
A Study of the Molecular Regulatory Network of VcTCP18 during Blueberry Bud Dormancy

 , , and
, , and
Abstract
1. Introduction
2. Results
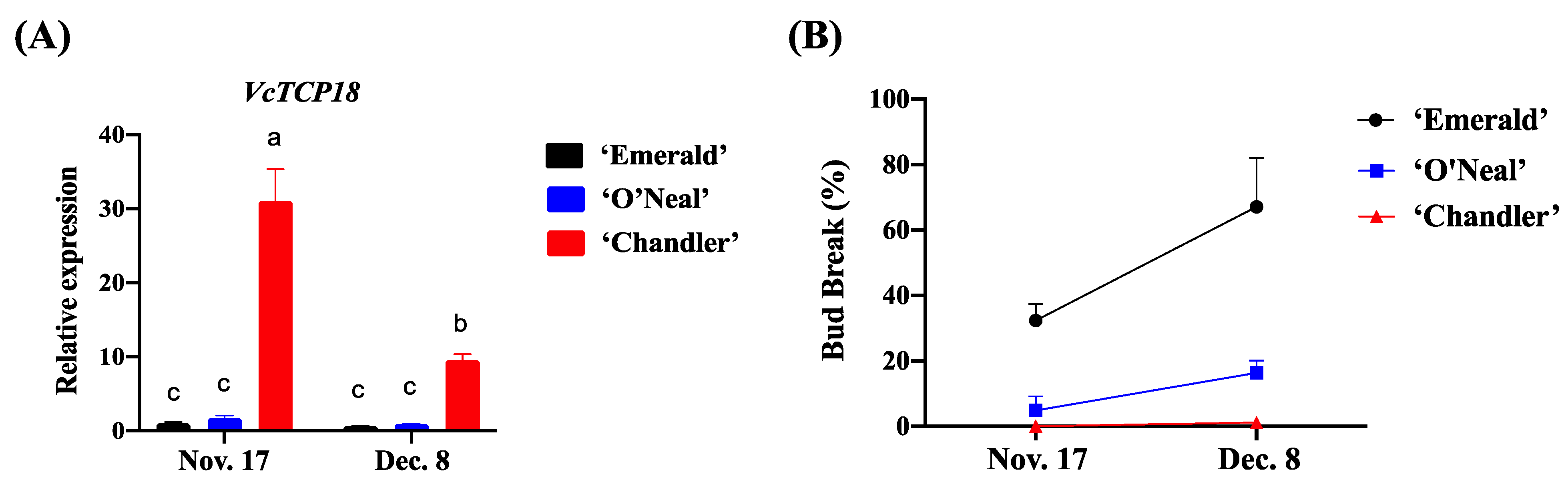
2.1. Expression Patterns of VcTCP18 in the Process of Para- and Endodormancy in Different Blueberry Cultivars
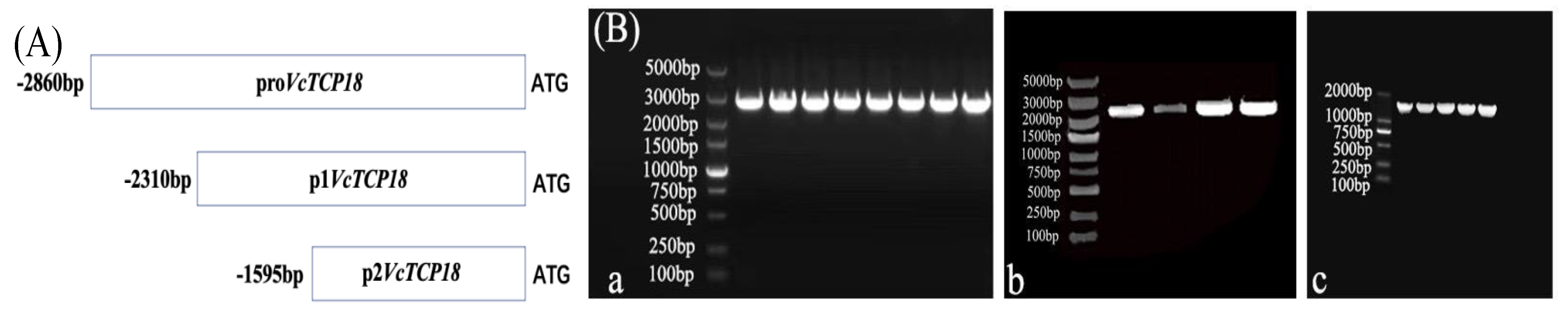
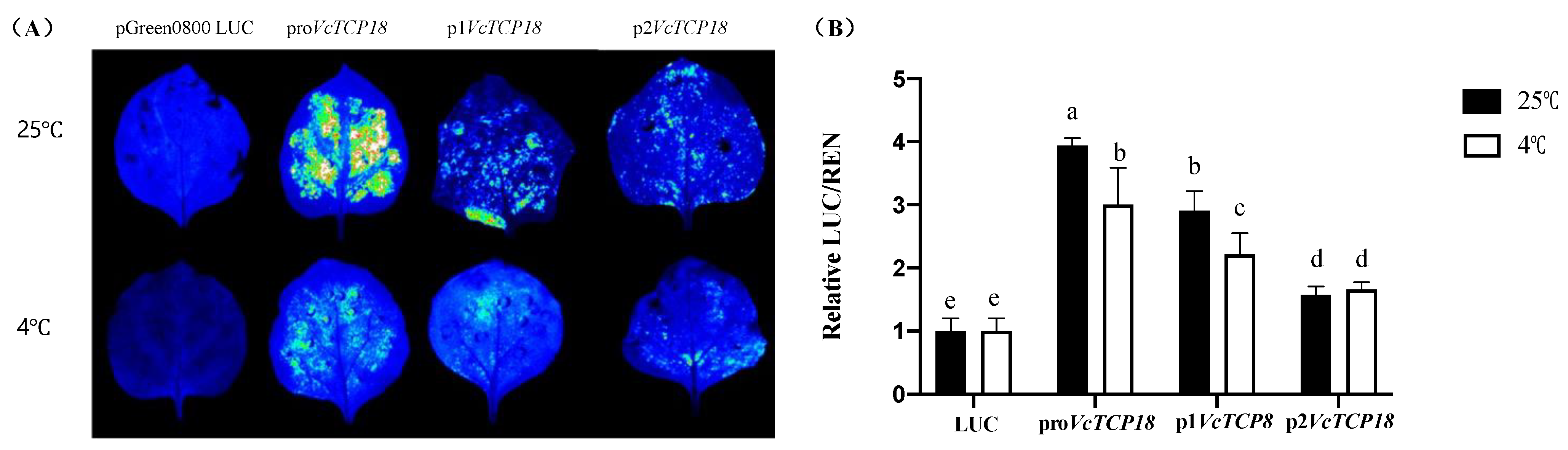
2.2. Promoter Activity Analysis of the VcTCP18 Gene
2.3. Screening of A Yeast One-Hybrid Library
2.4. Verification of the Positive Clones by Rotation
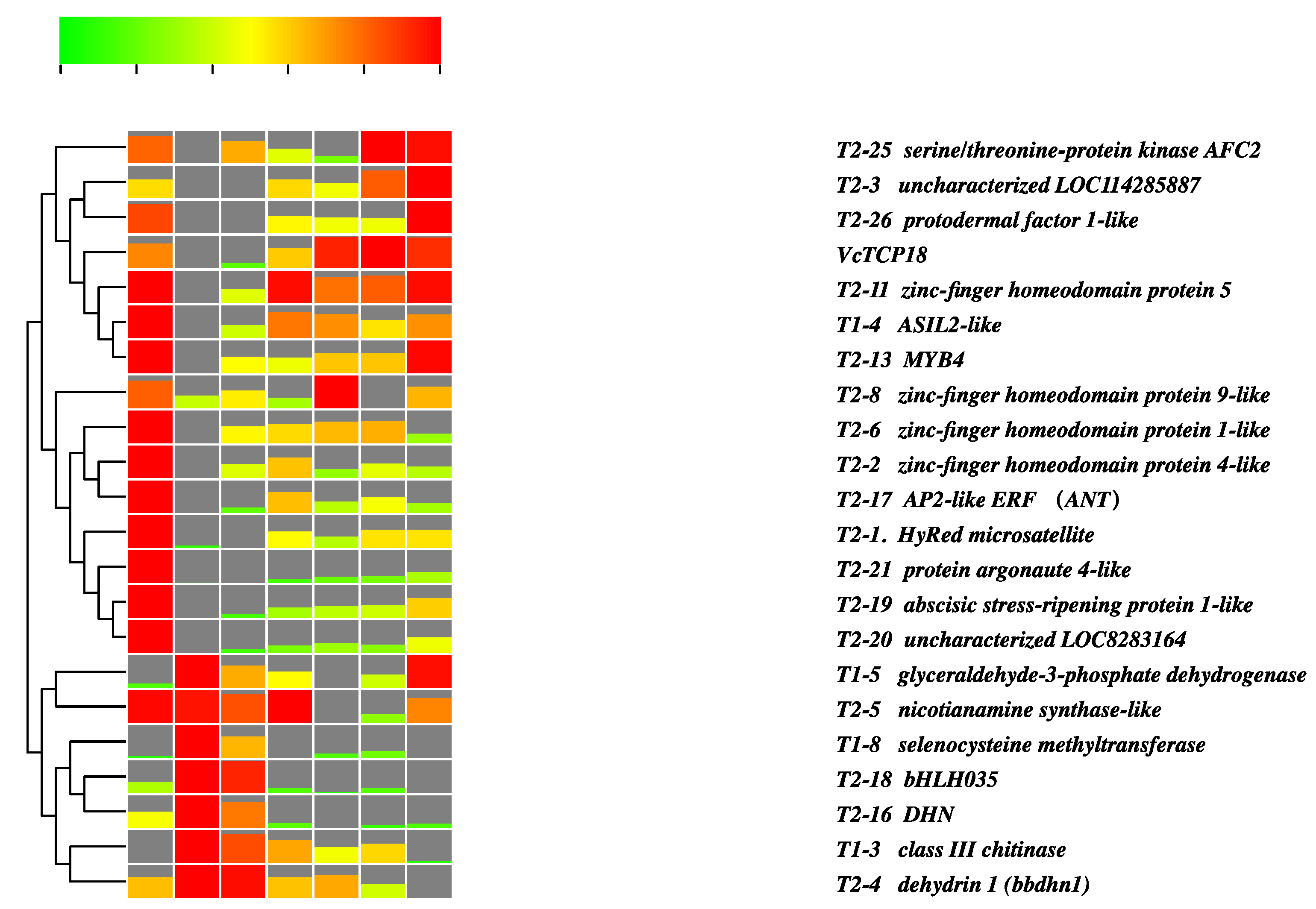
2.5. Analysis of the Expression of the Screened Genes
3. Discussion
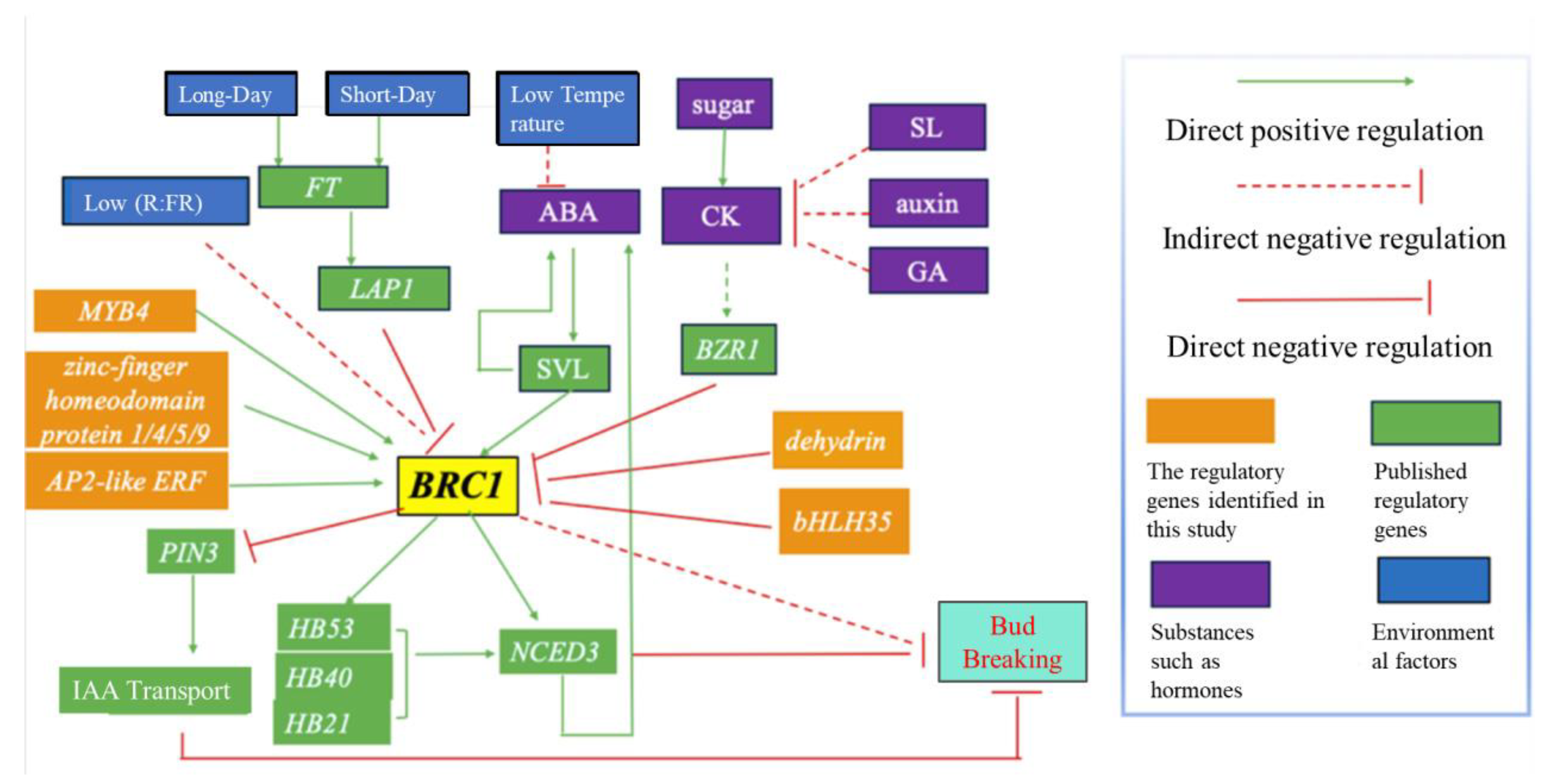
3.1. Bud Break and BRC1
3.2. BRC1 and Its Upstream Regulatory Genes
4. Materials and Methods
4.1. Plant Materials
4.2. qRT-PCR Analysis
4.3. Prediction of the Promoter’s Cis-Acting Element
4.4. Promoter Activity Analysis
4.5. Screening of the Y1H Library
4.6. Identification of Positive Clones and Extraction of the Prey Plasmids
4.7. Positive Clone Rotation Verification
4.8. Expression Pattern of the Upstream Regulator Genes in the Transcriptome
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lang, G.A.; Early, J.D.; Martin, G.C.; Darnell, R.L. Endo-, Para-, and Ecodormancy: Physiological Terminology and Classification for Dormancy Research. Hortscience 1987, 22, 271–277. [Google Scholar]
- Richards, R.A. Selectable Traits to Increase Crop Photosynthesis and Yield of Grain Crops. J. Exp. Bot. 2000, 51, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, J. Genes Controlling Plant Architecture. Curr. Opin. Biotechnol. 2006, 17, 123–129. [Google Scholar] [CrossRef]
