
Chemical and Molecular Characterization of Wound-Induced Suberization in Poplar (Populus alba × P. tremula) Stem Bark


Abstract
:1. Introduction
2. Results
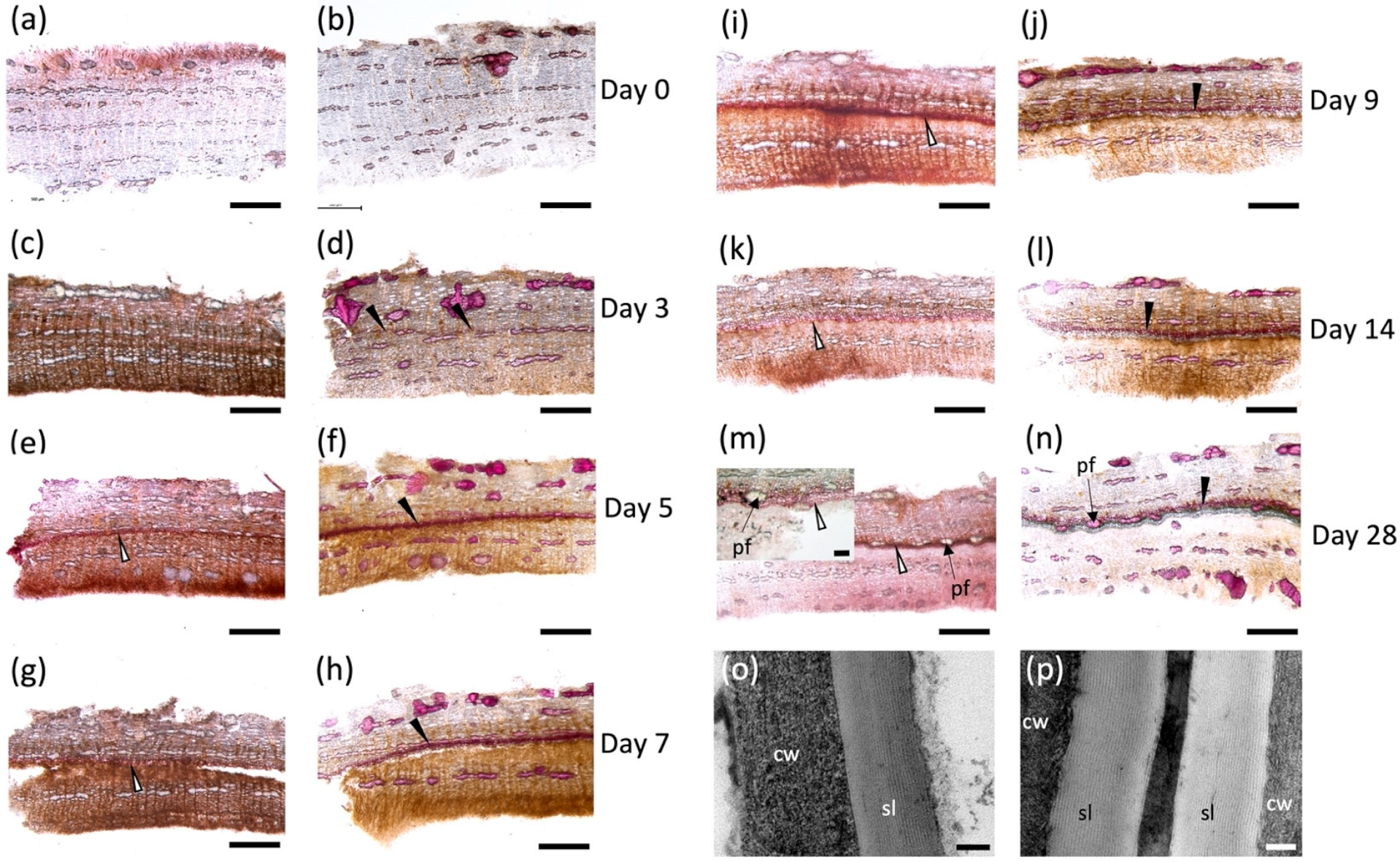
2.1. Histochemistry of the Wound-Healing Suberization Process in Poplar Bark
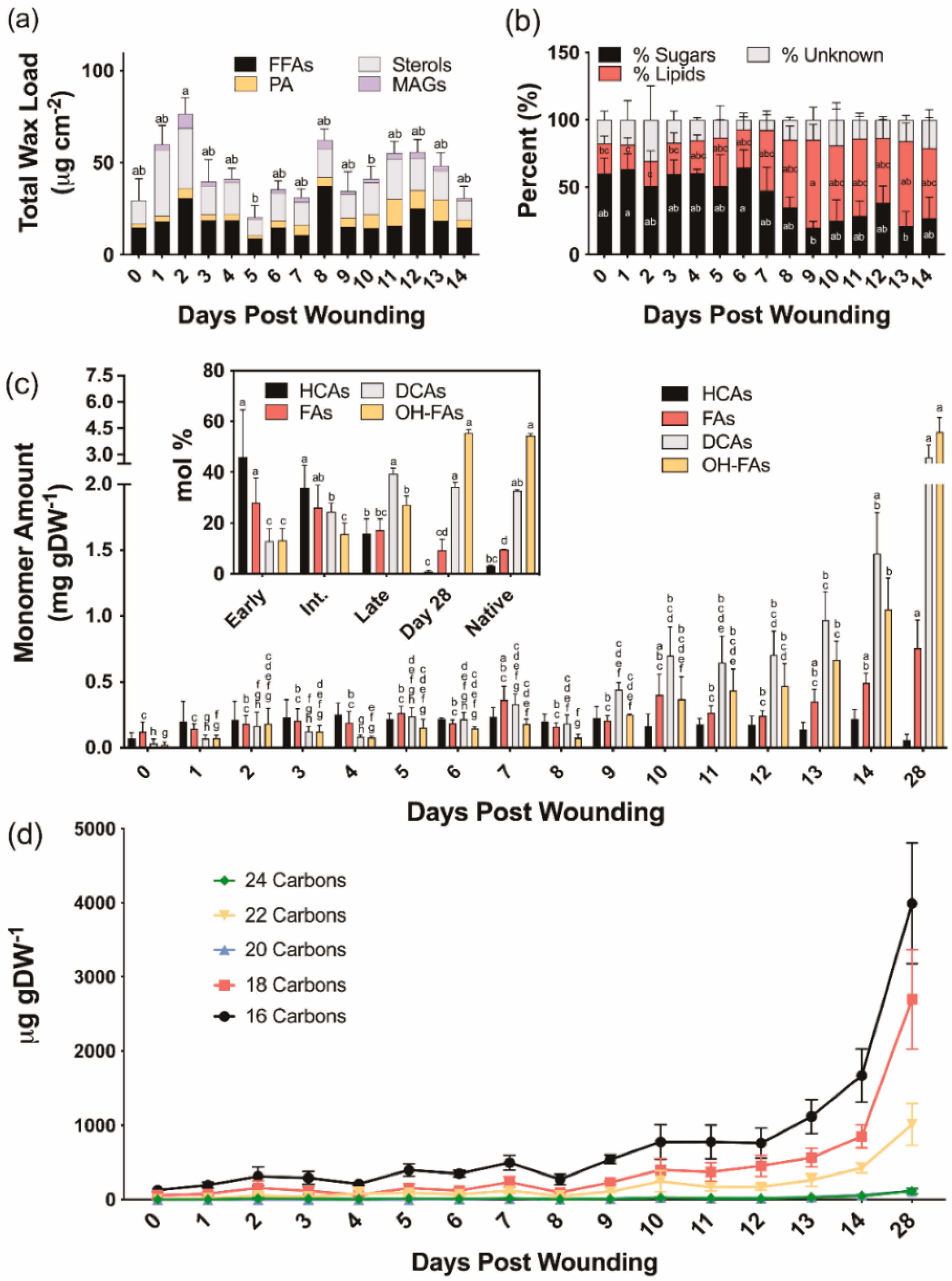
2.2. Time Course of Suberin Deposition during Wound Periderm Formation
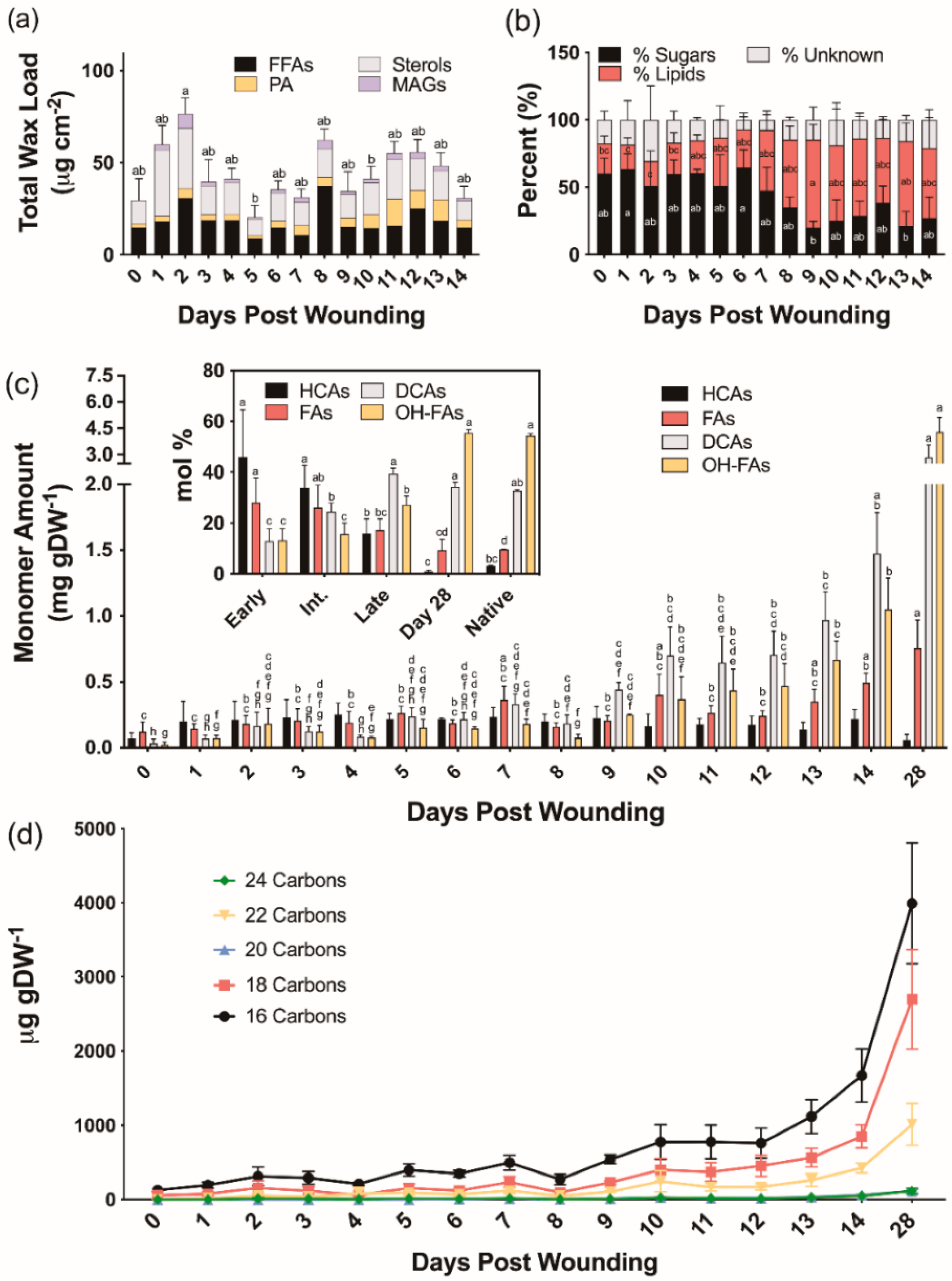
2.2.1. Suberin-Associated Waxes
2.2.2. Suberin Aliphatic Polyester Monomers
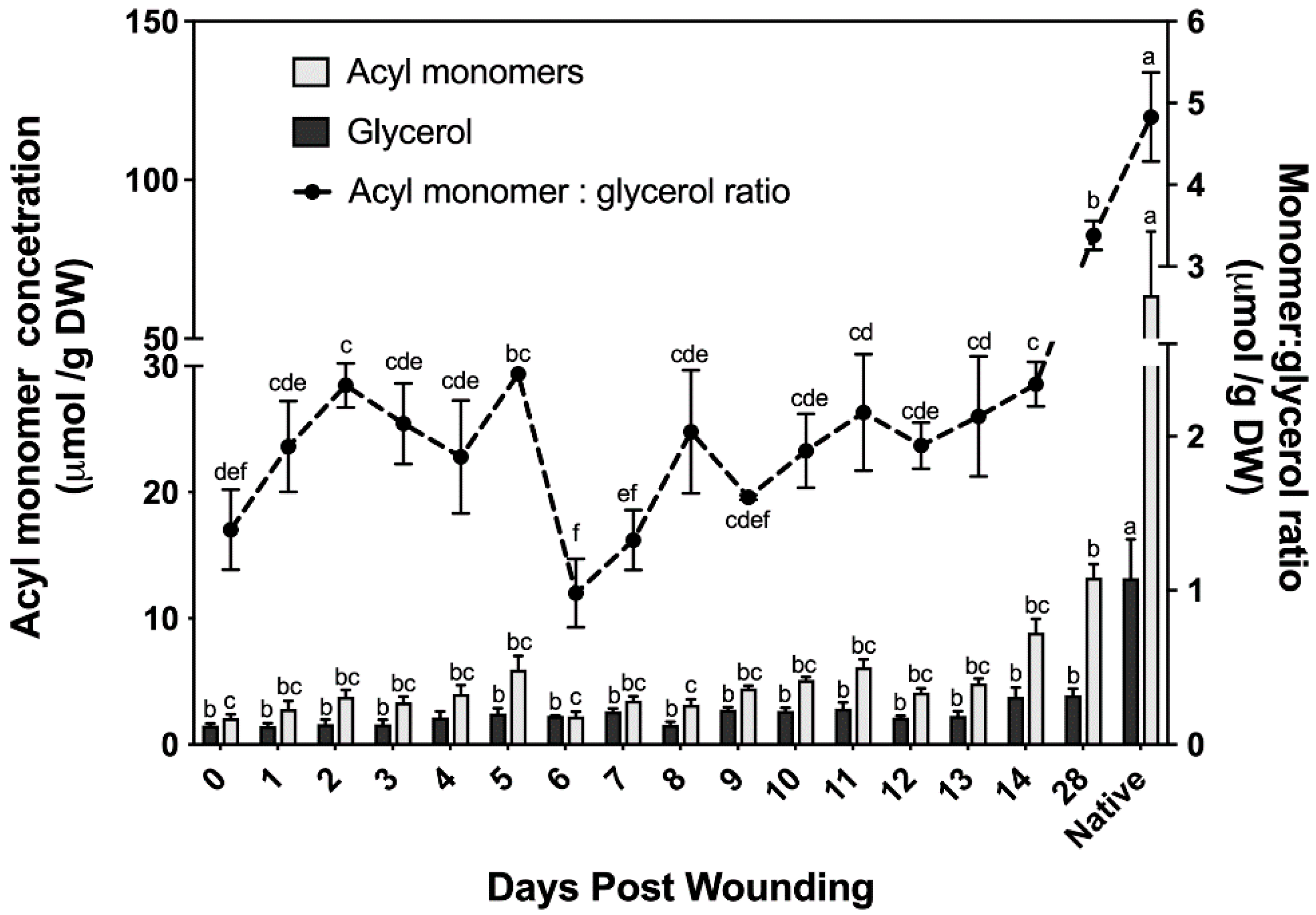
2.2.3. Suberin Glycerol
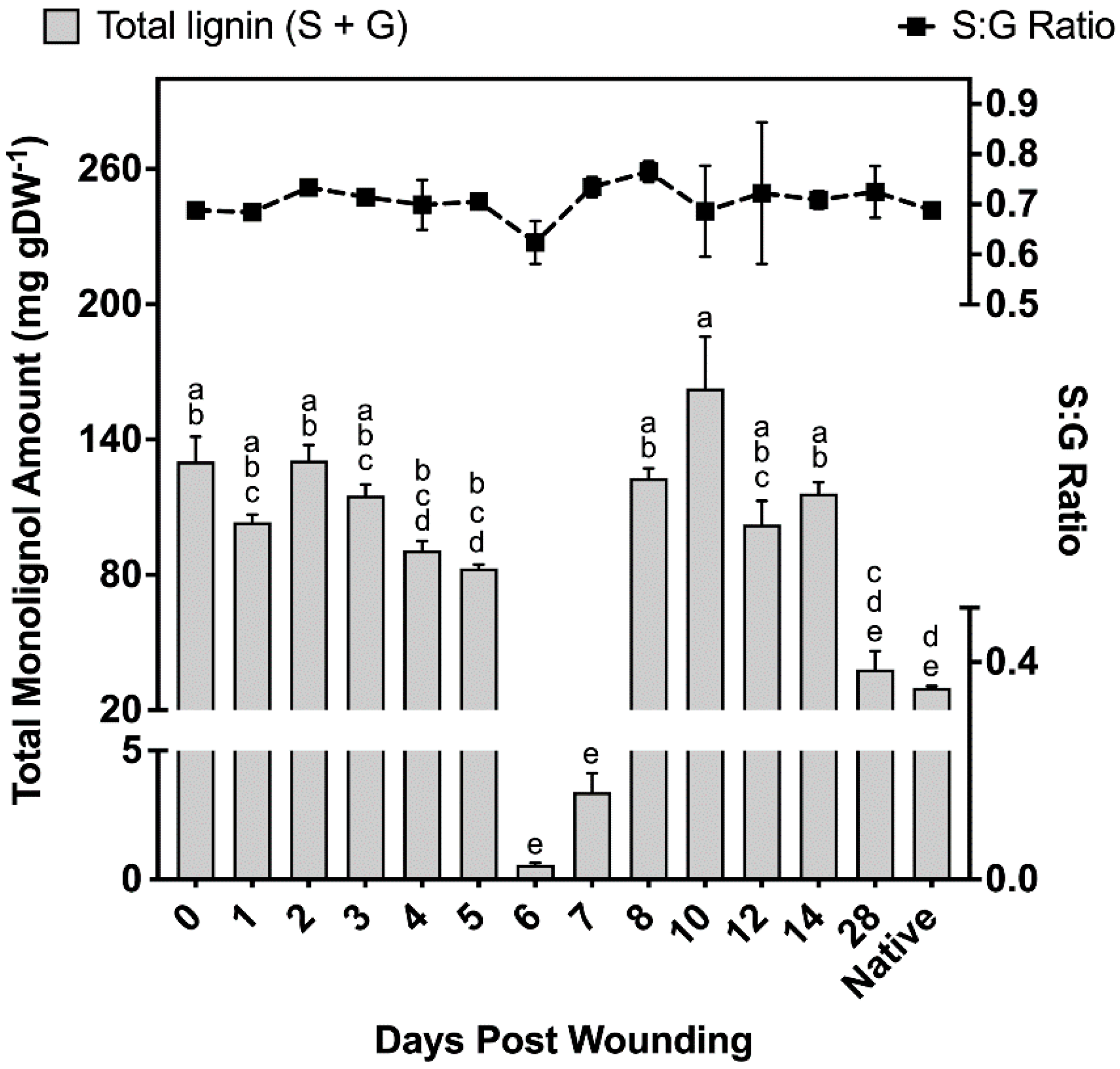
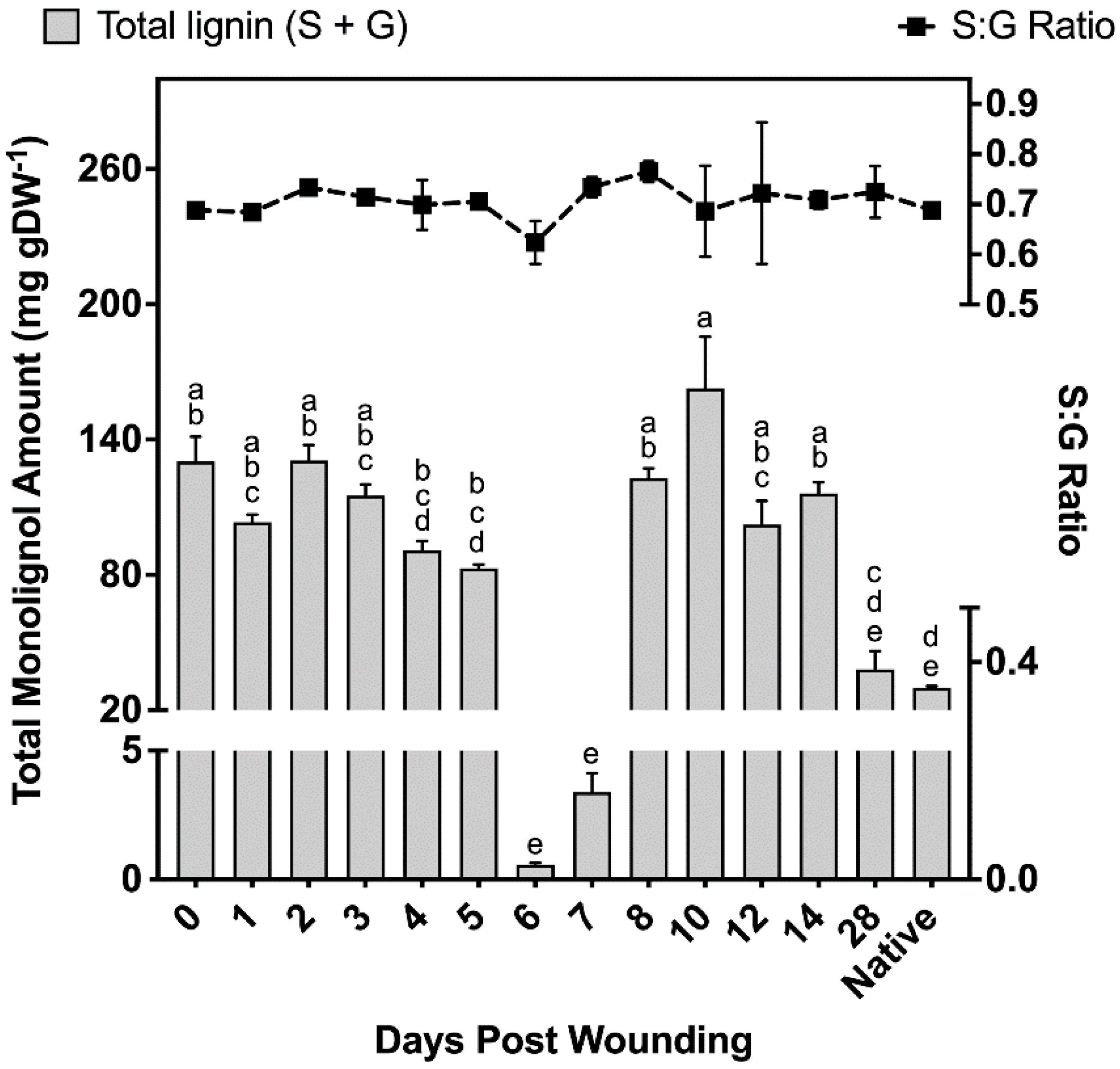
2.2.4. Polyphenolics Analysis
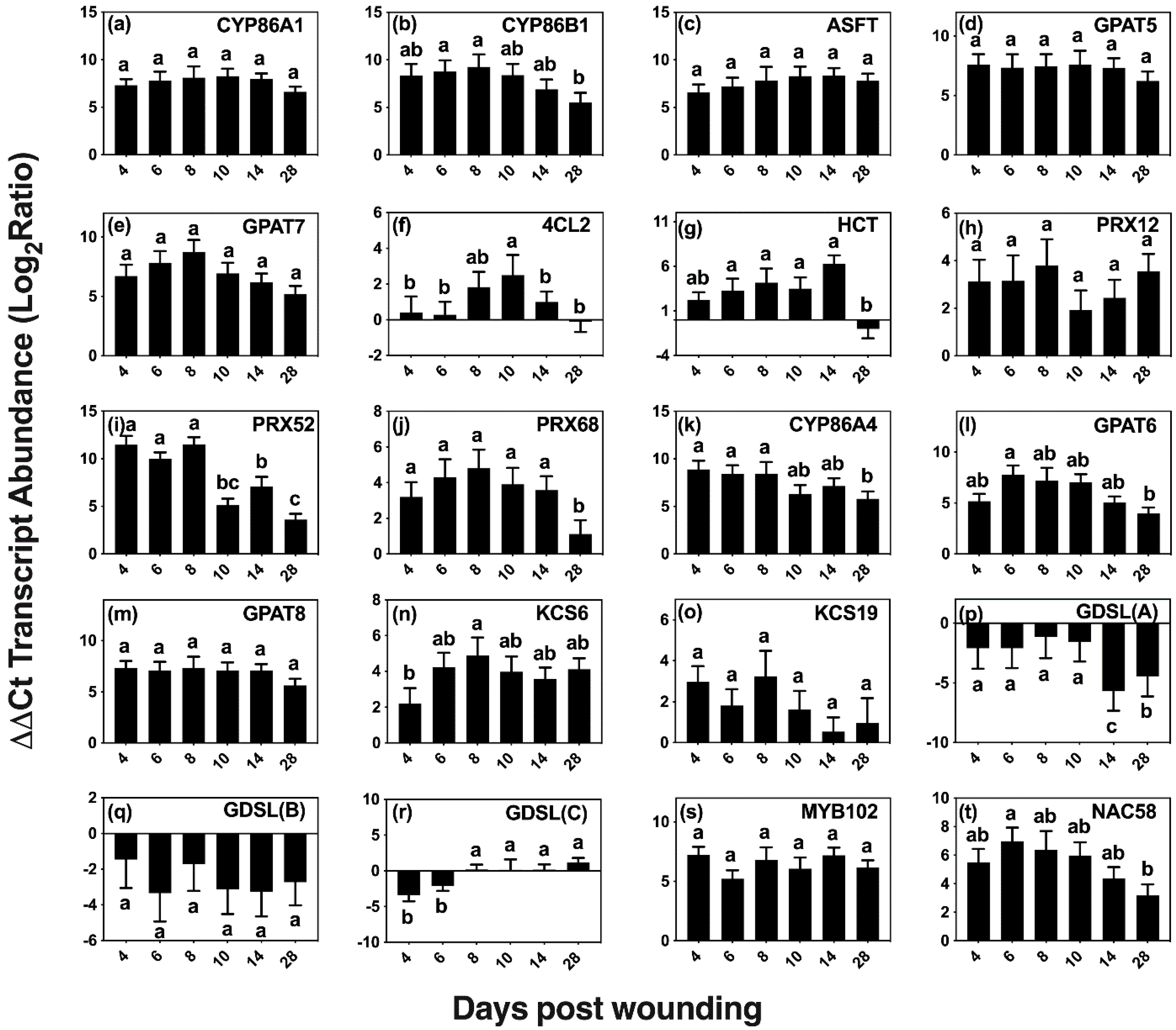
2.3. Time Course of Expression of Predicted Genes of Suberin Biosynthesis
3. Discussion
3.1. Microscopic Analysis of Bark Wound-Healing Suberization
3.2. Polyphenolic Suberin Biosynthesis and Composition
3.3. Aliphatic Suberin Biosynthesis and Composition
4. Materials and Methods
4.1. Plant Materials
4.2. Bark Wounding
4.3. RNA Extraction and cDNA Synthesis
4.4. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.5. Total Lipids and Wax Analysis
4.6. Lipid Polyester Analysis
4.7. Glycerol Quantification
4.8. Glycerol Determination Using a Commercial Kit
4.9. Polyphenolic Analysis: Modified Microscale Green Solvent Thioacidolysis
4.10. Microscopy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ginzberg, I. Wound-peridem formation. In Induced Plant Resistance to Herbivory; Schalle, A., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 131–146. [Google Scholar]
- Jin, L.; Cai, Q.; Huang, W.; Dastmalchi, K.; Rigau, J.; Molinas, M.; Figueras, M.; Serra, O.; Stark, R.E. Potato native and wound periderms are differently affected by down-regulation of FHT, a suberin feruloyl transferase. Phytochemistry 2018, 147, 30–48. [Google Scholar] [CrossRef] [PubMed]
- Bernards, M.A. Demystifying suberin. Can. J. Bot. 2002, 80, 227–240. [Google Scholar] [CrossRef]
- Graça, J. Suberin: A biopolyester of plants’ skin. Macromol. Biosci. 2007, 7, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Graça, J. Suberin: The biopolyester at the frontier of plants. Front. Chem. 2015, 3, 62. [Google Scholar] [CrossRef]
- Woolfson, K.N.; Esfandiari, M.; Bernards, M.A. Suberin biosynthesis, assembly, and regulation. Plants 2022, 11, 555. [Google Scholar] [CrossRef]
- Pollard, M.; Beisson, F.; Li, Y.; Ohlrogge, J.B. Building lipid barriers: Biosynthesis of cutin and suberin. Trends Plant Sci. 2008, 13, 236–246. [Google Scholar] [CrossRef]
- Kolattukudy, P.E. Biochemistry and function of cutin and suberin. Can. J. Bot. 1984, 62, 2918–2933. [Google Scholar] [CrossRef]
- Schreiber, L.; Franke, R.; Hartmann, K. Wax and suberin development of native and wound periderm of potato (Solanum Tuberosum L.) and its relation to peridermal transpiration. Planta 2005, 220, 520–530. [Google Scholar] [CrossRef]
- Lulai, E.C.; Neubauer, J.D. Wound-induced suberization genes are differentially expressed, spatially and temporally, during closing layer and wound periderm formation. Postharvest Biol. Technol. 2014, 90, 24–33. [Google Scholar] [CrossRef]
- Lulai, E.C.; Corsini, D.L. Differential Deposition of suberin phenolic and aliphatic domains and their roles in resistance to infection during potato tuber (Solanum tuberosum L.) wound-healing. Physiol. Mol. Plant Pathol. 1998, 53, 209–222. [Google Scholar] [CrossRef]
