
Exploration of the Genetic Diversity of Solina Wheat and Its Implication for Grain Quality

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Grain Samples
2.2. Genotyping
2.3. Solina Dataset Pre-Processing and Filtering
2.4. Public CIMMYT Dataset Importing, Pre-Processing, and Filtering
2.5. Commercial Quality
2.6. Statistical Analysis
3. Results
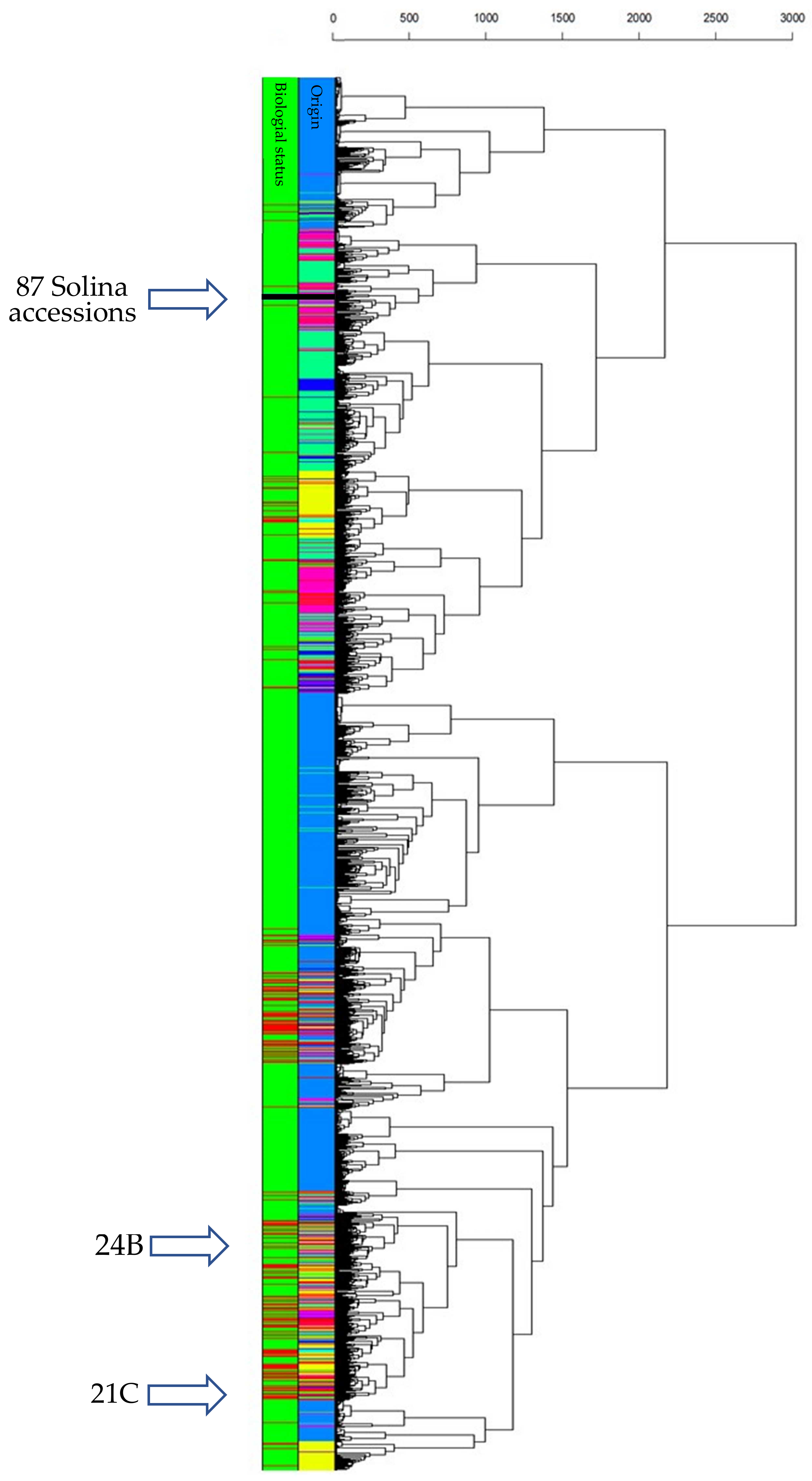
3.1. Solina Datasets: DArTseq Marker Characteristics
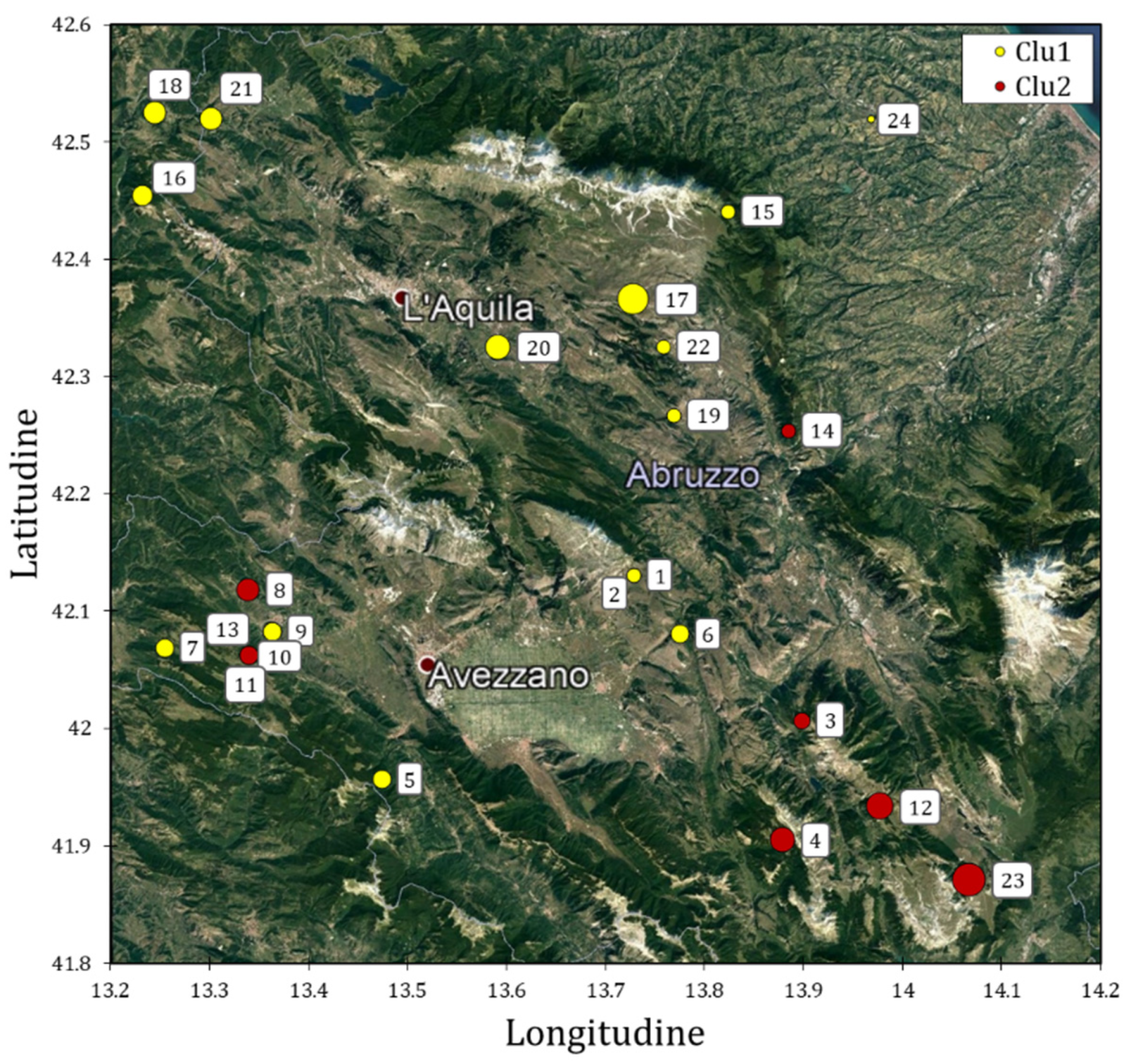
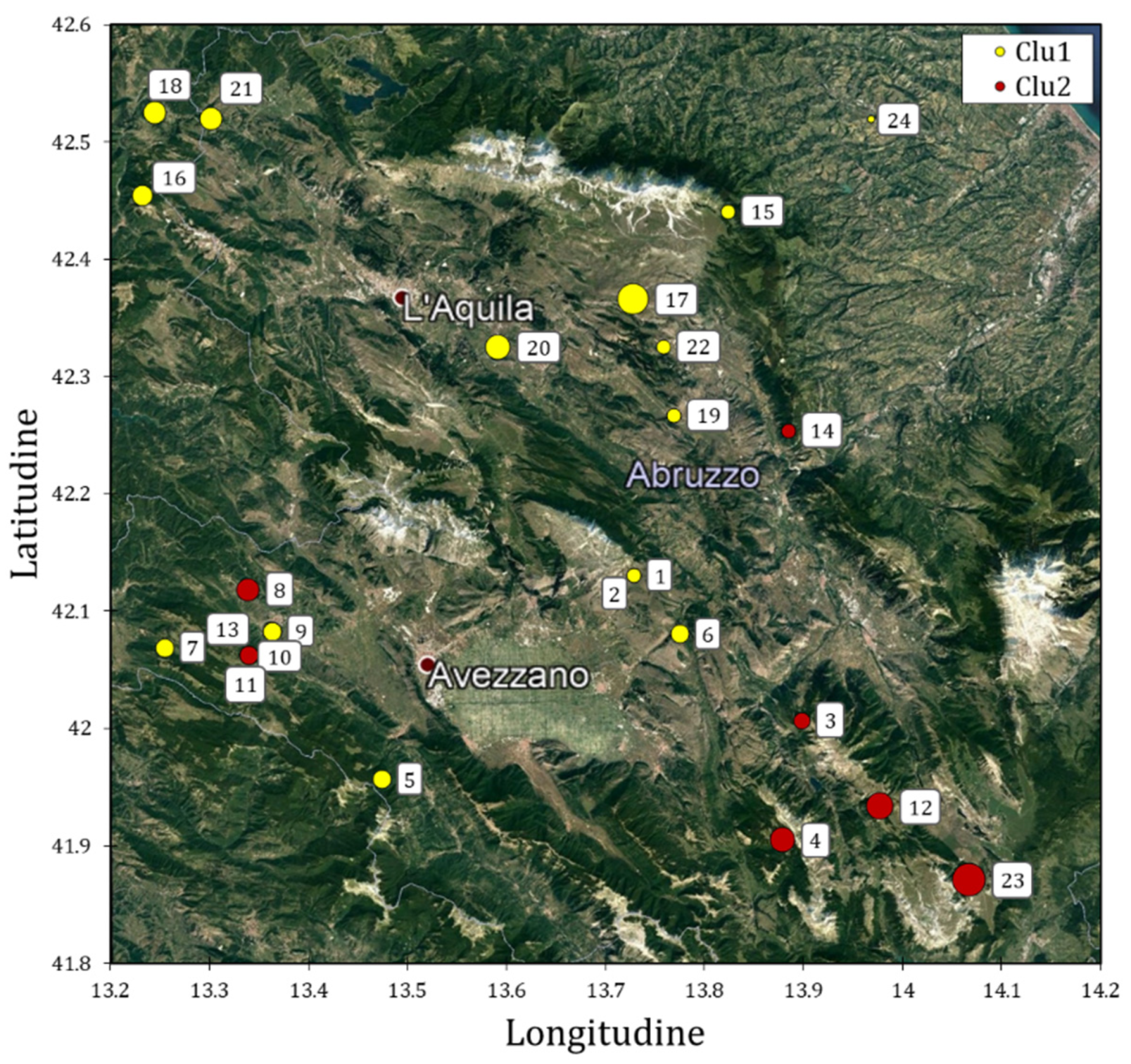
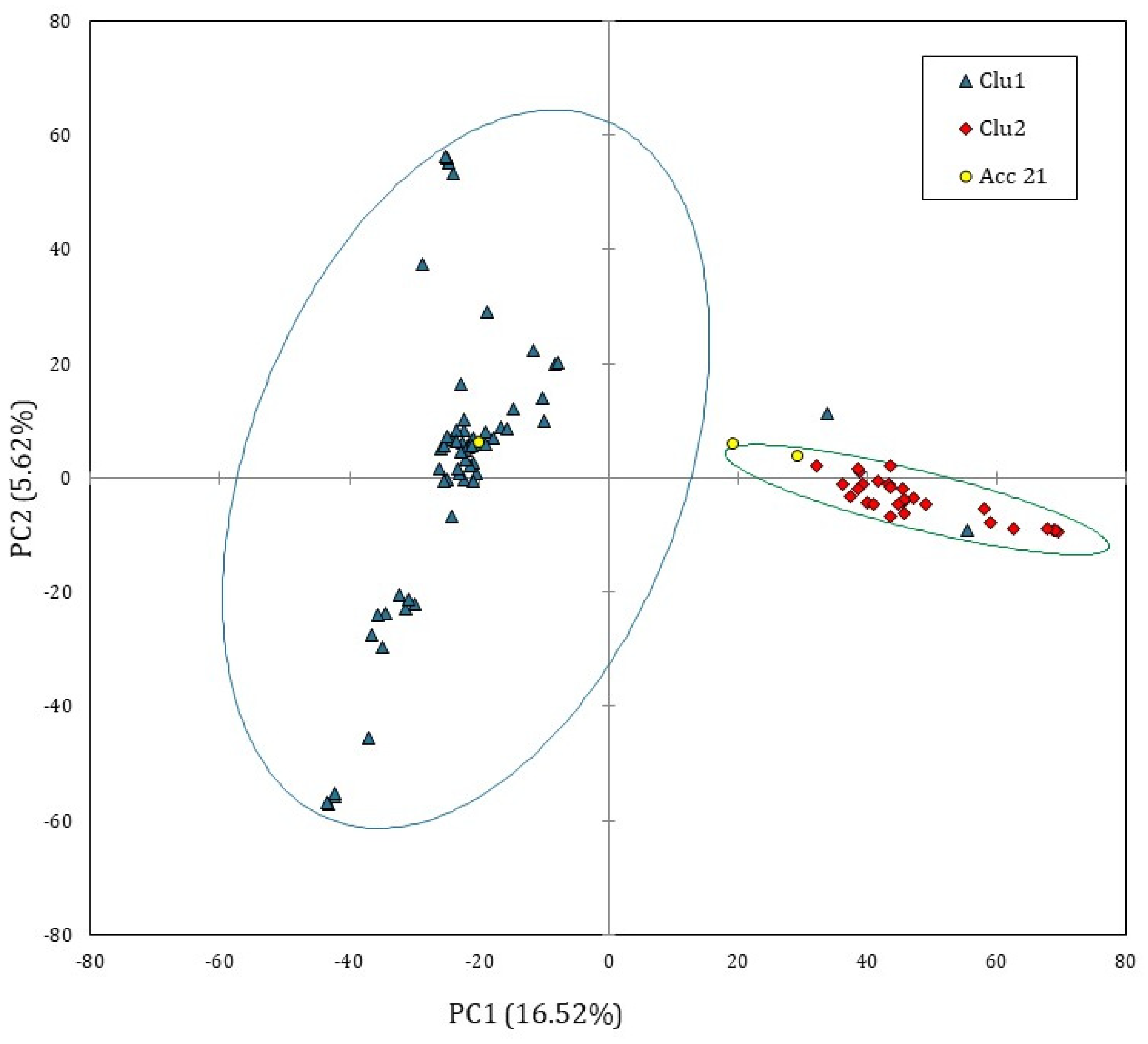
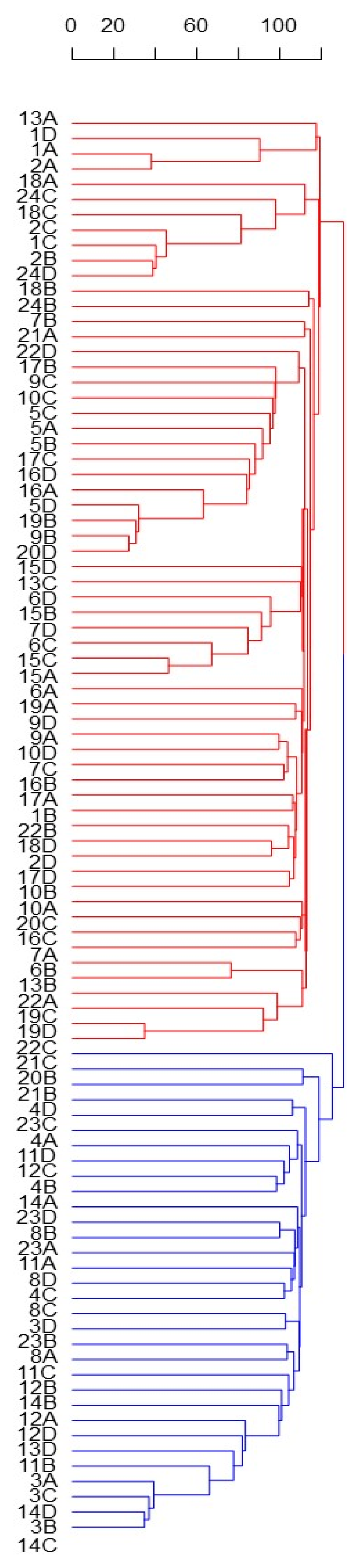
3.2. Solina Population Structure
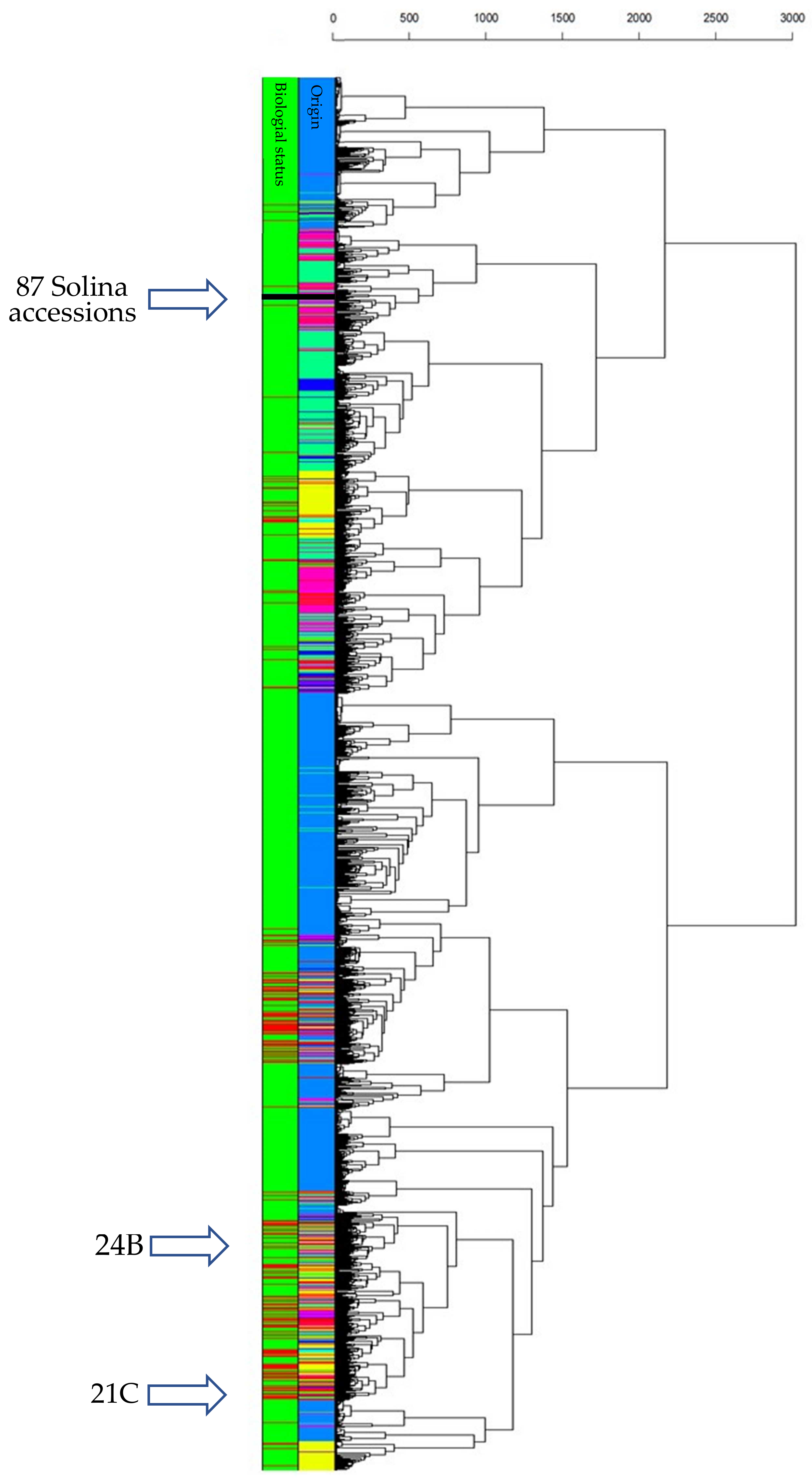
3.3. CIMMYT/Solina Dataset: DArTseq Marker and Population Structure
3.4. Commercial Quality Parameters
4. Discussion
4.1. Population Structure Analysis
4.2. Commercial Quality Parameters
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Manzi, A. Cereali. In Origine e Storia Delle Piante Coltivate in Abruzzo; Casa Editrice Rocco Carabba: Lanciano, Italy, 2006; pp. 23–42. [Google Scholar]
- Porfiri, O.; Silveri, D. La “Solina” e altre varietà locali di cereali ancora coltivate in Abruzzo: I risultati di una campagna di collezione e caratterizzazione promossa dall’ARSSA. In Atti VI Convegno Nazionale Biodiversità; Tecnomack: Bari, Italy, 2004; pp. 843–852. [Google Scholar]
