
Phytochemistry, Biological, and Pharmacological Properties of Abies alba Mill.


 ,
,
Abstract
1. Introduction
2. Parts Used and Extraction Methods
3. Phytochemistry
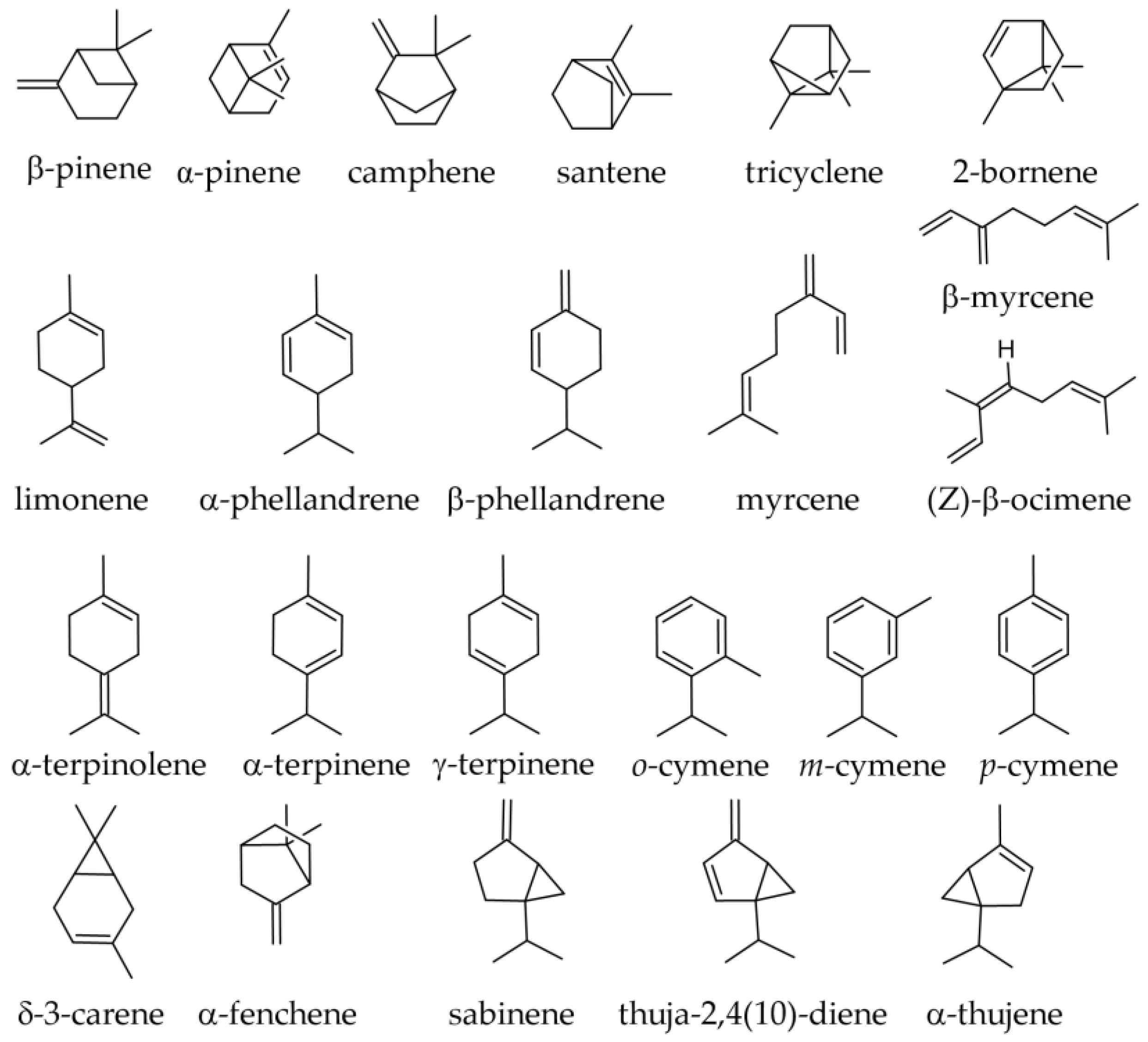
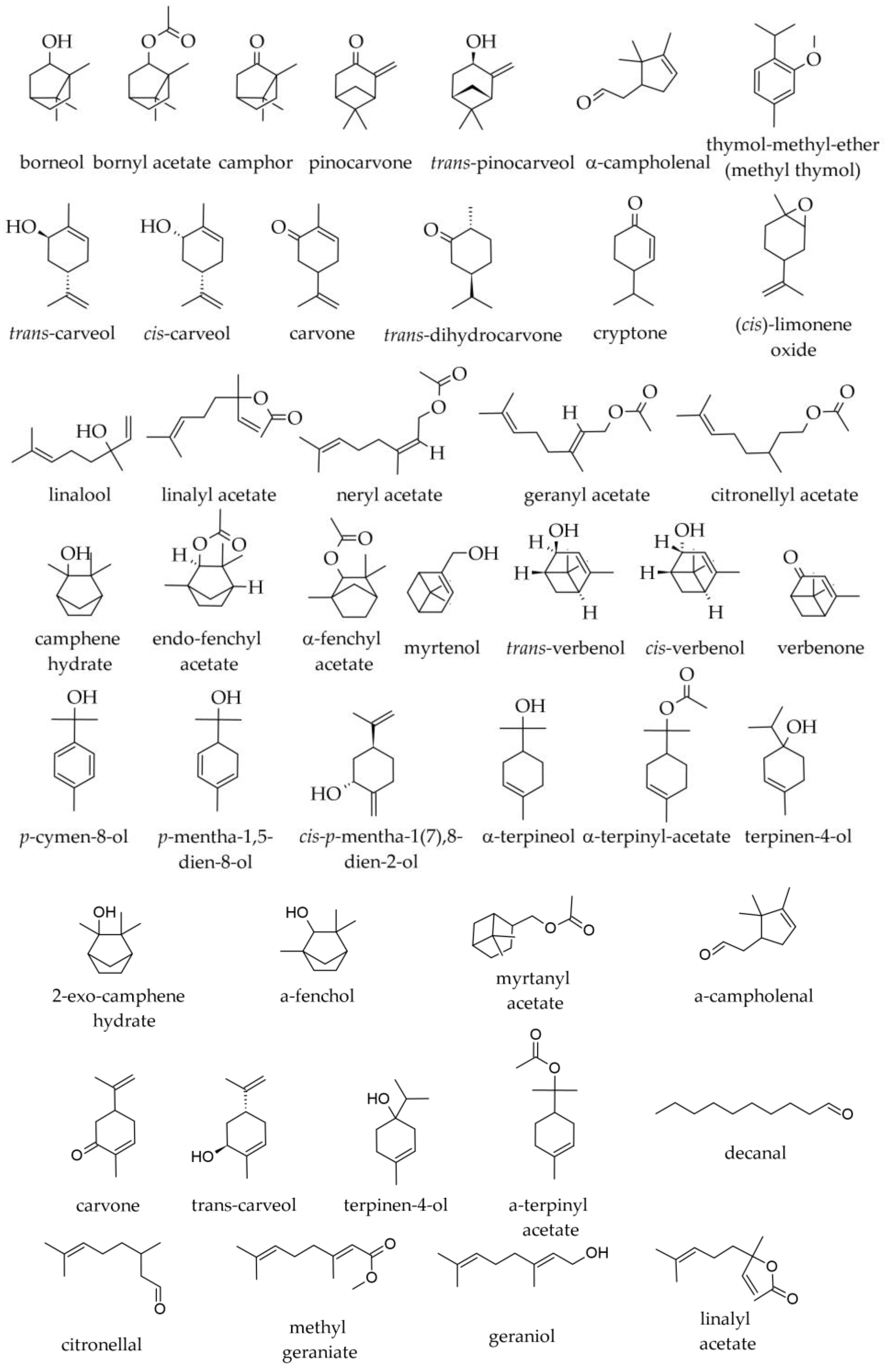
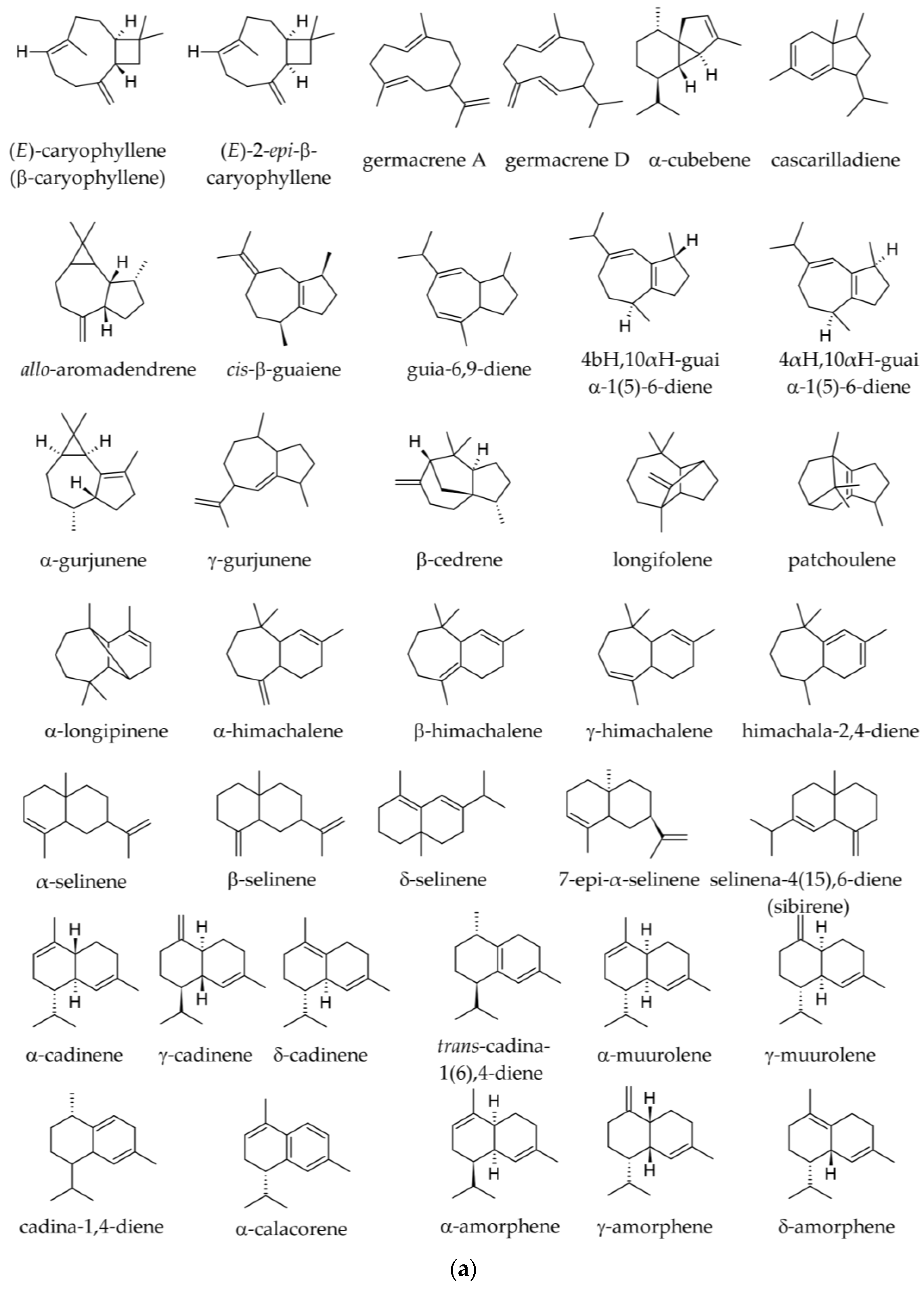
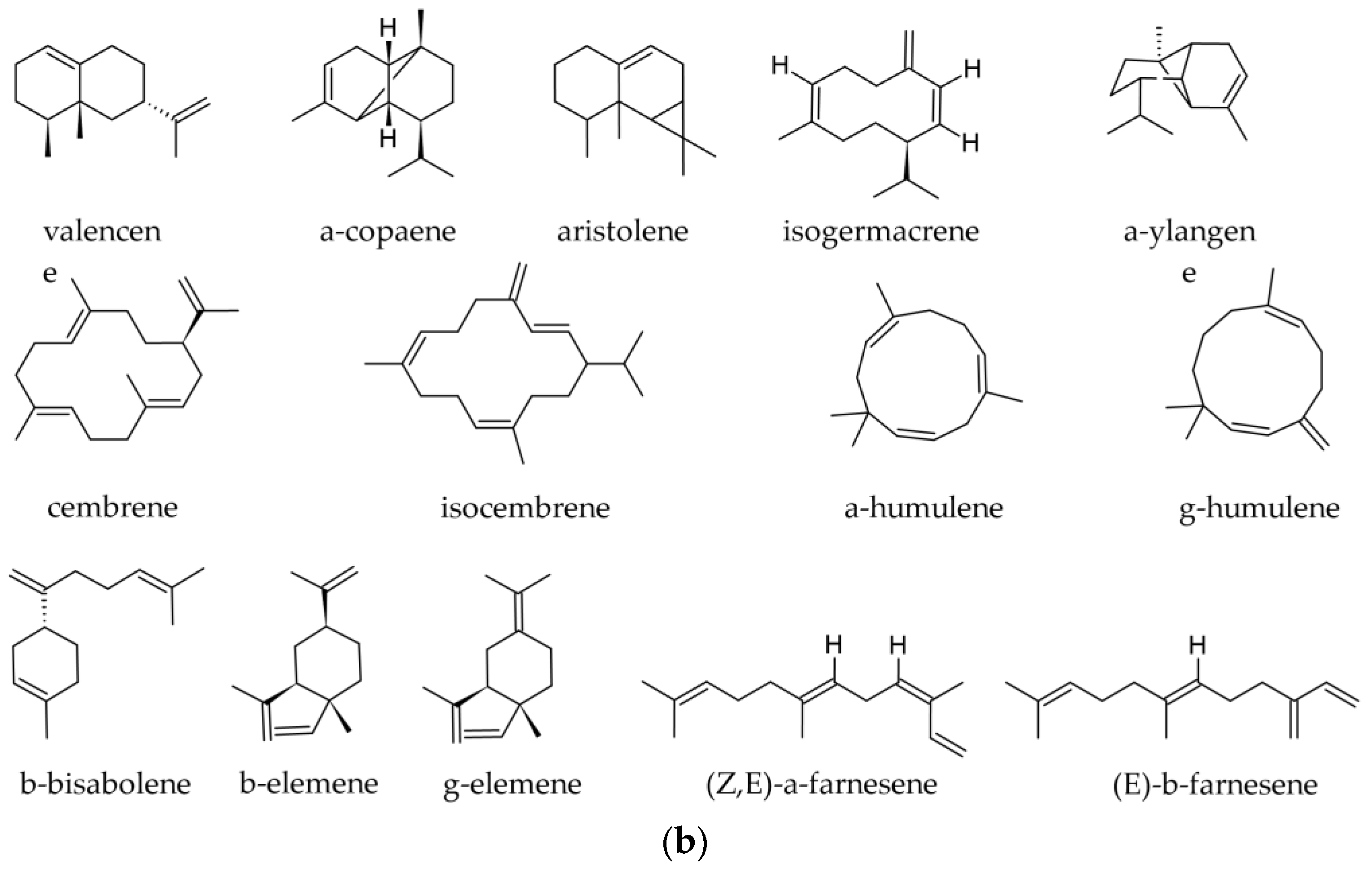
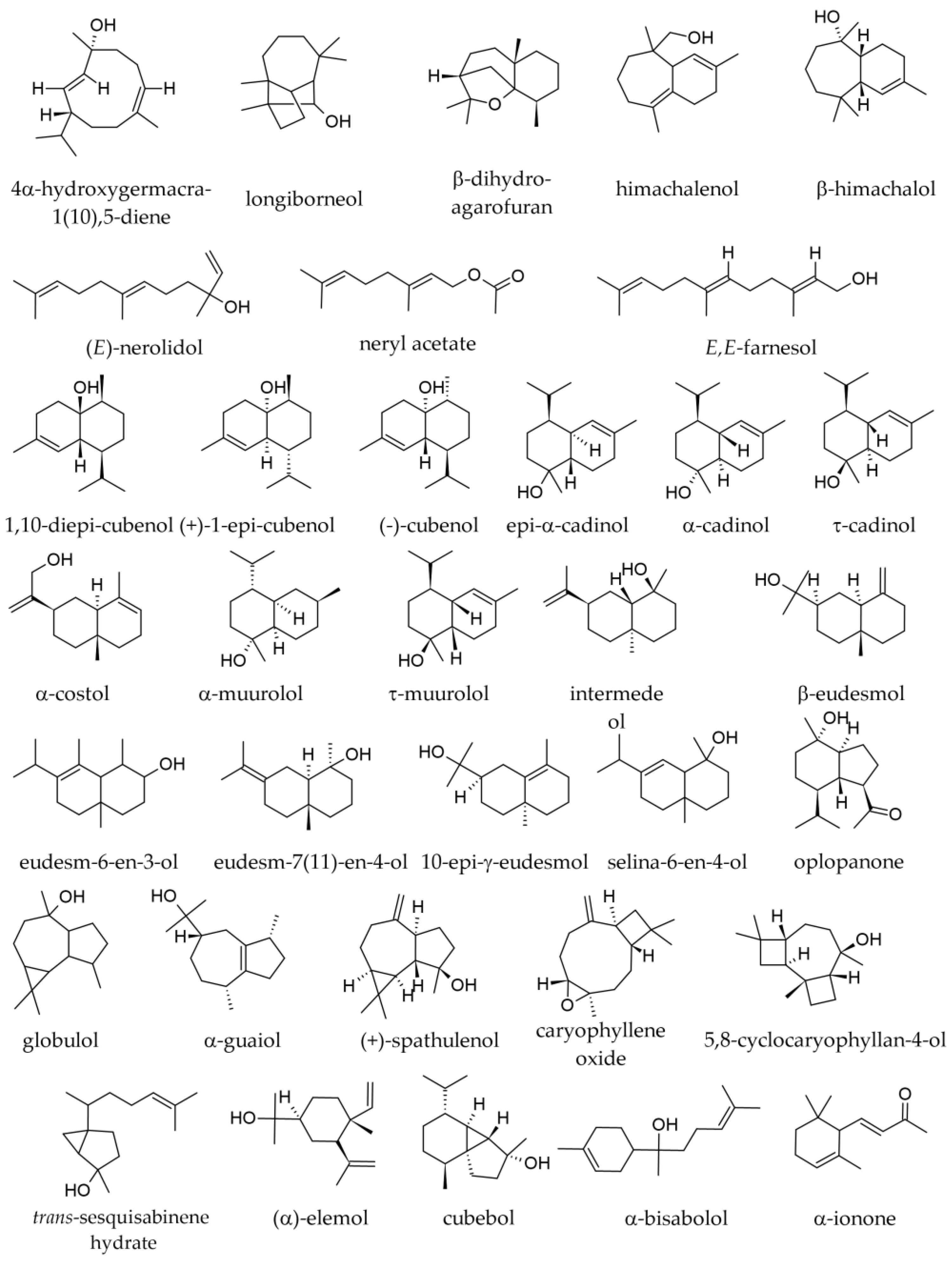
3.1. Terpenoids and Other Components of Essential Oils
3.1.1. Essential Oil from Leaves
3.1.2. Essential Oil from Twigs and Branches
3.1.3. Supercritical Fluid Extracts from Twigs
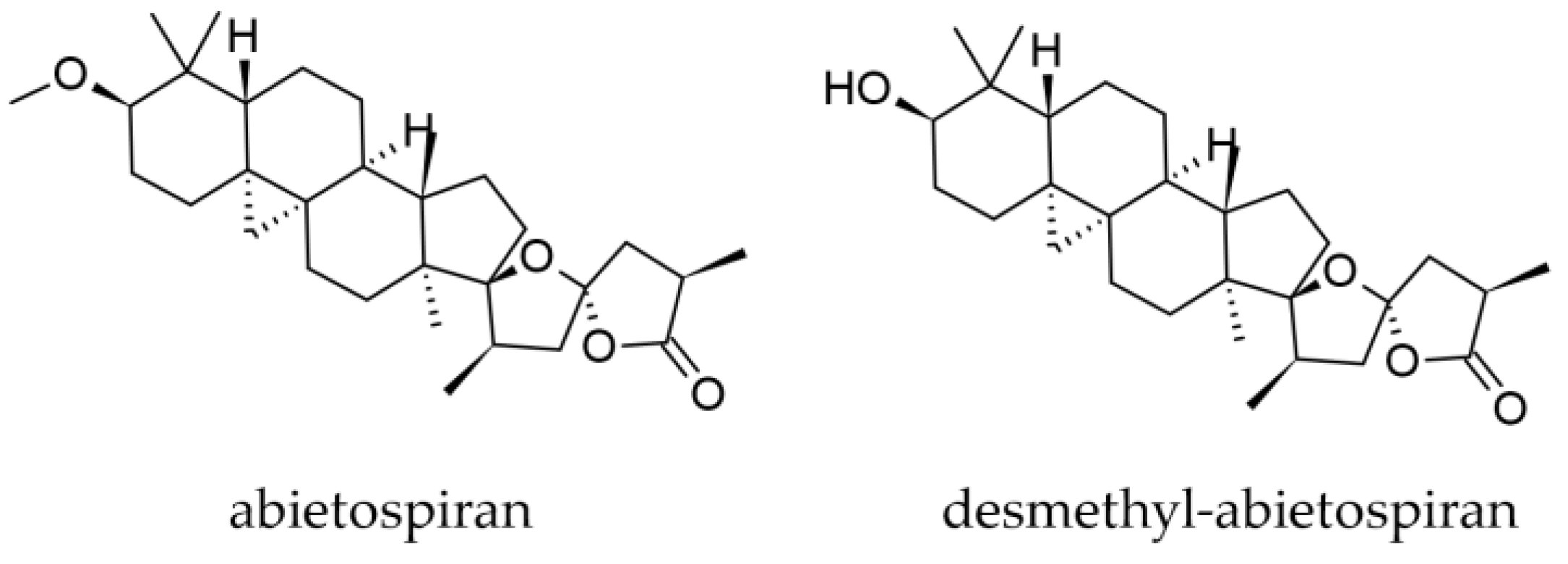
3.1.4. Bark Oleoresin and Bark Triterpenoids
3.1.5. Cone Essential Oils
3.1.6. Seed Essential Oils
3.1.7. Seed Hydrolates
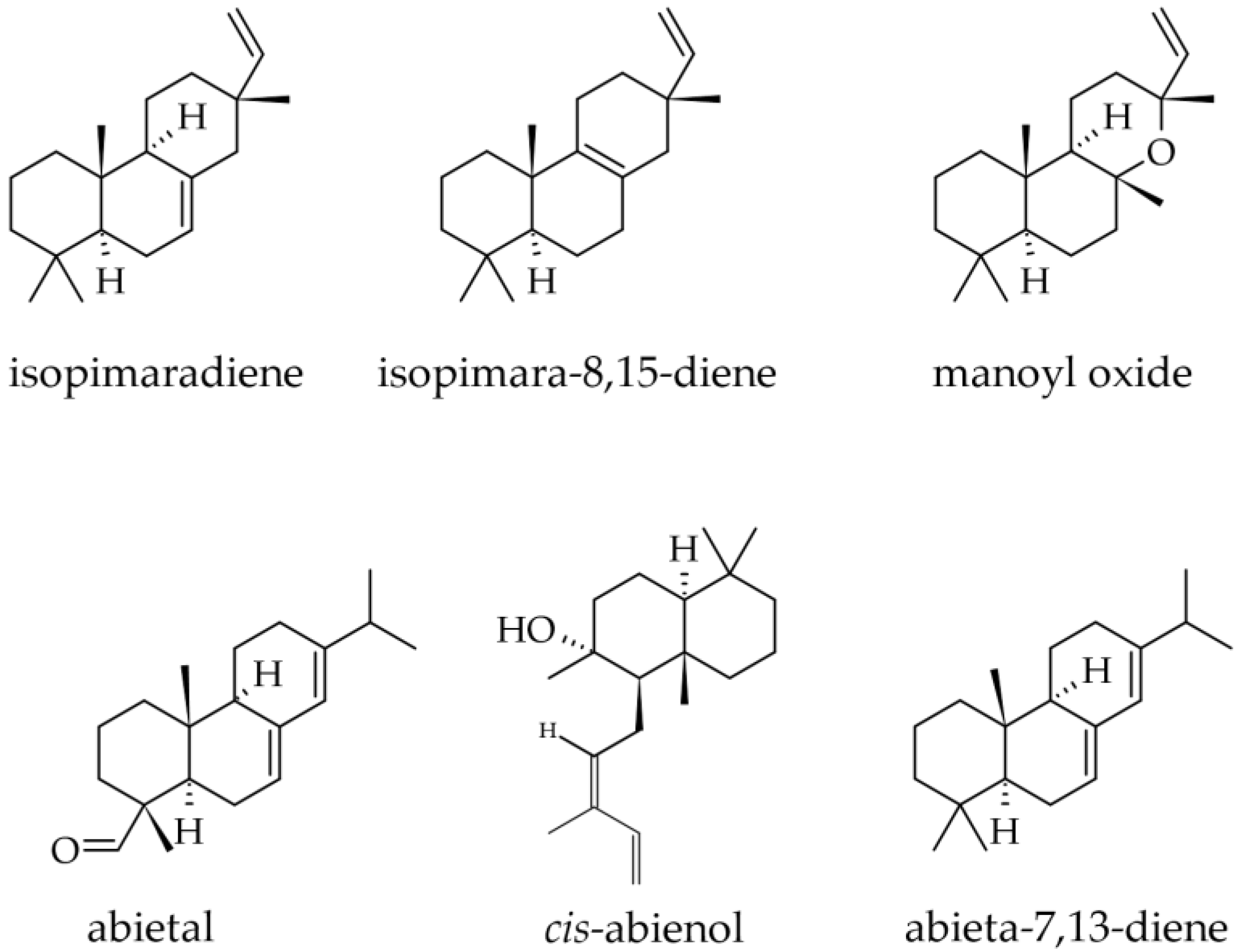
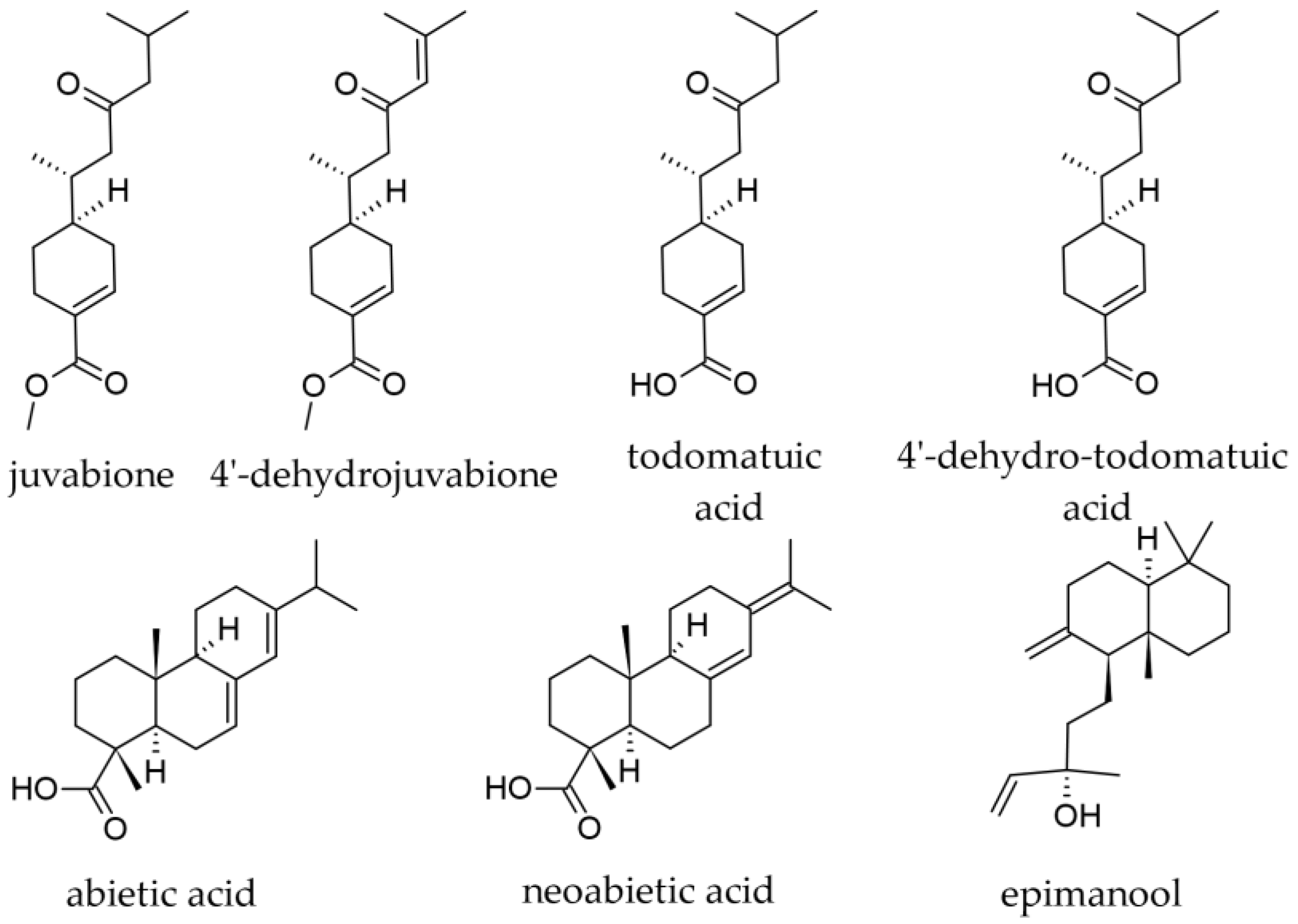
3.1.8. Wood Terpenoids
3.2. Lignans
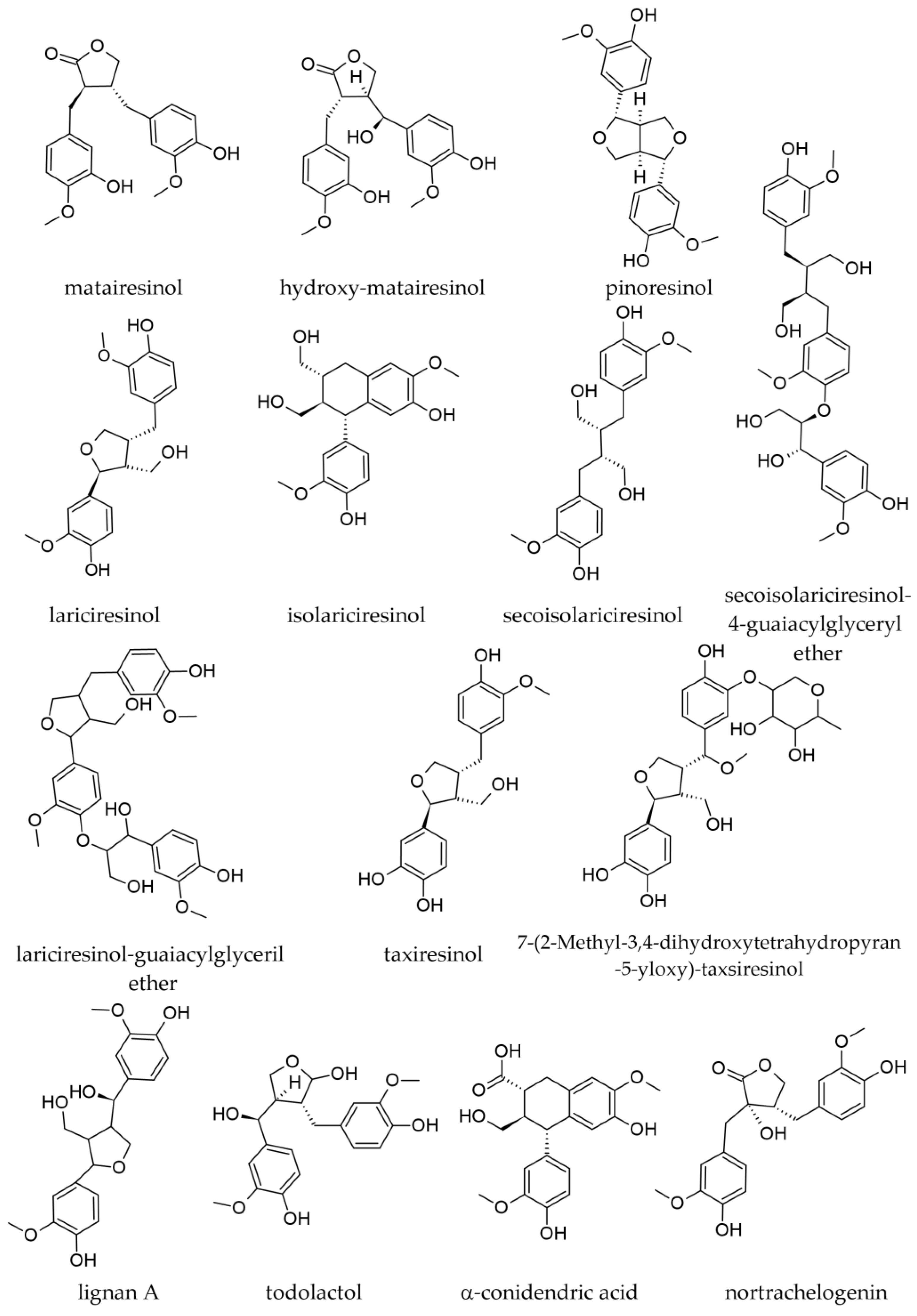
3.2.1. Bark Lignans
3.2.2. Wood Lignans
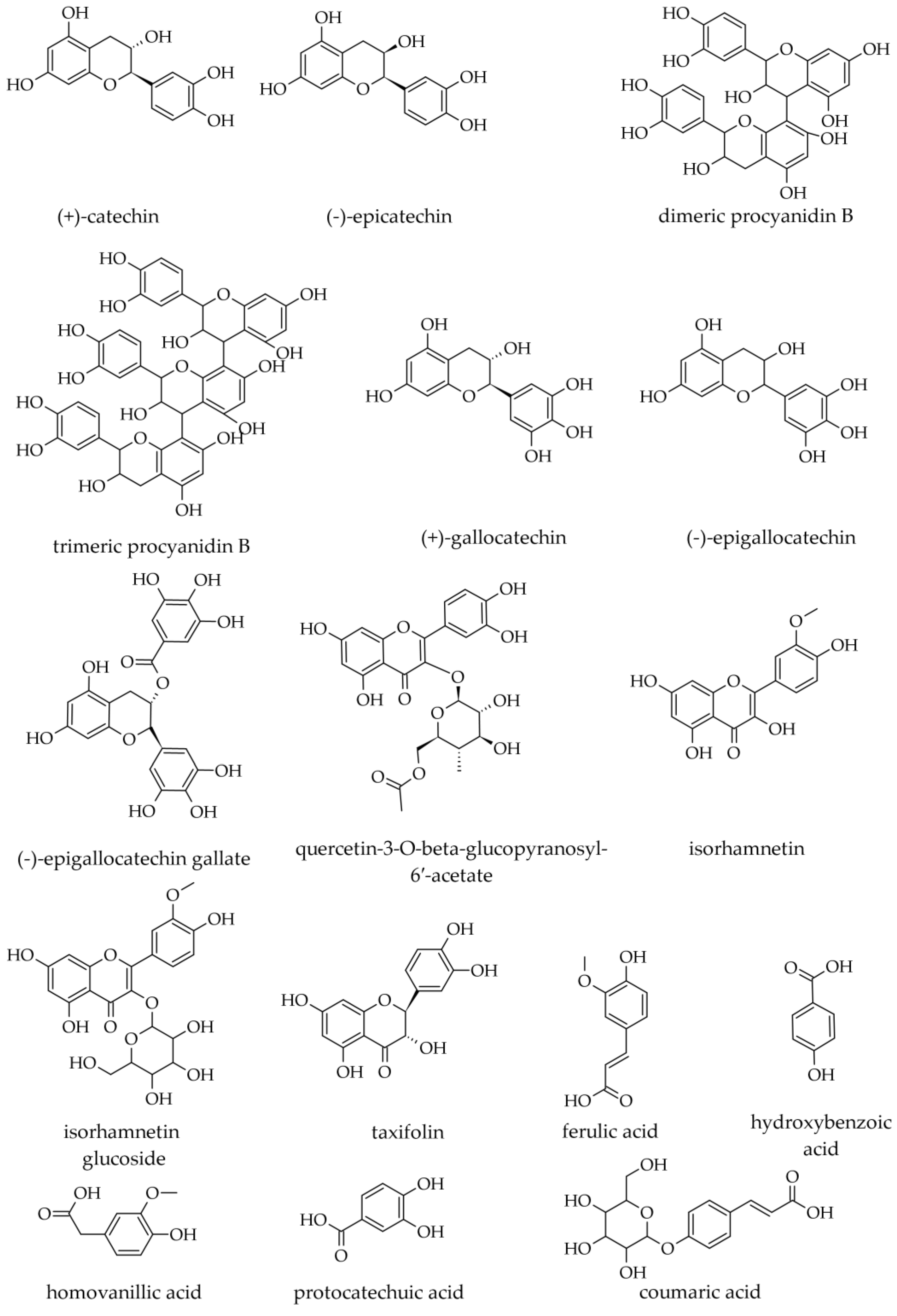
3.3. Phenolics
3.3.1. Bark Phenolic Compounds (Including Flavonoids)
3.3.2. Wood Phenolic Compounds (Including Flavonoids)
3.4. Other Compounds
3.4.1. Lipids and Other Lipophilic Compounds
3.4.2. Carbohydrates
3.4.3. Inorganic Compounds
4. Ethnopharmacology
5. Biological and Pharmacological Effects
5.1. Antioxidant Effects
5.1.1. Antioxidant Effects of Essential Oils
5.1.2. Antioxidant Effects of Various Extracts
5.2. Antimicrobial Activity
5.2.1. Antimicrobial Activity of Essential Oils
5.2.2. Antimicrobial Activity of Various A. alba Extracts
5.3. Cytotoxicity and Toxicity
5.3.1. Cytotoxic Activity of A. alba Essential Oils
5.3.2. Cytotoxic Activity of Various A. alba Extracts
5.4. Influence on Chondrogenesis
5.5. Prebiotic Effects
5.6. Antidiabetic Effects
5.7. Hepatic Steatosis
5.8. Cardiovascular Effects
5.9. Anti-Psoriatic Effects
5.10. Antitumour Effects
5.11. Pharmacokinetics
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Duquesnoy, E.; Castola, V.; Casanova, J. Composition and Chemical Variability of the Twig Oil of Abies alba Miller from Corsica. Flavour Fragr. J. 2007, 22, 293–299. [Google Scholar] [CrossRef]
- Nikolić, J.S.; Zlatković, B.K.; Jovanović, S.Č.; Stojanović, G.S.; Marin, P.D.; Mitić, Z.S. Needle Volatiles as Chemophenetic Markers in Differentiation of Natural Populations of Abies alba, A. x borisii-regis, and A. cephalonica. Phytochemistry 2021, 183, 112612. [Google Scholar] [CrossRef] [PubMed]
- WFO. World Flora Online. 2023. Available online: http://www.worldfloraonline.org (accessed on 12 July 2023).
- Benković, E.T.; Grohar, T.; Žigon, D.; Švajger, U.; Janeš, D.; Kreft, S.; Štrukelj, B. Chemical Composition of the Silver Fir (Abies alba) Bark Extract Abigenol® and Its Antioxidant Activity. Ind. Crop. Prod. 2014, 52, 23–28. [Google Scholar] [CrossRef]
- Kersten, B.; Rellstab, C.; Schroeder, H.; Brodbeck, S.; Fladung, M.; Krutovsky, K.V.; Gugerli, F. The Mitochondrial Genome Sequence of Abies alba Mill. Reveals a High Structural and Combinatorial Variation. BMC Genomics 2022, 23, 776. [Google Scholar] [CrossRef]
- Bianchi, S.; Gloess, A.N.; Kroslakova, I.; Mayer, I.; Pichelin, F. Analysis of the Structure of Condensed Tannins in Water Extracts from Bark Tissues of Norway Spruce (Picea abies [Karst.]) and Silver Fir (Abies alba [Mill.]) Using MALDI-TOF Mass Spectrometry. Ind. Crop. Prod. 2014, 61, 430–437. [Google Scholar] [CrossRef]
- Salem, M.Z.M.; Nasser, R.A.; Zeidler, A.; Elansary, H.O.; Aref, I.M.; Böhm, M.; Ali, H.M.; Ahmed, A.I. Methylated Fatty Acids from Heartwood and Bark of Pinus Sylvestris, Abies alba, Picea abies, and Larix decidua: Effect of Strong Acid Treatment. BioResources 2015, 10, 7715–7724. [Google Scholar] [CrossRef]
