.55 ± 0.02 * | 2.42 | 2.08–2.75 | −0.02 | −0.65 | 6.06 |
| 2020 | 2.45 ± 0.03 | 2.57 ± 0.08 | 2.48 | 2.12–2.97 | 0.00 | −0.21 | 6.29 | |
| 2022 | 2.52 ± 0.04 | 2.61 ± 0.09 | 2.46 | 2.09–2.98 | 0.21 | −0.02 | 6.30 | |
| GLWR | 2019 | 3.57 ± 0.02 | 3.63 ± 0.03 * | 3.72 | 3.15–4.55 | 0.33 | −0.35 | 7.60 |
| 2020 | 3.55 ± 0.06 | 3.70 ± 0.07 * | 3.67 | 2.99–4.68 | 0.39 | 0.00 | 8.02 | |
| 2022 | 3.49 ± 0.05 | 3.66 ± 0.05 * | 3.87 | 3.04–4.81 | 0.45 | 0.15 | 7.86 | |
| TGW(g) | 2019 | 17.36 ± 0.11 | 21.95 ± 0.34 ** | 20.20 | 15.19–28.69 | 0.32 | 0.13 | 11.08 |
| 2020 | 18.22 ± 0.17 | 23.40 ± 0.21 ** | 21.76 | 16.23–28.35 | 0.07 | −0.03 | 10.17 | |
| 2022 | 19.70 ± 0.35 | 24.47 ± 0.14 ** | 23.13 | 15.16–29.95 | −0.07 | 0.32 | 10.37 | |
| AS (mm2) | 2019 | 16.25 ± 0.06 | 17.96 ± 0.23 ** | 16.89 | 13.77–20.70 | 0.05 | −0.49 | 8.18 |
| 2020 | 16.53 ± 0.11 | 18.64 ± 0.30 ** | 17.53 | 13.95–21.07 | −0.10 | −0.17 | 8.14 | |
| 2022 | 16.95 ± 0.19 | 19.39 ± 0.25 ** | 18.24 | 14.76–22.81 | 0.15 | −0.21 | 8.23 | |
| PL (mm) | 2019 | 19.89 ± 0.10 | 21.36 ± 0.22 ** | 20.73 | 18.68–23.63 | 0.43 | −0.05 | 4.48 |
| 2020 | 20.08 ± 0.19 | 21.57 ± 0.18 ** | 17.53 | 18.97–23.57 | 0.04 | −0.02 | 4.16 | |
| 2022 | 21.19 ± 0.013 | 22.42 ± 0.37 ** | 22.00 | 19.93–24.53 | 0.17 | −0.06 | 4.00 | |
| Chromosome | Number of Bin Markers | Length (cM) | Average Genetic Distance between Markers (cM) | Average Physical Distance between Markers (kb) |
|---|---|---|---|---|
| 1 | 122 | 126.24 | 1.03 | 354.65 |
| 2 | 248 | 138.35 | 0.56 | 144.88 |
| 3 | 293 | 137.93 | 0.47 | 124.27 |
| 4 | 308 | 192.93 | 0.63 | 115.14 |
| 5 | 151 | 96.82 | 0.64 | 198.39 |
| 6 | 88 | 68.68 | 0.78 | 355.05 |
| 7 | 120 | 98.58 | 0.82 | 247.34 |
| 8 | 196 | 143.89 | 0.73 | 145.10 |
| 9 | 162 | 119.46 | 0.74 | 142.04 |
| 10 | 71 | 78.38 | 1.10 | 326.85 |
| 11 | 235 | 147.96 | 0.63 | 123.47 |
| 12 | 199 | 193.02 | 0.97 | 138.34 |
| Total | 2193 | 1542.24 | 0.76 | 201.29 |
| Trait | QTL | Chr. | Marker Interval | Physical Interval (bp) | QTL Interval Size (kb) | Environment | LOD | PVE (%) | ADD |
|---|---|---|---|---|---|---|---|---|---|
| GL | qGL-2 | 2 | Block1305–Block1318 | 29,677,627–30,291,539 | 613.9 | 2020 | 5.56 | 6.50 | 0.16 |
| 2 | Block1305–Block1318 | 29,677,627–30,291,539 | 613.9 | 2022 | 4.81 | 5.39 | 0.14 | ||
| qGL-7-1 | 7 | Block5298–Block5303 | 2,305,563–2,667,445 | 361.9 | 2022 | 3.43 | 2.16 | −0.09 | |
| qGL-7-2 | 7 | Block5433–Block5439 | 20,606,977–21,226,418 | 619.4 | 2019 | 4.68 | 5.72 | −0.15 | |
| 7 | Block5433–Block5439 | 20,606,977–21,226,418 | 619.4 | 2020 | 5.53 | 4.98 | −0.14 | ||
