
Influence of Variable Chloride/Sulfur Doses as Part of Potassium Fertilization on Nitrogen Use Efficiency by Coffee

 ,
,
Abstract
1. Introduction
2. Results
2.1. Influence of Cl and S Rates on Biomass Accumulation, Nutrient Uptake, and NUE under Greenhouse Conditions
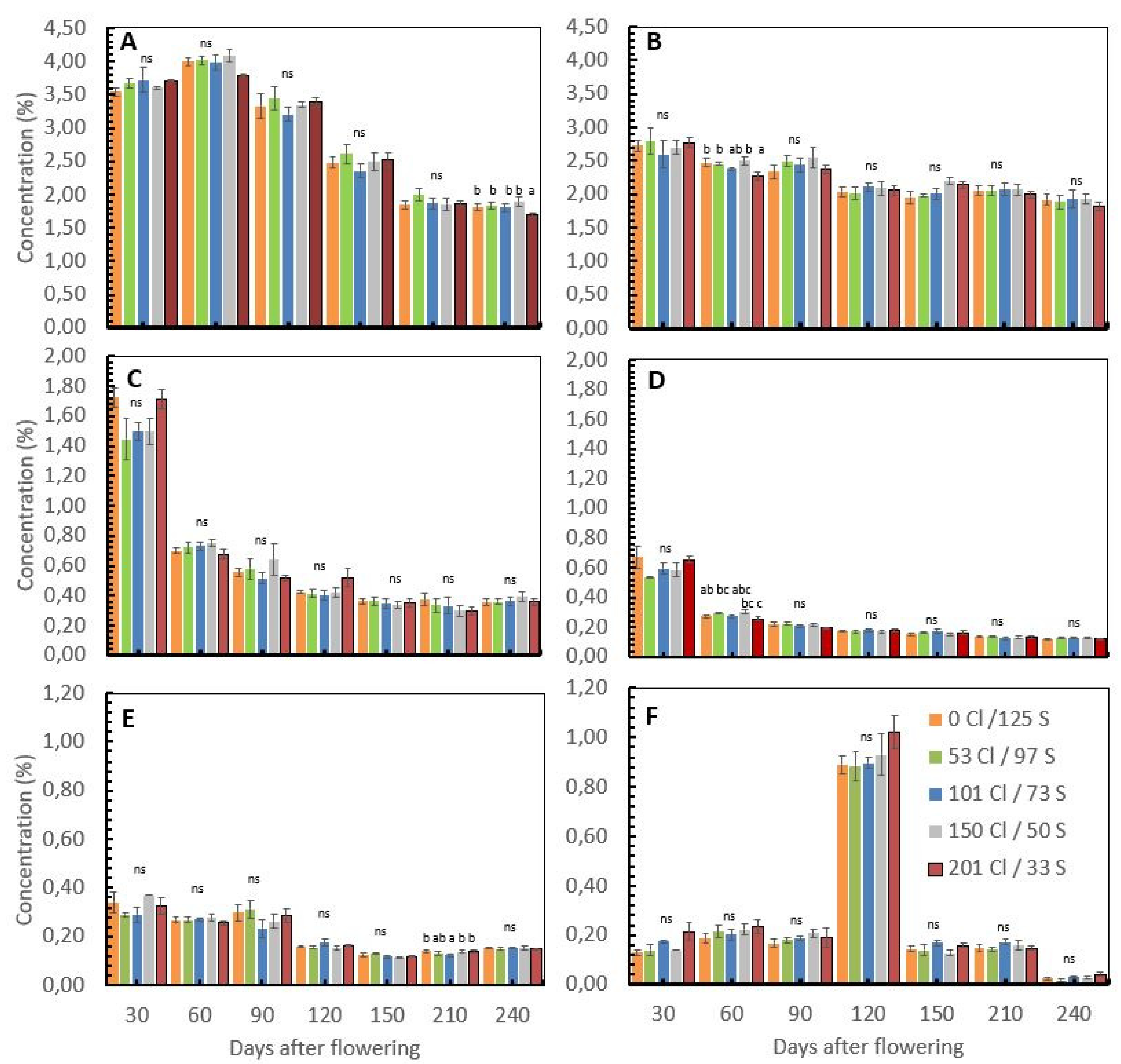
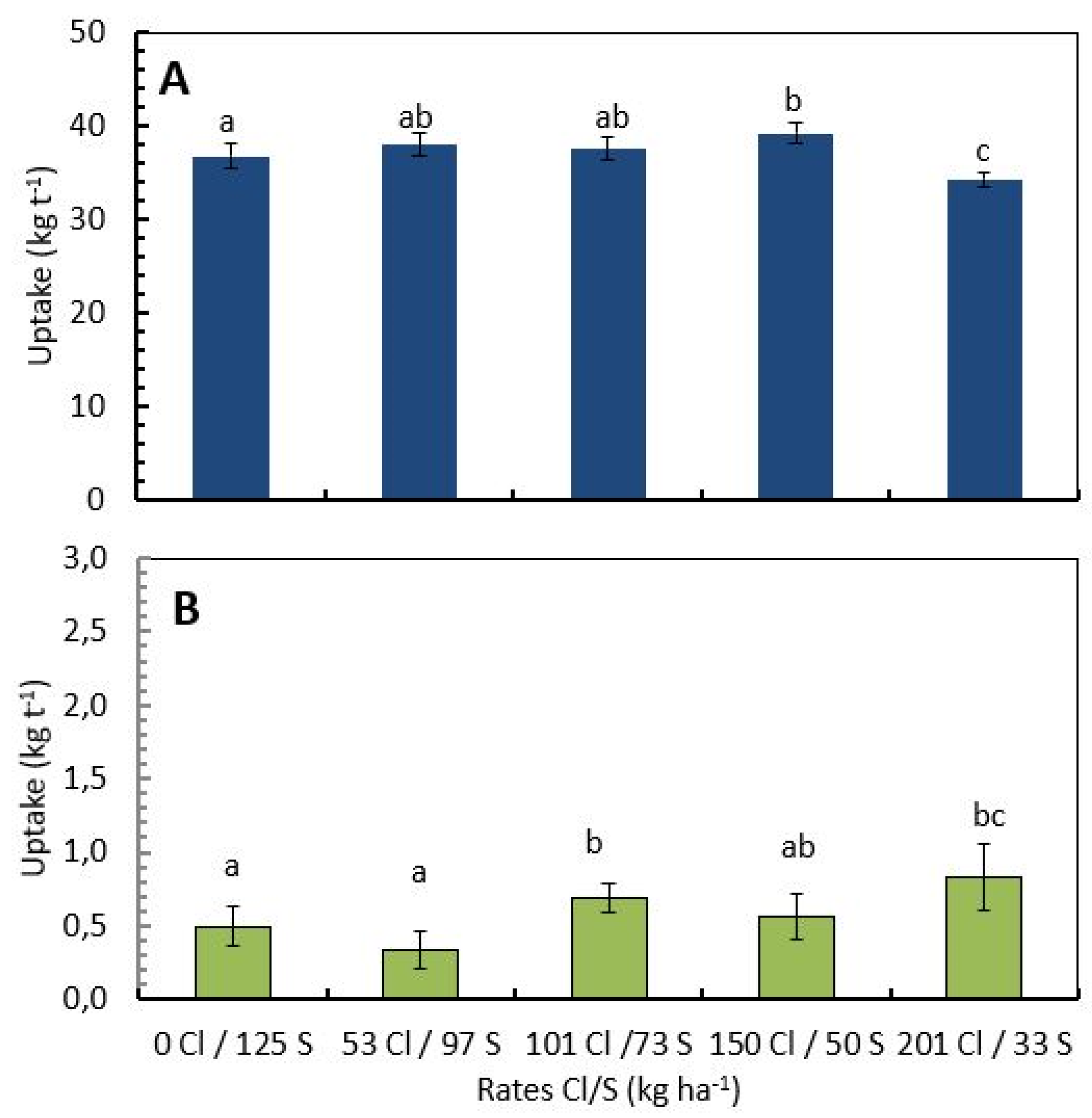
2.2. Cl/S Ratios Influence Nutrient Concentration and Uptake by Coffee Beans under Field Conditions
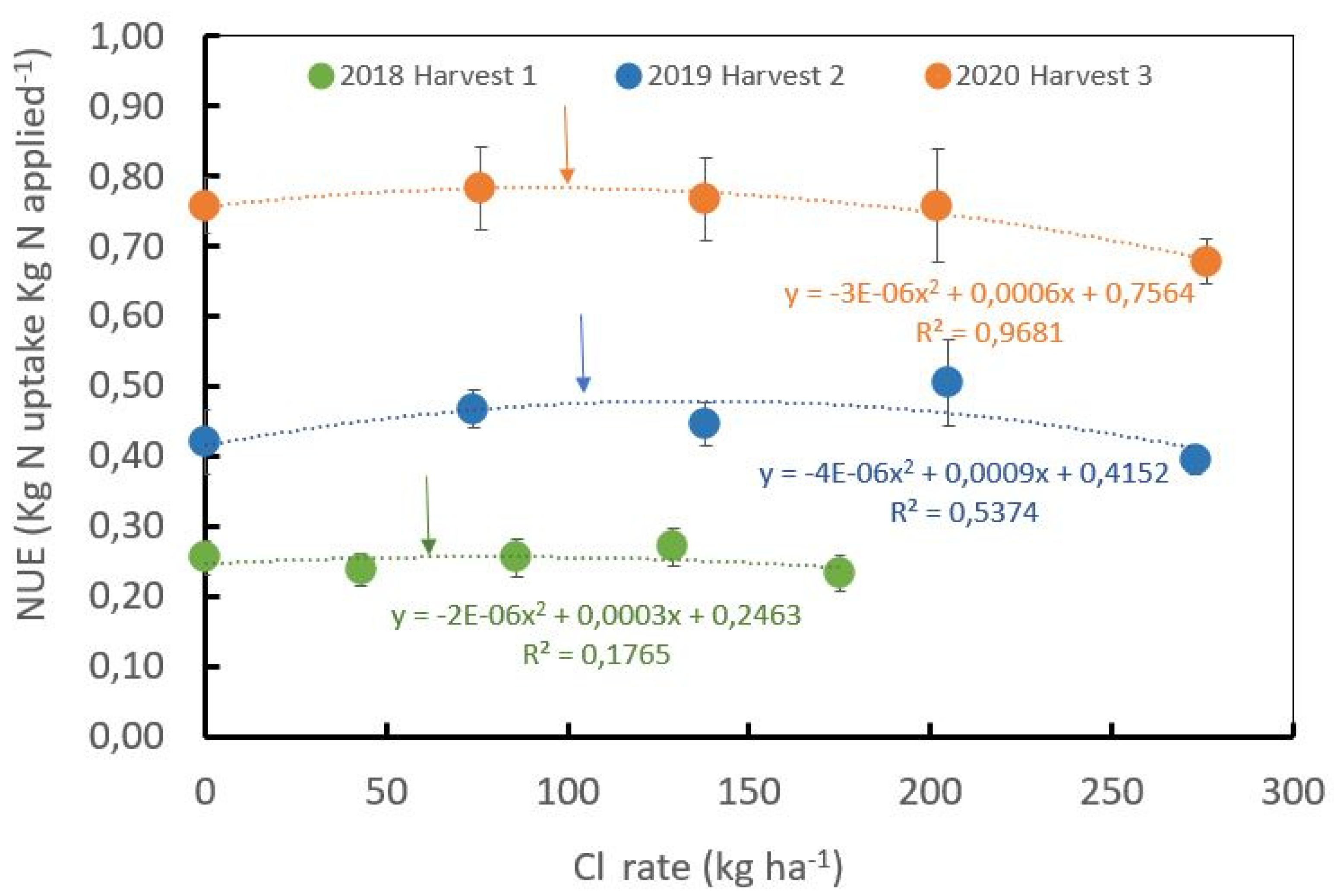
2.3. Cl/S Ratios Influence Nitrogen Use Efficiency for Coffee Beans at the Field Level
