
Evaluation of Fungal Volatile Organic Compounds for Control the Plant Parasitic Nematode Meloidogyne incognita

 , , ,
, , ,
Abstract
1. Introduction
2. Results
2.1. Nematicidal Activity on M. incognita Juveniles
2.2. Nematicidal Activity of VOCs on M. incognita Egg Masses
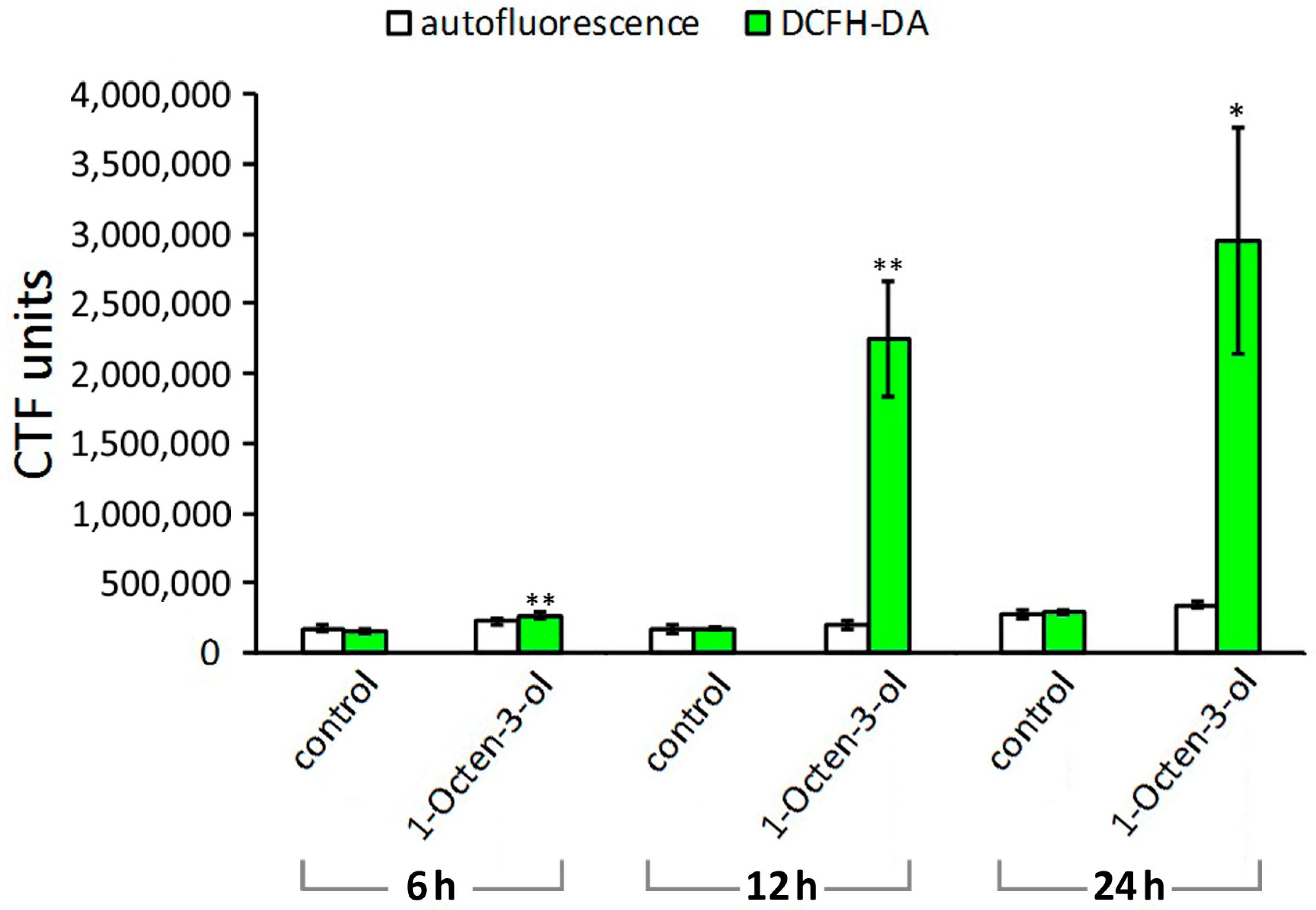
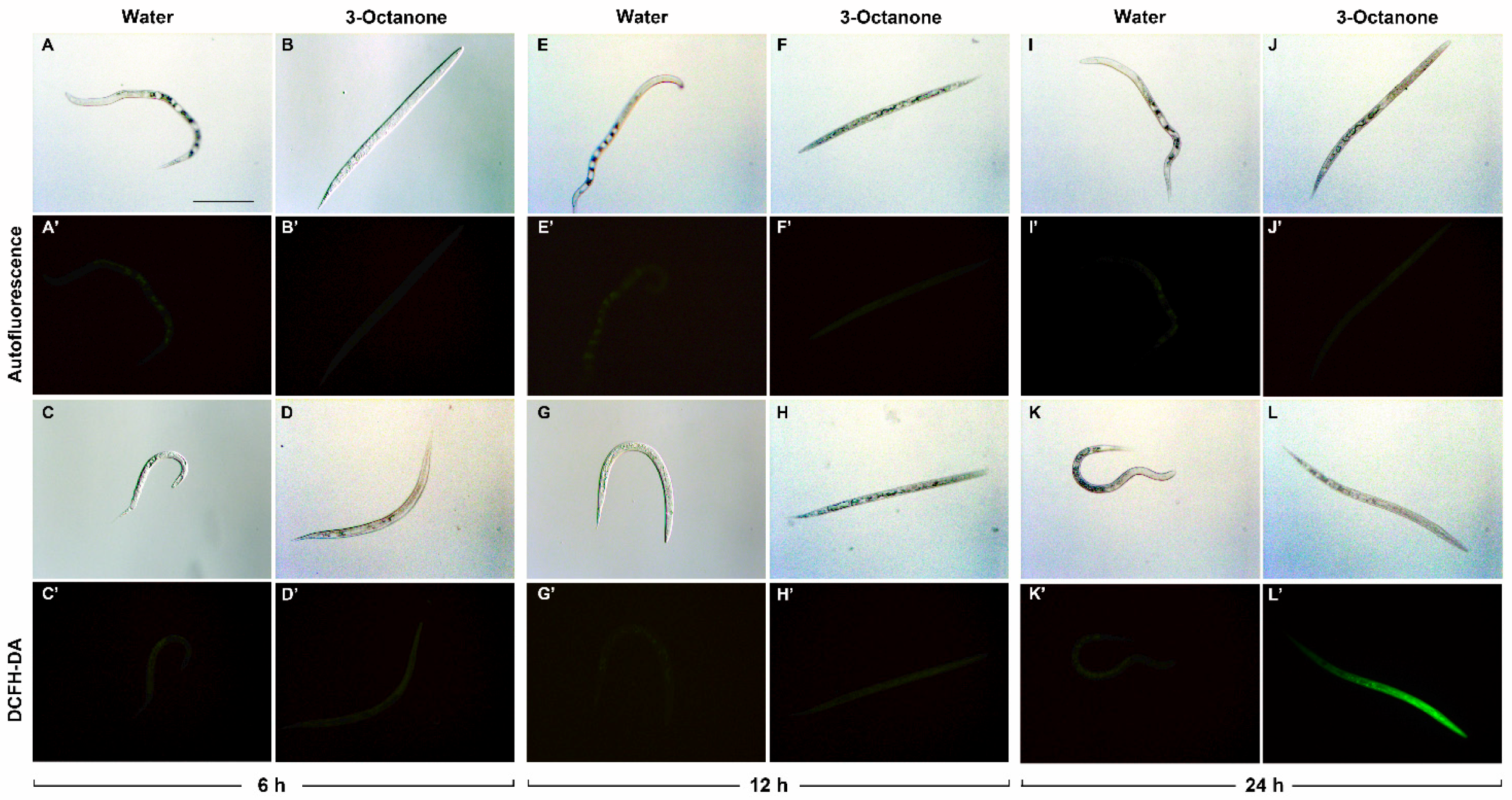
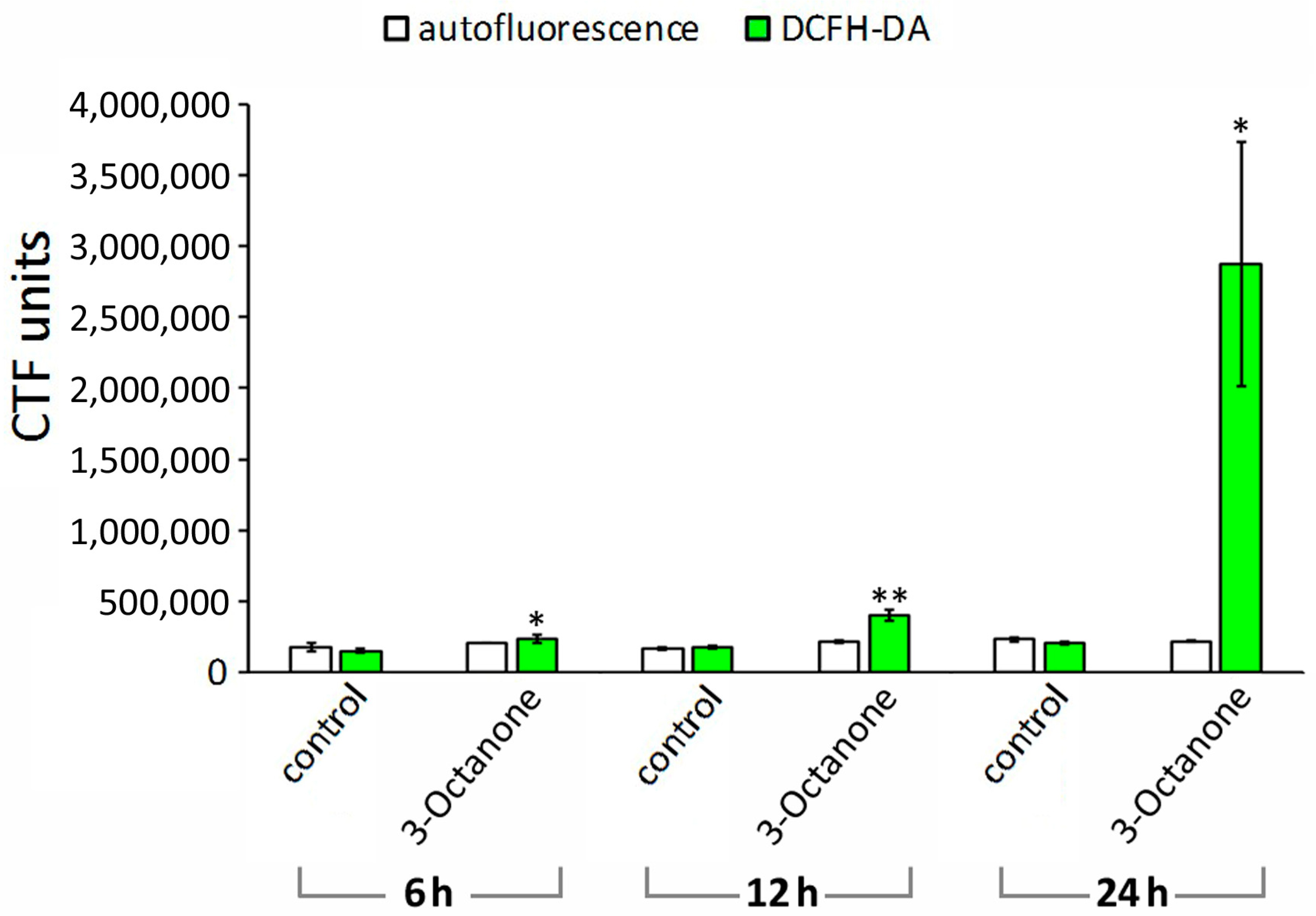
2.3. Reactive Oxygen Species Induction in M. incognita Juveniles by 1-Octen-3-ol and 3-Octanone
3. Discussion
4. Materials and Methods
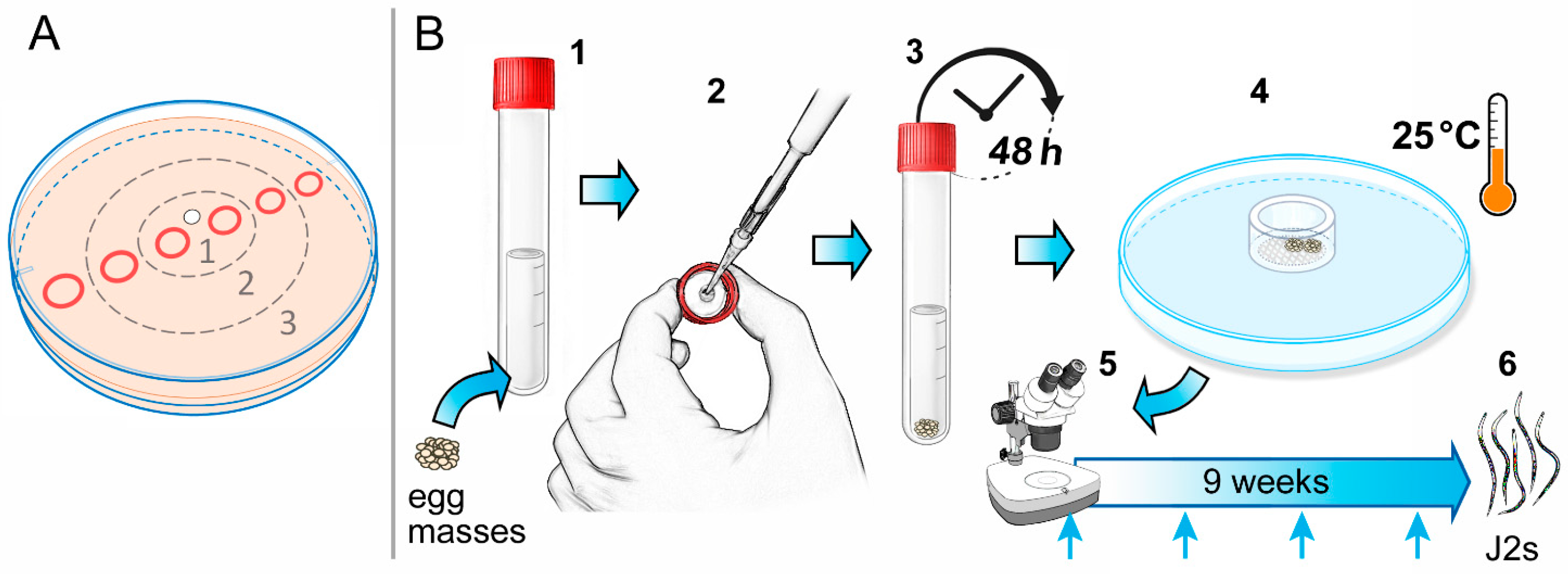
4.1. Reproduction of the Plant Parasitic Nematodes Meloidogyne incognita Race 2
4.2. Volatile Organic Compounds (VOCs)
4.3. In Vitro Tests
4.3.1. Evaluation of VOCs for Their Nematicidal Activity on M. incognita Juveniles
4.3.2. Evaluation of VOCs for Their Nematicidal Activity on M. incognita Egg Masses
4.3.3. ROS Production
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abad, P.; Gouzy, J.; Aury, J.M.; Castagnone-Sereno, P.; Danchin, E.G.J.; Deleury, E.; Perfus-Barbeoch, L.; Anthouard, V.; Artiguenave, F.; Blok, V.C.; et al. Genome sequence of the metazoan plant-parasitic nematode Meloidogyne incognita. Nat. Biotechnol. 2008, 26, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Seinhorst, J. The relation between nematode density and damage to plants. Nematologica 1965, 11, 137–154. [Google Scholar] [CrossRef]
- Ganaie, M.A.; Khan, T. Studies on the interactive effect of Meloidogyne incognita and Fusarium solani on Lycopersicon esculentum, Mill. Int. J. Bot. 2011, 7, 205–208. [Google Scholar] [CrossRef]
- Adam, M.; Heuer, H.; Hallmann, J. Bacterial antagonists of fungal pathogens also control root-knot nematodes by induced systemic resistance of tomato plants. PLoS ONE 2014, 9, e90402. [Google Scholar] [CrossRef]
- Lamelas, A.; Desgarennes, D.; López-Lima, D.; Villain, L.; Alonso-Sánchez, A.; Artacho, A.; Latorre, A.; Moya, A.; Carrión, G. The bacterial microbiome of Meloidogyne-based disease complex in coffee and tomato. Front. Plant Sci. 2020, 11, 136. [Google Scholar] [CrossRef]
- Sasanelli, N.; Ciccarese, F.; Papajová, I. Aphanocladium album by via sub-irrigation in the control of Pyrenochaeta lycopersici and Meloidogyne incognita on tomato in a plastic-house. Helminthologia 2008, 45, 137–142. [Google Scholar] [CrossRef]
