
Seasonal Variation in Selected Biochemical Traits in the Leaves of Co-Occurring Invasive and Native Plant Species under Mediterranean Conditions




Abstract
:1. Introduction
2. Materials and Methods
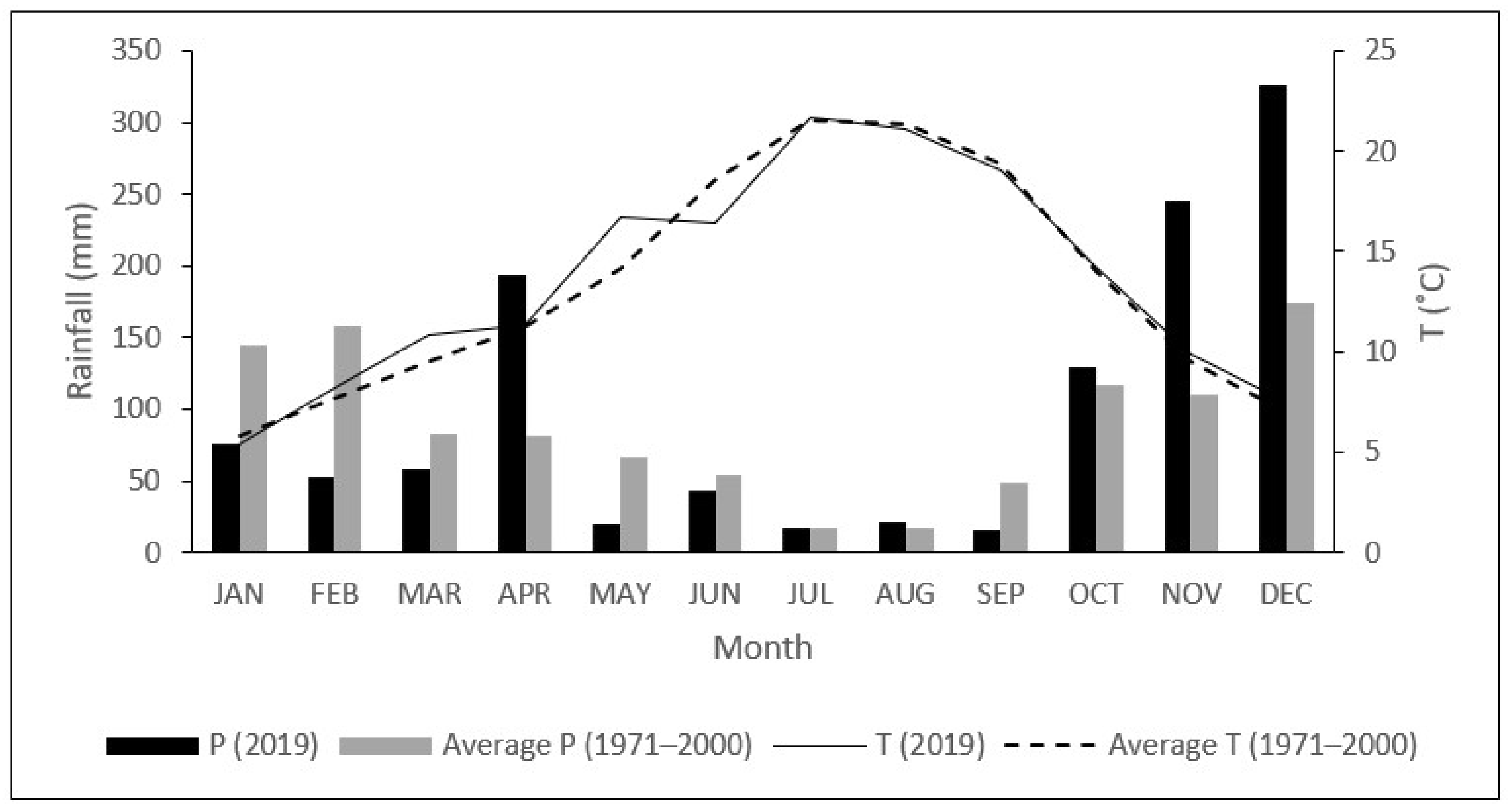
2.1. Study Site and Climatic Conditions
2.2. Sampling Site and Data Collection
2.3. Biochemical Determinations
2.3.1. Photosynthetic Pigments
2.3.2. Total Soluble Sugars and Starch Contents
2.3.3. Total Phenolics Content
2.3.4. Lipid Peroxidation
2.4. Statistical Analysis
3. Results
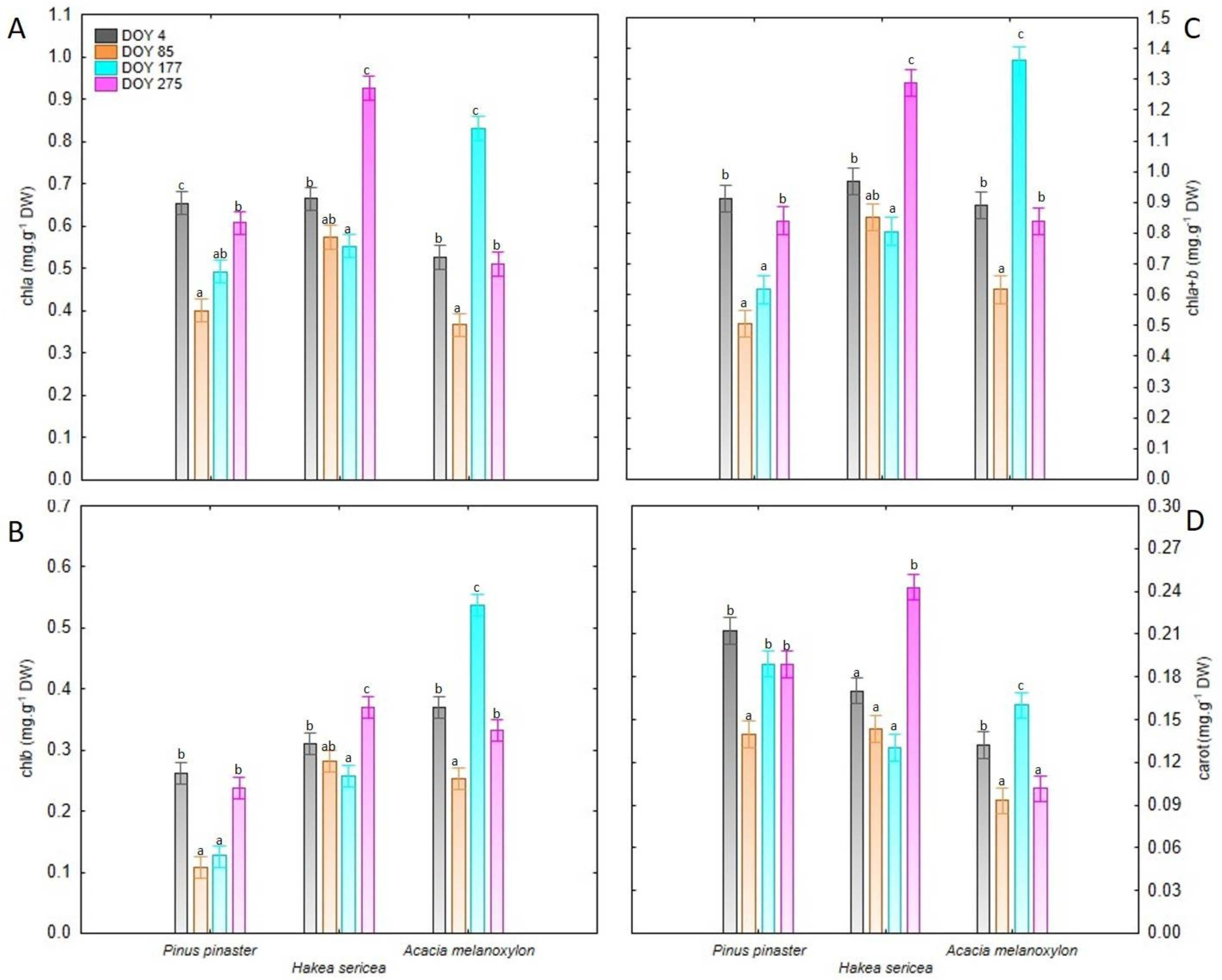
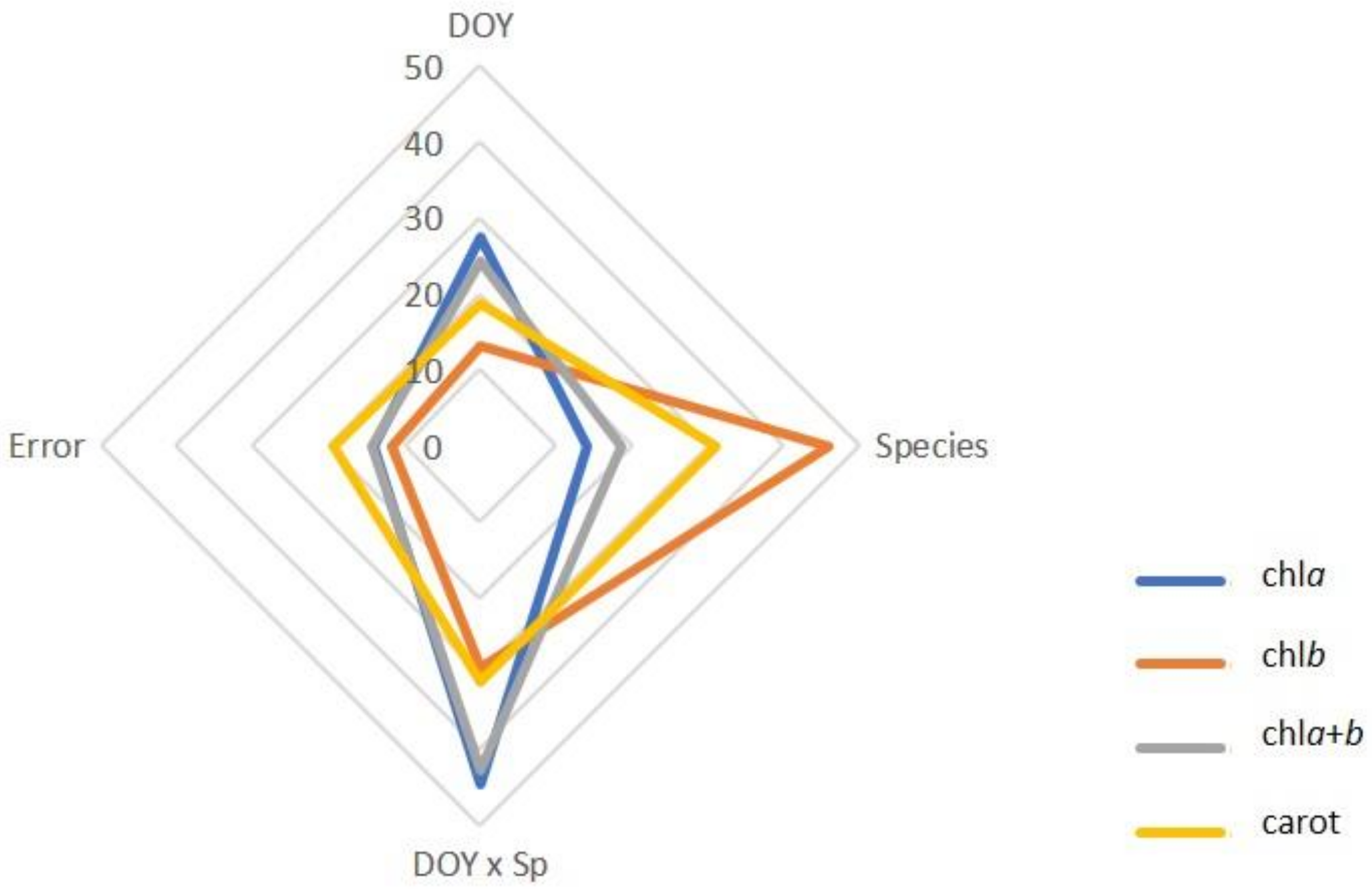
3.1. Leaf Photosynthetic Pigments
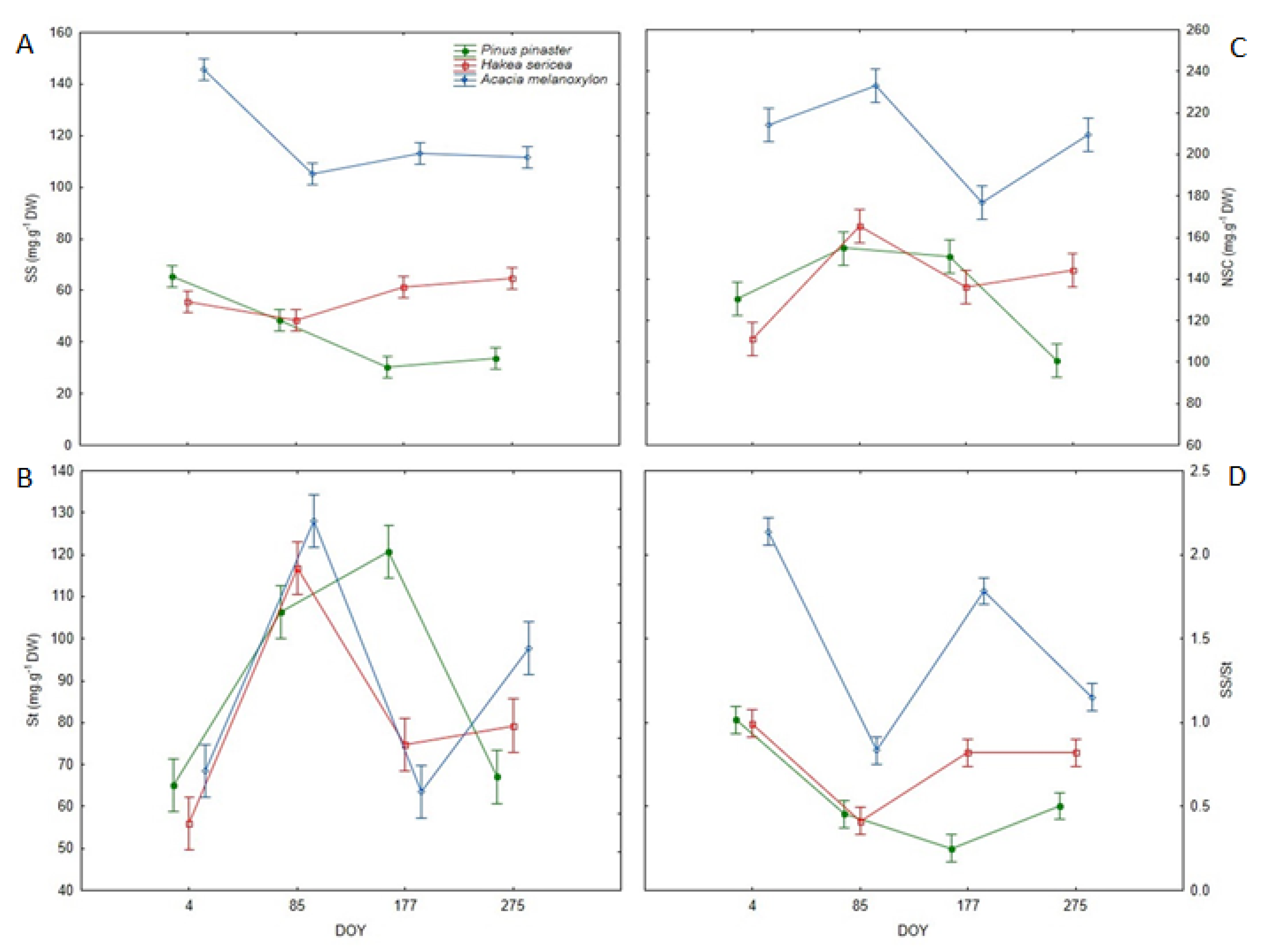
3.2. Non-Structural Carbohydrates
3.3. Total Phenolic and MDA Contents
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pyšek, P.; Blackburn, T.M.; García-Berthou, E.; Perglová, I.; Rabitsch, W. Displacement and local extinction of native and endemic species. In Impact of Biological Invasions on Ecosystem Services; Vilà, M., Hulme, P., Eds.; Springer: Berlin, Germany, 2017; Volume 12, pp. 157–175. [Google Scholar] [CrossRef]
- Bellard, C.; Cassey, P.; Blackburn, T.M. Alien species as a driver of recent extinctions. Biol. Lett. 2016, 12, 20150623. [Google Scholar] [CrossRef] [PubMed]
- Mazza, G.; Tricarico, E.; Genovesi, P.; Gherardi, F. Biological invaders are threats to human health: An overview. Ethol. Ecol. Evol. 2014, 26, 112–129. [Google Scholar] [CrossRef]
- Nentwig, W.; Mebs, D.; Vilà, M. Impact of non-native animals and plants on human health. In Impact of Biological Invasions on Ecosystem Services; Vilà, M., Hulme, P., Eds.; Springer: Berlin, Germany, 2017; Volume 12, pp. 277–293. [Google Scholar] [CrossRef]
- Haubrock, P.J.; Turbelin, A.J.; Cuthbert, R.N.; Novoa, A.; Taylor, N.G.; Angulo, E.; Ballesteros-Mejia, L.; Bodey, T.W.; Capinha, C.; Diagne, C.; et al. Economic costs of invasive alien species across Europe. NeoBiota 2021, 67, 153–190. [Google Scholar] [CrossRef]
- Pyšek, P.; Richardson, D.M. Traits associated with invasiveness in alien plants: Where do we stand? In Biological Invasions; Nentwig, W., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; Volume 193, pp. 97–125. [Google Scholar] [CrossRef]