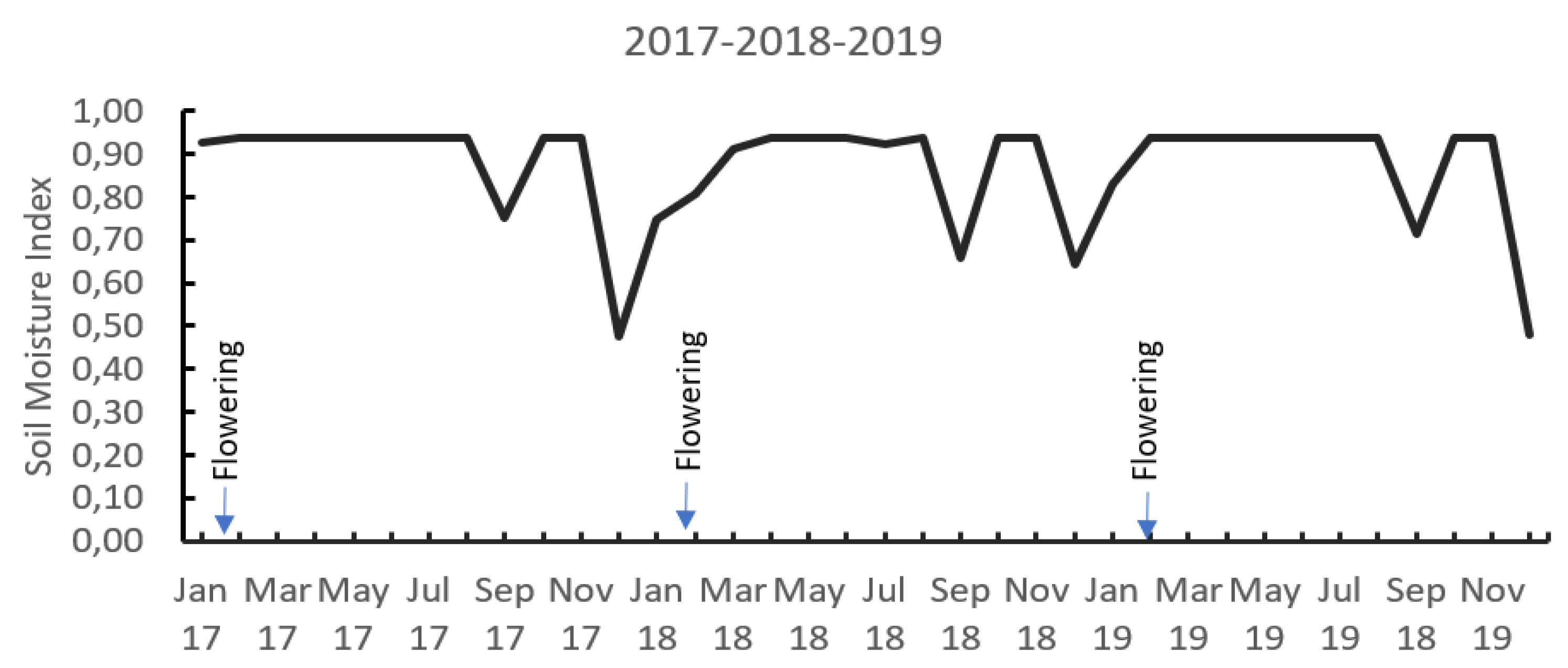
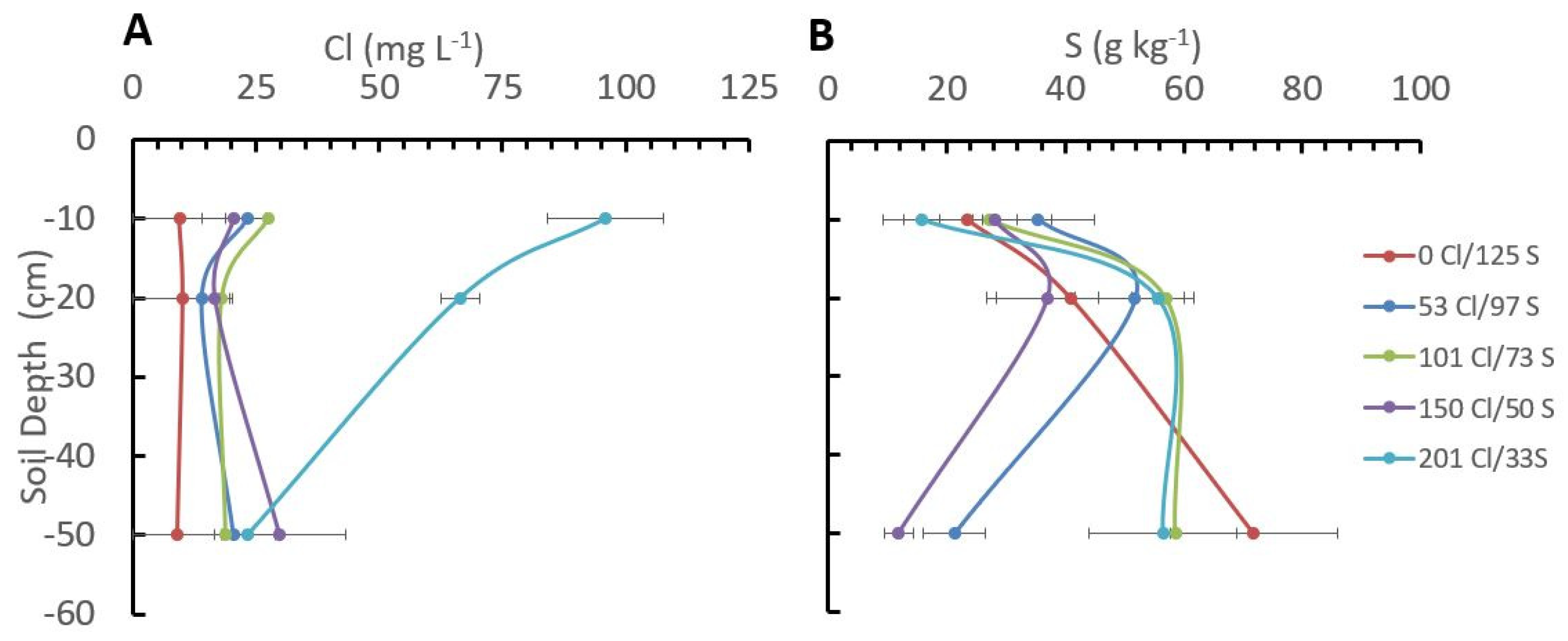
2.4. Cl/S Ratios Influence Cl and S Distribution in the Soil Profile
3. Discussion
3.1. Cl− Concentration on Tissues and Influence on Growth and Productivity
3.2. Cl− Influence on Nutrients Uptake and NUE
3.3. Cl− on the Soil
4. Materials and Methods
4.1. Greenhouse Trial
4.2. Field Trial
5. Conclusions
- -
- A balance between Cl− and S is necessary for coffee with the aim to improve N uptake and NUE. For greenhouse conditions, there should be between 60 and 180 mg·L−1 of Cl− and between 160 and 80 mg·L−1 of S. Plants without Cl− or without S significantly reduce the NUE. At the field level, the balances should be between 100 and 150 kg Cl− ha−1·year−1 and between 73 and 50 kg S·ha−1·year−1.
- -
- Cl− is applied to coffee at rates as a “macronutrient”, and the coffee plants can take higher amounts of Cl−, reaching concentrations on the leaves of higher than 0.69% without toxicity symptoms, but with a significant reduction in dry biomass accumulation. With the aim to keep a balance between biomass accumulation and NUE, the Cl− content in the leaves should be lower than 0.33%, placing the coffee into the group of glycophyte Cl−-sensitive plants.
- -
- Cl− rates at the field level that are higher than 150 kg Cl− ha−1·year−1 significantly increase the Cl− content in the soil and Cl− uptake by the coffee cherries and reduces N uptake by the coffee cherries, reducing the NUE on average by 9% to 14%.
- -
- In terms of the NUE, the Cl− content in the soil in coffee in the first 20 cm depth should not exceed the concentration of 30 mg·L−1.
