
Seasonal Changes in the Plant Growth-Inhibitory Effects of Rosemary Leaves on Lettuce Seedlings

 ,
,  ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
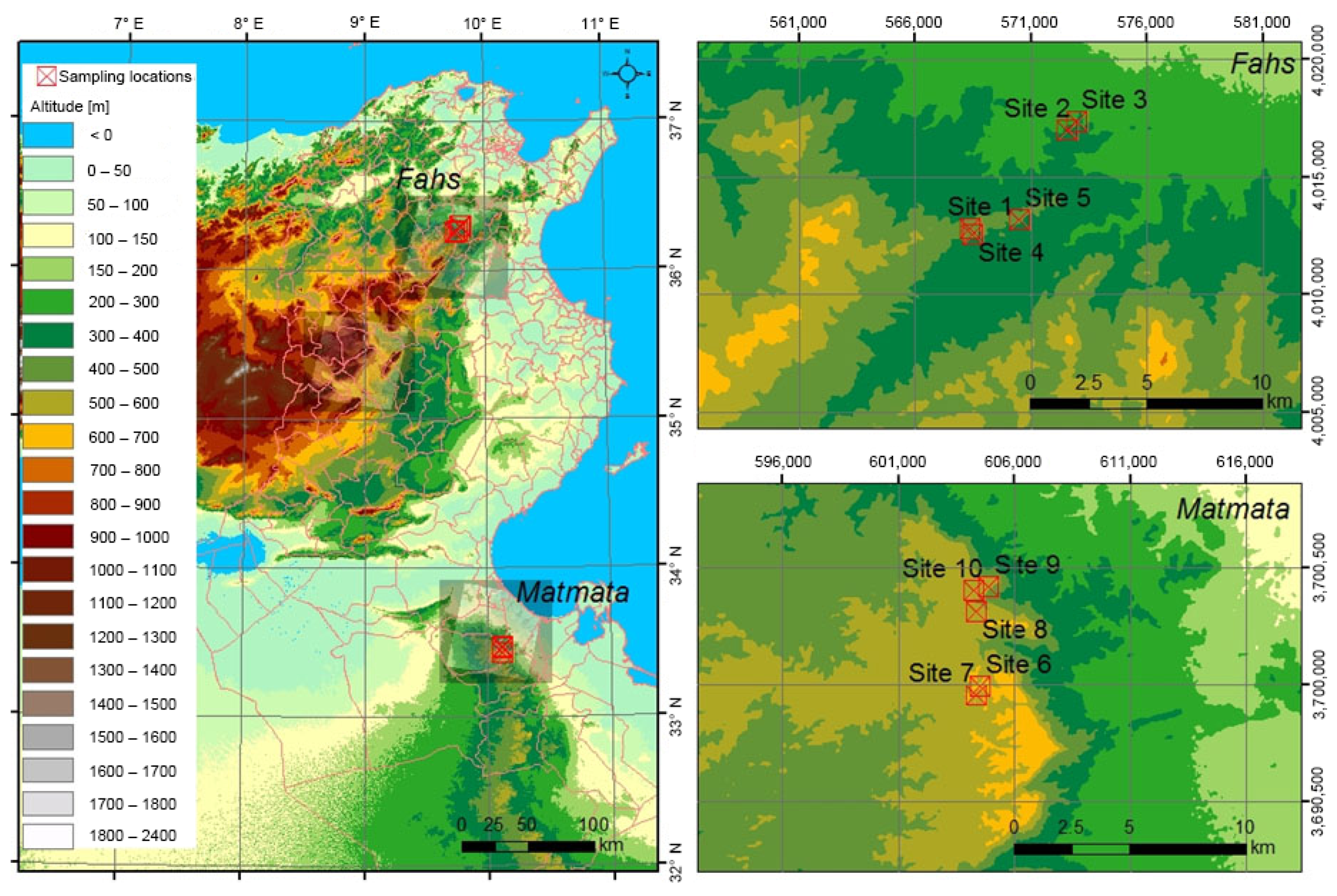
2.1. Collection of Plant Samples
2.2. Extraction Procedure
2.3. Chemicals and Reagents
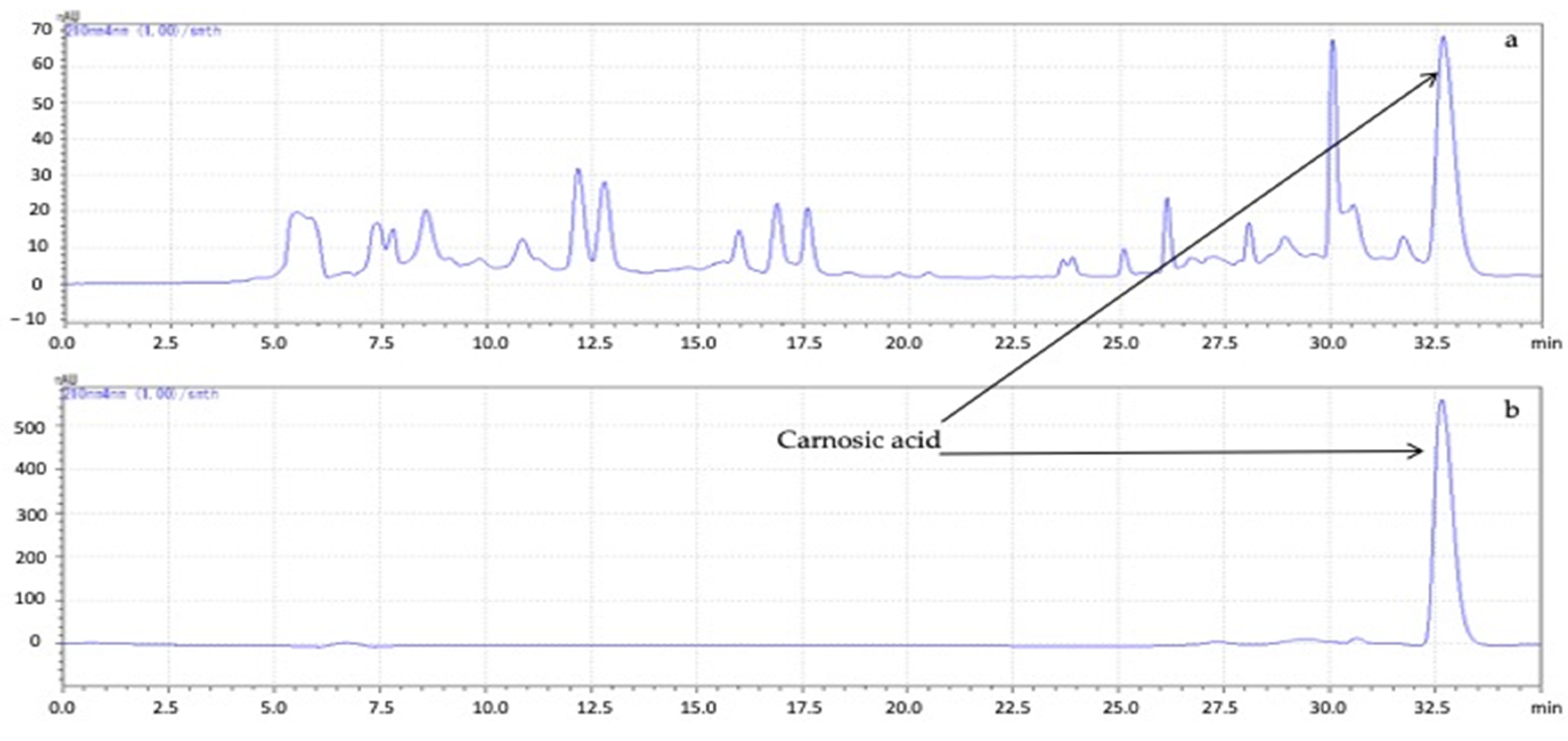