- Khan, A.R.; Goldringer, I.; Thomas, M. Management Practices and Breeding History of Varieties Strongly Determine the Fine Genetic Structure of Crop Populations: A Case Study Based on European Wheat Populations. Sustainability 2020, 12, 613. [Google Scholar] [CrossRef] [Green Version]
- Dawson, J.C.; Serpolay, E.; Giuliano, S.; Schermann, N.; Galic, N.; Chable, V.; Goldringer, I. Multi-trait evolution of farmer varieties of bread wheat after cultivation in contrasting organic farming systems in Europe. Genetica 2012, 140, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Piergiovanni, A.R. Evaluation of genetic variation and grain quality of old bread wheat varieties introduced in north-western Italian environments. Genet. Resour. Crop Evol. 2013, 60, 325–333. [Google Scholar] [CrossRef]
- Porfiri, O.; Torricelli, R.; Silveri, D.D.; Papa, R.; Barcaccia, G.; Negri, V. The Triticeae genetic resources of central Italy: Collection, evaluation and conservation. Hereditas 2004, 135, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Osman, A.; Chable, V. Inventory of initiatives on seeds of landraces in Europe. J. Agric. Environ. Ternational Dev. 2009, 103, 95–130. [Google Scholar]
- Serpolay-Besson, E.; Dawson, J.C.; Chable, V.; Van Bueren, E.L.; Osman, A.; Pino, S.; Silveri, D.; Goldringer, I. Diversity of different farmer and modern wheat varieties cultivated in contrasting organic farming conditions in western Europe and implications for European seed and variety legislation. Org. Agric. 2011, 1, 127–145. [Google Scholar] [CrossRef]
- Dawson, J.C.; Serpolay, E.; Giuliano, S.; Schermann, N.; Galic, N.; Berthellot, J.-F.; Chesneau, V.; Ferte, H.; Mercier, F.; Osman, A.; et al. Phenotypic diversity and evolution of farmer varieties of bread wheat on organic farms in Europe. Genet. Resour. Crop Evol. 2012, 60, 145–163. [Google Scholar] [CrossRef]
- Durazzo, A.; Casale, G.; Melini, V.; Maiani, G.; Acquistucci, R. Total polyphenol content and antioxidant properties of Solina (Triticum aestivum L.) and derivatives thereof. Ital. J. Food Sci. 2016, 28, 221. [Google Scholar]
- Tavoletti, S.; Foligni, R.; Mozzon, M.; Pasquini, M. Comparison between fatty acid profiles of old and modern varieties of T. turgidum and T. aestivum: A case study in central Italy. J. Cereal Sci. 2018, 82, 198–205. [Google Scholar] [CrossRef]
- Raggi, L.; Caproni, L.; Negri, V. Landrace added value and accessibility in Europe: What a collection of case studies tells us. Biodivers. Conserv. 2021, 30, 1031–1048. [Google Scholar] [CrossRef]