- Gregory, P.J.; Johnson, S.; Newton, A.C.; Ingram, J.S.I. Integrating pests and pathogens into the climate change/food security debate. J. Exp. Bot. 2009, 60, 2827–2838. [Google Scholar] [CrossRef]
- Orgiazzi, A.; Panagos, P.; Fernández-Ugalde, O.; Wojda, P.; Labouyrie, M.; Ballabio, C.; Franco, A.; Pistocchi, A.; Montanrella, L.; Jones, A. LUCAS coil biodiversity and LUCAS soil pesticides, new tools for research and policy development. Eur. J. Soil Sci. 2022, 73, e13299. [Google Scholar] [CrossRef]
- Lewis, W.J.; van Lenteren, J.C.; Phatak, S.C.; Tumlinson, J.H. A total system approach to sustainable pest management. Proc. Natl. Acad. Sci. USA 1997, 94, 12243–12248. [Google Scholar] [CrossRef]
- Rosskopf, E.; Di Gioia, F.; Hong, J.C.; Pisani, C.; Kokalis-Burelle, N. Organic amendments for pathogen and nematode control. Annu. Rev. Phytopathol. 2020, 58, 277–311. [Google Scholar] [CrossRef]
- Djian-Caporalino, C.; Mateille, T.; Bailly-Bechet, M.; Marteu, N.; Fazari, A.; Bautheac, P.; Raptopoulo, A.; Van Duong, L.; Tavoillot, J.; Martiny, B.; et al. Evaluating sorghums as green manure against root-knot nematodes. Crop Prot. 2019, 122, 142–150. [Google Scholar] [CrossRef]
- Brennan, R.J.B.; Glaze-Corcoran, S.; Wick, R.; Hashemi, M. Biofumigation: An alternative strategy for the control of plant parasitic nematodes. J. Integr. Agric. 2020, 19, 1680–1690. [Google Scholar] [CrossRef]
- Francis, C.A.; Clegg, M.D. Crop Rotations in sustainable production system. In Sustainable Agricultural Systems; CRC Press: Boca Raton, FL, USA, 2020; ISBN 9781003070474. [Google Scholar]
- Westerdahl, B. Evaluation of trap cropping for management of root-knot nematode on annual crops. Acta Hortic. 2020, 1270, 141–146. [Google Scholar] [CrossRef]
- Nyaku, S.T.; Amissah, N. Grafting: An effective strategy for nematode management in tomato genotypes. In Recent Advances in Tomato Breeding and Production; Nyaku, S.T., Danquah, A., Eds.; InTechOpen: London, UK, 2018. [Google Scholar] [CrossRef]
- Kumari, C.; Dutta, T.K.; Banakar, P.; Rao, U. Comparing the defence-related gene expression changes upon root-knot nematode attack in susceptible versus resistant cultivars of rice. Sci. Rep. 2016, 6, 22846. [Google Scholar] [CrossRef]