- Jiao, Y.; Wang, Y.; Xue, D.; Wang, J.; Yan, M.; Liu, G.; Dong, G.; Zeng, D.; Lu, Z.; Zhu, X.; et al. Regulation of OsSPL14 by OsmiR156 Defines Ideal Plant Architecture in Rice. Nat. Genet. 2010, 42, 541–545. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Morinaka, Y.; Ohnishi, T.; Sunohara, H.; Fujioka, S.; Ueguchi-Tanaka, M.; Mizutani, M.; Sakata, K.; Takatsuto, S.; Yoshida, S.; et al. Erect Leaves Caused by Brassinosteroid Deficiency Increase Biomass Production and Grain Yield in Rice. Nat. Biotechnol. Sci. Bus. Biotechnol. 2006, 24, 105–109. [Google Scholar] [CrossRef]
- Beauvieux, R.; Wenden, B.; Dirlewanger, E. Bud Dormancy in Perennial Fruit Tree Species: A Pivotal Role for Oxidative Cues. Front. Plant Sci. 2018, 9, 657. [Google Scholar] [CrossRef]
- Horvath, D.P.; Anderson, J.V.; Chao, W.S.; Foley, M.E. Knowing When to Grow: Signals Regulating Bud Dormancy. Trends Plant Sci. 2003, 8, 534–540. [Google Scholar] [CrossRef]
- Lobos, G.A. Breeding Blueberries for a Changing Global Environment: A Review. Front. Plant Sci. 2015, 6, 14. [Google Scholar] [CrossRef]
- Wang, B.; Smith, S.M.; Li, J. Genetic Regulation of Shoot Architecture. In Annual Review of Plant Biology; Merchant, S.S., Ed.; Annual Reviews: San Mateo, CA, USA, 2018; Volume 69, pp. 437–468. [Google Scholar] [CrossRef]
- Doebley, J.; Stec, A.; Gustus, C. Teosinte Branched1 and the Origin of Maize: Evidence for Epistasis and the Evolution of Dominance. Genetics 1995, 141, 333–346. [Google Scholar] [CrossRef]
- Wang, M.; Le Moigne, M.-A.; Bertheloot, J.; Crespel, L.; Perez-Garcia, M.-D.; Ogé, L.; Demotes-Mainard, S.; Hamama, L.; Davière, J.-M.; Sakr, S. BRANCHED1: A Key Hub of Shoot Branching. Front. Plant Sci. 2019, 10, 76. [Google Scholar] [CrossRef]
- Aguilar-Martínez, J.A.; Poza-Carrión, C.; Cubas, P. Arabidopsis BRANCHED1 Acts as an Integrator of Branching Signals within Axillary Buds. Plant Cell 2007, 19, 458–472. [Google Scholar] [CrossRef]
