

Lanthanum Supplementation Alleviates Tomato Root Growth Suppression under Low Light Stress

 , and
, and
Abstract
1. Introduction
2. Results
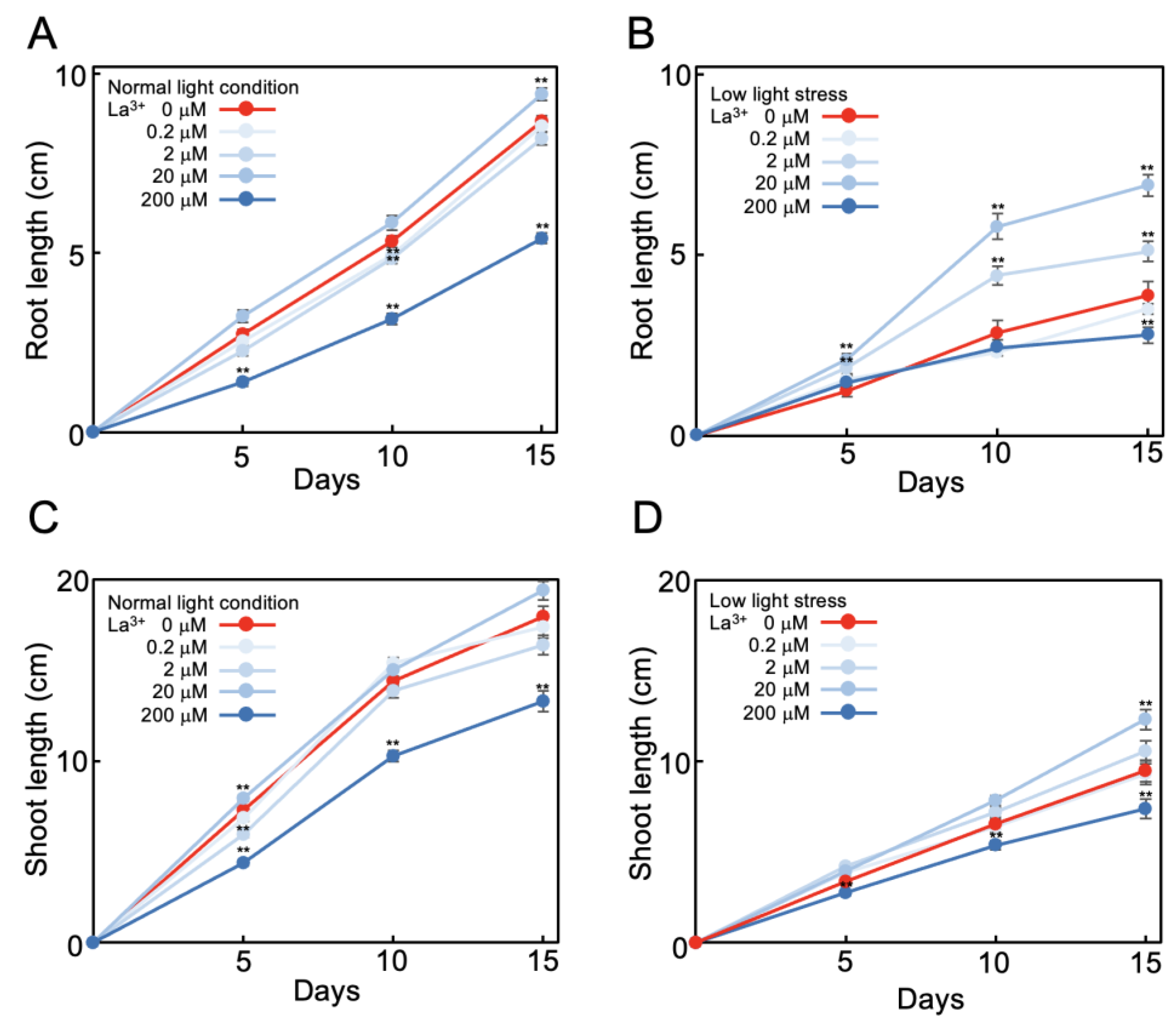
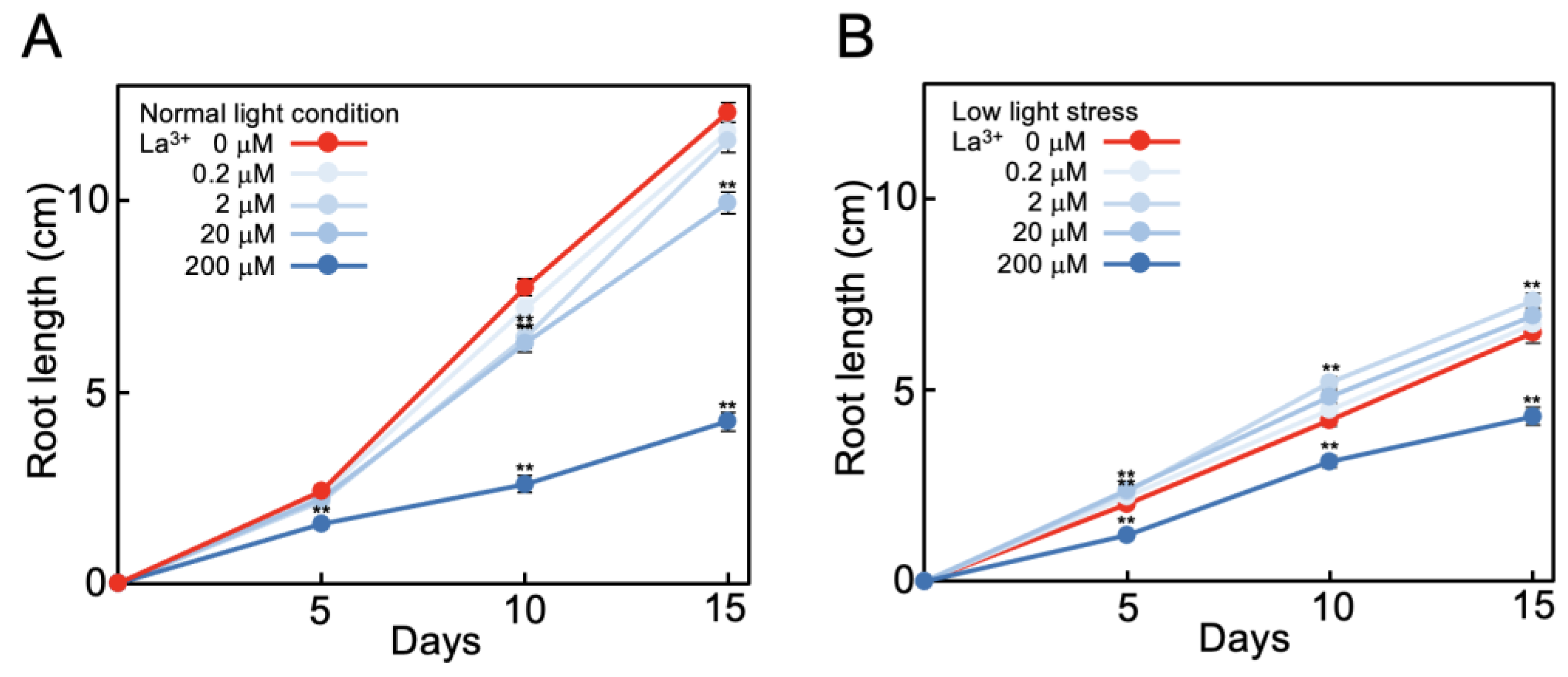
2.1. La3+ Supplementation Alleviates Suppression of Tomato Root Elongation under Low Light
2.2. Effect of La3+ Supplementation on Tomato Plant Growth under Low Light Stress
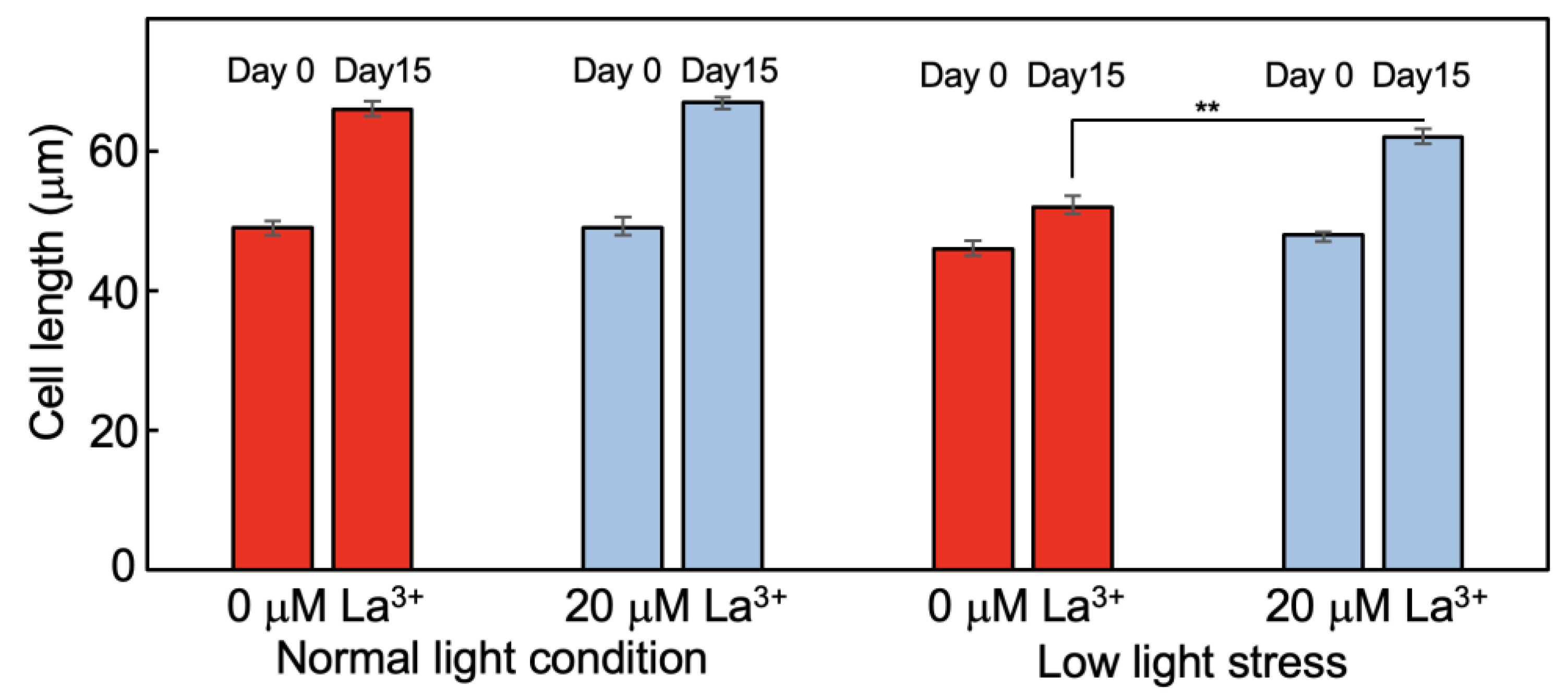
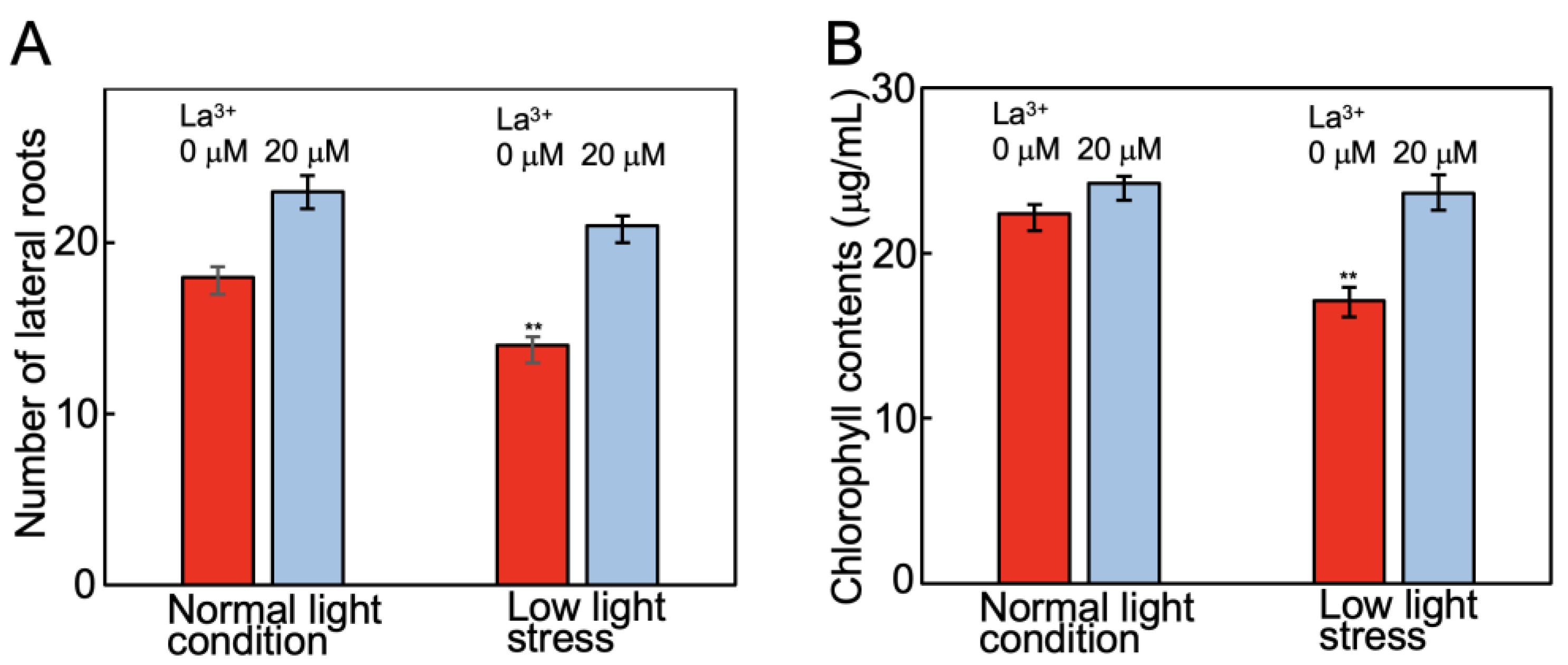
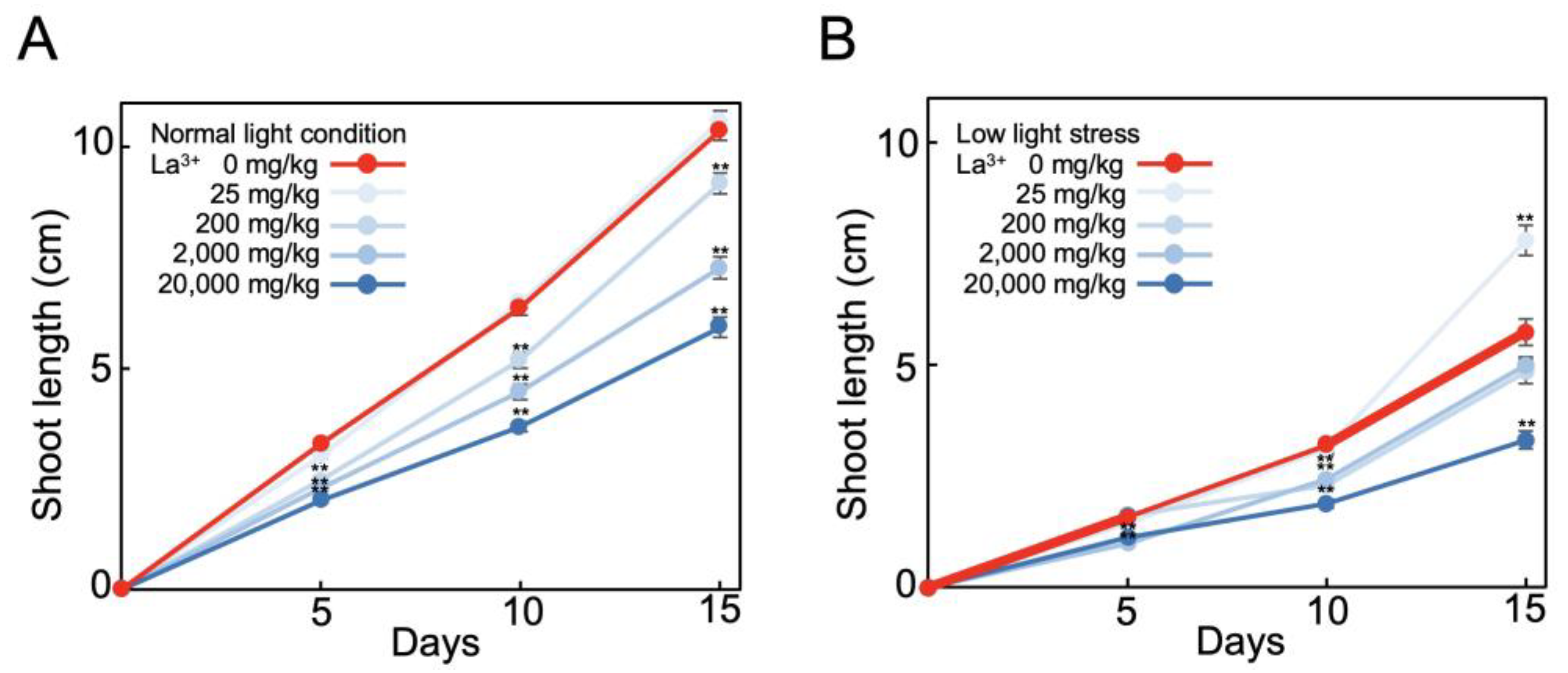
2.3. Analysis of Tomato Plants Grown in La3+-Supplemented Vermiculite
2.4. Effect of La3+ Supplementation on Arabidopsis Root Elongation under Low Light Stress
2.5. Gene Expression of Tomato Roots Grown with La3+ Supplementation under Low Light Stress
3. Discussion
4. Materials and Methods
4.1. Plant Growing Conditions
4.2. Tomato Root and Shoot Lengths and Chlorophyll Content Measurements
4.3. Observation of Tomato Root Cells
4.4. Gene Expression Analysis Using qRT-PCR
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tanaka, Y.; Hosaka, T.; Ochi, K. Rare earth elements activate the secondary metabolite-biosynthetic gene clusters in Streptomyces coelicolor A3(2). J. Antibiot. 2010, 63, 477–481. [Google Scholar] [CrossRef]
- Inaoka, T.; Ochi, K. Scandium stimulates the production of amylase and bacilysin in Bacillus subtilis. Appl. Environ. Microbiol. 2011, 77, 8181–8183. [Google Scholar] [CrossRef]
- Hibi, A.; Asai, K.; Arafuku, H.; Hamajima, M.; Iwama, T.; Kawai, K. Molecular structure of La3+-induced methanol dehydrogenase-like protein in Methylobacterium radiotolerans. J. Biosci. Bioeng. 2011, 111, 547–549. [Google Scholar] [CrossRef]
- Pol, A.; Barends, T.R.M.; Dietl, A.; Khadem, A.F.; Eygensteyn, J.; Jetten, M.S.M.; Op den Camp, H.J.M. Rare earth metals are essential for methanotrophic life in volcanic mudpots. Environ. Microbiol. 2013, 16, 255–264. [Google Scholar] [CrossRef]