| qGL-7-3 | 7 | Block5588–Block5589 | 28,557,384–28,724,630 | 167.2 | 2022 | 2.85 | 1.74 | −0.08 | |
| qGL-8 | 8 | Block6010–Block6012 | 23,379,921–23,447,755 | 67.8 | 2019 | 4.85 | 6.96 | −0.17 | |
| 8 | Block5989–Block6010 | 22,065,235–23,387,924 | 1322.7 | 2020 | 5.56 | 6.12 | −0.15 | ||
| 8 | Block5989–Block6010 | 22,065,235–23,387,924 | 1322.7 | 2022 | 4.81 | 3.46 | −0.11 | ||
| GW | qGW-3-1 | 3 | Block1966–Block1967 | 15,915,064–16,067,455 | 152.4 | 2022 | 3.00 | 3.29 | 0.04 |
| qGW-3-2 | 3 | Block1968–Block1974 | 16,148,731–16,617,377 | 468.6 | 2019 | 4.81 | 4.93 | 0.05 | |
| 3 | Block1970–Block1974 | 16,148,731–16,617,377 | 468.6 | 2022 | 3.00 | 3.90 | 0.05 | ||
| qGW-4-1 | 4 | Block3702–Block3701 | 19,871,103–19,942,280 | 71.2 | 2019 | 4.81 | 3.23 | −0.04 | |
| qGW-4-2 | 4 | Block3808–Block3815 | 20,702,635–20,878,645 | 176.0 | 2020 | 4.89 | 5.89 | −0.06 | |
| 4 | Block3808–Block3815 | 20,702,635–20,878,645 | 176.0 | 2022 | 3.00 | 3.29 | −0.04 | ||
| qGW-8 | 8 | Block6167–Block6169 | 27,555,403–27,911,017 | 355.6 | 2019 | 8.91 | 9.21 | 0.07 | |
| 8 | Block6167–Block6169 | 27,555,403–27,911,017 | 355.6 | 2020 | 15.71 | 14.10 | 0.09 | ||
| 8 | Block6167–Block6169 | 27,555,403–27,911,017 | 355.6 | 2022 | 11.57 | 10.73 | 0.07 | ||
| GLWR | qGLWR-2-1 | 2 | Block821 | 9,286,980–10,372,621 | 1085.6 | 2019 | 3.88 | 2.91 | 0.07 |
| qGLWR-2-2 | 2 | Block1221–Block1270 | 26,355,325–27,386,770 | 1031.4 | 2020 | 3.68 | 2.03 | 0.06 | |
| qGLWR-4 | 4 | Block3684–Block3688 | 19,157,777–19,344,941 | 187.2 | 2022 | 3.00 | 2.01 | 0.06 | |
| qGLWR-5 | 5 | Block4850–Block4851 | 27,634,637–27,858,389 | 223.8 | 2020 | 3.68 | 3.53 | 0.08 | |
| qGLWR-7 | 7 | Block5298–Block5299 | 2,305,563–2,328,233 | 22.7 | 2022 | 3.00 | 1.45 | −0.05 | |
| qGLWR-8 | 8 | Block6167–Block6169 | 27,555,403–27,911,017 | 355.6 | 2019 | 16.30 | 14.20 | −0.16 | |
| 8 | Block6167–Block6169 | 27,555,403–27,911,017 | 355.6 | 2020 | 15.51 | 13.83 | −0.16 | ||
| 8 | Block6167–Block6169 | 27,555,403–27,911,017 | 355.6 | 2022 | 16.12 | 12.46 | −0.16 | ||
| TGW | qTGW-2 | 2 | Block1305–Block1318 | 29,677,627–30,291,539 | 613.9 | 2020 | 4.85 | 4.97 | 0.41 |
| qTGW-3 | 3 | Block1970–Block1974 | 16,148,731–16,617,377 | 468.6 | 2019 | 3.29 | 2.66 | 0.05 | |
| 3 | Block1970–Block1974 | 16,148,731–16,617,377 | 468.6 | 2022 | 4.70 | 3.67 | 0.67 | ||
| qTGW-4-1 | 4 | Block3830–Block3834 | 20,994,282–21,294,165 | 299.9 | 2020 | 3.97 | 3.92 | −0.64 | |
| qTGW-4-2 | 4 | Block3948–Block3949 | 24,101,830–24,109,967 | 8.1 | 2022 | 4.89 | 3.76 | −0.68 | |