2.4. High-Performance Liquid Chromatography (HPLC) Analysis
2.5. Phytotoxic Activity Bioassay
2.6. Statistical Analysis
3. Results
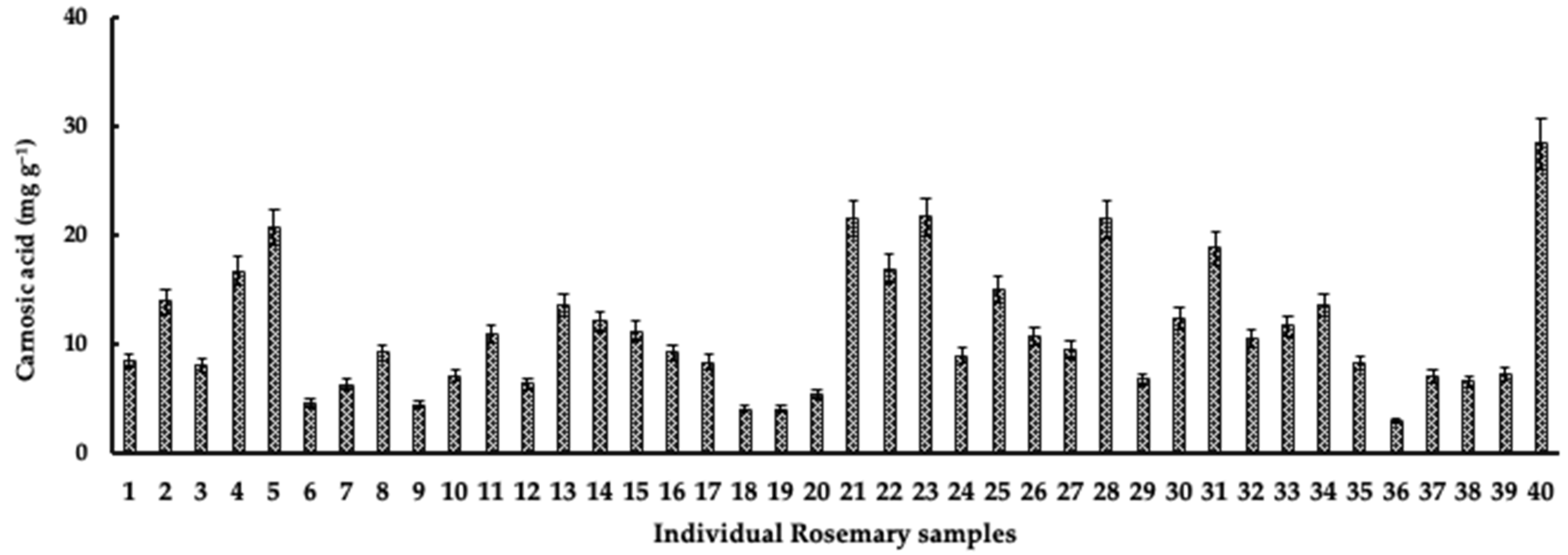
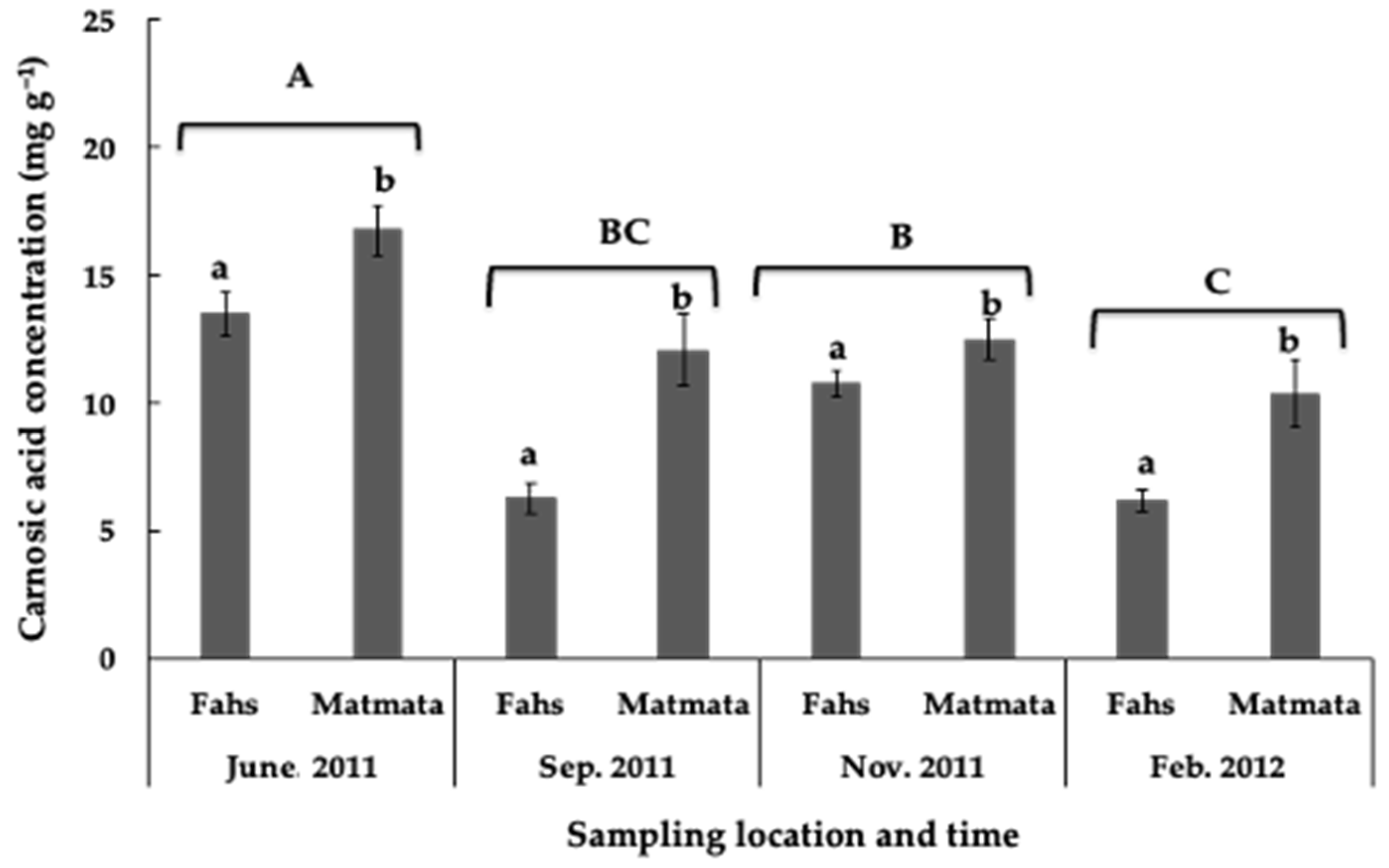
3.1. Variations in Carnosic Acid Concentration in Rosemary Leaves during the Growing Season
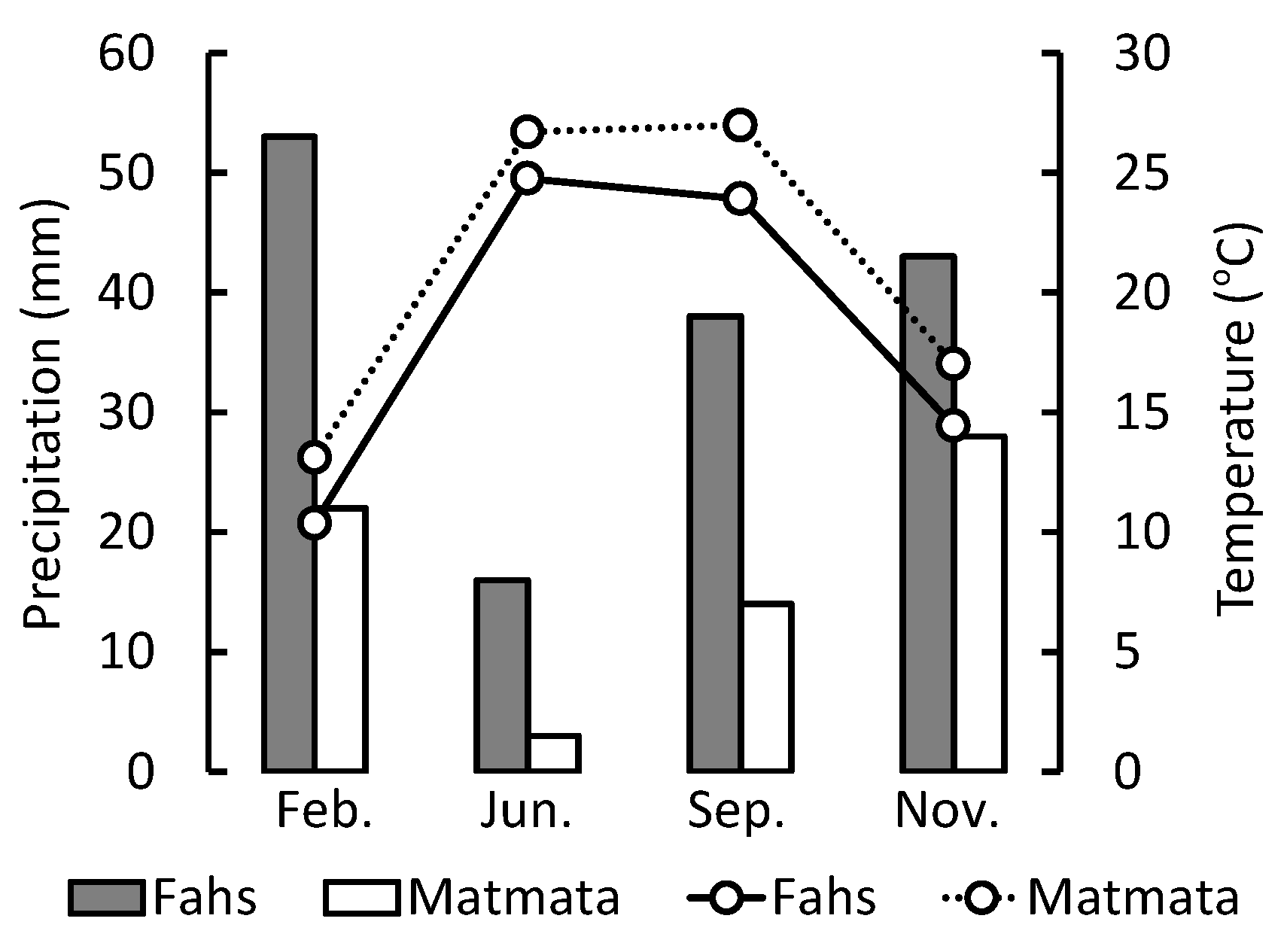