- Davidson, A.M.; Jennions, M.; Nicotra, A.B. Do invasive species show high phenotypic plasticity than native species and, if so, is it adapt? A meta-analysis. Ecol. Lett. 2011, 14, 419–431. [Google Scholar] [CrossRef]
- Marchante, E.; Marchante, H. Engaging society to fight invasive alien plants in Portugal—One of the main threats to biodiversity. In Biodiversity and Education for Sustainable Development; Castro, P., Azeiteiro, U., Bacelar-Nicolau, P., Leal Filho, W., Azul, A., Eds.; Springer: Cham, Switzerland, 2016; pp. 107–122. [Google Scholar] [CrossRef]
- Marchante, H.; Morais, M.; Freitas, H.; Marchante, E. Guia Prático para a Identificação de Plantas Invasoras em Portugal; Imprensa da Universidade de Coimbra: Coimbra, Portugal, 2014. [Google Scholar]
- Arnaud, A.; Chapman, D.; Roux, J.; Linnamagi, M.; Marchante, E.; Pasiecznik, N.; Pescott, O.; Singh, I.; Starfinger, U.; Vicente, J.; et al. Hakea sericea Schrad. & J.C. Wendl. Bull. OEPP/EPPO Bull. 2019, 49, 273–279. [Google Scholar]
- Pasiecznik, N. Hakea sericea. In Invasive Species Compendium; CAB International: South Oxfordshire, UK, 2022. [Google Scholar]
- CABI. Acacia melanoxylon. In Invasive Species Compendium; CAB International: South Oxfordshire, UK, 2022. [Google Scholar]
- González, L.; Souto, X.C.; Reigosa, M.J. Allelopathic effects of Acacia melanoxylon R. Br. phyllodes during their decomposition. For. Ecol. Manag. 1995, 77, 53–63. [Google Scholar] [CrossRef]
- Hussain, M.I.; El-Sheikh, M.A.; Reigosa, M.J. Allelopathic potential of aqueous extract from Acacia melanoxylon R. Br. on Lactuca sativa. Plants 2020, 9, 1228. [Google Scholar] [CrossRef]
- McDowell, S.C. Photosynthetic characteristics of invasive and noninvasive species of Rubus (Rosaceae). Am. J. Bot. 2002, 89, 1431–1438. [Google Scholar] [CrossRef]