| qTGW-4-3 | 4 | Block3955–Block3957 | 24,194,456–24,626,257 | 431.8 | 2019 | 4.54 | 2.93 | −0.56 | |
| qTGW-7-1 | 7 | Block5435–Block5490 | 20,956,479–22,680,384 | 1723.9 | 2020 | 3.97 | 4.23 | −0.67 | |
| qTGW-7-2 | 7 | Block5588–Block5589 | 28,557,384–28,724,630 | 167.2 | 2019 | 3.00 | 2.09 | −0.48 | |
| qTGW-8-1 | 8 | Block5806–Block5808 | 19,645,419–19,698,589 | 53.2 | 2019 | 4.79 | 3.76 | −0.64 | |
| 8 | Block5807–Block5808 | 19,672,283–19,698,589 | 263.1 | 2022 | 4.89 | 5.75 | −0.85 | ||
| qTGW-8-2 | 8 | Block5989–Block6012 | 22,065,235–23,447,755 | 1382.5 | 2020 | 4.85 | 4.60 | −0.70 | |
| AS | qAS-2-1 | 2 | Block1352–Block1354 | 31,122,113–31,447,924 | 325.8 | 2020 | 4.88 | 5.34 | 0.48 |
| qAS-2-2 | 2 | Block1376–Block1377 | 32,614,759–32,704,042 | 89.3 | 2019 | 3.60 | 4.07 | 0.41 | |
| qAS-3 | 3 | Block1970–Block1974 | 16,148,731–16,617,377 | 468.6 | 2019 | 3.78 | 3.38 | 0.37 | |
| 3 | Block1970–Block1974 | 16,148,731–16,617,377 | 468.6 | 2022 | 2.87 | 4.25 | 0.45 | ||
| qAS-4-1 | 4 | Block3830–Block3834 | 20,994,282–21,294,165 | 299.9 | 2019 | 3.39 | 3.42 | −0.38 | |
| 4 | Block3830–Block3834 | 20,994,282–21,294,165 | 299.9 | 2020 | 4.88 | 4.77 | −0.46 | ||
| qAS-4-2 | 4 | Block3948–Block3949 | 24,101,830–24,109,967 | 8.1 | 2022 | 3.00 | 3.55 | −0.42 | |
| qAS-4-3 | 4 | Block3955–Block3967 | 24,194,456–25,976,047 | 1781.6 | 2022 | 3.00 | 3.31 | −0.40 | |
| qAS-4-4 | 4 | Block4088–Block4087 | 30,322,633–30,361,362 | 38.7 | 2022 | 3.00 | 2.45 | −0.35 | |
| qAS-7 | 7 | Block5435–Block5463 | 20,956,479–21,435,416 | 478.9 | 2019 | 3.35 | 3.51 | −0.38 | |
| qAS-8 | 8 | Block6167–Block6169 | 27,555,403–27,911,017 | 355.6 | 2020 | 9.17 | 8.08 | 0.60 | |
| 8 | Block6167–Block6169 | 27,555,403–27,911,017 | 355.6 | 2022 | 5.70 | 6.30 | 0.55 | ||
| PL | qPL-2 | 2 | Block1305–Block1318 | 29,677,627–30,291,539 | 613.9 | 2020 | 5.03 | 6.91 | 0.34 |
| 2 | Block1305–Block1318 | 29,677,627–30,291,539 | 613.9 | 2022 | 4.93 | 5.79 | 0.31 | ||
| qPL-7 | 7 | Block5433–Block5439 | 20,606,977–21,226,418 | 619.4 | 2019 | 4.80 | 5.85 | −0.33 | |
| 7 | Block5433–Block5435 | 20,606,977–20,956,479 | 349.5 | 2020 | 5.03 | 5.83 | −0.31 | ||
| qPL-8 | 8 | Block5989–Block6010 | 22,065,235–23,387,924 | 1322.7 | 2019 | 4.93 | 6.06 | −0.34 | |
| 8 | Block5989–Block6012 | 22,065,235–23,447,755 | 1382.5 | 2020 | 5.03 | 4.44 | −0.27 | ||
| 8 | Block5989–Block6010 | 22,065,235–23,387,924 | 1322.7 | 2022 | 3.41 | 3.12 | −0.23 |
| S. No. | Locus | QTL | Marker Interval | Physical Interval (bp) | PVE (%) | Overlapped QTL Reported |