- Shen, J.; Zhang, Y.; Ge, D.; Wang, Z.; Song, W.; Gu, R.; Che, G.; Cheng, Z.; Liu, R.; Zhang, X. CsBRC1 Inhibits Axillary Bud Outgrowth by Directly Repressing the Auxin Efflux Carrier CsPIN3 in Cucumber. Proc. Natl. Acad. Sci. USA 2019, 116, 17105–17114. [Google Scholar] [CrossRef]
- Gomez-Roldan, V.; Fermas, S.; Brewer, P.B.; Puech-Pages, V.; Dun, E.A.; Al, E. Strigolactone Inhibition of Shoot Branching. Nature 2008, 455, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Maurya, J.P.; Azeez, A.; Miskolczi, P.; Tylewicz, S.; Stojkovič, K.; Delhomme, N.; Busov, V.; Bhalerao, R.P. A Genetic Network Mediating the Control of Bud Break in Hybrid Aspen. Nat. Commun. 2018, 9, 4173. [Google Scholar] [CrossRef] [PubMed]
- Maurya, J.P.; Singh, R.K.; Miskolczi, P.C.; Prasad, A.N.; Jonsson, K.; Wu, F.; Bhalerao, R.P. Branching Regulator BRC1 Mediates Photoperiodic Control of Seasonal Growth in Hybrid Aspen. Curr. Biol. 2020, 30, 122–126.e2. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, M.; Torres-Pérez, R.; Wahl, V.; Cruz-Oró, E.; Rodríguez-Buey, M.L.; Zamarreño, A.M.; Martín-Jouve, B.; García-Mina, J.M.; Oliveros, J.C.; Prat, S.; et al. Spatial Control of Potato Tuberization by the TCP Transcription Factor BRANCHED1b. Nat. Plants 2022, 8, 281–294. [Google Scholar] [CrossRef]
- Nicolas, M.; Rodríguez-Buey, M.L.; Franco-Zorrilla, J.M.; Cubas, P. A Recently Evolved Alternative Splice Site in the BRANCHED1a Gene Controls Potato Plant Architecture. Curr. Biol. 2015, 25, 1799–1809. [Google Scholar] [CrossRef]
- Li, Y.; An, S.; Cheng, Q.; Zong, Y.; Chen, W.; Guo, W.; Zhang, L. Analysis of Evolution, Expression and Genetic Transformation of TCP Transcription Factors in Blueberry Reveal That VcTCP18 Negatively Regulates the Release of Flower Bud Dormancy. Front. Plant Sci. 2021, 12, 697609. [Google Scholar] [CrossRef]