- Bonvicini, M. Genealogical Selection of the Wheat "Solina"; Istituto di Allevamento Vegetale per la Cerealicoltura: Bologna, Italy, 1936. [Google Scholar]
- Perrino, P.; Laghetti, G.; Hammer, K. Collection of plant genetic resources in Italy, 1987. Genet. Resour. Crop Evol. 1988, 36, 377–390. [Google Scholar] [CrossRef]
- Salles, J.-M.; Teillard, F.; Tichit, M.; Zanella, M. Land sparing versus land sharing: An economist’s perspective. Reg. Environ. Chang. 2017, 17, 1455–1465. [Google Scholar] [CrossRef] [Green Version]
- Boukid, F.; Folloni, S.; Sforza, S.; Vittadini, E.; Prandi, B. Current trends in ancient grains-based foodstuffs: Insights into nutritional aspects and technological applications. Compr. Rev. Food Sci. Food Saf. 2018, 17, 123–136. [Google Scholar] [CrossRef] [Green Version]
- Negri, V.; Maxted, N.; Veteläinen, M. European Landrace Conservation: An Introduction. In European Landraces: On Farm Conservation, Management and Use; Biodiversity Technical Bulletin no. 15; Veteläinen, M., Negri, V., Maxted, N., Eds.; European Cooperative Programme for Plant Genetic Resources: Rome, Italy, 2009. [Google Scholar]
- Marone, D.; Russo, M.; Mores, A.; Ficco, D.; Laidò, G.; Mastrangelo, A.; Borrelli, G. Importance of Landraces in Cereal Breeding for Stress Tolerance. Plants 2021, 10, 1267. [Google Scholar] [CrossRef] [PubMed]