- Sasanelli, N.; Konrat, A.; Migunova, V.; Toderas, I.; Iurcu-Straistaru, E.; Rusu, S.; Bivol, A.; Andoni, C.; Veronico, P. Review on control methods against plant parasitic nematodes applied in southern member states (C Zone) of the European Union. Agriculture 2021, 11, 602. [Google Scholar] [CrossRef]
- Cavoski, I.; Chami, Z.; Bouzebboudja, F.; Sasanelli, N.; Simeone, V.; Mondelli, D.; Miano, T.; Sarais, G.; Ntalli, N.; Caboni, P. Melia azedarach controls Meloidogyne incognita and triggers plant defense mechanisms on cucumber. Crop Prot. 2012, 35, 85–90. [Google Scholar] [CrossRef]
- Caboni, P.; Saba, M.; Tocco, G.; Casu, L.; Murgia, A.; Maxia, A.; Menkissoglu-Spiroudi, U.; Ntalli, N. Nematicidal activity of mint aqueous extracts against the root-knot nematode Meloidogyne incognita. J. Agric. Food Chem. 2013, 61, 9784–9788. [Google Scholar] [CrossRef]
- Renčo, M.; Sasanelli, N.; Maistrello, L. Plants as natural sources of nematicides. In Nematodes: Comparative Genomics, Disease Management and Ecological Importance; Davis, L.M., Ed.; Nova Science Publisher, Inc.: Hauppauge, NY, USA, 2014; pp. 115–141. ISBN 978-1-62948-764-9. [Google Scholar]
- Eloh, K.; Kpegba, K.; Sasanelli, N.; Koumaglo, H.K.; Caboni, P. Nematicidal activity of some essential plant oils from tropical West Africa. Int. J. Pest Manag. 2020, 66, 131–141. [Google Scholar] [CrossRef]
- Poiras, L.; Iurcu-Straistaru, E.; Poiras, N.; Burtseva, S.; Birsa, M.; Bivol, A.; Sasanelli, N. Phytosanitary expectation of pepper in greenhouses of Moldova and testing some biological agents against phytopathogens and plant parasitic nematodes. In Proceedings of the 47th Congress on Plant Protection, Chisinau, Republic of Moldova, 27–28 October 2015; pp. 146–149. [Google Scholar]
- Leoni, C.; Piancone, E.; Sasanelli, N.; Bruno, G.L.; Manzari, C.; Pesole, G.; Ceci, L.R.; Volpicella, M. Plant health and rhizosphere microbiome: Effects of the bionematicide Aphanocladium album in tomato plants infested by Meloidogyne javanica. Microorganisms 2020, 8, 1922. [Google Scholar] [CrossRef]
- Migunova, V.D.; Sasanelli, N. Bacteria as biocontrol tool against phytoparasitic nematodes. Plants 2021, 10, 389. [Google Scholar] [CrossRef]