- Janssen, B.J.; Drummond, R.S.; Snowden, K.C. Regulation of Axillary Shoot Development. Curr. Opin. Plant Biol. 2014, 17, 28–35. [Google Scholar] [CrossRef]
- Barbier, F.F.; Dun, E.A.; Kerr, S.C.; Chabikwa, T.G.; Beveridge, C.A. An Update on the Signals Controlling Shoot Branching. Trends Plant Sci. 2019, 24, 220–236. [Google Scholar] [CrossRef]
- Martín-Fontecha, E.S.; Tarancón, C.; Cubas, P. To Grow or Not to Grow, a Power-Saving Program Induced in Dormant Buds. Curr. Opin. Plant Biol. 2017, 41, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Clemence, H.; Amelie, R.; Maryse, L.; Eric, M.; Monique, S.; Nathalie, L.; Rémi, L.; Soulaiman, S.; Alain, V.; Sandrine, P.-T. Regulation of RhSUC2, a Sucrose Transporter, Is Correlated with the Light Control of Bud Burst in Rosa Sp. Plant Cell Environ. 2011, 34, 1776–1789. [Google Scholar]
- Catherine, R.; Jessica, B.; Nathalie, L.; Bruno, A.; Fabrice, F.; Soulaiman, S. Multiple Pathways Regulate Shoot Branching. Front. Plant Sci. 2015, 5, 741. [Google Scholar]
- Gao, Y.; Yang, Q.; Yan, X.; Wu, X.; Yang, F.; Li, J.; Wei, J.; Ni, J.; Ahmad, M.; Bai, S.; et al. High-Quality Genome Assembly of “Cuiguan” Pear (Pyrus Pyrifolia) as a Reference Genome for Identifying Regulatory Genes and Epigenetic Modifications Responsible for Bud Dormancy. Hortic. Res. 2021, 8, 197. [Google Scholar] [CrossRef] [PubMed]
- Ruttink, T.; Arend, M.; Morreel, K.; Bhalerao, R.P.; Boerjan, W.; Rohde, A. A Molecular Timetable for Apical Bud Formation and Dormancy Induction in Poplar. Plant Cell 2007, 19, 2370–2390. [Google Scholar] [CrossRef]
- Li, Y.; Ma, R.; Li, R.; Zhao, Q.; Zhang, Z.; Zong, Y.; Yao, L.; Chen, W.; Yang, L.; Liao, F.; et al. Comparative Transcriptomic Analysis Provides Insight into the Key Regulatory Pathways and Differentially Expressed Genes in Blueberry Flower Bud Endo- and Ecodormancy Release. Horticulturae 2022, 8, 176. [Google Scholar] [CrossRef]