- European Commission; Joint Research Centre. European Atlas of Forest Tree Species; Publications Office: Luxembourg, 2016. [Google Scholar]
- Farjon, A. A Handbook of the World’s Conifers; Koninklijke Brill NV: Leiden, The Netherlands; Boston, MA, USA, 2010; ISBN 978-90-474-3062-9. [Google Scholar]
- The CABI Encyclopedia of Forest Trees; CAB International, Ed.; CABI: Oxfordshire, UK; Boston, MA, USA, 2013; ISBN 978-1-78064-236-9. [Google Scholar]
- Idzojtic, M. Dendrology: Cones, Flowers, Fruits and Seeds of Europe; Elsevier: Wlatham, MA, USA, 2019; ISBN 978-0-444-64175-5. [Google Scholar]
- Petelka, J.; Plagg, B.; Säumel, I.; Zerbe, S. Traditional Medicinal Plants in South Tyrol (Northern Italy, Southern Alps): Biodiversity and Use. J. Ethnobiol. Ethnomed. 2020, 16, 74. [Google Scholar] [CrossRef]
- Schoss, K.; Benedetič, R.; Kreft, S. The Phenolic Content, Antioxidative Properties and Extractable Substances in Silver Fir (Abies alba Mill.) Branches Decrease with Distance from the Trunk. Plants 2022, 11, 333. [Google Scholar] [CrossRef]
- Parenti, O.; Albanese, L.; Guerrini, L.; Zanoni, B.; Zabini, F.; Meneguzzo, F. Whole Wheat Bread Enriched with Silver Fir (Abies alba Mill.) Needles Extract: Technological and Antioxidant Properties. J. Sci. Food Agric. 2022, 102, 3581–3589. [Google Scholar] [CrossRef]
- Tofan, L.; Paduraru, C.; Amalinei, R.L.M.; Bunia, I.; Miron, A. Valorization of Romanian Silver Fir Tree Bark (Abies alba Mill.) Wastes as Low-Cost Sorbent of Cu(II) Ions from Polluted Waters. Water Sci. Technol. 2016, 74, 2314–2324. [Google Scholar] [CrossRef]
- Albanese, L.; Bonetti, A.; D’Acqui, L.P.; Meneguzzo, F.; Zabini, F. Affordable Production of Antioxidant Aqueous Solutions by Hydrodynamic Cavitation Processing of Silver Fir (Abies alba Mill.) Needles. Foods 2019, 8, 65. [Google Scholar] [CrossRef]
- Butnaru, E.; Pamfil, D.; Stoleru, E.; Brebu, M. Characterization of Bark, Needles and Cones from Silver Fir (Abies alba Mill.) towards Valorization of Biomass Forestry Residues. Biomass Bioenergy 2022, 159, 106413. [Google Scholar] [CrossRef]
- Vek, V.; Šmidovnik, T.; Humar, M.; Poljanšek, I.; Oven, P. Comparison of the Content of Extractives in the Bark of the Trunk and the Bark of the Branches of Silver Fir (Abies alba Mill.). Molecules 2022, 28, 225. [Google Scholar] [CrossRef]
- Zeneli, G.; Tsitsimpikou, C.; Petrakis, P.V.; Naxakis, G.; Habili, D.; Roussis, V. Foliar and Cortex Oleoresin Variability of Silver Fir (Abies alba Mill.) in Albania. Z. Naturforschung C 2001, 56, 531–539. [Google Scholar] [CrossRef]
- Brennan, M.; Fritsch, C.; Cosgun, S.; Dumarcay, S.; Colin, F.; Gérardin, P. Yield and Compositions of Bark Phenolic Extractives from Three Commercially Significant Softwoods Show Intra- and Inter-Specific Variation. Plant Physiol. Biochem. PPB 2020, 155, 346–356. [Google Scholar] [CrossRef]
- Willför, S.; Nisula, L.; Hemming, J.; Reunanen, M.; Holmbom, B. Bioactive Phenolic Substances in Industrially Important Tree Species. Part 2: Knots and Stemwood of Fir Species. Holzforschung 2004, 58, 650–659. [Google Scholar] [CrossRef]
- Vek, V.; Keržič, E.; Poljanšek, I.; Eklund, P.; Humar, M.; Oven, P. Wood Extractives of Silver Fir and Their Antioxidant and Antifungal Properties. Molecules 2021, 26, 6412. [Google Scholar] [CrossRef]
- Brennan, M.; Hentges, D.; Cosgun, S.; Dumarcay, S.; Colin, F.; Gérardin, C.; Gérardin, P. Intraspecific Variability of Quantity and Chemical Composition of Ethanolic Knotwood Extracts along the Stems of Three Industrially Important Softwood Species: Abies alba, Picea abies and Pseudotsuga menziesii. Holzforschung 2021, 75, 168–179. [Google Scholar] [CrossRef]