- -
- The NUE in coffee could be improved by using agronomical management practices such as the selection of mineral potassium fertilizer sources, with the main aim to reduce the Cl− application rates and to limit them to no more than 100 to 150 kg Cl− ha−1 with a fine balance with S rates, as mentioned before.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- White, J.P.; Broadley, R.M. Chloride in Soils and its Uptake and Movement within the Plant. A Review. Ann. Bot. 2001, 88, 967–988. [Google Scholar] [CrossRef]
- Chen, E.; He, L.Z.; Yang, E.X.; Mishra, S.; Stoffella, J.P. Chlorine nutrition of higher plants: Progress and perspectives. J. Plant Nutr. 2010, 33, 943–952. [Google Scholar] [CrossRef]
- Colmenero-Flores, J.M.; Franco-Navarro, J.D.; Cubero-Front, P.; Peinado-Torrubia, P.; Rosales, M.A. Chloride as a beneficial Macronutrient in Higher Plants: New Roles and Regulation. Int. J. Mol. Sci. 2019, 20, 4686. [Google Scholar] [CrossRef]
- Broyer, T.C.; Carlton, A.B.; Johnson, C.M.; Stout, P.R. Chlorine—A micronutrient element for higher plants. Plant Physiol. 1954, 29, 526–532. [Google Scholar] [CrossRef]
- Marschner, P. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Elsevier: Amsterdam, The Netherlands; Academic Press: San Diego, CA, USA, 2012; 668p. [Google Scholar]
- Franco-Navarro, J.D.; Brúmos, J.; Rosales, M.A.; Cubero-Font, P.; Talón, M.; Colmero-Flores, J.M. Chloride regulates leaf cell size and water relations in tobacco plants. J. Exp. Bot. 2015, 67, 873–891. [Google Scholar] [CrossRef] [PubMed]
- Terry, N. Photosynthesis, Growth, and the Role of Chloride. Plant Physiol. 1977, 60, 69–75. [Google Scholar] [CrossRef]
- Rosales, M.A.; Franco-Navarro, J.D.; Peinado-Torrubia, P.; Diaz-Rueda, P.; Alvarez, R.; Colmero-Flores, J.M. Chloride Improves Nitrates Utilization and NUE in Plants. Front. Plant Sci. 2020, 11, 442. [Google Scholar] [CrossRef]
- Sollins, P.; Robertson, P.G.; Uehara, G. Nutrient mobility in variable- and permanent-charge soils. Biogeochemistry 1988, 6, 181–199. [Google Scholar] [CrossRef]
- Neto, A.P.; Favarin, J.L.; de Almeida, R.E.M.; dos Santos Dias, C.T.; Tezotto, T.; Ferreira, M.T. Changes of Nutritional Status during a Phenological Cycle of Coffee under High Nitrogen Supply by Fertigation. Commun. Soil Sci. Plant Anal. 2011, 42, 2414–2425. [Google Scholar] [CrossRef]
- Neto, A.P.; Favarin, J.L.; do Reis, A.R.; Tezotto, T.; de Almeida, R.E.M.; Levres, J.; Gallo, L.A. Nitrogen metabolism in coffee plants in response to nitrogen supply by fertigation. Theor. Exp. Plant Physiol. 2015, 27, 41–50. [Google Scholar] [CrossRef]
- Bruno, P.I.; Unkovich, J.M.; Bortolotto, P.R.; Bacchi, O.S.; Dourado, N.D.; Reichardt, K. Fertilizer nitrogen in fertigated coffee crop: Absorption changes in plant compartments over time. Field Crops Res. 2011, 124, 369–377. [Google Scholar] [CrossRef]
- Sadeghian, K.S.; Duque, O.H. Optimal doses of nutrients for coffee plantations in production: Economical considerations. Adv. Técnicos Cenicafé 2021, 533, 1–8. (In Spanish) [Google Scholar] [CrossRef]
- Favarin, J.L.; de Souza, L.T.; Mazzafera, P.; Dimenstein, L. Soil correction and fertilization of irrigated coffee plants in production. In Cafeicultura do Cerrado/Editores Técnicos Gladyston Rodrigues/Carvalho et al-Belo Horizonte; EPAMIG: Minas Gerais, Brazil, 2021; 564p. (In Portuguese) [Google Scholar]
- Cannavo, P.; Harmand, J.M.; Zeller, B.; Vaast, F.; Ramirez, J.E.; Dambrine, E. Low nitrogen use efficiency and high nitrate leaching in a highly fertilized Coffea arabica–Inga densiflora agroforestry system: A 15N labeled fertilizer study. Nutr. Cycl. Agroecosyst. 2013, 92, 377–394. [Google Scholar] [CrossRef]
- Salamanca, J.A.; Doane, T.; Horwath, R.W. Nitrogen Use Efficiency of Coffee at the Vegetative Stage as Influenced by Fertilizer Application Method. Front. Plant Sci. 2017, 8, 223. [Google Scholar] [CrossRef] [PubMed]