- Woolfson, K.N.; Haggitt, M.L.; Zhang, Y.; Kachura, A.; Bjelica, A.; Rincon, M.A.R.; Kaberi, K.M.; Bernards, M.A. Differential induction of polar and non-polar metabolism during wound-induced suberization in potato (Solanum tuberosum L.) tubers. Plant J. 2018, 93, 931–942. [Google Scholar] [CrossRef] [Green Version]
- Biggs, A.R. Anatomical and physiological responses of bark tissues to mechanical injury. In Defense Mechanisms of Woody Plants against Fungi; Blanchette, R.A., Biggs, R.A., Eds.; Springer Series in Wood Science; Springer: Berlin/Heidelberg, Germany, 1992; pp. 13–40. [Google Scholar]
- Biggs, A.R. Detection of impervious tissue in tree bark with selective histochemistry and fluorescence microscopy. Stain Technol. 1985, 60, 299–304. [Google Scholar] [CrossRef]
- Biggs, A.R. Phellogen regeneration in injured peach tree bark. Ann. Bot. 1996, 57, 463–470. [Google Scholar] [CrossRef]
- Woodward, S.; Pocock, S. Formation of the ligno-suberized barrier zone and wound periderm in four species of European broad-leaved trees. Eur. J. For. Res. 1996, 26, 97–105. [Google Scholar] [CrossRef]
- Esau, K. Anatomy of Seed Plants, 2nd ed.; John Wiley & Sons: New York, NY, USA, 1977. [Google Scholar]
- Serra, O.; Mähönen, A.P.; Hetherington, A.J.; Ragni, L. The making of plant armor: The periderm. Annu. Rev. Plant Biol. 2022, 73, 1–28. [Google Scholar] [CrossRef]
- Biggs, A.R. Intracellular suberin: Occurrence and detection in tree bark. IAWA J. 1984, 5, 243–248. [Google Scholar] [CrossRef]
- Biggs, A.R.; Stobbs, L.W. Fine Structure of the suberized cell walls in the boundary zone and necrophylactic periderm in wounded peach bark. Can. J. Bot. 1986, 64, 1606–1610. [Google Scholar] [CrossRef]
- Mullick, D.B. A new tissue essential to necrophylactic periderm formation in the bark of four conifers. Can. J. Bot. 1975, 53, 2443–2457. [Google Scholar] [CrossRef]
- Biggs, A.R.; Merrill, W.; Davis, D.D. Discussion: Response of bark tissues to injury and infection. Can. J. For. Res. 1984, 14, 351–356. [Google Scholar] [CrossRef]
- Rittinger, P.A.; Biggs, A.R.; Peirson, D.R. Histochemistry of lignin and suberin deposition in boundary-layers formed after wounding in various plant-species and organs. Can. J. Bot. 1987, 65, 1886–1892. [Google Scholar] [CrossRef]
- Lapierre, C.; Monties, B.; Rolando, C.; de Chirale, L. Thioacidolysis of lignin: Comparison with acidolysis. J. Wood Chem. Technol. 1985, 5, 277–292. [Google Scholar] [CrossRef]
- Biggs, A.R. Prediction of lignin and suberin deposition in boundary zone tissue of wounded tree bark using accumulated degree-days. J. Am. Soc. Hort. Sci. 1986, 111, 757–760. [Google Scholar]
- Tuskan, G.A.; DiFazio, S.; Jansson, S.; Bohlmann, J.; Grigoriev, I.; Hellsten, U.; Putnam, N.; Ralph, S.; Rombauts, S.; Salamov, A.; et al. The genome of black cottonwood, Populus Trichocarpa. Science 2006, 313, 1596–1604. [Google Scholar] [CrossRef] [Green Version]
- Ellis, B.; Jansson, S.; Strauss, S.H.; Tuskan, G.A. Why and how Populus became a “model tree”. In Genetics and Genomics of Populus; Jansson, S., Bhalerao, R., Groover, A., Eds.; Springer: New York, NY, USA, 2010; pp. 15–38. [Google Scholar]
- Slavov, G.T.; Zhelev, P. Salient biological features, systematics, and genetic variation of Populus. In Genetics and Genomics of Populus; Jansson, S., Bhalerao, R., Groover, A., Eds.; Plant Genetics and Genomics: Crops and Models; Springer: New York, NY, USA, 2010; Volume 8, pp. 15–38. [Google Scholar]