- Kebbi-Benkeder, Z.; Manso, R.; Gérardin, P.; Dumarçay, S.; Chopard, B.; Colin, F. Knot Extractives: A Model for Analysing the Eco-Physiological Factors That Control the within and between-Tree Variability. Trees 2017, 31, 1619–1633. [Google Scholar] [CrossRef]
- Sirše, M.; Fokter, S.K.; Štrukelj, B.; Zupan, J. Silver Fir (Abies alba L.) Polyphenolic Extract Shows Beneficial Influence on Chondrogenesis In Vitro under Normal and Inflammatory Conditions. Molecules 2020, 25, 4616. [Google Scholar] [CrossRef]
- Stojanov, S.; Ravnikar, M.; Berlec, A.; Kreft, S. Interaction between Silver Fir (Abies alba) Wood Water Extract and Lactobacilli. Pharm. 2021, 76, 614–617. [Google Scholar] [CrossRef]
- Drevenšek, G.; Lunder, M.; Benković, E.T.; Mikelj, A.; Štrukelj, B.; Kreft, S. Silver Fir (Abies alba) Trunk Extract Protects Guinea Pig Arteries from Impaired Functional Responses and Morphology Due to an Atherogenic Diet. Phytomedicine 2015, 22, 856–861. [Google Scholar] [CrossRef] [PubMed]
- Jończyk, J. Study of volatile oil from Abies alba Miller. I. Study of raw material. Acta Pol. Pharm. 1970, 27, 71–77. [Google Scholar] [PubMed]
- Jończyk, J. Study of volatile oil from sprigs of Abies alba Miller. II. Study of monoterpene fractions of oil. Acta Pol. Pharm. 1970, 27, 155–162. [Google Scholar]
- van Wyk, B.-E.; Wink, M. Medicinal Plants of the World: An Illustrated Scientific Guide to Important Medicina Plants and Their Uses, 2nd ed.; First Impression; Timber Press: Portland, OR, USA, 2017; ISBN 978-1-78639-325-8. [Google Scholar]
- Moukhtar, S.; Couret, C.; Rouil, L.; Simon, V. Biogenic Volatile Organic Compounds (BVOCs) Emissions from Abies alba in a French Forest. Sci. Total Environ. 2006, 354, 232–245. [Google Scholar] [CrossRef]
- Dormont, L.; Roques, A.; Malosse, C. Cone and Foliage Volatiles Emitted by Pinus Cembra and Some Related Conifer Species. Phytochemistry 1998, 49, 1269–1277. [Google Scholar] [CrossRef]
- Wajs-Bonikowska, A.; Sienkiewicz, M.; Stobiecka, A.; Maciąg, A.; Szoka, Ł.; Karna, E. Chemical Composition and Biological Activity of Abies alba and A. koreana Seed and Cone Essential Oils and Characterization of Their Seed Hydrolates. Chem. Biodivers. 2015, 12, 407–418. [Google Scholar] [CrossRef]
- Chalchat, J.-C.; Sidibé, L.; Maksimovic, Z.; Petrovic, S.; Gorunovic, M. Essential Oil of Abies alba Mill., Pinaceae, from the Pilot Production in Montenegro. J. Essent. Oil Res. 2001, 13, 288–289. [Google Scholar] [CrossRef]
- Yang, S.-A.; Jeon, S.-K.; Lee, E.-J.; Im, N.-K.; Jhee, K.-H.; Lee, S.-P.; Lee, I.-S. Radical Scavenging Activity of the Essential Oil of Silver Fir (Abies alba). J. Clin. Biochem. Nutr. 2009, 44, 253–259. [Google Scholar] [CrossRef]
- Roussis, V.; Couladis, M.; Tzakou, O.; Loukis, A.; Petrakis, P.V.; Dukic, N.M.; Jancic, R. A Comparative Study on the Needle Volatile Constituents of Three Abies Species Grown in South Balkans. J. Essent. Oil Res. 2000, 12, 41–46. [Google Scholar] [CrossRef]
- Garzoli, S.; Masci, V.L.; Caradonna, V.; Tiezzi, A.; Giacomello, P.; Ovidi, E. Liquid and Vapor Phase of Four Conifer-Derived Essential Oils: Comparison of Chemical Compositions and Antimicrobial and Antioxidant Properties. Pharmaceuticals 2021, 14, 134. [Google Scholar] [CrossRef]
- Kubeczka, K.-H.; Schultze, W. Biology and Chemistry of Conifer Oils. Flavour Fragr. J. 1987, 2, 137–148. [Google Scholar] [CrossRef]