- Feng, C.; Andreasson, E.; Maslak, A.; Mock, H.P.; Mattsson, O.; Mundy, J. Arabidopsis MYB68 in Development and Responses to Environmental Cues. Plant Sci. 2004, 167, 1099–1107. [Google Scholar] [CrossRef]
- Dai, X.; Xu, Y.; Ma, Q.; Xu, W.; Wang, T.; Xue, Y.; Chong, K. Overexpression of an R1R2R3 MYB Gene, OsMYB3R—2, Increases Tolerance to Freezing, Drought, and Salt Stress in Transgenic Arabidopsis. Plant Physiol. 2007, 143, 1739–1751. [Google Scholar] [CrossRef]
- Chen, B.-J.; Wang, Y.; Hu, Y.-L.; Wu, Q.; Lin, Z.-P. Cloning and Characterization of a Drought-Inducible MYB Gene from Boea Crassifolia. Plant Sci. 2005, 168, 493–500. [Google Scholar] [CrossRef]
- Cominelli, E.; Galbiati, M.; Vavasseur, A.; Conti, L.; Sala, T.; Vuylsteke, M.; Leonhardt, N.; Dellaporta, S.L.; Tonelli, C. A Guard-Cell-Specific MYB Transcription Factor Regulates Stomatal Movements and Plant Drought Tolerance. Curr. Biol. 2005, 15, 1196–1200. [Google Scholar] [CrossRef]
- Pasquali, G.; Biricolti, S.; Locatelli, F.; Baldoni, E.; Mattana, M. Osmyb4 Expression Improves Adaptive Responses to Drought and Cold Stress in Transgenic Apples. Plant Cell Rep. 2008, 27, 1677–1686. [Google Scholar] [CrossRef]
- Park, M.-R.; Yun, K.-Y.; Mohanty, B.; Herath, V.; Xu, F.; Wijaya, E.; Bajic, V.B.; Yun, S.-J.; De Los Reyes, B.G. Supra-Optimal Expression of the Cold-Regulated OsMyb4 Transcription Factor in Transgenic Rice Changes the Complexity of Transcriptional Network with Major Effects on Stress Tolerance and Panicle Development. Plant Cell Environ. 2010, 33, 2209–2230. [Google Scholar] [CrossRef]
- Yang, C.Y.; Chen, Y.C.; Jauh, G.Y.; Wang, C.-S. A Lily ASR Protein Involves Abscisic Acid Signaling and Confers Drought and Salt Resistance in Arabidopsis. Plant Physiol. 2005, 139, 836–846. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Yang, L.; Zhang, B.; Song, X.; Li, Y.; Wang, S. Molecular Cloning of Sugarcane ASR Gene(SoASR)and Its Expression Analysis. Biotechnol. Bull. 2013, 2, 93–99. [Google Scholar]