- Brundrett, M.C.; Kendrick, B.; Peterson, C.A. Efficient lipid staining in plant-material with sudan red-7b or fluoral yellow-088 in polyethylene glycol-glycerol. Biotech. Histochem. 1991, 66, 111–116. [Google Scholar] [CrossRef]
- Rains, M.K.; de Silva, N.D.G.; Molina, I. Reconstructing the suberin pathway in poplar by chemical and transcriptomic analysis of bark tissues. Tree Physiol. 2018, 38, 340–361. [Google Scholar] [CrossRef]
- Yang, W.; Pollard, M.; Li-Beisson, Y.; Ohlrogge, J. Quantitative analysis of glycerol in dicarboxylic acid-rich cutins provides insights into Arabidopsis cutin structure. Phytochemistry 2016, 130, 159–169. [Google Scholar] [CrossRef] [Green Version]
- Zeier, J.; Schreiber, L. Chemical composition of hypodermal and endodermal cell walls and xylem vessels isolated from Clivia Miniata (Identification of the biopolymers lignin and suberin). Plant Physiol. 1997, 113, 1223–1231. [Google Scholar] [CrossRef] [Green Version]
- Lapierre, C.; Pollet, B.; Négrel, J. The phenolic domain of potato suberin: Structural comparison with lignins. Phytochemistry 1996, 42, 949–953. [Google Scholar] [CrossRef]
- Capello, C.; Fischer, U.; Hungerbühler, K. What is a green solvent? A comprehensive framework for the environmental assessment of solvents. Green Chem. 2007, 9, 927–934. [Google Scholar] [CrossRef]
- Derosa, C.T.; Wilbur, S.; Holler, J.; Richter, P.; Stevens, Y.-W. Health evaluation of 1,4-dioxane. Toxicol. Ind. Health 1996, 12, 1–43. [Google Scholar] [CrossRef]
- Foster, C.E.; Martin, T.M.; Pauly, M. Comprehensive compositional analysis of plant cell walls (lignocellulosic biomass) Part I: Lignin. J. Vis. Exp. 2010, 37, 1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, A.R.; Mansfield, S.D. Rapid analysis of poplar lignin monomer composition by a streamlined thioacidolysis procedure and near-infrared reflectance-based prediction modeling. Plant J. 2009, 58, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Yamagiwa, N.; Torisawa, Y. Cyclopentyl methyl ether as a new and alternative process solvent. Org. Process Res. Dev. 2007, 11, 251–258. [Google Scholar] [CrossRef]
- Ehlting, J.; Büttner, D.; Wang, Q.; Douglas, C.J.; Somssich, I.E.; Kombrink, E. Three 4-coumarate:coenzyme A ligases in Arabidopsis thaliana represent two evolutionarily divergent classes in angiosperms. Plant J. 1999, 19, 9–20. [Google Scholar] [CrossRef]
- Hoffmann, L.; Besseau, S.; Geoffroy, P.; Ritzenthaler, C.; Meyer, D.; Lapierre, C.; Pollet, B.; Legrand, M. Silencing of hydroxycinnamoyl-coenzyme a shikimate/quinate hydroxycinnamoyltransferase affects phenylpropanoid biosynthesis. Plant Cell 2004, 16, 1446–1465. [Google Scholar] [CrossRef] [Green Version]
- Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin biosynthesis and structure. Plant Physiol. 2010, 153, 895–905. [Google Scholar] [CrossRef] [Green Version]
- Barceló, A.R. Lignification in plant cell walls. Int. Rev. Cytol. 1997, 176, 87–132. [Google Scholar]
- Bernards, M.A.; Fleming, W.D.; Llewellyn, D.B.; Priefer, R.; Yang, X.; Sabatino, A.; Plourde, G.L. Biochemical characterization of the suberization-associated anionic peroxidase of potato. Plant Physiol. 1999, 121, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Bernards, M.A.; Summerhurst, D.K.; Razem, F.A. Oxidases, peroxidases and hydrogen peroxide: The suberin connection. Phytochem. Rev. 2004, 3, 113–126. [Google Scholar] [CrossRef]