- Souza, A.; Sandrin, C.Z.; Calió, M.F.A.; Meirelles, S.T.; Pivello, V.R.; Figueiredo-Ribeiro, R.C.L. Seasonal variation of soluble carbohydrates and starch in Echinolaena inflexa, a native grass species from the Brazilian savanna, and in the invasive grass Melinis minutiflora. Braz. J. Biol. 2010, 70, 395–404. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Wang, L.; Tian, B.; Ding, J.; Siemann, E. Introduced populations of an invasive tree have higher soluble sugars but lower starch and cellulose. Front. Plant Sci. 2020, 11, 587414. [Google Scholar] [CrossRef]
- Oliveira, M.T.; Matzek, V.; Medeiros, C.D.; Rivas, R.; Falcão, H.M.; Santos, M.G. Stress tolerance and ecophysiological ability of an invader and a native species in a seasonally dry tropical forest. PLoS ONE 2014, 9, e105514. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Zhang, T.-J.; Li, B.-H.; Liang, W.-J.; Zhang, Q.-L.; Cai, M.-L.; Peng, C.-L. Strong response of stem photosynthesis to defoliation in Mikania micrantha highlights the contribution of phenotypic plasticity to plant invasiveness. Front. Plant Sci. 2021, 12, 638796. [Google Scholar] [CrossRef] [PubMed]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef] [PubMed]
- Colautti, R.I.; Barrett, S.C.H. Rapid adaptation to climate facilitates range expansion of an invasive plant. Science 2013, 342, 364–366. [Google Scholar] [CrossRef]
- Ramírez-Briones, E.; Rodríguez-Macías, R.; Salcedo-Pérez, E.; Ramírez-Chávez, E.; Molina-Torres, J.; Tiessen, A.; Ordaz-Ortiz, J.; Martínez-Gallardo, N.; Délano-Frier, J.P.; Zañudo-Hernández, J. Seasonal changes in the metabolic profiles and biological activity in leaves of Diospyros digyna and D. rekoi “Zapote” trees. Plants 2019, 8, 449. [Google Scholar] [CrossRef] [Green Version]
- Plazaola, G.; Ignacio, J.; Becerril, J. Seasonal changes in photosynthetic pigments and antioxidants in beech (Fagus sylvatica) in a Mediterranean climate: Implications for tree decline diagnosis. Funct. Plant Biol. 2001, 28, 225–232. [Google Scholar] [CrossRef]
- Martinez Vilalta, J.; Sala, A.; Asensio, D.; Galiano, L.; Hoch, G.; Palacio, S.; Piper, F.; Lloret, F. Dynamics of non-structural carbohydrates in terrestrial plants: A global synthesis. Ecol. Monogr. 2016, 86, 495–516. [Google Scholar] [CrossRef]