- Thomas, G.; Withall, D.; Birkett, M. Harnessing microbial volatiles to replace pesticides and fertilizers. Microb. Biotechnol. 2020, 13, 1366–1376. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.W.; Inamdar, A.A. Are some fungal volatile organic compounds (VOCs) mycotoxins? Toxins 2015, 7, 3785–3804. [Google Scholar] [CrossRef] [PubMed]
- Khoja, S.; Eltayef, K.M.; Baxter, I.; Myrta, A.; Bull, J.C.; Butt, T. Volatiles of the entomopathogenic fungus, Metarhizium brunneum, attract and kill plant parasitic nematodes. Biol. Control. 2021, 152, 104472. [Google Scholar] [CrossRef] [PubMed]
- Bourdon, P.-A.; Zottele, M.; Baxter, I.; Myrta, A.; Midthassel, A.; Wechselberger, K.F.; Khoja, S.; Bull, J.C.; Hermann, S.; Butt, T.M. Fumigation of three major soil pests (Agriotes lineatus, Diabrotica virgifera virgifera, Phyllopertha horticola) with 3-octanone and 1-octen-3-ol enantiomers. Biocontrol Sci. Technol. 2022, 32, 863–876. [Google Scholar] [CrossRef]
- Xu, Y.-Y.; Lu, H.; Wang, X.; Zhang, K.-Q.; Li, G.-H. Effect of volatile organic compounds from bacteria on nematodes. Chem. Biodivers. 2015, 12, 1415–1421. [Google Scholar] [CrossRef]
- Zhai, Y.; Shao, Z.; Cai, M.; Zheng, L.; Li, G.; Huang, D.; Cheng, W.; Thomashow, L.S.; Weller, D.M.; Yu, Z.; et al. Multiple modes of nematode control by volatiles of Pseudomonas putida 1A00316 from Antarctic soil against Meloidogyne incognita. Front. Microbiol. 2018, 9, 253. [Google Scholar] [CrossRef]
- Bui, H.X.; Hadi, B.A.; Oliva, R.; Schroeder, N.E. Beneficial bacterial volatile compounds for the control of root-knot nematode and bacterial leaf blight on rice. Crop Prot. 2020, 135, 104792. [Google Scholar] [CrossRef]
- Bui, H.X.; Desaeger, J.A. Volatile compounds as potential bio-fumigants against plant-parasitic nematodes—A mini review. J. Nematol. 2021, 53, e2021-14. [Google Scholar] [CrossRef]
- Crespo, R.; Pedrini, N.; Juárez, M.P.; Dal Bello, G.M. Volatile organic compounds released by the entomopathogenic fungus Beauveria bassiana. Microbiol. Res. 2008, 163, 148–151. [Google Scholar] [CrossRef]
- Butt, T.; Coates, C.; Dubovskiy, I.; Ratcliffe, N. Entomopathogenic Fungi: New insights into host–pathogen interactions. Adv. Genet. 2016, 94, 307–364. [Google Scholar] [CrossRef]