- Bernards, M.A.; Razem, F.A. The poly(phenolic) domain of potato suberin: A non-lignin cell wall bio-polymer. Phytochemistry 2001, 57, 1115–1122. [Google Scholar] [CrossRef]
- Molina, I.; Li-Beisson, Y.; Beisson, F.; Ohlrogge, J.B.; Pollard, M. Identification of an Arabidopsis feruloyl-coenzyme A transferase required for suberin synthesis. Plant Physiol. 2009, 151, 1317–1328. [Google Scholar] [CrossRef] [Green Version]
- Gou, J.-Y.; Yu, X.-H.; Liu, C.-J. A Hydroxycinnamoyltransferase responsible for synthesizing suberin aromatics in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 18855–18860. [Google Scholar] [CrossRef] [Green Version]
- Höfer, R.; Briesen, I.; Beck, M.; Pinot, F.; Schreiber, L.; Franke, R. The Arabidopsis cytochrome P450 CYP86A1 encodes a fatty acid ω-hydroxylase involved in suberin monomer biosynthesis. J. Exp. Bot. 2008, 59, 2347–2360. [Google Scholar] [CrossRef] [Green Version]
- Compagnon, V.; Diehl, P.; Benveniste, I.; Meyer, D.; Schaller, H.; Schreiber, L.; Franke, R.; Pinot, F. CYP86B1 is required for very long chain-hydroxyacid and dicarboxylic acid synthesis in root and seed suberin polyester. Plant Physiol. 2009, 150, 1831–1843. [Google Scholar] [CrossRef] [Green Version]
- Beisson, F.; Li, Y.; Bonaventure, G.; Pollard, M.; Ohlrogge, J.B. The acyltransferase GPAT5 is required for the synthesis of suberin in seed coat and root of Arabidopsis. Plant Cell 2007, 19, 351–368. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Simpson, J.P.; Li-Beisson, Y.; Beisson, F.; Pollard, M.; Ohlrogge, J.B. A land-plant-specific glycerol-3-phosphate acyltransferase family in Arabidopsis: Substrate specificity, sn-2 preference, and evolution. Plant Physiol. 2012, 160, 638–652. [Google Scholar] [CrossRef] [Green Version]
- Li-Beisson, Y.; Pollard, M.; Sauveplane, V.; Pinot, F.; Ohlrogge, J.; Beisson, F. Nanoridges that characterize the surface morphology of flowers require the synthesis of cutin polyester. Proc. Natl. Acad. Sci. USA 2009, 106, 22008–22013. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Beisson, F.; Koo, A.J.K.; Molina, I.; Pollard, M.; Ohlrogge, J. Identification of acyltransferases required for cutin biosynthesis and production of cutin with suberin-like monomers. Proc. Natl. Acad. Sci. USA 2007, 104, 18339–18344. [Google Scholar] [CrossRef] [Green Version]
- Fiebig, A.; Mayfield, J.A.; Miley, N.L.; Chau, S.; Fischer, R.L.; Preuss, D. Alterations in CER6, a gene identical to CUT1, differentially affect long-chain lipid content on the surface of pollen and stems. Plant Cell 2000, 12, 2001–2008. [Google Scholar] [CrossRef] [Green Version]
- Yeats, T.H.; Huang, W.; Chatterjee, S.; Viart, H.M.; Clausen, M.H.; Stark, R.E.; Rose, J.K.C. Tomato CUTIN DEFICIENT 1 (CD1) and putative orthologs comprise an ancient family of cutin synthase-like (CUS) proteins that are conserved among land plants. Plant J. 2014, 77, 667–675. [Google Scholar] [CrossRef]
- Yeats, T.H.; Martin, L.B.B.; Viart, H.M.-F.; Isaacson, T.; He, Y.; Zhao, L.; Matas, A.J.; Buda, G.J.; Domozych, D.S.; Clausen, M.H.; et al. The identification of cutin synthase: Formation of the plant polyester cutin. Nat. Chem. Biol. 2012, 8, 609–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girard, A.-L.; Mounet, F.; Lemaire-Chamley, M.; Gaillard, C.; Elmorjani, K.; Vivancos, J.; Runavot, J.