- Lloret, F.; Sapes, G.; Rosas, T.; Galiano, L.; Saura-Mas, S.; Sala, A.; Martínez-Vilalta, J. Non-structural carbohydrate dynamics associated with drought-induced die-off in woody species of a shrubland community. Ann. Bot. 2018, 121, 1383–1396. [Google Scholar] [CrossRef] [Green Version]
- Tóth, V.R.; Villa, P.; Pinardi, M.; Bresciani, M. Aspects of invasiveness of Ludwigia and Nelumbo in shallow temperate fluvial lakes. Front. Plant Sci. 2019, 10, 647. [Google Scholar] [CrossRef]
- Rivas, R.; Barros, V.; Falcão, H.; Frosi, G.; Arruda, E.; Santos, M. Ecophysiological traits of invasive C3 species Calotropis procera to maintain high photosynthetic performance under high VPD and low soil water balance in semi-arid and seacoast zones. Front. Plant Sci. 2020, 11, 717. [Google Scholar] [CrossRef]
- Davidson, A.M.; Le, S.T.; Cooper, K.B.; Lange, E.; Zwieniecki, M. No time to rest: Seasonal dynamics of non-structural carbohydrates in twigs of three Mediterranean tree species suggest year-round activity. Sci. Rep. 2021, 11, 5181. [Google Scholar] [CrossRef] [PubMed]
- Tixier, A.; Guzmán-Delgado, P.; Sperling, O.; Roxas, A.A.; Laca, E.; Zwieniecki, M.A. Comparison of phenological traits, growth patterns, and seasonal dynamics of non-structural carbohydrate in Mediterranean tree crop species. Sci. Rep. 2020, 10, 347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gleason, S.M.; Ares, A. Photosynthesis, carbohydrate storage and survival of a native and an introduced tree species in relation to light and defoliation. Tree Physiol. 2004, 24, 1087–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramírez-Briones, E.; Rodríguez-Macías, R.; Salcedo-Pérez, E.; Martínez-Gallardo, N.; Tiessen, A.; Molina-Torres, J.; Délano-Frier, J.P.; Zãnudo-Hernández, J. Seasonal variation in non-structural carbohydrates, sucrolytic activity and secondary metabolites in deciduous and perennial Diospyros species sampled in Western Mexico. PLoS ONE 2017, 12, e0187235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gori, A.; Tattini, M.; Centritto, M.; Ferrini, F.; Marino, G.; Mori, J.; Guidi, L.; Brunetti, C. Seasonal and daily variations in primary and secondary metabolism of three maquis shrubs unveil different adaptive responses to Mediterranean climate. Conserv. Physiol. 