- Frisvad, J. Penicillium: Penicillium/Penicillia in food production. In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C.A., Tortorello, M.L., Eds.; Academic Press: Oxford, UK, 2014; pp. 14–18. [Google Scholar] [CrossRef]
- Combet, E.; Eastwood, D.C.; Burton, K.S.; Henderson, J. Eight-carbon volatiles in mushrooms and fungi: Properties, analysis, and biosynthesis. Mycoscience 2006, 47, 317–326. [Google Scholar] [CrossRef]
- Lee, S.J.; Umano, K.; Shibamoto, T.; Lee, K.G. Identification of volatile components in basil (Ocimum basilicum L.) and thyme leaves (Thymus vulgaris L.) and their antioxidant properties. Food Chem. 2005, 91, 131–137. [Google Scholar] [CrossRef]
- Khoja, S.; Eltayef, K.M.; Baxter, I.; Bull, J.C.; Loveridge, E.J.; Butt, T. Fungal volatile organic compounds show promise as potent molluscicides. Pest Manag. Sci. 2019, 75, 3392–3404. [Google Scholar] [CrossRef]
- Chitarra, G.S.; Abee, T.; Rombouts, F.M.; Posthumus, M.A.; Dijksterhuis, J. Germination of Penicillium paneum Conidia Is Regulated by 1-Octen-3-ol, a Volatile Self-Inhibitor. Appl. Environ. Microbiol. 2004, 70, 2823–2829. [Google Scholar] [CrossRef]
- Xu, P.; Zhu, F.; Buss, G.K.; Leal, W.S. 1-Octen-3-ol—The attractant that repels. F1000Research 2015, 4, 156. [Google Scholar] [CrossRef]
- Herrera, J.M.; Pizzolitto, R.P.; Zunino, M.P.; Dambolena, J.S.; Zygadlo, J.A. Effect of fungal volatile organic compounds on a fungus and an insect that damage stored maize. J. Stored Prod. Res. 2015, 62, 74–80. [Google Scholar] [CrossRef]
- Kishimoto, K.; Matsui, K.; Ozawa, R.; Takabayashi, J. Volatile 1-octen-3-ol induces a defensive response in Arabidopsis thaliana. J. Gen. Plant Pathol. 2007, 73, 35–37. [Google Scholar] [CrossRef]
- Yang, T.; Xin, Y.; Liu, T.; Li, Z.; Liu, X.; Wu, Y.; Wang, M.; Xiang, M. Bacterial volatile-mediated suppression of root-knot nematode (Meloidogyne incognita). Plant Dis. 2022, 106, 1358–1365. [Google Scholar] [CrossRef]
- Diyapoglu, A.; Oner, M.; Meng, M. Application potential of bacterial volatile organic compounds in the control of root-knot nematodes. Molecules 2022, 27, 4355. [Google Scholar] [CrossRef]
- Deng, X.; Wang, X.; Li, G. Nematicidal effects of volatile organic compounds from microorganisms and plants on plant-parasitic nematodes. Microorganisms 2022, 10, 1201. [Google Scholar] [CrossRef]