- Mitić, Z.S.; Stojanović-Radić, Z.Z.; Jovanović, S.Č.; Cvetković, V.J.; Nikolić, J.S.; Ickovski, J.D.; Mitrović, T.L.; Nikolić, B.M.; Zlatković, B.K.; Stojanović, G.S. Essential Oils of Three Balkan Abies Species: Chemical Profiles, Antimicrobial Activity and Toxicity toward Artemia salina and Drosophila melanogaster. Chem. Biodivers. 2022, 19, e202200235. [Google Scholar] [CrossRef]
- Valková, V.; Ďúranová, H.; Vukovic, N.L.; Vukic, M.; Kluz, M.; Kačániová, M. Assessment of Chemical Composition and Anti-Penicillium Activity of Vapours of Essential Oils from Abies alba and Two Melaleuca Species in Food Model Systems. Molecules 2022, 27, 3101. [Google Scholar] [CrossRef]
- Lanzerstorfer, A.; Hackl, M.; Schlömer, M.; Rest, B.; Deutsch-Grasl, E.; Lanzerstorfer, C. The Influence of Air-Dispersed Essential Oils from Lemon (Citrus limon) and Silver Fir (Abies alba) on Airborne Bacteria and Fungi in Hospital Rooms. J. Environ. Sci. Health Part A 2019, 54, 256–260. [Google Scholar] [CrossRef]
- De Groot, A.C.; Schmidt, E. Essential Oils Contact Allergy and Chemical Composition; Routledge: London, UK, 2021; ISBN 978-1-00-320435-0. [Google Scholar]
- Duquesnoy, E.; Marongiu, B.; Castola, V.; Piras, A.; Porcedda, S.; Casanova, J. Combined Analysis by GC (RI), GC-MS and 13C NMR of the Supercritical Fluid Extract of Abies alba Twigs. Nat. Prod. Commun. 2010, 5, 1995–1998. [Google Scholar] [CrossRef]
- Salem, M.Z.M.; Zeidler, A.; Böhm, M.; Mohamed, M.E.A.; Ali, H.M. GC/MS Analysis of Oil Extractives from Wood and Bark of Pinus Sylvestris, Abies alba, Picea abies, and Larix decidua. BioResources 2015, 10, 7725–7737. [Google Scholar] [CrossRef]
- Lang, K.J. Abies alba Mill.: Differentiation of Provenances and Provenance Groups by the Monoterpene Patterns in the Cortex Resin of Twigs. Biochem. Syst. Ecol. 1994, 22, 53–63. [Google Scholar] [CrossRef]
- O’Neill, J.A.; Gallagher, O.P.; Devine, K.J.; Jones, P.W.; Maguire, A.R. Desmethylabietospiran, a Naturally Occurring Self-Gelation Agent. J. Nat. Prod. 2005, 68, 125–128. [Google Scholar] [CrossRef]
- Steglich, W.; Klaar, M.; Zechlin, L.; Hecht, H.J. Abietospiran, the Triterpene from the Bark of the White Fir (Abies alba). Angew. Chem. Int. Ed. Engl. 1979, 18, 698. [Google Scholar] [CrossRef]
- Wajs, A.; Urbańska, J.; Zaleśkiewicz, E.; Bonikowski, R. Composition of Essential Oil from Seeds and Cones of Abies alba. Nat. Prod. Commun. 2010, 5, 1291–1294. [Google Scholar] [CrossRef] [PubMed]
- Bazdi, B.; Oller-López, J.L.; Cuerva, J.M.; Oltra, J.E.; Mansour, A.I. Composition of the Essential Oil from the Seeds of Abies Marocana. J. Essent. Oil Res. 2006, 18, 160–161. [Google Scholar] [CrossRef]
- Patyra, A.; Dudek, M.K.; Kiss, A.K. LC-DAD-ESI-MS/MS and NMR Analysis of Conifer Wood Specialized Metabolites. Cells 2022, 11, 3332. [Google Scholar] [CrossRef] [PubMed]
- Kebbi-Benkeder, Z.; Colin, F.; Dumarçay, S.; Gérardin, P. Quantification and Characterization of Knotwood Extractives of 12 European Softwood and Hardwood Species. Ann. For. Sci. 2015, 72, 277–284. [Google Scholar] [CrossRef]
- Tavčar Benković, E.; Žigon, D.; Mihailović, V.; Petelinc, T.; Jamnik, P.; Kreft, S. Identification, in Vitro and in Vivo Antioxidant Activity, and Gastrointestinal Stability of Lignans from Silver Fir (Abies alba) Wood Extract. J. Wood Chem. Technol. 2017, 37, 467–477. [Google Scholar] [CrossRef]
- Bianchi, S.; Kroslakova, I.; Janzon, R.; Mayer, I.; Saake, B.; Pichelin, F. Characterization of Condensed Tannins and Carbohydrates in Hot Water Bark Extracts of European Softwood Species. Phytochemistry 2015, 120, 53–61. [Google Scholar] [CrossRef]
- Brennan, M.; Fritsch, C.; Cosgun, S.; Dumarcay, S.; Colin, F.; Gérardin, P. Quantitative and Qualitative Composition of Bark Polyphenols Changes Longitudinally with Bark Maturity in Abies alba Mill. Ann. For. Sci. 2020, 77, 9. [Google Scholar] [CrossRef]