- Akbari, M.; Wenzl, P.; Caig, V.; Carling, J.; Xia, L.; Yang, S.; Uszynski, G.; Mohler, V.; Lehmensiek, A.; Kuchel, H.; et al. Diversity arrays technology (DArT) for high-throughput profiling of the hexaploid wheat genome. Theor. Appl. Genet. 2006, 113, 1409–1420. [Google Scholar] [CrossRef]
- Baloch, F.S.; Alsaleh, A.; Shahid, M.Q.; Çiftçi, V.; de Miera, L.E.S.; Aasim, M.; Nadeem, M.A.; Aktaş, H.; Özkan, H.; Hatipoğlu, R. A Whole Genome DArTseq and SNP Analysis for Genetic Diversity Assessment in Durum Wheat from Central Fertile Crescent. PLoS ONE 2017, 12, e0167821. [Google Scholar] [CrossRef] [Green Version]
- Edet, O.U.; Gorafi, Y.S.; Nasuda, S.; Tsujimoto, H. DArTseq-based analysis of genomic relationships among species of tribe Triticeae. Sci. Rep. 2018, 8, 16397. [Google Scholar] [CrossRef]
- Pascual, L.; Ruiz, M.; López-Fernández, M.; Pérez-Peña, H.; Benavente, E.; Vázquez, J.F.; Sansaloni, C.; Giraldo, P. Genomic analysis of Spanish wheat landraces reveals their variability and potential for breeding. BMC Genom. 2020, 21, 122. [Google Scholar] [CrossRef] [Green Version]
- Mahboubi, M.; Mehrabi, R.; Naji, A.M.; Talebi, R. Whole-genome diversity, population structure and linkage disequilibrium analysis of globally diverse wheat genotypes using genotyping-by-sequencing DArTseq platform. 3 Biotech 2020, 10, 48. [Google Scholar] [CrossRef]