- Xu, H.; Yang, Y.; Xie, L.; Li, X.; Feng, C.; Chen, J.; Xu, C. Involvement of Multiple Types of Dehydrins in the Freezing Response in Loquat (Eriobotrya Japonica). PLoS ONE 2014, 9, e87575. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Li, X.; Xu, C.; Chen, J. A Study on Cold Resistance Function of Dehydrin Gene EjDHN5 from Loqua. Acta Hortic. Sin. 2017, 44, 268–278. [Google Scholar] [CrossRef]
- Hancock, J.F. Temperate Fruit Crop Breeding: Germplasm to Genomics; Springer: Berlin, Germany, 2008. [Google Scholar]
- Ehlenfeldt, M.K.; Draper, A.D.; Clark, J.R. Performance of Southern Highbush Blueberry Cultivars Released by the U.S. Department of Agriculture and Cooperating State Agricultural Experiment Stations. Horttechnology 1995, 27, 127–130. [Google Scholar] [CrossRef]
- Lyrene, P.M. “Emerald” Southern Highbush Blueberry. Hortscience A Publ. Am. Soc. Hortic. Sci. 2008, 43, 1606–1607. [Google Scholar] [CrossRef]
- Hao, X.; Yang, Y.; Yue, C.; Wang, L.; Horvath, D.P.; Wang, X. Comprehensive Transcriptome Analyses Reveal Differential Gene Expression Profiles of Camellia Sinensis Axillary Buds at Para-, Endo-, Ecodormancy, and Bud Flush Stages. Front. Plant Sci. 2017, 8, 1–19. [Google Scholar] [CrossRef]
- Yang, L.; Chen, M.; Cai, K.; Zhang, L.; Zhu, Y.; Ye, Q.; Lu, M.; Liao, F.; Chen, W.; Guo, W. VcFAS, VcSUN and VcOVATE Orchestrated the Fruit Morphogenesis in Southern Highbush Blueberry during the Pre-Anthesis and Fruit Development. Sci. Horticulturae 2018, 240, 109–115. [Google Scholar] [CrossRef]