- Leone, K.; Micheletto, M.; Di Maira, G.; Tedesco, E.; Benetti, F.; Zaloker, U. Role of a Novel Silver Fir (Abies alba) Extract, Abigenol®/AlbiPhenol®, in Modulating Cardiovascular Disorders: Key Factors. Antioxidants 2022, 11, 618. [Google Scholar] [CrossRef]
- Wajs-Bonikowska, A.; Smeds, A.; Willför, S. Chemical Composition and Content of Lipophilic Seed Extractives of Some Abies and Picea Species. Chem. Biodivers. 2016, 13, 1194–1201. [Google Scholar] [CrossRef]
- Bianchi, S.; Giovannini, L. Inhibition of MTOR/S6K1/4E-BP1 Signaling by Nutraceutical SIRT1 Modulators. Nutr. Cancer 2018, 70, 490–501. [Google Scholar] [CrossRef]
- Novotný, R.; Černý, D.; Šrámek, V. Nutrition of Silver Fir (Abies alba Mill) Growing at the Upper Limit of Its Occurrence in the Šumava National Park and Protected Landscape Area. J. For. Sci. 2010, 56, 381–388. [Google Scholar] [CrossRef]
- Szymura, T.H. Concentration of Elements in Silver Fir (Abies alba Mill.) Needles as a Function of Needles’ Age. Trees 2009, 23, 211–217. [Google Scholar] [CrossRef]
- Durzan, D.J. Arginine, Scurvy and Cartier’s “Tree of Life”. J. Ethnobiol. Ethnomedicine 2009, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Radulescu, V.; Ilies, D.-C.; Voiculescu, I.; Iovu-Adrian, B.; Craciunescu, A. Determination of Ascorbic Acid in Shoots from Different Coniferous Species by HPLC. Farmacia 2013, 61, 1158–1166. [Google Scholar]
- Adams, M.; Berset, C.; Kessler, M.; Hamburger, M. Medicinal Herbs for the Treatment of Rheumatic Disorders—A Survey of European Herbals from the 16th and 17th Century. J. Ethnopharmacol. 2009, 121, 343–359. [Google Scholar] [CrossRef]
- Papp, N.; Purger, D.; Czigle, S.; Czégényi, D.; Stranczinger, S.; Tóth, M.; Dénes, T.; Kocsis, M.; Takácsi-Nagy, A.; Filep, R. The Importance of Pine Species in the Ethnomedicine of Transylvania (Romania). Plants 2022, 11, 2331. [Google Scholar] [CrossRef]
- Jarić, S.; Kostić, O.; Mataruga, Z.; Pavlović, D.; Pavlović, M.; Mitrović, M.; Pavlović, P. Traditional Wound-Healing Plants Used in the Balkan Region (Southeast Europe). J. Ethnopharmacol. 2018, 211, 311–328. [Google Scholar] [CrossRef]
- Mazzei, R.; De Marco, E.V.; Gallo, O.; Tagarelli, G. Italian Folk Plant-Based Remedies to Heal Headache (XIX–XX Century). J. Ethnopharmacol. 2018, 210, 417–433. [Google Scholar] [CrossRef]
- Monari, S.; Ferri, M.; Salinitro, M.; Tassoni, A. Ethnobotanical Review and Dataset Compiling on Wild and Cultivated Plants Traditionally Used as Medicinal Remedies in Italy. Plants 2022, 11, 2041. [Google Scholar] [CrossRef]
- Gilca, M.; Tiplica, G.S.; Salavastru, C.M. Traditional and Ethnobotanical Dermatology Practices in Romania and Other Eastern European Countries. Clin. Dermatol. 2018, 36, 338–352. [Google Scholar] [CrossRef]
- Bonet, M.À.; Vallès, J. Pharmaceutical Ethnobotany in the Montseny Biosphere Reserve (Catalonia, Iberian Peninsula). General Results and New or Rarely Reported Medicinal Plants. J. Pharm. Pharmacol. 2010, 55, 259–270. [Google Scholar] [CrossRef]
- Bacanlı, M.; Başaran, A.A.; Başaran, N. The Antioxidant and Antigenotoxic Properties of Citrus Phenolics Limonene and Naringin. Food Chem. Toxicol. 2015, 81, 160–170. [Google Scholar] [CrossRef]
- Wei, A.; Shibamoto, T. Antioxidant Activities and Volatile Constituents of Various Essential Oils. J. Agric. Food Chem. 2007, 55, 1737–1742. [Google Scholar] [CrossRef]