- Tehseen, M.M.; Istipliler, D.; Kehel, Z.; Sansaloni, C.P.; da Silva Lopes, M.; Kurtulus, E. Genetic diversity and population structure analysis of Triticum aestivum L. Landrace panel from Afghanistan. Genes 2021, 12, 340. [Google Scholar] [CrossRef]
- Sansaloni, C.; Franco, J.; Santos, B.; Percival-Alwyn, L.; Singh, S.; Petroli, C.; Campos, J.; Dreher, K.; Payne, T.; Marshall, D.; et al. Diversity analysis of 80,000 wheat accessions reveals consequences and opportunities of selection footprints. Nat. Commun. 2020, 11, 4572. [Google Scholar] [CrossRef] [PubMed]
- Tumino, G.; Voorrips, R.E.; Rizza, F.; Badeck, F.-W.; Morcia, C.; Ghizzoni, R.; Germeier, C.U.; Paulo, M.J.C.; Terzi, V.; Smulders, M.J. Population structure and genome-wide association analysis for frost tolerance in oat using continuous SNP array signal intensity ratios. Theor. Appl. Genet. 2016, 129, 1711–1724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, S.; Dubcovsky, J.; Dvorak, J.; Luo, M.-C.; Baenziger, S.P.; Matnyazov, R.; Clark, D.R.; Talbert, L.; Anderson, J.; Dreisigacker, S.; et al. Population- and genome-specific patterns of linkage disequilibrium and SNP variation in spring and winter wheat (Triticum aestivum L.). BMC Genom. 2010, 11, 727. [Google Scholar] [CrossRef] [Green Version]