- Cotruvo, J.A., Jr.; Featherston, E.R.; Mattocks, J.A.; Ho, J.V.; Laremore, T.N. Lanmodulin: A highly selective lanthanide-binding protein from a lanthanide-utilizing bacterium. J. Am. Chem. Soc. 2018, 140, 15056–15061. [Google Scholar] [CrossRef]
- Kovaříková, M.; Tomášková, I.; Soudek, P. Rare earth elements in plants. Biol. Plant. 2019, 63, 20–32. [Google Scholar] [CrossRef]
- Tommasi, F.; Thomas, P.J.; Pagano, G.; Perono, G.A.; Oral, R.; Lyons, D.M.; Toscanesi, M.; Trifuoggi, M. Review of rare earth elements as fertilizers and feed additives: A knowledge gap analysis. Arch. Environ. Contam. Toxicol. 2021, 81, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Shen, L.; Feng, C.; Yang, R.; Qu, J.; Ju, H.; Zhang, Y. Distribution of rare earth elements (REEs) and their roles in plant growth: A review. Environ. Pollut. 2022, 298, 118540. [Google Scholar] [CrossRef] [PubMed]
- Evans, W.H. The influence of the carbonates of the rare earths (cerium, lanthanum, yttrium) on growth and cell-division in hyacinths. Biochem. J. 1913, 7, 349–355. [Google Scholar] [CrossRef]
- Drobkov, A.A. Influence of cerium, lanthanum, and samarium on development of peas. Proc. USSR Acad. Sci. 1941, 32, 669–670. [Google Scholar]
- Harmet, K.H. Rapid growth responses of Avena coleoptile segments to lanthanum and other cations. Plant Physiol. 1979, 64, 1094–1098. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Li, D.; Peng, A. Application of rare-earth elements in the agriculture of China and its environmental behavior in soil. J. Soils Sediments 2001, 1, 124–129. [Google Scholar] [CrossRef]
- Liu, D.; Wang, X.; Zhang, X.; Gao, Z. Effects of lanthanum on growth and accumulation in roots of rice seedlings. Plant Soil Environ. 2013, 59, 196–200. [Google Scholar] [CrossRef]