- Kačániová, M.; Vukovič, N.; Horská, E.; Salamon, I.; Bobková, A.; Hleba, L.; Fiskelová, M.; Vatľák, A.; Petrová, J.; Bobko, M. Antibacterial Activity against Clostridium Genus and Antiradical Activity of the Essential Oils from Different Origin. J. Environ. Sci. Health B 2014, 49, 505–512. [Google Scholar] [CrossRef]
- Vasincu, A.; Creţu, E.; Geangalău, I.; Amalinei, R.L.M.; Miron, A. Polyphenolic Content and Antioxidant Activity of an Extractive Fraction from Abies alba Bark. Rev. Med. Chir. Soc. Med. Nat. Iasi 2013, 117, 545–550. [Google Scholar]
- Cunningham, E. What Has Happened to the ORAC Database? J. Acad. Nutr. Diet. 2013, 113, 740. [Google Scholar] [CrossRef]
- Lunder, M.; Roškar, I.; Hošek, J.; Štrukelj, B. Silver Fir (Abies alba) Extracts Inhibit Enzymes Involved in Blood Glucose Management and Protect against Oxidative Stress in High Glucose Environment. Plant Foods Hum. Nutr. 2019, 74, 47–53. [Google Scholar] [CrossRef]
- Bağci, E.; Diğrak, M. Antimicrobial Activity of Essential Oils of Some Abies (Fir) Species from Turkey. Flavour Fragr. J. 1996, 11, 251–256. [Google Scholar] [CrossRef]
- Bueno, J. In Vitro Antimicrobial Activity of Natural Products Using Minimum Inhibitory Concentrations: Looking for New Chemical Entities or Predicting Clinical Response. Med. Aromat. Plants 2012, 1, 1000113. [Google Scholar] [CrossRef]
- Jeršek, B.; Poklar Ulrih, N.; Skrt, M.; Gavarić, N.; Božin, B.; Smole Možina, S. Effects of Selected Essential Oils on the Growth and Production of Ochratoxin A by Penicillium Verrucosum. Arh. Hig. Rada Toksikol. 2014, 65, 199–208. [Google Scholar] [CrossRef]
- Salamon, I.; Kryvtsova, M.; Bucko, D.; Tarawneh, A.H. Chemical Characterization and Antimicrobial Activity of Some Essential Oils after Their Industrial Large-Scale Distillation. J. Microbiol. Biotechnol. Food Sci. 2021, 2021, 984–988. [Google Scholar] [CrossRef]
- Şandru, D.M. Antimicrobial Effect of Escherichia Coli on Essential Oils Derived from Romanian Aromatic Plants. Acta Univ. Cibiniensis Ser. E Food Technol. 2015, 19, 87–92. [Google Scholar] [CrossRef]
- Liu, T.-T.; Yang, T.-S. Antimicrobial Impact of the Components of Essential Oil of Litsea Cubeba from Taiwan and Antimicrobial Activity of the Oil in Food Systems. Int. J. Food Microbiol. 2012, 156, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Di Pasqua, R.; Betts, G.; Hoskins, N.; Edwards, M.; Ercolini, D.; Mauriello, G. Membrane Toxicity of Antimicrobial Compounds from Essential Oils. J. Agric. Food Chem. 2007, 55, 4863–4870. [Google Scholar] [CrossRef] [PubMed]
- Van Vuuren, S.F.; Viljoen, A.M. Antimicrobial Activity of Limonene Enantiomers and 1,8-Cineole Alone and in Combination. Flavour Fragr. J. 2007, 22, 540–544. [Google Scholar] [CrossRef]
- da Silva, A.C.R.; Lopes, P.M.; de Azevedo, M.M.B.; Costa, D.C.M.; Alviano, C.S.; Alviano, D.S. Biological Activities of A-Pinene and β-Pinene Enantiomers. Molecules 2012, 17, 6305–6316. [Google Scholar] [CrossRef]
- Nissen, L.; Zatta, A.; Stefanini, I.; Grandi, S.; Sgorbati, B.; Biavati, B.; Monti, A. Characterization and Antimicrobial Activity of Essential Oils of Industrial Hemp Varieties (Cannabis sativa L.). Fitoterapia 2010, 81, 413–419. [Google Scholar] [CrossRef]
- Hachlafi, N.E.; Aanniz, T.; Menyiy, N.E.; Baaboua, A.E.; Omari, N.E.; Balahbib, A.; Shariati, M.A.; Zengin, G.; Fikri-Benbrahim, K.; Bouyahya, A. In Vitro and in Vivo Biological Investigations of Camphene and Its Mechanism Insights: A Review. Food Rev. Int. 2021, 39, 1799–1826. [Google Scholar] [CrossRef]
- Runyoro, D.; Ngassapa, O.; Vagionas, K.; Aligiannis, N.; Graikou, K.; Chinou, I. Chemical Composition and Antimicrobial Activity of the Essential Oils of Four Ocimum Species Growing in Tanzania. Food Chem. 2010, 119, 311–316. [Google Scholar] [CrossRef]