- Chapin, F.S.; Autumn, K.; Pugnaire, F.I. Evolution of Suites of Traits in Response to Environmental Stress. Am. Nat. 1993, 142, S78–S92. [Google Scholar] [CrossRef]
- Matsuoka, Y. Evolution of polyploidy Triticum wheats under cultivation: The role of domestication, natural hybridization and alloplypolid speciation in their diversification. Plant Cell Physiol. 2011, 52, 750–764. [Google Scholar] [CrossRef] [Green Version]
- Casañas, F.; Simó, J.; Casals, J.; Prohens, J. Toward an evolved concept of landrace. Front. Plant Sci. 2017, 8, 145. [Google Scholar] [CrossRef] [Green Version]
- Zeven, A.C. Landraces: A review of definitions and classifications. Euphytica 1998, 104, 127–139. [Google Scholar] [CrossRef]
- Williams, R.M.; O’Brien, L.; Eagles, H.A.; Solah, V.; Jayasena, V. The influences of genotype, environment, and genotype×environment interaction on wheat quality. Aust. J. Agric. Res. 2008, 59, 95–111. [Google Scholar] [CrossRef]
- Ashraf, M. Stress-Induced Changes in Wheat Grain Composition and Quality. Crit. Rev. Food Sci. Nutr. 2014, 54, 1576–1583. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effect | Type | TKW | TW | Protein % |
|---|---|---|---|---|
| Clu1/Clu2 | Fixed | 0.018 | 0.017 | ns |
| Farm (Clu1/Clu2) | Random | <0.001 | <0.001 | <0.001 |
| Effect | Type | TKW | TW | Protein % | Yield |