- Die, J.V.; Rowland, L.J.; Niedz, R.P. Superior Cross-Species Reference Genes: A Blueberry Case Study. PLoS ONE 2013, 8, e73354. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, L.; Wang, Z.; Zong, Y.; Yu, L.; Li, Y.; Liao, F.; Chen, M.; Cai, K.; Guo, W. Comparative Anatomical and Transcriptomic Insights into Vaccinium Corymbosum Flower Bud and Fruit throughout Development. BMC Plant Biol. 2021, 21, 289. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a Database of Plant Cis-Acting Regulatory Elements and a Portal to Tools for in Silico Analysis of Promoter Sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Chen, K.-S.; Li, F.; Xu, C.-J.; Zhang, S.-L.; Fu, C.-X. An efficient macro-method of genomic DNA isolation from Actinidia chinensis leaves. Yi Chuan 2004, 26, 529–531. [Google Scholar] [PubMed]
- Liu, Z.N.; Yuan, L.; Sundaresan, V.; Yu, X.L. Screening and identification of CKI1 upstream transcription regulators in Arabidopsis. Yi Chuan 2019, 41, 430–438. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serial Number | Gene Name | NCBI Login Number | Identity | E-Value |
|---|---|---|---|---|
| 3 | Class III chitinase | GU944517.1 | 84.00% | 0.0 |
| 4 | Transcription factor ASIL2-like (LOC109148174) | XM_019296010.1 | 80.89% | 1 × 10−58 |
| 5 | Glyceraldehyde-3-phosphate dehydrogenase GAPCP1 | XM_028206951.1 | 81.20% | 0.0 |
| 8 | Selenocysteine methyltransferase | DQ480337.1 | 88.27% | 0.0 |
| Serial Number | Gene Name | NCBI Login Number | Identity | E-Value |
|---|---|---|---|---|
| 1 | HyRed microsatellite | KP279095.1 | 84.83% | 2 × 10−25 |
| 2 | Zinc-finger homeodomain protein 4-like | XM_016035315.2 | 76.37% | 2 × 10−88 |
| 3 | Uncharacterized LOC114285887 | XM_028228973.1 | 82.82% | 2 × 10−88 |
| 4 | Dehydrin 1 (bbdhn1) | AF030180.1 | 96.09% | 0.0 |
| 5 | Nicotianamine synthase-like | XM_028241468.1 | 83.10% | 1 × 10−141 |
| 6 | Zinc-finger homeodomain protein 1-like | XM_018974724.1 | 85.57% | 9 × 10−48 |