3.2. Influence of Precipitation, Elevation, and Temperature on Carnosic Acid Concentration in Rosemary Leaves
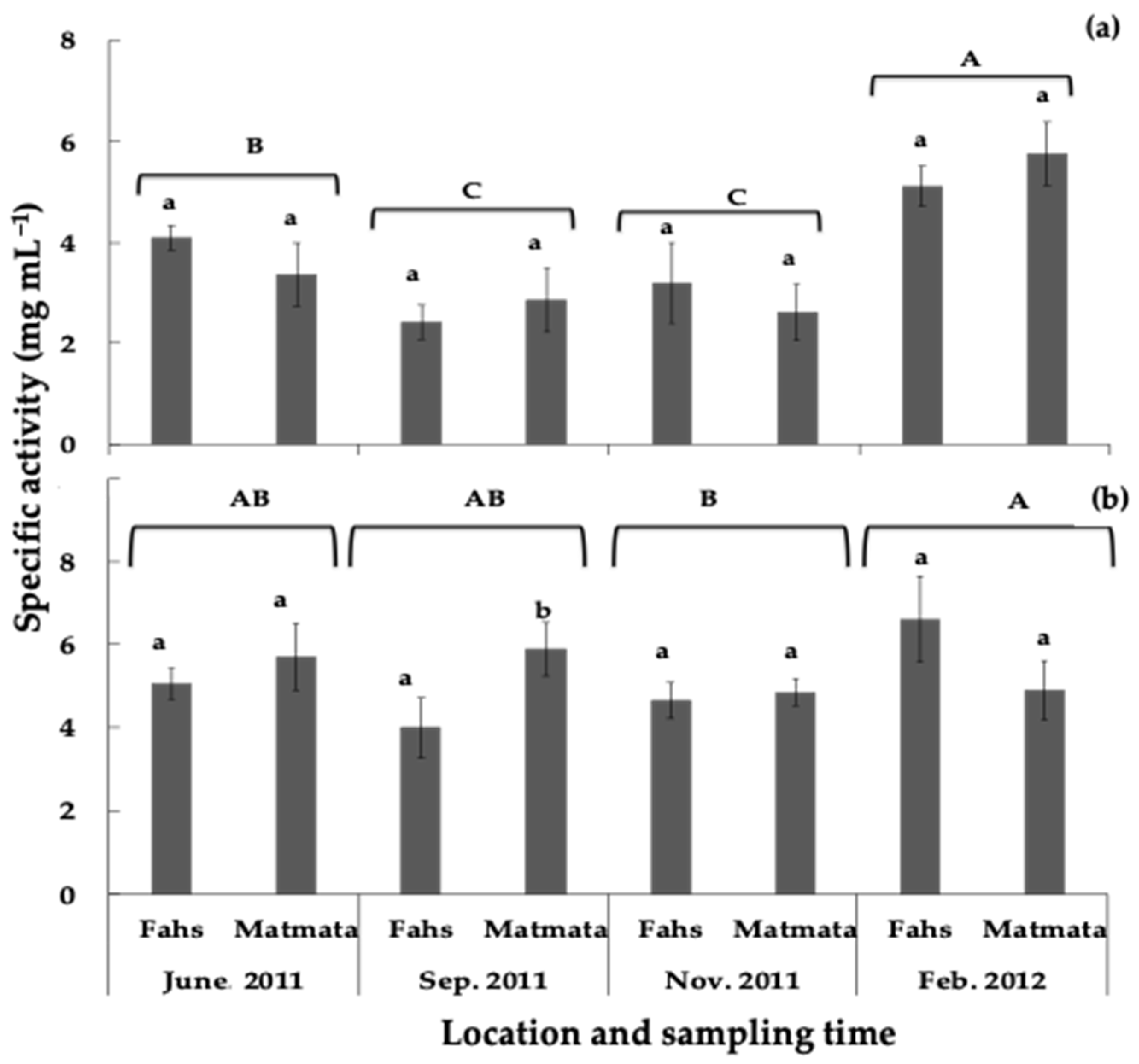
3.3. Effects of Seasonality on the Plant Growth-Inhibitory Potential of Rosemary Leaves
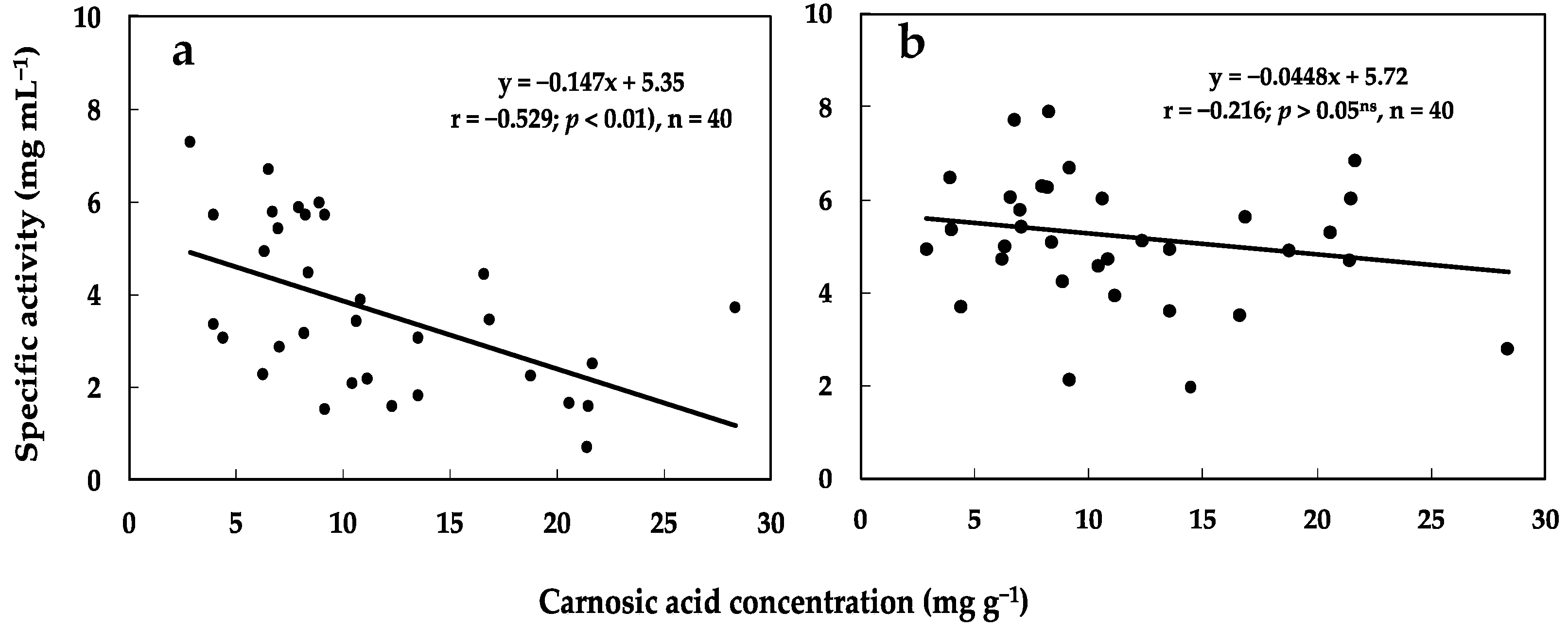
3.4. Correlation between Carnosic Acid Concentration and Phytotoxicity of Rosemary Leaves
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bastiaans, L.; Kropff, M.J.; Goudriaan, J.; van Laar, H.H. Design of Weed Management Systems with a Reduced Reliance on Herbicides Poses New Challenges and Prerequisites for Modelling Crop-Weed Interactions. Field Crops Res. 2000, 67, 161–179. [Google Scholar] [CrossRef]
- Heap, I. Global Perspective of Herbicide-Resistant Weeds. Pest Manag. Sci. 2014, 70, 1306–1315. [Google Scholar] [CrossRef] [PubMed]
- Atwood, D.; Paisley-Jones, C. 2008–2012 Market Estimates; The United States Environmental Protection Agency: Washington, DC, USA, 2017.