- Silva, E.B.; Nogueira, F.D.; Guimaräes, P.T.G.; Furtini Neto, A.E. Responds of the coffee to the potassium fertilization. Pesq. Agropec. Bras. 2001, 36, 133–1337. (In Portuguese) [Google Scholar] [CrossRef]
- Sadeghian, K.S. Soil fertility and coffee nutrition in Colombia. Boletín Técnico Cenicafé. 2008, 43. (In Spanish) [Google Scholar]
- Cuzato-Mancuso, M.A.; Peres, S.R.; Costa, C.C.A.; Amaral, C.G.S. Effect of the potassium sources and rates on arabica coffee yield, nutrition, and macronutrient export. Rev. Bras. Cienc. Solo. 2014, 38, 1448–1456. [Google Scholar] [CrossRef]
- Mengel, K.; Kirkby, E.A. Principles of Plant Nutrition; CH-3048; International Potash Institute: Worblaufen-Bern, Switzerland, 1978; 593p. [Google Scholar]
- Santos, C.; Malta, M.R.; Gonzalves, M.G.M.; Borém, F.M.; Pozza, A.A.A.; Martinez, H.E.P.; de Souza, T.I.; Chagas, W.F.T.; de Melo, M.E.A.; Oliveira, D.P.; et al. Chloride Applied via Fertilizer Affects Plant Nutrition and Coffee Quality. Plants 2023, 12, 885. [Google Scholar] [CrossRef]
- Ramirez, B.V.H.; Küsters, J. Calcium and Potassium nutrition increase the water use efficiency in coffee: A promising strategy to adapt to climate change. Hydrology 2021, 8, 75. [Google Scholar] [CrossRef]
- Xu, G.; Magen, H.; Tarchitzky, J.; Kafkafi, U. Advances in chloride nutrition of plants. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: San Diego, CA, USA, 2000; Volume 68, pp. 97–150. [Google Scholar]
- Silva, E.B.; Nogueira, F.D.; Guimaräes, P.T.G.; Chagas, S.J.R.; Costa, L. Sources and rates of potassium influence on productivity and quality of the coffee. Pesq. Agropec. Bras. 1999, 34, 133–1337. (In Portuguese) [Google Scholar]
- Diaz-Zorita, M.; Duarte, A.G.; Barraco, M. Effects of Chloride Fertilization on Wheat (Triticum aestivum L.) Productivity in the Sandy Pampas Region, Argentina. Agron. J. 2004, 96, 839–844. [Google Scholar] [CrossRef]
- Randle, M.W. Chloride Requirements in Onion: Clarifying a Widespread Misunderstanding. Better Crops 2004, 88, 101. [Google Scholar]
- Schwenke, G.D.; Simpfendorfer, S.R.; Collard, B.C.Y. Confirmation of chloride deficiency as the cause of leaf spotting in durum wheat grown in the Australian northern grains regions. Crop Pasture Sci. 2015, 66, 122–134. [Google Scholar] [CrossRef]
- Smith, G.S.; Clark, C.J.; Holland, P.T. Chlorine requirements of kiwifruit (Actinidia deliciosa). New Phytol. 1987, 106, 71–80. [Google Scholar] [CrossRef]
- Braconnier, S.; Dauzac, J. Chloride and stomatal conductance in coconut. Plant Physiol. Biochem. 1990, 28, 105–111. [Google Scholar]
- Brown, H.P.; Zhao, J.F.; Dobermann, A. What is a plant nutrient? Changing definitions to advance science and innovation in plant nutrition. Plant Soil 2022, 476, 11–23. [Google Scholar] [CrossRef]
- Broadley, M.; Brown, H.P.; Cakmak, I.; Ma, J.F.; Rengel, Z.; Zhao, J.F. Beneficial Elements. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marchner, P., Ed.; Academic Press: San Diego, CA, USA, 2012; pp. 249–269. [Google Scholar]
- De Wit, C.T.; Dijkshoorn, W.; Noggle, J.C. Ionic Balance and Growth of Plants; Verlagen van Landbouwkundige Onderzoekingen: Wageningen, The Netherlands, 1963; 68p. [Google Scholar]
- Diatlofft, E.; Roberts, M.; Sanders, D.; Roberts, K.S. Characterization of Anion Channels in the Plasma Membrane of Arabidopsis Epidermal Roots Cells and the Identifications of a Citrate-Permeable Channel Induced by Phosphate Starvation. Plant. Phys. 2004, 136, 4136–4149. [Google Scholar] [CrossRef]
- Carillo, P.; Rouphael, Y. Nitrate Uptake and Use Efficiency: Pros and Cons of Chloride interference in the Vegetable Crops. Front. Plant. Sci. 2022, 13, 899552. [Google Scholar] [CrossRef]