2019, 7, coz070. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Lelarge-Trouverie, C.; Mhamdi, A. The metabolomics of oxidative stress. Phytochemistry 2015, 112, 33–53. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wen, K.-S.; Ruan, X.; Zhao, Y.-X.; Wei, F.; Wang, Q. Response of plant secondary metabolites to environmental factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [Green Version]
- Sesták, Z.; Castky, J.; Jarvis, P.G. Plant Photosynthetic Production: Manual of Methods; Dr. W. Junk Publishers: The Hague, The Netherlands, 1971. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Irigoyen, J.J.; Emerich, D.W.; Sánchez-Díaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativa) plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Osaki, M.; Shinano, T.; Tadano, T. Redistribution of carbon and nitrogen compounds from the shoot to the harvesting organs during maturation in field crops. Soil Sci. Plant Nutr. 1991, 37, 117–128. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.Á. Colorometry of total phenolics with phosphomolybdicphosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Heat, R.L.; Packer, L. Peroxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar]
- Llorens, L.; Peñuelas, J.; Filella, I. Diurnal and seasonal variations in the photosynthetic performance and water relations of two co-occurring Mediterranean shrubs, Erica multiflora and Globularia alypum. Physiol. Plant. 2003, 118, 84–95. [Google Scholar] [CrossRef] [Green Version]
- Ain-Lhout, F.; Díaz, M.; Zunzunegui, M.; Rodríguez, H.; Novo, F.; Vargas, M.A. Seasonal differences in photochemical efficiency and chlorophyll and carotenoid contents in six Mediterranean shrub species under field conditions. Photosynthetica 2004, 42, 399–407. [Google Scholar] [CrossRef]
- Deligöz, A.; Bayar, E.; Genç, M.; Karatepe, Y.; Kırdar, E.; Cankara, F.G. Seasonal and needle age-related variations in the biochemical characteristics of Pinus nigra subsp. pallasiana (Lamb.) Holmboe. J. For. Sci. 2018, 64, 379–386. [Google Scholar] [CrossRef] [Green Version]