|---|---|---|---|---|---|
| Plot | Random | ns | ns | ns | ns |
| Year | Fixed | ns | <0.001 | ns | 0.011 |
| Farm | Fixed | <0.001 | ns | ns | <0.001 |
| Clu1/Clu2 | Fixed | 0.003 | 0.044 | 0.003 | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Flaviis, R.; Tumino, G.; Terzi, V.; Morcia, C.; Santarelli, V.; Sacchetti, G.; Mastrocola, D. Exploration of the Genetic Diversity of Solina Wheat and Its Implication for Grain Quality. Plants 2022, 11, 1170. https://doi.org/10.3390/plants11091170
De Flaviis R, Tumino G, Terzi V, Morcia C, Santarelli V, Sacchetti G, Mastrocola D. Exploration of the Genetic Diversity of Solina Wheat and Its Implication for Grain Quality. Plants. 2022; 11(9):1170. https://doi.org/10.3390/plants11091170
Chicago/Turabian StyleDe Flaviis, Riccardo, Giorgio Tumino, Valeria Terzi, Caterina Morcia, Veronica Santarelli, Giampiero Sacchetti, and Dino Mastrocola. 2022. "Exploration of the Genetic Diversity of Solina Wheat and Its Implication for Grain Quality" Plants 11, no. 9: 1170. https://doi.org/10.3390/plants11091170
APA StyleDe Flaviis, R., Tumino, G., Terzi, V., Morcia, C., Santarelli, V., Sacchetti, G., & Mastrocola, D. (2022). Exploration of the Genetic Diversity of Solina Wheat and Its Implication for Grain Quality. Plants, 11(9), 1170. https://doi.org/10.3390/plants11091170