|---|---|---|---|---|---|---|
| 1 | Locus 1 | qGL-2 (2020, 2022); qTGW-2 (2020); qPL-2 (2020, 2022) | Block1305–Block1318 | 29,677,627–30,291,539 | 4.97–6.91 | |
| 2 | Locus 2 | qAS-2-1 (2020) | Block1352–Block1354 | 31,122,113–31,447,924 | 5.34 | PGL2 |
| 3 | Locus 3 | qGW-3-2 (2019, 2022); qTGW-3 (2019, 2022); qAS-3 (2019, 2022) | Block1968–Block1974 | 16,148,731–16,617,377 | 2.66–4.93 | |
| 4 | Locus 4 | qGW-4-2 (2020, 2022) | Block3808–Block3815 | 20,702,635–20,878,645 | 3.29–5.89 | |
| 5 | Locus 5 | qTGW-4-1 (2020); qAS-4-1 (2019, 2020) | Block3830–Block3834 | 20,994,282–21,294,165 | 3.42–4.77 | |
| 6 | Locus 6 | qTGW-4-2 (2022); qAS-4-2 (2022) | Block3948–Block3949 | 24,101,830–24,109,967 | 3.55–3.76 | |
| 7 | Locus 7 | qTGW-4-3 (2019); qAS-4-3 (2022) | Block3955–Block3967 | 24,194,456–25,976,047 | 2.93–3.31 | |
| 8 | Locus 8 | qGLWR-5 (2020) | Block4850–Block4851 | 27,634,637–27,858,389 | 3.53 | OsPUP7 |
| 9 | Locus 9 | qGL-7-1 (2022); qGLWR-7 (2022) | Block5298–Block5303 | 2,305,563–2,667,445 | 1.45–2.16 | |
| 10 | Locus 10 | qGL-7-2 (2019, 2020); qTGW-7-1 (2020); qAS-7 (2019); qPL-7 (2019, 2020) | Block5433–Block5490 | 20,606,977–22,680,384 | 3.51–5.85 | |
| 11 | Locus 11 | qGL-7-3 (2022); qTGW-7-2 (2019) | Block5588–Block5589 | 28,557,384–28,724,630 | 1.74–2.09 | |
| 12 | Locus 12 | qTGW-8-1 (2019, 2022) | Block5807–Block5808 | 19,672,283–19,698,589 | 3.76–5.75 | |
| 13 | Locus 13 | qGL-8 (2019, 2020, 2022); qTGW-8-2 (2020); qPL-8 (2019, 2020, 2022) | Block5989–Block6012 | 22,065,235–23,447,755 | 3.42–6.96 | GGC2 |
| 14 | Locus 14 | qGW-8 (2019, 2020, 2022); qGLWR-8 (2019, 2020, 2022); qAS-8 (2020, 2022) | Block6167–Block6169 | 27,555,403–27,911,017 | 6.30–14.20 |
| Trait | QTL | Environment | Number of RILs of Marker Type aa | Number of RILs of Marker Type bb | Donors of Positive Allele | Phenotypic Value | Difference | |
|---|---|---|---|---|---|---|---|---|
| Marker Type aa | Marker Type bb | |||||||
| Locus 1 | ||||||||
| GL (mm) | - | 2019 | 82 | 120 | bb | 8.82 ± 0.37 | 9.00 ± 0.45 | 0.17 ** |
| qGL-2 | 2020 | 8.85 ± 0.37 | 9.13 ± 0.42 | 0.28 ** | ||||
| qGL-2 | 2022 | 9.27 ± 0.38 | 9.54 ± 0.40 | 0.28 ** | ||||
| TGW(g) | - | 2019 | 19.82 ± 2.09 | 20.42 ± 2.34 | 0.60 | |||
| qTGW-2 | 2020 | 21.11 ± 2.18 | 22.18 ± 2.18 | 1.07 ** | ||||
| - | 2022 | 22.57 ± 2.53 | 23.51 ± 2.17 | 0.94 ** | ||||
| PL (mm) | - | 2019 | 20.51 ± 0.79 | 20.90 ± 0.99 | 0.39 ** | |||
| qPL-2 | 2020 | 20.81 ± 0.80 | 21.43 ± 0.86 | 0.62 ** | ||||
| qPL-2 | 2022 | 21.65 ± 0.84 | 22.26 ± 0.82 | 0.61 ** | ||||