- González-Rodríguez, Á.M.; Brito, P.; Lorenzo, J.R.; Jiménez, M.S. Photosynthetic performance in Pinus canariensis at semiarid treeline: Phenotype variability to cope with stressful environment. Forests 2019, 10, 845. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Singh, H.; Sharma, S.K.; Nautiyal, R. Seasonal variation in biochemical responses of bamboo clones in the sub-tropical climate of Indian Himalayan foothills. Heliyon 2021, 7, e06859. [Google Scholar] [CrossRef] [PubMed]
- Havaux, M. Carotenoid oxidation products as stress signals in plants. Plant J. 2014, 79, 597–606. [Google Scholar] [CrossRef]
- Hernández, I.; Alegre, L.; Munné-Bosch, S. Drought-induced changes in flavonoids and other low molecular weight antioxidants in Cistus clusii grown under Mediterranean field conditions. Tree Physiol. 2004, 24, 1303–1311. [Google Scholar] [CrossRef] [Green Version]
- Scartazza, A.; Moscatello, S.; Matteucci, G.; Battistelli, A.; Brugnoli, E. Seasonal and inter-annual dynamics of growth, non-structural carbohydrates and C stable isotopes in a Mediterranean beech forest. Tree Physiol. 2013, 33, 730–742. [Google Scholar] [CrossRef] [Green Version]
- Dong, S.; Beckles, D.M. Dynamic changes in the starch-sugar interconversion within plant source and sink tissues promote a better abiotic stress response. J. Plant Physiol. 2019, 234–235, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Cavender-Bares, J.; Cortes, P.; Rambal, S.; Joffre, R.; Miles, B.; Rocheteau, A. Summer and winter sensitivity of leaves and xylem to minimum freezing temperatures: A comparison of co-occurring Mediterranean oaks that differ in leaf lifespan. New Phytol. 2005, 168, 597–612. [Google Scholar] [CrossRef] [PubMed]
- Ugarte, R.M.; Escudero, A.; Gavilán, R.G. Assessing the role of selected osmolytes in Mediterranean high-mountain specialists. Front. Ecol. Evol. 2021, 9, 576122. [Google Scholar] [CrossRef]
- Liu, W.; Su, J.; Lang, X.; Huang, X. Non-structural carbohydrates regulated by season and species in the subtropical monsoon broad-leaved evergreen forest of Yunnan Province, China. Sci. Rep. 2018, 8, 1083. [Google Scholar] [CrossRef] [PubMed]