- Eisenback, J.D.; Hunt, D.J. General morphology. In Root-knot Nematodes; Perry, R.N., Moens, M., Starr, F.J., Eds.; CAB International: Wallingford, UK, 2009; p. 23. [Google Scholar]
- Terra, W.C.; Campos, V.P.; Martins, S.J.; Costa, L.S.A.S.; da Silva, J.C.P.; Barros, A.F.; Lopez, L.E.; Santos, T.C.N.; Smant, G.; Oliveira, D.F. Volatile organic molecules from Fusarium oxysporum strain 21 with nematicidal activity against Meloidogyne incognita. Crop Prot. 2018, 106, 125–131. [Google Scholar] [CrossRef]
- Mei, X.; Wang, X.; Li, G. Pathogenicity and volatile nematicidal metabolites from Duddingtonia flagrans against Meloidogyne incognita. Microorganisms 2021, 9, 2268. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Yu, C.; Shao, Z.; Cai, M.; Li, G.; Zheng, L.; Yu, Z.; Zhang, J. Identification and characterization of nematicidal volatile organic compounds from deep-sea Virgibacillus dokdonensis MCCC 1A00493. Molecules 2020, 25, 744. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Yang, J.; Nie, Q.; Huang, D.; Yu, C.; Zheng, L.; Cai, M.; Thomashow, L.S.; Weller, D.M.; Yu, Z.; et al. Volatile organic compounds from Paenibacillus polymyxa KM2501-1 control Meloidogyne incognita by multiple strategies. Sci. Rep. 2017, 7, 16213. [Google Scholar] [CrossRef]
- Hummadi, E.H.; Cetin, Y.; Demirbek, M.; Kardar, N.M.; Khan, S.; Coates, C.J.; Eastwood, D.C.; Dudley, E.; Maffeis, T.; Loveridge, J.; et al. Antimicrobial volatiles of the insect pathogen Metarhizium brunneum. J. Fungi 2022, 8, 326. [Google Scholar] [CrossRef]
- Macedo, G.E.; Vieira, P.D.B.; Rodrigues, N.R.; Gomes, K.K.; Rodrigues, J.F.; Franco, J.L.; Posser, T. Effect of fungal indoor air pollutant 1-octen-3-ol on levels of reactive oxygen species and nitric oxide as well as dehydrogenases activities in Drosophila melanogaster males. J. Toxicol. Environ. Health Part A 2022, 85, 573–585. [Google Scholar] [CrossRef]
- Ayaz, M.; Ali, Q.; Farzand, A.; Khan, A.R.; Ling, H.; Gao, X. Nematicidal volatiles from Bacillus atrophaeus GBSC56 Promote growth and stimulate induced systemic resistance in tomato against Meloidogyne incognita. Int. J. Mol. Sci. 2021, 22, 5049. [Google Scholar] [CrossRef]