- Raman, A.; Weir, U.; Bloomfield, S.F. Antimicrobial Effects of Tea-Tree Oil and Its Major Components on Staphylococcus aureus, Staph. Epidermidis and Propionibacterium Acnes. Lett. Appl. Microbiol. 1995, 21, 242–245. [Google Scholar] [CrossRef]
- Cosentino, S.; Tuberoso, C.I.G.; Pisano, B.; Satta, M.; Mascia, V.; Arzedi, E.; Palmas, F. In-Vitro Antimicrobial Activity and Chemical Composition of Sardinian Thymus Essential Oils. Lett. Appl. Microbiol. 1999, 29, 130–135. [Google Scholar] [CrossRef]
- Tabanca, N.; Kırımer, N.; Demirci, B.; Demirci, F.; Başer, K.H.C. Composition and Antimicrobial Activity of the Essential Oils of Micromeria cristata subsp. Phrygia and the Enantiomeric Distribution of Borneol. J. Agric. Food Chem. 2001, 49, 4300–4303. [Google Scholar] [CrossRef]
- Nikolova, M.T.; Yordanov, P.; Slavov, S.; Berkov, S. Antifungal Activity of Plant Extracts against Phytopathogenic Fungi: Antifungal Activity of Plant Extracts. J. Biosci. Biotechnol. 2017, 6, 155–161. [Google Scholar]
- Valimaa, A.; Honkalampihamalainen, U.; Pietarinen, S.; Willfor, S.; Holmbom, B.; Vonwright, A. Antimicrobial and Cytotoxic Knotwood Extracts and Related Pure Compounds and Their Effects on Food-Associated Microorganisms. Int. J. Food Microbiol. 2007, 115, 235–243. [Google Scholar] [CrossRef]
- Kim, K.-T.; Rioux, L.-E.; Turgeon, S.L. Alpha-Amylase and Alpha-Glucosidase Inhibition Is Differentially Modulated by Fucoidan Obtained from Fucus Vesiculosus and Ascophyllum Nodosum. Phytochemistry 2014, 98, 27–33. [Google Scholar] [CrossRef]
- Debeljak, J.; Ferk, P.; Cokolic, M.; Zavratnik, A.; Tavcar Benkovic, E.; Kreft, S.; Strukelj, B. Randomised, Double Blind, Cross-over, Placebo and Active Controlled Human Pharmacodynamic Study on the Influence of Silver Fir Wood Extract (Belinal) on Post-Prandial Glycemic Response. Pharmazie 2016, 71, 566–569. [Google Scholar] [CrossRef]
- Drevenšek, G.; Lunder, M.; Benković, E.T.; Štrukelj, B.; Kreft, S. Cardioprotective Effects of Silver Fir (Abies alba) Extract in Ischemic-Reperfused Isolated Rat Hearts. Food Nutr. Res. 2016, 60, 29623. [Google Scholar] [CrossRef]
- Zorko, M.S.; Štrukelj, B.; Švajger, U.; Kreft, S.; Lunder, T. Efficacy of a Polyphenolic Extract from Silver Fir (Abies alba) Bark on Psoriasis: A Randomised, Double-Blind, Placebo-Controlled Trial. Pharm. 2018, 73, 56–60. [Google Scholar] [CrossRef]
- Karkabounas, S.; Assimakopoulos, D.; Malamas, M.; Skaltsounis, A.L.; Leonce, S.; Zelovitis, J.; Stefanou, D.; Evangelou, A. Antiproliferative and Anticarcinogenic Effects of an Aqueous Preparation of Abies alba and Viscum Album Se Abies, on a L-1210 Malignant Cell Line and Tumor-Bearing Wistar Rats. Anticancer Res. 2000, 20, 4391–4395. [Google Scholar]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
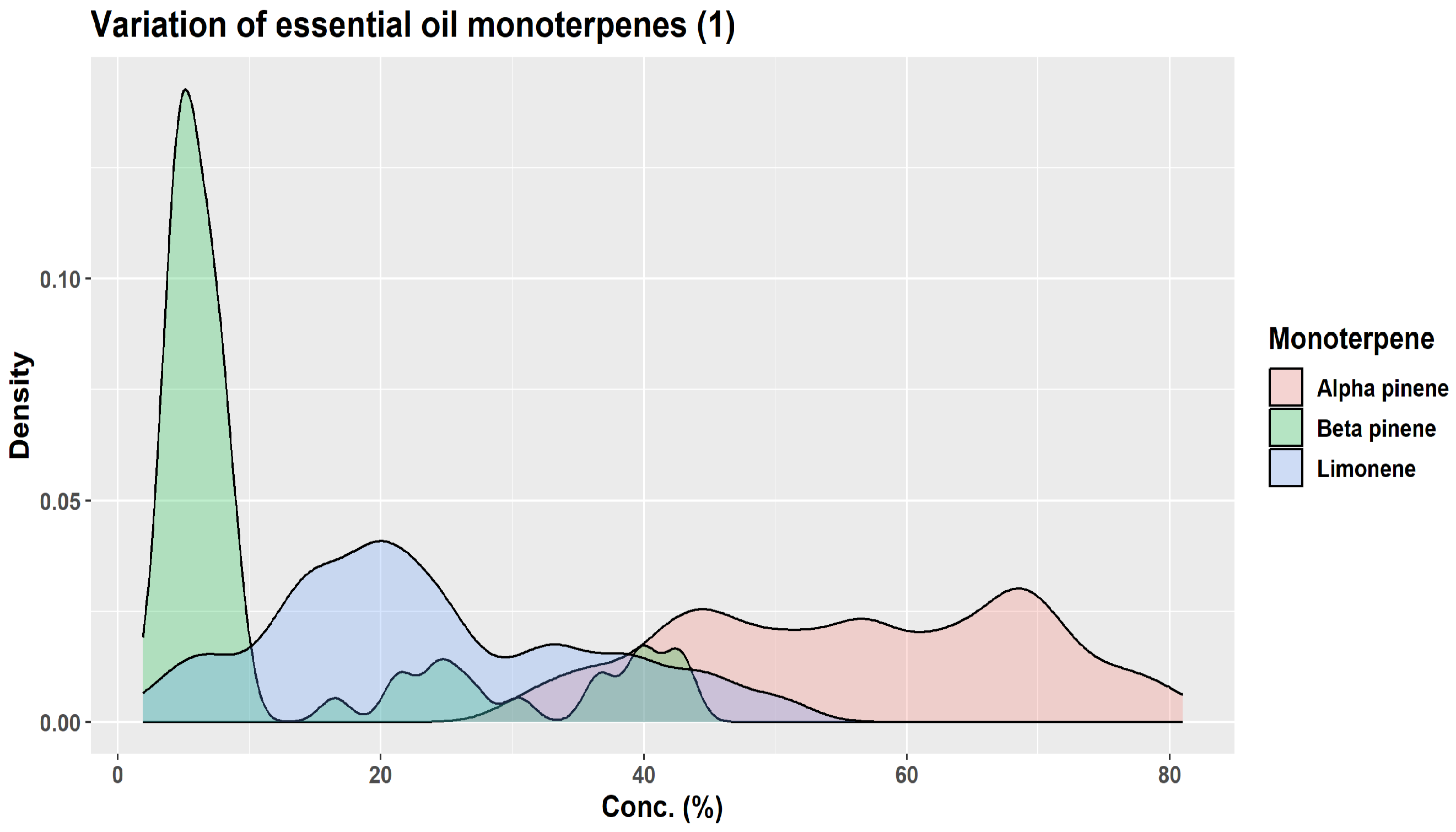
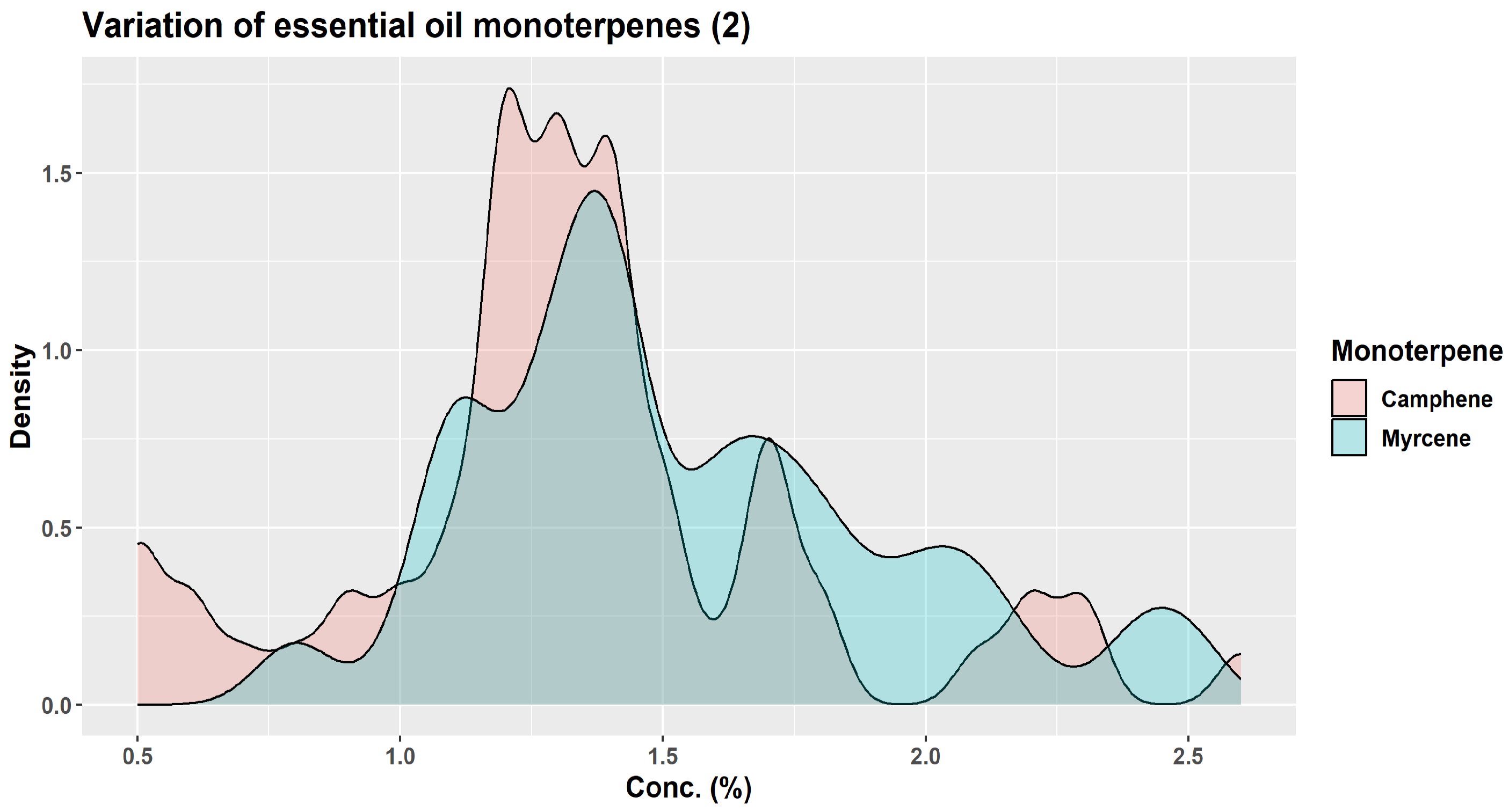
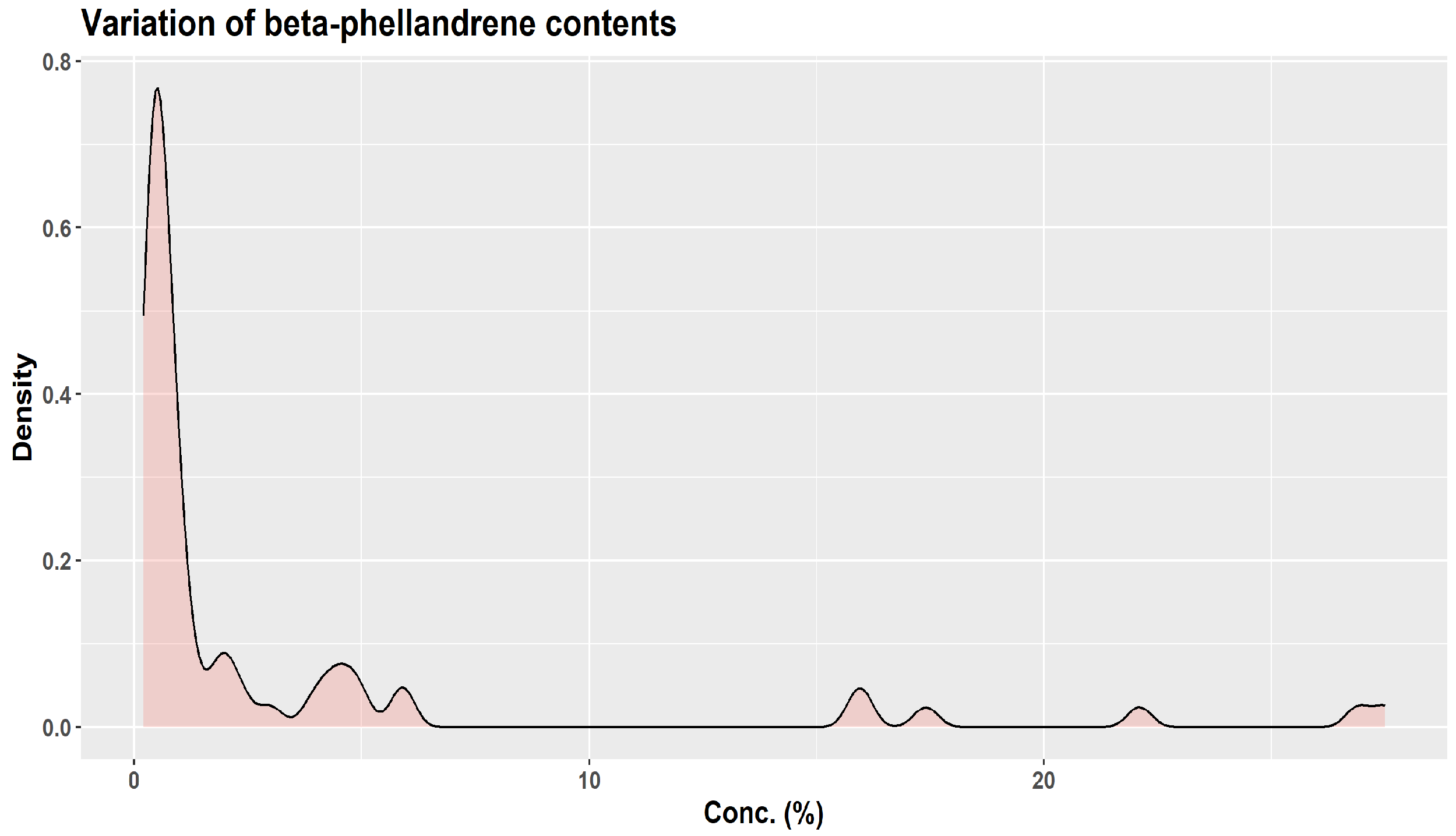
| Monoterpene | Proportion in Winter Samples (%) | Proportion in Summer Samples (%) |
|---|---|---|
| β-pinene | 24.7–31.6 | 14.5–18.4 |
| camphene | 10.8–17.2 | 3.0–5.4 |
| limonene | 9.6–17.5 | 6.5–10.2 |
| α-pinene | 9.5–11.7 | 8.4–15.2 |
| santene | 2.1–3.2 | 0.3–1.0 |
| tricyclene | 1.3–1.8 | 0.3–0.7 |
| myrcene | 1.0–1.2 | 0.6–0.8 |
| α-terpinolene | 0.6–0.8 | 0.8–1.7 |
| α-phellandrene | 0.1–0.2 | 0.1–0.2 |
| Z-ß-ocimene | traces—0.2 | traces |
| Oxygenated Monoterpene | Proportion in Winter Samples (%) | Proportion in Summer Samples (%) |
|---|---|---|
| Bornyl acetate | 5.68–17.6% | 1.6–2.4% |
| α-Terpineol | 1.21–2.0% | 1.21–2.0% |
| Borneol | 0.4–1.7% | 2.3–5.5% |