- Palacio, S.; Maestro, M.; Montserrat-Martí, G. Seasonal dynamics of non-structural carbohydrates in two species of Mediterranean sub-shrubs with different leaf phenology. Environ. Exp. Bot. 2007, 59, 34–42. [Google Scholar] [CrossRef]
- Funk, J.L. The physiology of invasive plants in low-resource environments. Conserv. Physiol. 2013, 1, cot026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potijun, S.; Yaisamlee, C.; Sirikhachornkit, A. Pigment production under cold stress in the green microalga Chlamydomonas reinhardtii. Agriculture 2021, 11, 564. [Google Scholar] [CrossRef]
- Zunzunegui, M.; Ruiz-Valdepeñas, E.; Sert, M.A.; Díaz-Barradas, M.C.; Gallego-Fernández, J.B. Field comparison of ecophysiological traits between an invader and a native species in a Mediterranean coastal dune. Plant Physiol. Biochem. 2020, 146, 278–286. [Google Scholar] [CrossRef]
- Kumar, A.; Memo, M.; Mastinu, A. Plant behaviour: An evolutionary response to the environment? Plant Biol. 2020, 22, 961–970. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | chla (mg.g−1 DW) | chlb (mg.g−1 DW) | chla+b (mg.g−1 DW) | carot (mg.g−1 DW) | |
| Time | DOY 4 | 0.615 ± 0.024 b | 0.314 ± 0.017 b | 0.924 ± 0.032 b | 0.171 ± 0.009 b |
| DOY 85 | 0.447 ± 0.026 a | 0.214 ± 0.020 a | 0.658 ± 0.041 a | 0.125 ± 0.007 a | |
| DOY 177 | 0.625 ± 0.037 bc | 0.307 ± 0.042 b | 0.928 ± 0.079 b | 0.160 ± 0.008 b | |
| DOY 275 | 0.681 ± 0.045 c | 0.313 ± 0.016 b | 0.990 ± 0.056 b | 0.178 ± 0.015 b | |
| Species | Pinus pinaster | 0.539 ± 0.026 A | 0.183 ± 0.018 A | 0.719 ± 0.043 A | 0.182 ± 0.007 B |
| Hakea sericea | 0.680 ± 0.033 B | 0.305 ± 0.010 B | 0.979 ± 0.043 B | 0.172 ± 0.010 B | |
| Acacia melanoxylon | 0.559 ± 0.036 A | 0.373 ± 0.023 C | 0.927 ± 0.059 B | 0.122 ± 0.006 A | |
| Two-way Repeated measures ANOVA | |||||
| DOY | *** | *** | *** | *** | |
| Species (Sp) | *** | *** | *** | *** | |
| DOY × Sp | *** | *** | *** | *** | |
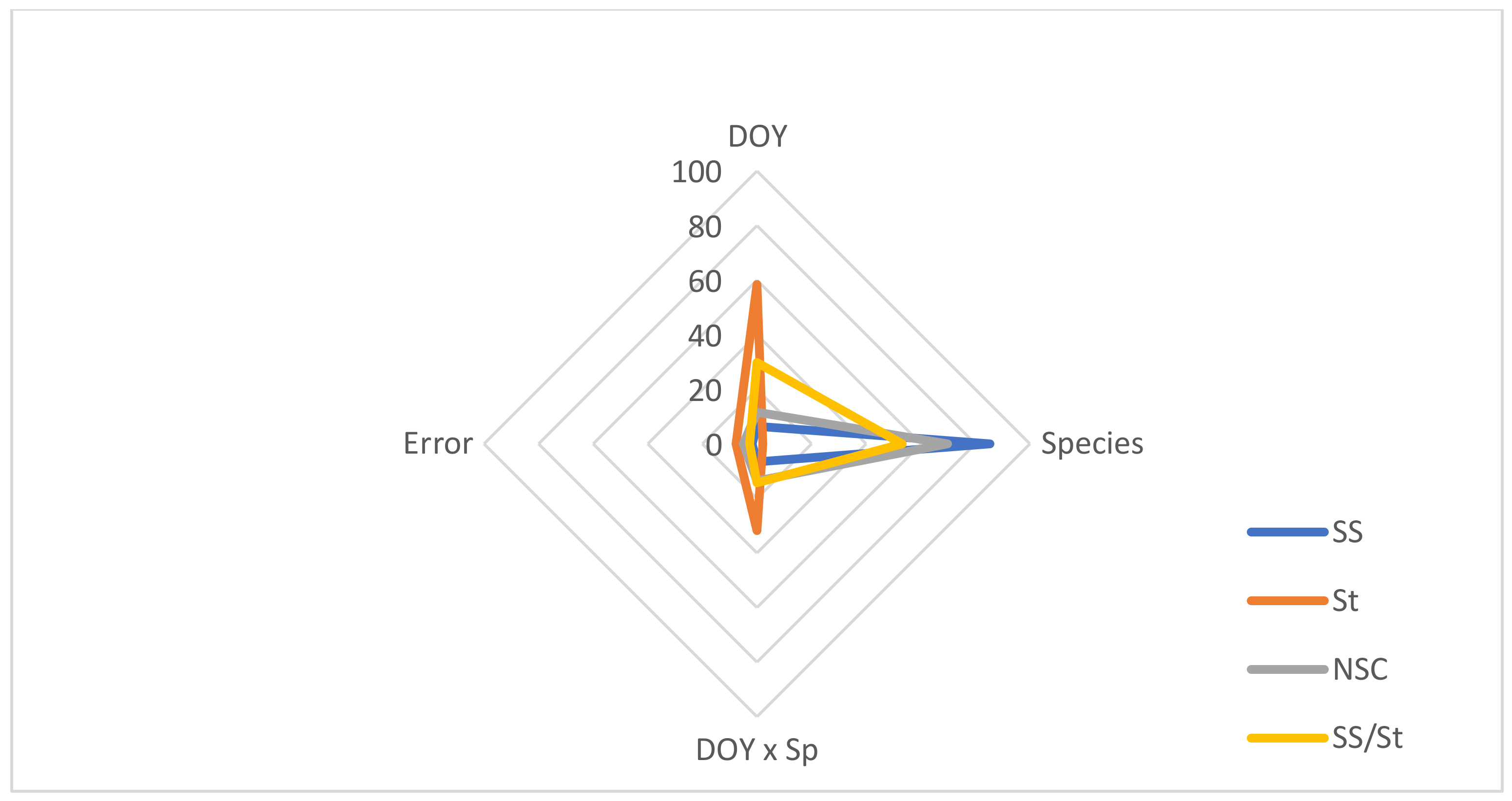
| Source of Variation | SS | St | NSC | SS/St | |||||