- Rice, E.L. Allelopathy. In Physiological Ecology; Kozlowski, T.T., Ed.; Academic Press: New York, NY, USA, 1984. [Google Scholar]
- Willis, R.J. What Is Allelopathy? In The History of Allelopathy; Springer: Dordrecht, The Netherlands, 2007; pp. 1–13. [Google Scholar]
- Inderjit. Soil Microorganisms: An Important Determinant of Allelopathic Activity. Plant Soil 2005, 274, 227–236. [Google Scholar] [CrossRef]
- Teoh, E.S. Medicinal Orchids of Asia; Springer: Cham, Switzerland, 2016; pp. 1–752. [Google Scholar] [CrossRef]
- Wink, M. Modes of Action of Herbal Medicines, and Plant Secondary Metabolites. Medicines 2015, 2, 251–286. [Google Scholar] [CrossRef] [PubMed]
- Wink, M. Secondary Metabolites, the Role in Plant Diversification Of. Encycl. Evol. Biol. 2016, 4, 1–9. [Google Scholar] [CrossRef]
- Cheng, F.; Cheng, Z. Corrigendum: Research Progress on the Use of Plant Allelopathy in Agriculture and the Physiological and Ecological Mechanisms of Allelopathy. Front. Plant Sci. 2016, 7, 1697. [Google Scholar] [CrossRef]
- Soltys, D.; Krasuska, U.; Bogatek, R.; Gniazdowsk, A. Allelochemicals as Bioherbicides—Present and Perspectives. In Herbicides Current Research and Case Studies in Use; IntechOpen: London, UK, 2013. [Google Scholar] [CrossRef]
- Macías, F.A.; Mejías, F.J.R.; Molinillo, J.M.G. Recent Advances in Allelopathy for Weed Control: From Knowledge to Applications. Pest Manag. Sci. 2019, 75, 2413–2436. [Google Scholar] [CrossRef]
- Zeng, R.H.; Mallik, A.U.; Luo, S.M. Allelopathy in Sustainable Agriculture and Forestry; Springer: New York, NY, USA, 2008. [Google Scholar]
- Jabran, K.; Mahajan, G.; Sardana, V.; Chauhan, B.S. Allelopathy for Weed Control in Agricultural Systems. Crop Prot. 2015, 72, 57–65. [Google Scholar] [CrossRef]
- Dayan, F.E.; Owens, D.K.; Duke, S.O. Rationale for a Natural Products Approach to Herbicide Discovery. Pest Manag. Sci. 2012, 68, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Heil, M.; Bostock, R.M. Induced Systemic Resistance (ISR) against Pathogens in the Context of Induced Plant Defences. Ann. Bot. 2002, 89, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Lur, H.; Hsu, C.; Wu, C.; Lee, C.; Lao, C.; Wu, Y.; Chang, S.; Wang, C. Changes in Temperature, Cultivation Timing and Grain Quality of Rice in Taiwan in Recent Years. Crop Environ. Bioinform. 2009, 6, 175–182. [Google Scholar]
- Gray, D.E.; Pallardy, S.G.; Garrett, H.E.; Rottinghaus, G.E. Effect of Acute Drought Stress and Time of Harvest on Phytochemistry and Dry Weight of St. John’s Wort Leaves and Flowers. Planta Med. 2003, 69, 1024–1030. [Google Scholar] [CrossRef]
- Catalán, P.; Vázquez-de-Aldana, B.R.; de las Heras, P.; Fernández-Seral, A.; Pérez-Corona, M.E. Comparing the Allelopathic Potential of Exotic and Native Plant Species on Understory Plants: Are Exotic Plants Better Armed? An. Biol. 2013, 35, 65–74. [Google Scholar] [CrossRef]
- Lou, Y.; Davis, A.S.; Yannarell, A.C. Interactions between Allelochemicals and the Microbial Community Affect Weed Suppression Following Cover Crop Residue Incorporation into Soil. Plant Soil 2016, 399, 357–371. [Google Scholar] [CrossRef]
- Gagliardo, R.W.; Chilton, W.S. Soil Transformation of 2(3H)-Benzoxazolone of Rye into Phytotoxic 2-Amino-3H-Phenoxazin-3-One. J. Chem. Ecol. 1992, 18, 1683–1691. [Google Scholar] [CrossRef]
- Silva, E.R.; Overbeck, G.E.; Soares, G.L.G. Phytotoxicity of Volatiles from Fresh and Dry Leaves of Two Asteraceae Shrubs: Evaluation of Seasonal Effects. S. Afr. J. Bot. 2014, 93, 14–18. [Google Scholar] [CrossRef]
- Frizzo, C.D.; Atti-Serafini, L.; Laguna, S.E.; Cassel, E.; Lorenzo, D.; Dellacassa, E. Essential Oil Variability in Baccharis uncinella DC and Baccharis dracunculifolia DC Growing Wild in Southern Brazil, Bolivia, and Uruguay. Flavour Fragr. J. 2008, 23, 99–106. [Google Scholar] [CrossRef]
- Anese, S.; Grisi, P.U.; de Jesus Jatobá, L.; Imatomi, M.; de Cassia Pereira, V.; Juliano Gualtieri, S.C. Seasonal Variation in Phytotoxicity of Drimys brasiliensis Miers. Idesia 2014, 32, 109–116. [Google Scholar] [CrossRef][Green Version]
- Missouri Botanical Garden. Salvia rosmarinus—Plant Finder. Available online: http://www.missouribotanicalgarden.org/PlantFinder/PlantFinderDetails.aspx?kempercode=b968 (accessed on 20 August 2021).