| Locus 10 | ||||||||
| GL (mm) | qGL-7-2 | 2019 | 88 | 118 | aa | 9.07 ± 0.42 | 8.80 ± 0.39 | 0.27 ** |
| qGL-7-2 | 2020 | 9.17 ± 0.42 | 8.91 ± 0.38 | 0.26 ** | ||||
| - | 2022 | 9.55 ± 0.42 | 9.34 ± 0.39 | 0.21 ** | ||||
| TGW(g) | - | 2019 | 20.87 ± 2.14 | 19.69 ± 2.18 | 1.18 ** | |||
| qTGW-7-1 | 2020 | 22.49 ± 2.19 | 21.2 ± 2.09 | 1.29 ** | ||||
| - | 2022 | 23.61 ± 2.50 | 22.72 ± 2.24 | 0.90 ** | ||||
| AS (mm2) | qAS-7 | 2019 | 17.27 ± 1.26 | 16.60 ± 1.41 | 0.66 ** | |||
| - | 2020 | 17.89 ± 1.39 | 17.25 ± 1.40 | 0.64 ** | ||||
| - | 2022 | 18.51 ± 1.62 | 18.01 ± 1.38 | 0.49 * | ||||
| PL (mm) | qPL-7 | 2019 | 21.08 ± 0.90 | 20.47 ± 0.87 | 0.60 ** | |||
| qPL-7 | 2020 | 21.51 ± 0.87 | 20.93 ± 0.80 | 0.58 ** | ||||
| - | 2022 | 22.26 ± 0.89 | 21.80 ± 0.83 | 0.46 ** | ||||
| Locus 13 | ||||||||
| GL (mm) | qGL-8 | 2019 | 83 | 122 | aa | 9.11 ± 0.43 | 8.79 ± 0.37 | 0.32 ** |
| qGL-8 | 2020 | 9.19 ± 0.38 | 8.90 ± 0.41 | 0.30 ** | ||||
| qGL-8 | 2022 | 9.56 ± 0.42 | 9.35 ± 0.39 | 0.21 ** | ||||
| TGW(g) | - | 2019 | 20.75 ± 2.46 | 19.83 ± 2.02 | 0.92 ** | |||
| qTGW-8-2 | 2020 | 22.23 ± 2.44 | 21.45 ± 2.01 | 0.77 * | ||||
| - | 2022 | 23.58 ± 2.60 | 22.86 ± 2.22 | 0.73 * | ||||
| PL (mm) | qPL-8 | 2019 | 21.12 ± 0.95 | 20.47 ± 0.82 | 0.65 ** | |||
| qPL-8 | 2020 | 21.51 ± 0.79 | 20.94 ± 0.87 | 0.57 ** | ||||
| qPL-8 | 2022 | 22.25 ± 0.88 | 21.84 ± 0.84 | 0.41 ** | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, M.; Luo, T.; Huang, D.; Ma, Z.; Liu, C.; Qin, Y.; Wu, Z.; Zhou, X.; Lu, Y.; Yan, L.; et al. Construction of High-Density Genetic Map and QTL Mapping for Grain Shape in the Rice RIL Population. Plants 2023, 12, 2911. https://doi.org/10.3390/plants12162911
Wei M, Luo T, Huang D, Ma Z, Liu C, Qin Y, Wu Z, Zhou X, Lu Y, Yan L, et al. Construction of High-Density Genetic Map and QTL Mapping for Grain Shape in the Rice RIL Population. Plants. 2023; 12(16):2911. https://doi.org/10.3390/plants12162911
Chicago/Turabian StyleWei, Minyi, Tongping Luo, Dahui Huang, Zengfeng Ma, Chi Liu, Yuanyuan Qin, Zishuai Wu, Xiaolong Zhou, Yingping Lu, Liuhui Yan, and et al. 2023. "Construction of High-Density Genetic Map and QTL Mapping for Grain Shape in the Rice RIL Population" Plants 12, no. 16: 2911. https://doi.org/10.3390/plants12162911
APA StyleWei, M., Luo, T., Huang, D., Ma, Z., Liu, C., Qin, Y., Wu, Z., Zhou, X., Lu, Y., Yan, L., Qin, G., & Zhang, Y. (2023). Construction of High-Density Genetic Map and QTL Mapping for Grain Shape in the Rice RIL Population. Plants, 12(16), 2911. https://doi.org/10.3390/plants12162911