| 8 | Zinc-finger homeodomain protein 9-like | XM_030657350.1 | 77.68% | 6 × 10−46 |
| 11 | Zinc-finger homeodomain protein 5 | XM_010062653.2 | 87.36% | 2 × 10−46 |
| 13 | MYB4 | KT225482.1 | 99.24% | 0.0 |
| 16 | DHN | KF192819.1 | 99.60% | 0.0 |
| 17 | AP2-like ethylene-responsive transcription factor (ANT) | XM_028207531.1 | 86.88% | 0.0 |
| 18 | bHLH035 | KU933649.1 | 95.48% | 0.0 |
| 19 | Abscisic stress-ripening protein 1-like | XM_035075404.1 | 82.20% | 2 × 10−54 |
| 20 | Uncharacterized LOC8283164 | XM_002524251.3 | 80.15% | 7 × 10−45 |
| 21 | Protein argonaute 4-like | XM_028251957.1 | 86.89% | 0.0 |
| 25 | Serine/threonine-protein kinase AFC2 | XM_027915241.1 | 79.02% | 5 × 10−74 |
| 26 | protodermal factor 1-like | XM_028230087.1 | 77.40 | 1 × 10−47 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, R.; Ma, R.; Zheng, Y.; Zhao, Q.; Zong, Y.; Zhu, Y.; Chen, W.; Li, Y.; Guo, W. A Study of the Molecular Regulatory Network of VcTCP18 during Blueberry Bud Dormancy. Plants 2023, 12, 2595. https://doi.org/10.3390/plants12142595
Li R, Ma R, Zheng Y, Zhao Q, Zong Y, Zhu Y, Chen W, Li Y, Guo W. A Study of the Molecular Regulatory Network of VcTCP18 during Blueberry Bud Dormancy. Plants. 2023; 12(14):2595. https://doi.org/10.3390/plants12142595
Chicago/Turabian StyleLi, Ruixue, Rui Ma, Yuling Zheng, Qi Zhao, Yu Zong, Youyin Zhu, Wenrong Chen, Yongqiang Li, and Weidong Guo. 2023. "A Study of the Molecular Regulatory Network of VcTCP18 during Blueberry Bud Dormancy" Plants 12, no. 14: 2595. https://doi.org/10.3390/plants12142595
APA StyleLi, R., Ma, R., Zheng, Y., Zhao, Q., Zong, Y., Zhu, Y., Chen, W., Li, Y., & Guo, W. (2023). A Study of the Molecular Regulatory Network of VcTCP18 during Blueberry Bud Dormancy. Plants, 12(14), 2595. https://doi.org/10.3390/plants12142595