- Ramírez-Olvera, S.M.; Trejo-Téllez, L.I.; García-Morales, S.; Pérez-Sato, J.A.; Gómez-Merino, F.C. Cerium enhances germination and shoot growth, and alters mineral nutrient concentration in rice. PLoS ONE 2018, 13, e0194691. [Google Scholar] [CrossRef]
- Diatloff, E.; Asher, C.J.; Smith, F.W. The effects of rare earth elements on the growth and nutrition of plants. Mater. Sci. Forum 1999, 315, 354–360. [Google Scholar] [CrossRef]
- Chen, W.; Tao, Y.; Gu, Y.; Zhao, G. Effect of lanthanide chloride on photosynthesis and dry matter accumulation in tobacco seedlings. Biol. Trace Elem. Res. 2001, 79, 169–176. [Google Scholar] [CrossRef]
- Hu, H.; Wang, L.; Li, Y.; Sun, J.; Zhou, Q.; Huang, X. Insight into mechanism of lanthanum (Ⅲ) induced damage to plant photosynthesis. Ecotoxicol. Environ. Saf. 2016, 127, 43–50. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, R.; Zhang, P.; Sun, L.; Xu, J. Involvement of reactive oxygen species in lanthanum-induced inhibition of primary root growth. J. Exp. Bot. 2016, 67, 6149–6159. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Chen, Y.; Wang, X.; Deng, X. Effects of redox potential and pH value on the release of rare earth elements from soil. Chemosphere 2001, 44, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Chen, A.; Peng, S.; Wei, Z.; Liu, G. Identification and application of amino acids as chelators in phytoremediation of rare earth elements lanthanum and yttrium. Plant Soil 2013, 373, 329–338. [Google Scholar] [CrossRef]
- Lai, Y.; Wang, Q.; Yang, L.; Huang, B. Subcellular distribution of rare earth elements and characterization of their binding species in a newly discovered hyperaccumulator Pronephrium simplex. Talanta 2006, 70, 26–31. [Google Scholar] [CrossRef]
- Wang, L.; Cheng, M.; Yang, Q.; Li, J.; Wang, X.; Zhou, Q.; Nagawa, S.; Xia, B.; Xu, T.; Huang, R.; et al. Arabinogalactan protein-rare earth element complexes activate plant endocytosis. Proc. Natl. Acad. Sci. USA 2019, 116, 14349–14357. [Google Scholar] [CrossRef]