- Kafkafi, U.; Valoras, N.; Letey, J. Chloride interaction with nitrate and phosphate nutrition in tomato (Lycopersicon esculentum L.). J. Plant. Nutr. 2008, 5, 1369–1385. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Hsu, P.K.; Tsay, Y.F. Uptake, allocation and signaling of nitrate. Trends Plant Sci. 2012, 17, 458–467. [Google Scholar] [CrossRef]
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Moller, I.S.; White, P. Functions of macronutrients. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marchner, P., Ed.; Academic Press: San Diego, CA, USA, 2012; pp. 135–189. [Google Scholar]
- Wege, S.; Gilliham, M.; Henderson, S.W. Chloride: Not simply a “Cheap osmoticum” but a beneficial plant macronutrient. J. Exp. Bot. 2017, 68, 3057–3069. [Google Scholar] [CrossRef]
- Rognes, S.E. Anion regulation of lupin asparagine synthetase-Chloride activation of the glutamine-utilizing reactions. Phytochemistry 1980, 19, 2287–2293. [Google Scholar] [CrossRef]
- Imas, P.; Bansal, S.K. Use of MOP as fertilizer—Some Myths and Reality. Fertil. News 2004, 49, 51–55. [Google Scholar]
- Fixen, P.E.; Buchenau, W.G.; Gelderman, H.R.; Schumacher, E.T.; Gerwing, E.T.; Cholick, A.F.; Farber, G.B. Influences of Soil and Applied Chloride on Several Wheat Parameters. Agron. J. 1989, 78, 736–740. [Google Scholar] [CrossRef]
- Arcila, P.J.; Buhr, L.; Bleiholder, H.; Hack, H.; Meier, U.; Wicke, H. Application of the extended BBCH scale for the description of the growing stages of coffee (Coffea spp.). Ann. Appl. Biol. 2002, 141, 19–27. [Google Scholar] [CrossRef]
- Alvarado, G.; Posada, H.E.; Cortina, H.A. CASTILLO a New Coffee Variety with Rust Resistance. Adv. Técnicos Cenicafé 2005, 337, 1–8. (In Spanish) [Google Scholar]
- Riaño, H.N.M.; Arcila, P.J.; Jaramillo, R.A.; Chaves, C.B. Dry matter accumulation and nutrient extraction by Coffea arabica L cv. Colombia in three localities of the central coffee zone. Cenicafé 2004, 55, 265–276. (In Spanish) [Google Scholar]
- Sadeghian, K.S.; Mejia, M.B.; Arcila, P.J. Elemental composition of coffee fruits and extraction of nutrients by the harvest in the coffee zone of Colombia. Cenicafé 2006, 57, 251–261. (In Spanish) [Google Scholar]
- Sadeghian, K.S.; Mejia, M.B.; González, O.H. Accumulation of nitrogen, phosphorus, and potassium in coffee fruits. Adv. Técnicos Cenicafé 2013, 429, 1–8. (In Spanish) [Google Scholar]
- Congreves, K.A.; Otchere, O.; Ferland, D.; Farzadfar, S.; Williams, S.; Arcand, M.M. Nitrogen Use Efficiency Definitions of Today and Tomorrow. Front. Plant Sci. 2021, 12, 637108. [Google Scholar] [CrossRef]
- Dobermann, A. Nutrient Uses Efficient Measurement and Management; Agronomy & Horticulture; University of Nebraska: Lincoln, NE, USA, 2007; p. 1442. Available online: https://digitalcommons.unl.edu/agronomyfacpub/1442 (accessed on 10 January 2023).
- Rendón, S.J.R.; Giraldo, H. Distribution of roots in coffee variety Castillo® under two spatial arrangements. Cenicafé 2019, 70, 7–17. (In Spanish) [Google Scholar] [CrossRef]
- Ramirez, B.V.H.; Jaramillo, R.A.; Arcila, P.J. Indices to evaluate water status in coffee plantations. Cenicafé 2010, 61, 55–66. (In Spanish) [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cl/S Rates | Total DW | Root DW | N | Cl | N Uptake | S Uptake | Total Shoot Cl Uptake | NUTE ¶ | NUE ¥ |
|---|---|---|---|---|---|---|---|---|---|
| mg L−1 | g Plant−1 | Content in Leaves % | mg Plant−1 | g2 DW· mg−1 N | g DW·g−1 of N | ||||
| 0/200 | 82.1 d | 27.0 c | 2.58 a | 0.03 a | 1234.7 b | 174.2 c | 19.3 a | 0.047 c | 0.83 c |
| 60/160 | 74.1 bc | 20.3 bc | 2.68 a | 0.16 b | 1230.0 b | 137.2 c | 89.1 b | 0.044 c | 0.86 cb |
| 120/120 | 79.4 cd | 23.3 bc | 2.59 a | 0.33 c | 1236.5 b | 106.0 c | 173.4 c | 0.046 c | 0.86 cb |
| 180/80 | 74.8 bc | 18.5 b | 2.69 a | 0.55 d | 1281.1 c | 90.4 b | 276.3 d | 0.038 b | 0.89 b |
| 240/40 | 72.1 b | 16.2 ab | 2.67 a | 0.69 e | 1306.3 c | 84.9 b | 361.7 e | 0.043 c | 0.89 b |