-L.; Quemener, B.; Petit, J.; Germain, V.; et al. Tomato GDSL1 is required for cutin deposition in the fruit cuticle. Plant Cell 2012, 24, 3119–3134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosma, D.K.; Murmu, J.; Razeq, F.M.; Santos, P.; Bourgault, R.; Molina, I.; Rowland, O. AtMYB41 activates ectopic suberin synthesis and assembly in multiple plant species and cell types. Plant J. 2014, 80, 216–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denekamp, M.; Smeekens, S.C. Integration of wounding and osmotic stress signals determines the expression of the AtMYB102 transcription factor gene. Plant Physiol. 2003, 132, 1415–1423. [Google Scholar] [CrossRef] [Green Version]
- Wahrenburg, Z.; Benesch, E.; Lowe, C.; Jimenez, J.; Vulavala, V.K.R.; Lü, S.; Hammerschmidt, R.; Douches, D.; Yim, W.C.; Santos, P.; et al. Transcriptional regulation of wound suberin deposition in potato cultivars with differential wound healing capacity. Plant J. 2021, 107, 77–99. [Google Scholar] [CrossRef]
- Nuruzzaman, M.; Sharoni, A.M.; Kikuchi, S. Roles of NAC transcription factors in the regulation of biotic and abiotic stress responses in plants. Front. Microbiol. 2013, 4, 248. [Google Scholar] [CrossRef] [Green Version]
- Verdaguer, R.; Soler, M.; Serra, O.; Garrote, A.; Fernández, S.; Company-Arumí, D.; Anticó, E.; Molinas, M.; Figueras, M. Silencing of the potato StNAC103 gene enhances the accumulation of suberin polyester and associated wax in tuber skin. J. Exp. Bot. 2016, 67, 5415–5427. [Google Scholar] [CrossRef] [Green Version]
- Li-Beisson, Y.; Shorrosh, B.; Beisson, F.; Andersson, M.X.; Arondel, V.; Bates, P.D.; Baud, S.; Bird, D.; DeBono, A.; Durrett, T.P.; et al. Acyl-lipid metabolism. The Arabidopsis Book 2013, 11, e0161. [Google Scholar] [CrossRef] [Green Version]
- Kosma, D.K.; Molina, I.; Ohlrogge, J.B.; Pollard, M. Identification of an Arabidopsis fatty alcohol:caffeoyl-coenzyme a acyltransferase required for the synthesis of alkyl hydroxycinnamates in root waxes. Plant Physiol. 2012, 160, 237–248. [Google Scholar] [CrossRef] [Green Version]
- Molina, I.; Kosma, D. Role of HXXXD-motif/BAHD acyltransferases in the biosynthesis of extracellular lipids. Plant Cell Rep. 2015, 34, 587–601. [Google Scholar] [CrossRef]
- Thomson, N.; Evert, R.F.; Kelman, A. Wound healing in whole potato tubers: A cytochemical, fluorescence, and ultrastructural Analysis of ut and bruise wounds. Can. J. Bot. 1995, 73, 1436–1450. [Google Scholar] [CrossRef]
- Yang, W.-L.; Bernards, M.A. Wound-induced metabolism in potato (Solanum tuberosum) tubers: Biosynthesis of aliphatic domain monomers. Plant Signal. Behav. 2006, 1, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Biggs, A.R. Suberized boundary zones and the chronology of wound response in tree bark. Phytopathology 1985, 75, 1191–1195. [Google Scholar] [CrossRef]
- Lulai, E.C.; Campbell, L.G.; Fugate, K.K.; McCue, K.F. Biological differences that distinguish the 2 major stages of wound healing in potato tubers. Plant Signal. Behav. 2016, 11, e1256531. [Google Scholar] [CrossRef] [Green Version]
- Baldacci-Cresp, F.; Moussawi, J.; Leplé, J.-C.; Acker, R.V.; Kohler, A.; Candiracci, J.; Twyffels, L.; Spokevicius, A.V.; Bossinger, G.; Laurans, F.; et al. PtaRHE1, a Populus tremula × Populus alba RING-H2 protein of the ATL family, has a regulatory role in secondary phloem fibre development. Plant J. 2015, 82, 978–990. [Google Scholar] [CrossRef]
- Yue, F.; Lu, F.; Sun, R.-C.; Ralph, J. Syntheses of lignin-derived thioacidolysis monomers and their uses as quantitation standards. J. Agric. Food Chem. 2012, 60, 922–928. [Google Scholar] [CrossRef]