- Song, W.; Hong, F.; Wan, Z.; Zhou, Y. Effects of lanthanum and europium on rooting of plantlet Eriobotrya japonica Lindl. in vitro. J. Rare Earths 2002, 20, 658–662. [Google Scholar]
- Fan, Z.; Zhang, K.; Wang, F.; Zhao, X.; Bai, R.; Liu, B. Effect of rare earth elements on growth and determination of secondary metabolites under in vitro conditions in Salvia miltiorrhiza. HortScience 2020, 55, 310–316. [Google Scholar] [CrossRef]
- Ben, Y.; Cheng, M.; Liu, Y.; Wang, L.; Yang, Q.; Huang, X. The stimulatory effect and mechanism of low-dose lanthanum on soybean leaf cells. J. Hazard. Mater. 2023, 441, 129924. [Google Scholar] [CrossRef]
- Yuan, X.; Wang, Q.; Zhao, B.; Wang, Y. Improved cell growth and total flavonoids of Saussurea medusa on solid culture medium supplemented with rare earth elements. Biotechnol. Lett. 2002, 24, 1889–1892. [Google Scholar] [CrossRef]
- Ouyang, J.; Wang, X.; Zhao, B.; Yuan, X.; Wang, Y. Effects of rare earth elements on the growth of Cistanche deserticola cells and the production of phenylethanoid glycosides. J. Biotechnol. 2003, 102, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, J.; Zhou, Q.; Yang, G.; Ding, X.L.; Li, X.; Cai, C.X.; Zhang, Z.; Wei, H.Y.; Lu, T.H.; et al. Rare earth elements activate endocytosis in plant cells. Proc. Natl. Acad. Sci. USA 2014, 111, 12936–12941. [Google Scholar] [CrossRef]
- Cheng, M.; Wang, L.; Zhou, Q.; Chao, D.; Nagawa, S.; He, D.; Zhang, J.; Li, H.; Tan, L.; Gu, Z.; et al. Lanthanum(Ⅲ) triggers AtrbohD- and jasmonic acid-dependent systemic endocytosis in plants. Nat. Commun. 2021, 12, 4327. [Google Scholar] [CrossRef]
- Du, Y.; Ji, X.; Zhang, J.; Li, J.; Sun, H.; Zhao, Q. Research progress on the impacts of low light intensity on rice growth and development. Chin. J. Eco-Agric. 2013, 21, 1307–1317. [Google Scholar] [CrossRef]