|---|---|---|---|---|---|---|---|---|---|
| d.f. | F | p | F | p | F | p | F | p | |
| DOY | 3 | 71.937 | *** | 129.692 | *** | 41.940 | *** | 203.702 | *** |
| Species (Sp) | 2 | 1454.332 | *** | 18.372 | *** | 677.127 | *** | 702.720 | *** |
| DOY × Sp | 6 | 37.566 | *** | 35.260 | *** | 24.655 | *** | 49.402 | *** |
| Biochemical Trait | DOY | Species | ||
|---|---|---|---|---|
| Pinus pinaster | Hakea sericea | Acacia melanoxylon | ||
| Total phenolics (mg g−1 DW) | 4 | 47.848 ± 2.756 b | 27.719 ± 0.944 a | 32.161 ± 0.311 a |
| 85 | 23.523 ± 1.691 ab | 20.745 ± 0.860 a | 27.672 ± 0.889 b | |
| 177 | 40.447 ± 0.808 b | 35.965 ± 2.410 b | 30.209 ± 0.711 a | |
| 275 | 32.115 ± 1.059 a | 32.778 ± 0.932 a | 32.124 ± 0.726 a | |
| MDA (nmol.mg−1 DW) | 4 | 0.605 ± 0.007 c | 0.115 ± 0.002 a | 0.264 ± 0.009 b |
| 85 | 0.272 ± 0.019 b | 0.045 ± 0.001 a | 0.376 ± 0.003 c | |
| 177 | 0.663 ± 0.026 c | 0.085 ± 0.002 a | 0.258 ± 0.005 b | |
| 275 | 0.326 ± 0.011 b | 0.058 ± 0.001 a | 0.310 ± 0.003 b | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morais, M.C.; Cabral, J.A.; Gonçalves, B. Seasonal Variation in Selected Biochemical Traits in the Leaves of Co-Occurring Invasive and Native Plant Species under Mediterranean Conditions. Plants 2022, 11, 1171. https://doi.org/10.3390/plants11091171
Morais MC, Cabral JA, Gonçalves B. Seasonal Variation in Selected Biochemical Traits in the Leaves of Co-Occurring Invasive and Native Plant Species under Mediterranean Conditions. Plants. 2022; 11(9):1171. https://doi.org/10.3390/plants11091171
Chicago/Turabian StyleMorais, Maria Cristina, João Alexandre Cabral, and Berta Gonçalves. 2022. "Seasonal Variation in Selected Biochemical Traits in the Leaves of Co-Occurring Invasive and Native Plant Species under Mediterranean Conditions" Plants 11, no. 9: 1171. https://doi.org/10.3390/plants11091171
APA StyleMorais, M. C., Cabral, J. A., & Gonçalves, B. (2022). Seasonal Variation in Selected Biochemical Traits in the Leaves of Co-Occurring Invasive and Native Plant Species under Mediterranean Conditions. Plants, 11(9), 1171. https://doi.org/10.3390/plants11091171