- Doster, M.A.; Bostock, R.M. Quantification of lignin formation in almond bark in response to wounding and infection by Phytophthora species. Phytopathology 1988, 78, 473–477. [Google Scholar] [CrossRef]
- Trockenbrodt, M. Light and electron microscopic investigations on wound reactions in the bark of Salix caprea L. and Tilia tomentosa MOENCH. Flora 1994, 189, 131–140. [Google Scholar] [CrossRef]
- Ralph, S.G.; Chun, H.J.E.; Cooper, D.; Kirkpatrick, R.; Kolosova, N.; Gunter, L.; Tuskan, G.A.; Douglas, C.J.; Holt, R.A.; Jones, S.J.; et al. Analysis of 4,664 high-quality sequence-finished poplar full-length cdna clones and their utility for the discovery of genes responding to insect feeding. BMC Genom. 2008, 9, 57. [Google Scholar] [CrossRef] [Green Version]
- Cosio, C.; Dunand, C. Specific functions of individual class III peroxidase genes. J. Exp. Bot. 2009, 60, 391–408. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Li, Q.; Tunlaya-Anukit, S.; Shi, R.; Sun, Y.-H.; Wang, J.P.; Liu, J.; Loziuk, P.; Edmunds, C.W.; Miller, Z.D.; et al. A cell wall-bound anionic peroxidase, PTRPO21, is involved in lignin polymerization in Populus trichocarpa. Tree Genet. Genomes 2016, 12, 22. [Google Scholar] [CrossRef]
- Alassimone, J.; Fujita, S.; Doblas, V.G.; van Dop, M.; Barberon, M.; Kalmbach, L.; Vermeer, J.E.M.; Rojas-Murcia, N.; Santuari, L.; Hardtke, C.S.; et al. Polarly localized kinase SGN1 is required for Casparian strip integrity and positioning. Nat. Plants 2016, 2, 1–10. [Google Scholar] [CrossRef]
- Rojas-Murcia, N.; Hématy, K.; Lee, Y.; Emonet, A.; Ursache, R.; Fujita, S.; Bellis, D.D.; Geldner, N. High-order mutants reveal an essential requirement for peroxidases but not laccases in casparian strip lignification. Proc. Natl. Acad. Sci. USA 2020, 117, 29166–29177. [Google Scholar] [CrossRef]
- Serra, O.; Hohn, C.; Franke, R.; Prat, S.; Molinas, M.; Figueras, M. A feruloyl transferase involved in the biosynthesis of suberin and suberin-associated wax is required for maturation and sealing properties of potato periderm. Plant J. 2010, 62, 277–290. [Google Scholar] [CrossRef]
- Hatfield, R.D.; Rancour, D.M.; Marita, J.M. Grass cell walls: A story of cross-linking. Front. Plant Sci. 2017, 7, 203–215. [Google Scholar] [CrossRef] [Green Version]
- Serra, O.; Soler, M.; Hohn, C.; Franke, R.; Schreiber, L.; Prat, S.; Molinas, M.; Figueras, M. Silencing of StKCS6 in potato periderm leads to reduced chain lengths of suberin and wax compounds and increased peridermal transpiration. J. Exp. Bot. 2009, 60, 697–707. [Google Scholar] [CrossRef] [Green Version]
- Moire, L.; Schmutz, A.; Buchala, A.; Yan, B.; Stark, R.E.; Ryser, U. Glycerol is a suberin monomer. New experimental evidence for an old hypothesis. Plant Physiol. 1999, 119, 1137–1146. [Google Scholar] [CrossRef] [Green Version]
- Soler, M.; Verdaguer, R.; Fernández-Piñán, S.; Company-Arumí, D.; Boher, P.; Góngora-Castillo, E.; Valls, M.; Anticó, E.; Molinas, M.; Serra, O.; et al. Silencing against the conserved NAC domain of the potato stnac103 reveals new nac candidates to repress the suberin associated waxes in phellem. Plant Sci. 2019, 291, 110360. [Google Scholar] [CrossRef]
- Soler, M.; Serra, O.; Fluch, S.; Molinas, M.; Figueras, M. A Potato skin SSH library yields new candidate genes for suberin biosynthesis and periderm formation. Planta 2011, 233, 933–945. [Google Scholar] [CrossRef]