- Lu, T.; Yu, H.; Li, Q.; Chai, L.; Jiang, W. Improving plant growth and alleviating photosynthetic inhibition and oxidative stress from low-light stress with exogenous GR24 in tomato (Solanum lycopersicum L.) seedling. Front. Plant Sci. 2019, 10, 490. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, T.; Hirai, M.; Chino, M.; Komeda, Y.; Naito, S. Effects of sulfur nutrition on expression of the soybean seed storage protein genes in transgenic petunia. Plant Physiol. 1992, 99, 263–268. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Voncken, J.H.L. Physical and chemical properties of the rare earths. In The rare earth elements; Springer: Cham, Switzerland, 2016; pp. 53–72. [Google Scholar]
- Liu, Y.; Zhang, J. Lanthanum promotes bahiagrass (Paspalum notatum) roots growth by improving root activity, photosynthesis and respiration. Plants 2022, 11, 382. [Google Scholar] [CrossRef]
- Song, K.; Gao, J.; Li, S.; Sun, Y.; Sun, H.; An, B.; Hu, T.; He, X. Experimental and theoretical study of the effects of rare earth elements on growth and chlorophyll of alfalfa (Medicago sativa L.) seedling. Front. Plant Sci. 2021, 12, 731838. [Google Scholar] [CrossRef]
- García-Jiménez, A.; Gómez-Merino, F.C.; Tejeda-Sartorius, O.; Trejo-Téllez, L.I. Lanthanum affects bell pepper seeding quality depending on the genotype and time of exposure by differentially modifying plant height, stem diameter and concentrations of chlorophylls, sugars, amino acids, and proteins. Front. Plant Sci. 2017, 8, 308. [Google Scholar] [CrossRef]
- Abou El-Nour, H.H.; Attia, R.M. Evaluate the effects of rare earth elements on sweet pepper seeds germination process, seedlings growth and plants productivity. GSC Adv. Res. Rev. 2022, 13, 23–38. [Google Scholar] [CrossRef]