| 300/0 | 37.9 a | 10.2 a | 4.43 b | 2.98 f | 1095.8 a | 19.4 a | 587.2 f | 0.029 a | 0.73 a |
| p value | *** | ** | *** | *** | *** | *** | *** | *** | *** |
| Nutrient Concentration (%) | Nutrient Uptake at Harvest Time (t·ha−1) | |||||||
|---|---|---|---|---|---|---|---|---|
| Nutrient | Days After Flowering | |||||||
| 30 | 60 | 90 | 120 | 150 | 210 | 240 | 240 | |
| p Value | ||||||||
| N | 0.67 | 0.13 | 0.48 | 0.37 | 0.61 | - | 0.099 | 0.028 ** |
| K | 0.90 | 0.02 ** | 0.64 | 0.93 | 0.03 ** | 0.94 | 0.83 | 0.239 |
| Ca | 0.13 | 0.50 | 0.46 | 0.25 | 0.91 | 0.42 | 0.82 | 0.220 |
| Mg | 0.38 | 0.04 ** | 0.52 | 0.85 | 0.49 | 0.83 | 0.19 | 0.398 |
| S | 0.38 | 0.68 | 0.18 | 0.36 | 0.60 | 0.081 * | 0.54 | 0.459 |
| Cl | 0.14 | 0.69 | 0.78 | 0.53 | 0.11 | 0.21 | 0.27 | 0.006 ** |
| Cl/S Rate ± kg·ha−1 | Yield + | N-Uptake per Ton | Total N− Uptake per ha | Mean N. Applied per ha ++ | NUE +++ |
|---|---|---|---|---|---|
| t·ha−1 | kg·t−1 | kg·ha−1 year−1 | kg·kg−1 | ||
| 0/125 | 4.02 ±2.05) | 36.7 (±2.04) | 147.4 (±74.5) | 308 (±20.75) | 0.48 (±0.23) |
| 53/97 | 4.01 (±1.85) | 39.1 (±2.43) | 157.3 (±75.3) | 308 (±20.75) | 0.51 (±0.24) |
| 101/73 | 4.03 (±1.96) | 37.5 (±2.32) | 151.4 (±75.9) | 308 (±20.75) | 0.49 (±0.23) |
| 150/50 | 4.03 (±2.11) | 37.9 (±2.08) | 152.8 (±78.5) | 308 (±20.75) | 0.50 (±0.24) |
| 201/33 | 3.92 (±1.87) | 34.3 (±1.31) | 134.3 (±64.21) | 308 (±20.75) | 0.44 (±0.20) |
| p value | ns | * | * | ns | |
| Year | T·min (°C) | T·max (°C) | T. med (°C) | R.H (%) | Rainfall (mm) | Sunshine (h) | Rainfall in the Trial Area (mm) |
|---|---|---|---|---|---|---|---|
| 2017 | 16.9 | 24.6 | 20.2 | 77.1 | 1.575 | 1.1467 | 2.483 |
| 2018 | 16.6 ± | 25.1 ± | 20.3 ± | 72.6 ± | 1.234 | - | 2.053 |
| 2019 | 16.1 | 23.8 | 19.4 | - | 1.426 | 1.314,0 | 2.319 |
| 2020 | - | - | - | - | 1.206 | - | 2.100 |
| Mean | 16.5 | 24.5 | 20.0 | 74.8 | 1.360 | 1.2303 | 2.238 |
| 1955–2010 | 16.2 | 24.5 | 19.7 | - | 1.330 | 1.2500 | - |
| Treatment | 2017 | 2018 | 2019 | 2020 | Average |
|---|---|---|---|---|---|
| Cl/S Rates (kg·ha−1) | |||||
| 1 | 0/41 | 0/96 | 0/174 | 0/188 | 0/125 |
| 2 | 21/30 | 43/62 | 74/140 | 74/156 | 53/97 |
| 3 | 40/20 | 86/41 | 138/109 | 138/123 | 101/73 |
| 4 | 61/10 | 129/22 | 205/78 | 202/91 | 150/50 |
| 5 | 80/0 | 175/30 | 276/44 | 276/59 | 201/33 |
| 1 to 5 | K2O Rates (kg·ha−1) | ||||
| 110 | 230 | 360 | 360 | 265 | |
| N Rates (kg·ha−1) | |||||
| 160 | 163 | 280 | 320 | 231 | |
| P2O5 Rates (kg·ha−1) | |||||
| 90 | 46 | 92 | 92 | 80 | |
| CaO Rates (kg·ha−1) | |||||
| 130 | 86 | 104 | 121 | 110 | |
| MgO Rates (kg·ha−1) | |||||
| 29 | 50 | 54 | 72 | 51 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramirez-Builes, V.H.; Küsters, J.; Thiele, E.; Leal-Varon, L.A.; Arteta-Vizcaino, J. Influence of Variable Chloride/Sulfur Doses as Part of Potassium Fertilization on Nitrogen Use Efficiency by Coffee. Plants 2023, 12, 2033. https://doi.org/10.3390/plants12102033
Ramirez-Builes VH, Küsters J, Thiele E, Leal-Varon LA, Arteta-Vizcaino J. Influence of Variable Chloride/Sulfur Doses as Part of Potassium Fertilization on Nitrogen Use Efficiency by Coffee. Plants. 2023; 12(10):2033. https://doi.org/10.3390/plants12102033
Chicago/Turabian StyleRamirez-Builes, Victor Hugo, Jürgen Küsters, Ellen Thiele, Luis Alfredo Leal-Varon, and Jorge Arteta-Vizcaino. 2023. "Influence of Variable Chloride/Sulfur Doses as Part of Potassium Fertilization on Nitrogen Use Efficiency by Coffee" Plants 12, no. 10: 2033. https://doi.org/10.3390/plants12102033
APA StyleRamirez-Builes, V. H., Küsters, J., Thiele, E., Leal-Varon, L. A., & Arteta-Vizcaino, J. (2023). Influence of Variable Chloride/Sulfur Doses as Part of Potassium Fertilization on Nitrogen Use Efficiency by Coffee. Plants, 12(10), 2033. https://doi.org/10.3390/plants12102033