- Legay, S.; Guerriero, G.; André, C.; Guignard, C.; Cocco, E.; Charton, S.; Boutry, M.; Rowland, O.; Hausman, J.F. MdMYB93 is a regulator of suberin deposition in russeted apple fruit skins. New Phytol. 2016, 212, 977–991. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A New mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. Relative Quantification. In Real-Time PCR; Dorak, M.T., Ed.; Taylor & Francis: New York, NY, USA, 2006; pp. 63–82. [Google Scholar]
- Pettengill, E.A.; Parmentier-Line, C.; Coleman, G.D. Evaluation of QPCR reference genes in two genotypes of Populus for use in photoperiod and low-temperature studies. BMC Res. Notes 2012, 5, 366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, L.-J.; Alabady, M.S.; Mohebbi, M.; Tsai, C.-J. Exploiting genome variation to improve next-generation sequencing data analysis and genome editing efficiency in Populus tremula × alba 717-1B4. Tree Genet. Genomes 2015, 11, 82. [Google Scholar] [CrossRef]
- Koressaar, T.; Remm, M. Enhancements and modifications of primer design program Primer3. Bioinformatics 2007, 23, 1289–1291. [Google Scholar] [CrossRef] [Green Version]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, 1006–1007. [Google Scholar] [CrossRef] [Green Version]
- Molina, I.; Bonaventure, G.; Ohlrogge, J.; Pollard, M. The lipid polyester composition of Arabidopsis thaliana and Brassica napus seeds. Phytochemistry 2006, 67, 2597–2610. [Google Scholar] [CrossRef]
- Jacobsen, K.R.; Fisher, D.G.; Maretzki, A.; Moore, P.H. Developmental-changes in the anatomy of the sugarcane stem in relation to phloem unloading and sucrose storage. Bot. Acta 1992, 105, 70–80. [Google Scholar] [CrossRef]
- Graça, J.; Pereira, H. Glyceryl-acyl and aryl-acyl dimers in Pseudotsuga Menziesii bark suberin. Holzforschung 1999, 53, 397–402. [Google Scholar] [CrossRef]
- Graça, J.; Pereira, H. Cork Suberin: A glyceryl based polyester. Holzforschung 1997, 51, 225–234. [Google Scholar] [CrossRef]
- Graça, J.; Pereira, H. Feruloyl Esters of Omega-Hydroxyacids in Cork Suberin. J. Wood Chem. Technol. 1998, 18, 207–217. [Google Scholar] [CrossRef]
- Graça, J.; Pereira, H. Diglycerol alkenedioates in suberin: Building units of a poly(acylglycerol) polyester. Biomacromolecules 2000, 1, 519–522. [Google Scholar] [CrossRef]
- Graça, J.; Pereira, H. Methanolysis of bark suberins: Analysis of glycerol and acid monomers. Phytochem. Anal. 2000, 11, 45–51. [Google Scholar] [CrossRef]
- Graça, J.; Pereira, H. Suberin structure in potato periderm: Glycerol, long-chain monomers, and glyceryl and feruloyl dimers. J. Agric. Food Chem. 2000, 48, 5476–5483. [Google Scholar] [CrossRef]
- Schmutz, A.; Buchala, A.J.; Ryser, U. Changing the dimensions of suberin lamellae of green cotton fibers with a specific inhibitor of the endoplasmic reticulum-associated fatty acid elongases. Plant Physiol. 1996, 110, 403–411. [Google Scholar] [CrossRef] [Green Version]
- Schmutz, A.; Jenny, T.; Amrhein, N.; Ryser, U. Caffeic acid and glycerol are constituents of the suberin layers in green cotton fibres. Planta 1993, 189, 453–460. [Google Scholar] [CrossRef]
- Bonaventure, G.; Beisson, F.; Ohlrogge, J.; Pollard, M. Analysis of the Aliphatic Monomer Composition of Polyesters Associated with Arabidopsis Epidermis: Occurrence of Octadeca-Cis-6, Cis-9-Diene-1,18-Dioate as the Major Component. Plant J. 2004, 40, 920–930. [Google Scholar] [CrossRef]