- Royal Botanic Gardens. Salvia rosmarinus Spenn. | Plants of the World Online | Kew Science. Available online: http://plantsoftheworldonline.org/taxon/urn:lsid:ipni.org:names:457138-1 (accessed on 20 August 2021).
- UniProt. Taxanomy-Rosmarinus officinalis (Rosemary) (Salvia rosmarinus). Available online: https://www.uniprot.org/taxonomy/39367 (accessed on 20 August 2021).
- Sasaki, K.; el Omri, A.; Kondo, S.; Han, J.; Isoda, H. Rosmarinus officinalis Polyphenols Produce Anti-Depressant like Effect through Monoaminergic and Cholinergic Functions Modulation. Behav. Brain Res. 2013, 238, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Kontogianni, V.G.; Tomic, G.; Nikolic, I.; Nerantzaki, A.A.; Sayyad, N.; Stosic-Grujicic, S.; Stojanovic, I.; Gerothanassis, I.P.; Tzakos, A.G. Phytochemical Profile of Rosmarinus officinalis and Salvia officinalis Extracts and Correlation to their Antioxidant and Anti-Proliferative Activity. Food Chem. 2013, 136, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Bakirel, T.; Bakirel, U.; Keleş, O.Ü.; Ülgen, S.G.; Yardibi, H. In Vivo Assessment of Antidiabetic and Antioxidant Activities of Rosemary (Rosmarinus officinalis) in Alloxan-Diabetic Rabbits. J. Ethnopharmacol. 2008, 116, 64–73. [Google Scholar] [CrossRef]
- Thorsen, M.A.; Hildebrandt, K.S. Quantitative Determination of Phenolic Diterpenes in Rosemary Extracts. J. Chromatogr. A 2003, 995, 119–125. [Google Scholar] [CrossRef]
- Erkan, N.; Ayranci, G.; Ayranci, E. Antioxidant Activities of Rosemary (Rosmarinus officinalis L.) Extract, Blackseed (Nigella sativa L.) Essential Oil, Carnosic Acid, Rosmarinic Acid and Sesamol. Food Chem. 2008, 110, 76–82. [Google Scholar] [CrossRef]
- Amaral, G.P.; Mizdal, C.R.; Stefanello, S.T.; Mendez, A.S.L.; Puntel, R.L.; de Campos, M.M.A.; Soares, F.A.A.; Fachinetto, R. Antibacterial and Antioxidant Effects of Rosmarinus officinalis L. Extract and Its Fractions. J. Tradit. Complement. Med. 2019, 9, 383–392. [Google Scholar] [CrossRef]
- Appiah, K.S.; Mardani, H.K.; Omari, R.A.; Eziah, V.Y.; Ofosu-Anim, J.; Onwona-Agyeman, S.; Amoatey, C.A.; Kawada, K.; Katsura, K.; Oikawa, Y.; et al. Involvement of Carnosic Acid in the Phytotoxicity of Rosmarinus officinalis Leaves. Toxins 2018, 10, 498. [Google Scholar] [CrossRef] [PubMed]
- Sekine, T.; Appiah, K.S.; Azizi, M.; Fujii, Y. Plant Growth Inhibitory Activities and Volatile Active Compounds of 53 Spices and Herbs. Plants 2020, 9, 264. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.B.; Rai, D.K.; Brunton, N.P.; Martin-Diana, A.B.; Barry-Ryan, A.C. Characterization of Phenolic Composition in Lamiaceae Spices by LC-ESI-MS/MS. J. Agric. Food Chem. 2010, 58, 10576–10581. [Google Scholar] [CrossRef] [PubMed]
- Luis, J.C.; Johnson, C.B. Seasonal Variations of Rosmarinic and Carnosic Acids in Rosemary Extracts. Analysis of their in Vitro Antiradical Activity. Span. J. Agric. Res. 2005, 3, 106–112. [Google Scholar] [CrossRef]
- Achour, S.; Khelifi, E.; Attia, Y.; Ferjani, E.; Noureddine Hellal, A. Concentration of Antioxidant Polyphenols from Thymus capitatus Extracts by Membrane Process Technology. J. Food Sci. 2012, 77, 6. [Google Scholar] [CrossRef] [PubMed]
- Richheimer, S.L.; Bernart, M.W.; King, G.A.; Kent, M.C.; Bailey, D.T. Antioxidant Activity of Lipid-Soluble Phenolic Diterpenes from Rosemary. J. Am. Oil Chem. Soc. 1996, 73, 507–514. [Google Scholar] [CrossRef]
- Wellwood, C.R.L.; Cole, R.A. Relevance of Carnosic Acid Concentrations to the Selection of Rosemary, Rosmarinus officinalis (L.), Accessions for Optimization of Antioxidant Yield. J. Agric. Food Chem. 2004, 52, 6101–6107. [Google Scholar] [CrossRef] [PubMed]
- del Baño, M.J.; Lorente, J.; Castillo, J.; Benavente-García, O.; del Río, J.A.; Ortuño, A.; Quirin, K.W.; Gerard, D. Phenolic Diterpenes, Flavones, and Rosmarinic Acid Distribution during the Development of Leaves, Flowers, Stems, and Roots of Rosmarinus officinalis. Antioxidant Activity. J. Agric. Food Chem. 2003, 51, 4247–4253. [Google Scholar] [CrossRef] [PubMed]