- He, Y.; Loh, C. Cerium and lanthanum promote floral initiation and reproductive growth of Arabidopsis thaliana. Plant Sci. 2000, 159, 117–124. [Google Scholar] [CrossRef]
- Xiong, S.; Xiong, Z.; Chen, Y.; Huang, H. Interactive effects of lanthanum and cadmium on plant growth and mineral element uptake in crisped-leaf mustard under hydroponic conditions. J. Plant Nutr. 2006, 29, 1889–1902. [Google Scholar] [CrossRef]
- Buckingham, S.; Maheswaran, J.; Meehan, B.; Peverill, K. The role of applications of rare earth elements in enhancement of crop and pasture production. Mater. Sci. Forum 1999, 315, 339–347. [Google Scholar] [CrossRef]
- Reddy, N.; Maheswaran, J.; Meehan, B.; Peverill, K. The application of rare earth elements in enhancement of crop production in Australia, Part 2. In Proceedings of the 4th International Conference on Rare Earth Development and Application; Yu, Z., Yan, C., Xu, G., Niu, J., Chen, Z., Eds.; China Scientific Books: WanChai, Hong Kong, 2001; pp. 251–254. [Google Scholar]
- Franklin, K.A.; Lee, S.H.; Patel, D.; Kumar, S.V.; Spartz, A.K.; Gu, C.; Ye, S.; Yu, P.; Breen, G.; Cohen, J.D.; et al. Phytochrome-interacting factor 4 (PIF4) regulates auxin biosynthesis at high temperature. Proc. Natl. Acad. Sci. USA 2011, 108, 20231–20235. [Google Scholar] [CrossRef]
- Li, L.; Ljung, K.; Breton, G.; Schmitz, R.J.; Pruneda-Paz, J.; Cowing-Zitron, C.; Cole, B.J.; Ivans, L.J.; Pedmale, U.V.; Jung, H.; et al. Linking photoreceptor excitation to changes in plant architecture. Genes Dev. 2012, 26, 785–790. [Google Scholar] [CrossRef]
- Bielach, A.; Hrtyan, M.; Tognetti, V.B. Plants under stress: Involvement of auxin and cytokinin. Int. J. Mol. Sci. 2017, 18, 1427. [Google Scholar] [CrossRef] [PubMed]
- Korver, R.A.; Koevoets, I.T.; Testerink, C. Out of shape during stress: A key role for auxin. Trends Plant Sci. 2018, 23, 783–793. [Google Scholar] [CrossRef]