- Rolando, C.; Monties, B.; Lapierre, C. Thioacidolysis. In Methods in Lignin Chemistry; Lin, S.Y., Dence, C.W., Eds.; Springer Series in Wood Science; Springer: Berlin/Heidelberg, Germany, 1992; pp. 334–349. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Poplar ID | Best TAIR 10 BLAST | Annotation | Left Primer | Right Primer |
|---|---|---|---|---|
| Potri.001G036900 | AT3G21240 | 4CL2 | ATTCATCAGGGACTACAGGGTT | GCAACACACACAAAATCACATCT |
| Potri.014G166600 | AT5G41040 | ASFT | TGAAACAAGGAAAGCCAACTCT | CCTCAACAAAAACAGCACCTTC |
| Potri.004G133500 | AT3G48750 | CDC2 | TTACTTCTTTGCCCGACTTCA | TATTCATGCTCCAAAGCACTCC |
| Potri.009G043700 | AT5G58860 | CYP86A1 | GCGTTTATATCCGTCAGTACCACAA | GCTCTTCATCCGTCCAACAGAATAA |
| Potri.014G085800 | AT1G01600 | CYP86A4 | CTTTCCTTCAACACGTAGCACTCAA | TTGATCAGGACCGTGCAAATTTCAT |
| Potri.005G092200 | AT5G23190 | CYP86B1 | GCAGGAGATAAAGAAGATGGACTA | TATTGCCTCCATCCTTCCCA |
| Potri.019G024800 | AT5G18430 | GDSL (A)/CUS3 | TATTGTCAAACTTGTGCCCTAACCG | ATGATGGTGCTGAGGTTCATAGGAT |
| Potri.001G191400 | AT3G16370 | GDSL (B) | AGATGCTCTGGTTCCTGCTATCATA | GGGAGGGTAATTGGCCTTGAATATC |
| Potri.002G253400 | AT4G28780 | GDSL (C)/CUS4 | TGCTTTCTTTGTGTTTGGTGACTCA | GGGAAGTTGAAGCCATTAGAGAAGC |
| Potri.010G201200 | AT3G11430 | GPAT5 | TGGCAATGAATTACAGGGTTGGTTT | GGCAACTGATTCAAGAAAGTCACCT |
| Potri.008G058200 | AT5G06090 | GPAT7 | TTGAGTCAGTTGCCAGTAGAAGCTA | TCGTGCATTCAAATCCTAACGTAGC |
| Potri.014G085500 | AT4G00400 | GPAT8 | CCTTGATCGCTTGCCAGAAGAAATA | TCGAATCCTAACACTTCACCCAAC |
| Potri.018G105500 | AT5G48930 | HCT | AGAGAGACACCCAACAGGAC | TAGAACGGCACAAGAACATCAC |
| Potri.006G218000 | AT2G26250 | KCS10 | TGTCACGCCCAAGGTCTATTTATCT | ATCAAACTTGCCTGATTTCCTTGCT |
| Potri.013G119600 | AT5G04530 | KCS19 | TAGAGGGAAGAGAGGAATCTGCAAC | AGAACAGAGACACCGAAGAGACAAT |
| Potri.010G125300 | AT1G68530 | KCS6 | AACACCTCATCGTCCTCGCTTT | CAAACCCTGTCTCCTCTCCTCATC |
| Potri.011G041600 | AT4G21440 | MYB 102 | TCCAATAACAGCAACCAGACCTTTG | TAGCTATCCCTCTCATCTTCAGTGC |
| Potri.015G046800 | AT3G18400 | NAC 58 | AACCATCCTCTCAGCAATCTCTAGG | CCACTTGATGAACTGGCAATACGAT |
| Potri.005G195700 | AT1G71695 | PRX12 | TTACCTCAGACCAAGACCTGTACTC | CTGACAATACGCTAAGCTGTGACAT |
| Potri.013G083600 | AT5G05340 | PRX52 | CCACCTCCAACTTCTAACTTGAACC | ATATATGCGTGCCCTAAAGTTCGTG |
| Potri.013G156500 | AT5G58400 | PRX68 | ACTCCTCAGCTCAACTCTCAACAAA | AAAGAACAAGCGAACAAGAGAAGCA |
| Potri.009G093200 | AT4G34270 | TIP41 like | TAACTGGCTGGAAACAAGAGG | TTTCACTACCACAATAAGGCGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rains, M.K.; Caron, C.; Regan, S.; Molina, I. Chemical and Molecular Characterization of Wound-Induced Suberization in Poplar (Populus alba × P. tremula) Stem Bark. Plants 2022, 11, 1143. https://doi.org/10.3390/plants11091143
Rains MK, Caron C, Regan S, Molina I. Chemical and Molecular Characterization of Wound-Induced Suberization in Poplar (Populus alba × P. tremula) Stem Bark. Plants. 2022; 11(9):1143. https://doi.org/10.3390/plants11091143
Chicago/Turabian StyleRains, Meghan K., Christine Caron, Sharon Regan, and Isabel Molina. 2022. "Chemical and Molecular Characterization of Wound-Induced Suberization in Poplar (Populus alba × P. tremula) Stem Bark" Plants 11, no. 9: 1143. https://doi.org/10.3390/plants11091143
APA StyleRains, M. K., Caron, C., Regan, S., & Molina, I. (2022). Chemical and Molecular Characterization of Wound-Induced Suberization in Poplar (Populus alba × P. tremula) Stem Bark. Plants, 11(9), 1143. https://doi.org/10.3390/plants11091143