- Hartman, K.M.; Sasser, J.N. Identification of Meloidogyne species on the basis of differential host test and perineal pattern morphology. In An Advanced Treatise on Meloidogyne; Sasser, J.N., Carter, C.C., Eds.; North Carolina State University: Raleigh, NC, USA, 1985; Volume 2, pp. 69–77. [Google Scholar]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 18827, 1-Octen-3-OL PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/1-Octen-3-OL (accessed on 20 February 2023).
- National Center for Biotechnology Information. PubChem Compound Summary for CID 246728, 3-Octanone PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/3-Octanone (accessed on 20 February 2023).
- Puntener, W.; Zahner, O. Manual for Field Trials in Plant Protection; Ciba-Geigy: Basel, Switzerland, 1981; p. 205. [Google Scholar]
- Hussey, R.S.; Barker, K.R. A comparison of methods of collecting inocula of Meloidogyne spp. including a new technique. Plant Dis. Rep. 1973, 57, 1025–1028. [Google Scholar]
- Finney, D.J. Estimation of the median effective dose. In Probit Analysis, 3rd ed.; Cambridge University Press: Cambridge, UK, 1971; pp. 20–49. [Google Scholar]
- Stevens, W.L. Tables of the angular transformation. Biometrika 1953, 40, 70–73. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exposure Time | Doses (µL) | LSD | |||||
|---|---|---|---|---|---|---|---|
| 2.5 | 5.0 | 10.0 | 20.0 | 0.05 | 0.01 | ||
| 45 min | 38.0 * ± 9.8 ** | 40.0 ± 11.8 | 46.6 ± 16.2 | 60.0 ± 8.9 | 22.6 | 33.0 | |
| 1.5 h | 46.7 ± 18.9 | 48.7 ± 20.5 | 50.0 ± 20.5 | 63.7 ± 17.0 | 36.3 | 52.8 | |
| 3 h | 65.7 ± 15.5 | 68.0 ± 12.6 | 74.6 ± 20.5 | 77.7 ± 12.3 | 29.8 | 42.7 | |
| 6 h | 74.3 ± 6.5 | 69.3 ± 16.8 | 82.3 ± 3.2 | 82.7 ± 3.2 | 17.5 | 25.5 | |
| 12 h | 89.3 ± 3.2 | 96.0 ± 3.5 | 97.3 ± 1.2 | 99.3 ± 1.2 | 4.7 | 6.8 | |
| 24 h | 99.7 ± 0.6 | 100.0 ± 0.0 | 99.3 ± 1.2 | 100.0 ± 0.0 | 1.2 | 1.8 | |
| LSD | 0.05 | 19.9 | 23.1 | 24.3 | 16.8 | --- | --- |
| 0.01 | 27.8 | 32.4 | 34.1 | 23.5 | --- | --- | |
| Exposure Time | Doses (µL) | LSD | |||||
|---|---|---|---|---|---|---|---|
| 2.5 | 5.0 | 10.0 | 20.0 | 0.05 | 0.01 | ||
| 45 min | 18.0 * ± 9.2 ** | 52.3 ± 11.7 | 55.7 ± 10.3 | 61.0 ± 5.6 | 17.8 | 25.9 | |
| 1.5 h | 4.3 ± 2.9 | 33.0 ± 9.5 | 60.0 ± 2.0 | 65.0 ± 3.6 | 10.1 | 14.8 | |
| 3 h | 10.3 ± 6.1 | 51.3 ± 11.0 | 92.7 ± 1.5 | 98.7 ± 1.5 | 12.0 | 17.5 | |