- Ben, Y.; Cheng, M.; Wang, L.; Zhou, Q.; Yang, Z.; Huang, X. Low-dose lanthanum activates endocytosis, aggravating accumulation of lanthanum or/and lead and disrupting homeostasis of essential elements in the leaf cells of four edible plants. Ecotoxicol. Environ. Saf. 2021, 221, 112429. [Google Scholar] [CrossRef]
- Majda, M.; Robert, S. The role of auxin in cell wall expansion. Int. J. Mol. Sci. 2018, 19, 951. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Iribe, A.; De-la-Peña, C. Auxins, the hidden player in chloroplast development. Plant Cell Rep. 2020, 39, 1595–1608. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Cheng, M.; Chu, Y.; Li, X.; Chen, D.D.Y.; Huang, X.; Zhou, Q. Responses of plant calmodulin to endocytosis induced by rare earth elements. Chemosphere 2016, 154, 408–415. [Google Scholar] [CrossRef]
- Du, M.; Spalding, E.P.; Gray, W.M. Rapid auxin-mediated cell expansion. Annu. Rev. Plant Biol. 2020, 71, 379–402. [Google Scholar] [CrossRef]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: Verification of the concentration of chlorophyll standards by atomic absorption spectroscopy. Biochim. Biophys. Acta 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Takenaka, Y.; Kato, K.; Ogawa-Ohnishi, M.; Tsuruhama, K.; Kajiura, H.; Yagyu, K.; Takeda, A.; Takeda, Y.; Kunieda, T.; Nishimura, I.H.; et al. Pectin RG-I rhamnosyltransferases represent a novel plant-specific glycosyltransferase family. Nat. Plants 2018, 4, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| REE Ion | Optimum Concentration (μM) | Elongation with REE under Low Light Relative to Normal Light, % * | Elongation with REE Relative to No REE under Low Light, % | Atomic Weight | Ion Radius (Å) | Density (g/cm3) | Number of Unpaired 4f Electrons |
|---|---|---|---|---|---|---|---|
| None | – | 45 | 100 | – | – | – | – |
| Sc3+ | 20 | 72 | 161 | 45.0 | 0.75 | 2.99 | 0 |
| Y3+ | 20 | 51 | 114 | 88.9 | 0.90 | 4.47 | 0 |