| α-Terpinyl acetate | 0.35–1.4% | 0.2–0.4% |
| Linalool acetate | 0.1–1.0% | Traces |
| Linalool | 0.1–0.92% | traces—0.2% |
| Camphene hydrate | traces—0.2% | 0.2–0.4% |
| p-Menth-1-en-9-ol-acetate | traces—0.2% | 0.2–0.7% |
| α-Campholenal | traces—0.1% | 0.2–0.5% |
| Camphor | traces—0.1% | 0.1–0.2% |
| cis-Pinocamphone | traces—0.1% | 0.1–0.3% |
| Terpin-4-ol | traces—0.1% | traces—0.4% |
| Endo-fenchol | Traces | 0.2–0.4% |
| Trans-pinocarveol | Traces | 0.4–1.0% |
| Sesquiterpene Hydrocarbon | Proportion in Winter Samples (%) | Proportion in Summer Samples (%) |
|---|---|---|
| E-caryophyllene | 0.6–1.2% | 7.2–12.8% |
| Neryl acetate | absent/traces | 0.2–0.6% |
| α-himachalene | traces—0.2% | traces—0.28% |
| α-humulene | 0.3–0.6% | 4.7–7.2% |
| 9-epi-E-caryophyllene (2-epi-(E)-β-caryophyllene) | traces | traces—0.4% |
| y-gurjunene | traces | traces—0.4% |
| y-muurolene | traces | 0.5–0.9% |
| Germacrene D | 0.1–0.3% | 0.1–0.3% |
| β-selinene | traces—0.2% | 0.6–1.0% |
| cis-β-guaiene | traces—0.4% | 0.3–1.2% |
| α-selinene | 0.0–0.2% | 0.5–1.0% |
| β-himachalene | 0.2–0.4% | 0.7–1.2% |
| Germacrene A | traces—0.1% | traces—0.4% |
| y-cadinene | traces—0.1% | 0.1–1.2% |
| δ-cadinene | 0.3–0.5% | 2.0–3.5% |
| Cadina-1,4-diene | 0.1% | 0.2–0.3% |
| α-cadinene | traces | 0.2–0.3% |
| α-calacorene | traces—0.3% | 0.1–0.3% |
| Compound | Winter Samples (% Composition) | Summer Samples (% Composition) |
|---|---|---|
| E-nerolidol | traces—0.4% | 0.1–0.4% |
| 10-epi-γ-eudesmol | 1.4–5.4% | 0.7–3.9% |
| epi-α-cadinol | traces—0.6% | 0.2–0.3% |
| cubenol | 0.2–0.5% | traces—0.1% |
| himachalol | 0.3–0.8% | traces—0.2% |
| α-cadinol | 0.3–0.42% | 2.7–4.2% |
| E,E-farnesol | traces—0.3% | 0.3–1.0% |
| Monoterpene Hydrocarbon | Proportion in Winter Samples (%) | Proportion in Summer Samples (%) |
|---|---|---|
| α-pinene | 12.2–37.2 | 23.1–47.1 |
| camphene | 0.3–0.9 | 0.4–0.9 |
| β-pinene | 14.9–36.6 | 18.3–30.0 |
| myrcene | 1.4–1.6 | 1.4–2.6 |
| limonene | 6.9–35.3 | 5.5–41.4 |
| α-terpinolene | traces—0.1 | traces |
| Compound | Proportion in Winter Samples (%) | Proportion in Summer Samples (%) |
|---|---|---|
| α-longipinene | traces—0.3% | traces—0.1% |
| neryl acetate | traces—0.2% | traces—0.1% |
| E-caryophyllene | 3.4–8.9% | 3.2–3.9% |
| α-himachalene | traces—0.3% | traces |
| α-humulene | 1.6–3.9 | 1.6–1.9 |
| y-muurolene | 0.3–0.6% | traces—0.1% |
| germacrene D | 5.9–15.2% | 6.9–12.1% |
| β-cis-guaijene | traces—0.7% | traces—0.1% |
| α-selinene | traces—0.3% | traces—0.2% |
| β-himachalene | traces—0.2% | traces—0.1% |
| γ-cadinene | 0.2–0.6% | 0.1–0.3% |
| δ-cadinene | 0.6–1.1% | traces—0.3% |
| α-cadinene | 0.3–0.6% | 0.2–0.4% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ancuceanu, R.; Hovaneț, M.V.; Miron, A.; Anghel, A.I.; Dinu, M. Phytochemistry, Biological, and Pharmacological Properties of Abies alba Mill. Plants 2023, 12, 2860. https://doi.org/10.3390/plants12152860
Ancuceanu R, Hovaneț MV, Miron A, Anghel AI, Dinu M. Phytochemistry, Biological, and Pharmacological Properties of Abies alba Mill. Plants. 2023; 12(15):2860. https://doi.org/10.3390/plants12152860
Chicago/Turabian StyleAncuceanu, Robert, Marilena Viorica Hovaneț, Anca Miron, Adriana Iuliana Anghel, and Mihaela Dinu. 2023. "Phytochemistry, Biological, and Pharmacological Properties of Abies alba Mill." Plants 12, no. 15: 2860. https://doi.org/10.3390/plants12152860
APA StyleAncuceanu, R., Hovaneț, M. V., Miron, A., Anghel, A. I., & Dinu, M. (2023). Phytochemistry, Biological, and Pharmacological Properties of Abies alba Mill. Plants, 12(15), 2860. https://doi.org/10.3390/plants12152860