| 6 h | 38.3 ± 10.8 | 64.7 ± 10.7 | 95.0 ± 1.0 | 99.0 ± 1.2 | 14.4 | 20.9 | |
| 12 h | 80.3 ± 3.1 | 85.3 ± 3.2 | 97.0 ± 1.0 | 99.3 ± 1.0 | 4.4 | 6.4 | |
| 24 h | 88.0 ± 3.0 | 92.3 ± 3.2 | 100.0 ± 0.0 | 100.0 ± 0.0 | 4.1 | 6.0 | |
| LSD | 0.05 | 11.8 | 16.0 | 7.7 | 5.1 | --- | --- |
| 0.01 | 16.5 | 22.4 | 10.8 | 7.1 | --- | --- | |
| Time | LD50 (μL) | Dose (μL) | LT50 (min) | ||
|---|---|---|---|---|---|
| 1-Octen-3-ol | 3-Octanone | 1-Octen-3-ol | 3-Octanone | ||
| 45 min | 10.1 (6–17) * | 8.5 (7–11) | 2.5 | 91.6 (73–114) | 399.3 (343–464) |
| 1.5 h | 5.1 (2–10) | 10.0 (8–12) | 5.0 | 86.2 (69–107) | 106.5 (80–141) |
| 3 h | 0.3 (0.01–6) | 4.9 (4–5) | 10.0 | 66.8 (53–85) | 43.1 (33–57) |
| 6 h | 0.2 (0.005–5) | 3.3 (3–4) | 20.0 | 40.1 (28–57) | 39.6 (31–51) |
| 12 h | 0.2 (0.03–1.5) | 0.8 (0.4–1.7) | |||
| 24 h | --- ** | 0.6 (0.2–1.9) | |||
| Average | 3.2 | 4.6 | 71.2 | 147.1 | |
| Correlations | y = bxm | y = bxm | y = bxm | y = bxm | |
| b = 3665 | b = 375.7 | b = 146.6 | b = 842.75 | ||
| m = −1.598 | m = −0.875 | m = −0.394 | m = −1.13 | ||
| r = −0.92 | r = −0.95 | r = −0.94 | r = −0.94 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veronico, P.; Sasanelli, N.; Troccoli, A.; Myrta, A.; Midthassel, A.; Butt, T. Evaluation of Fungal Volatile Organic Compounds for Control the Plant Parasitic Nematode Meloidogyne incognita. Plants 2023, 12, 1935. https://doi.org/10.3390/plants12101935
Veronico P, Sasanelli N, Troccoli A, Myrta A, Midthassel A, Butt T. Evaluation of Fungal Volatile Organic Compounds for Control the Plant Parasitic Nematode Meloidogyne incognita. Plants. 2023; 12(10):1935. https://doi.org/10.3390/plants12101935
Chicago/Turabian StyleVeronico, Pasqua, Nicola Sasanelli, Alberto Troccoli, Arben Myrta, Audun Midthassel, and Tariq Butt. 2023. "Evaluation of Fungal Volatile Organic Compounds for Control the Plant Parasitic Nematode Meloidogyne incognita" Plants 12, no. 10: 1935. https://doi.org/10.3390/plants12101935
APA StyleVeronico, P., Sasanelli, N., Troccoli, A., Myrta, A., Midthassel, A., & Butt, T. (2023). Evaluation of Fungal Volatile Organic Compounds for Control the Plant Parasitic Nematode Meloidogyne incognita. Plants, 12(10), 1935. https://doi.org/10.3390/plants12101935