| La3+ | 20 | 82 | 177 | 138.9 | 1.03 | 6.15 | 0 |
| Ce3+ | 2 | 65 | 144 | 140.1 | 1.01 | 8.16 | 1 |
| Pr3+ | 2 | 64 | 143 | 140.9 | 0.99 | 6.77 | 2 |
| Nd3+ | 2 | 63 | 140 | 144.2 | 0.98 | 7.01 | 3 |
| Pm3+ | not available | – | – | 145.0 | 0.97 | 7.26 | 4 |
| Sm3+ | 2 | 52 | 115 | 150.4 | 0.96 | 7.52 | 5 |
| Eu3+ | 0.2 | 48 | 107 | 151.9 | 0.95 | 5.24 | 6 |
| Gd3+ | 0.2 | 50 | 112 | 157.3 | 0.94 | 7.90 | 7 |
| Tb3+ | 20 | 50 | 112 | 158.9 | 0.92 | 8.23 | 6 |
| Dy3+ | 0.2 | 62 | 138 | 162.5 | 0.91 | 8.55 | 5 |
| Ho3+ | not available | – | – | 164.9 | 0.90 | 8.80 | 4 |
| Er3+ | 20 | 65 | 145 | 167.3 | 0.89 | 9.07 | 3 |
| Tm3+ | 0.2 | 60 | 133 | 168.9 | 0.88 | 9.32 | 2 |
| Yb3+ | 2 | 52 | 116 | 173.0 | 0.87 | 6.97 | 1 |
| Lu3+ | 2 | 52 | 115 | 175.0 | 0.86 | 9.84 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iguchi, S.; Tokunaga, T.; Kamon, E.; Takenaka, Y.; Koshimizu, S.; Watanabe, M.; Ishimizu, T. Lanthanum Supplementation Alleviates Tomato Root Growth Suppression under Low Light Stress. Plants 2023, 12, 2663. https://doi.org/10.3390/plants12142663
Iguchi S, Tokunaga T, Kamon E, Takenaka Y, Koshimizu S, Watanabe M, Ishimizu T. Lanthanum Supplementation Alleviates Tomato Root Growth Suppression under Low Light Stress. Plants. 2023; 12(14):2663. https://doi.org/10.3390/plants12142663
Chicago/Turabian StyleIguchi, Syo, Tatsuya Tokunaga, Eri Kamon, Yuto Takenaka, Shizuka Koshimizu, Masao Watanabe, and Takeshi Ishimizu. 2023. "Lanthanum Supplementation Alleviates Tomato Root Growth Suppression under Low Light Stress" Plants 12, no. 14: 2663. https://doi.org/10.3390/plants12142663
APA StyleIguchi, S., Tokunaga, T., Kamon, E., Takenaka, Y., Koshimizu, S., Watanabe, M., & Ishimizu, T. (2023). Lanthanum Supplementation Alleviates Tomato Root Growth Suppression under Low Light Stress. Plants, 12(14), 2663. https://doi.org/10.3390/plants12142663