- Jaouadi, S.; Lebreton, V.; Bout-Roumazeilles, V.; Siani, G.; Lakhdar, R.; Boussoffara, R.; Dezileau, L.; Kallel, N.; Mannai-Tayech, B.; Combourieu-Nebout, N. Environmental Changes, Climate and Anthropogenic Impact in South-East Tunisia during the Last 8 Kyr. Clim. Past 2016, 12, 1339–1359. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-Km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Borrás-Linares, I.; Stojanović, Z.; Quirantes-Piné, R.; Arráez-Román, D.; Švarc-Gajić, J.; Fernández-Gutiérrez, A.; Segura-Carretero, A. Rosmarinus officinalis Leaves as a Natural Source of Bioactive Compounds. Int. J. Mol. Sci. 2014, 15, 20585–20606. [Google Scholar] [CrossRef]
- Hidalgo, P.J.; Ubera, J.L.; Tena, M.T.; Valcárcel, M. Determination of the Carnosic Acid Content in Wild and Cultivated Rosmarinus officinalis. J. Agric. Food Chem. 1998, 46, 2624–2627. [Google Scholar] [CrossRef]
- Lemos, M.F.; Lemos, M.F.; Endringer, D.C.; Scherer, R. Seasonality Modifies Rosemary’s Composition and Biological Activity. Ind. Crops Prod. 2015, 70, 41–47. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Nogués, S.; Alegre, L. Daily Patterns of Photosynthesis of Two Mediterranean Shrubs in Response to Water Deficit. In Photosynthesis: Mechanisms and Effects; Springer: Dordrecht, The Netherlands, 1998; pp. 4015–4018. [Google Scholar]
- Munné-Bosch, S.; Alegre, L. Changes in Carotenoids, Tocopherols and Diterpenes during Drought and Recovery, and the Biological Significance of Chlorophyll Loss in Rosmarinus officinalis Plants. Planta 2000, 210, 925–931. [Google Scholar] [CrossRef]
- Kasahara, H.; Hanada, A.; Kuzuyama, T.; Takagi, M.; Kamiya, Y.; Yamaguchi, S. Contribution of the Mevalonate and Methylerythritol Phosphate Pathways to the Biosynthesis of Gibberellins in Arabidopsis. J. Biol. Chem. 2002, 277, 45188–45194. [Google Scholar] [CrossRef]
- Birtić, S.; Dussort, P.; Pierre, F.X.; Bily, A.C.; Roller, M. Carnosic Acid. Phytochemistry 2015, 115, 9–19. [Google Scholar] [CrossRef]
- Ishikura, Y.; Kojima, Y.; Terrazawa, M. Effects of Phenolic Compounds on Seed Germination of Shirakamba Birch, Betula platyphylla var. Japonica. Eurasian J. For. Res. 2001, 2, 17–25. [Google Scholar]
- Loussouarn, M.; Krieger-Liszkay, A.; Svilar, L.; Bily, A.; Birtić, S.; Havaux, M. Carnosic Acid and Carnosol, Two Major Antioxidants of Rosemary, Act through Different Mechanisms. Plant Physiol. 2017, 175, 1381–1394. [Google Scholar] [CrossRef] [PubMed]
- Araniti, F.; Costas-Gil, A.; Cabeiras-Freijanes, L.; Lupini, A.; Sunseri, F.; Reigosa, M.J.; Abenavoli, M.R.; Sánchez-Moreiras, A.M. Rosmarinic Acid Induces Programmed Cell Death in Arabidopsis Seedlings through Reactive Oxygen Species and Mitochondrial Dysfunction. PLoS ONE 2018, 13, e0208802. [Google Scholar] [CrossRef] [PubMed]
- Rudrappa, T.; Bonsall, J.; Gallagher, J.L.; Seliskar, D.M.; Bais, H.P. Root-Secreted Allelochemical in the Noxious Weed Phragmites australis Deploys a Reactive Oxygen Species Response and Microtubule Assembly Disruption to Execute Rhizotoxicity. J. Chem. Ecol. 2007, 33, 1898–1918. [Google Scholar] [CrossRef]
- dos Santos, W.D.; Ferrarese, M.L.L.; Nakamura, C.V.; Mourão, K.S.M.; Mangolin, C.A.; Ferrarese-Filho, O. Soybean (Glycine max) Root Lignification Induced by Ferulic Acid. The Possible Mode of Action. J. Chem. Ecol. 2008, 34, 1230–1241. [Google Scholar] [CrossRef] [PubMed]
- Bubna, G.A.; Lima, R.B.; Zanardo, D.Y.L.; dos Santos, W.D.; Ferrarese, M.D.L.L.; Ferrarese-Filho, O. Exogenous Caffeic Acid Inhibits the Growth and Enhances the Lignification of the Roots of Soybean (Glycine max). J. Plant Physiol. 2011, 168, 1627–1633. [Google Scholar] [CrossRef]
- Koitabashi, R.; Suzuki, T.; Kawazu, T.; Sakai, A.; Kuroiwa, H.; Kuroiwa, T. 1, 8-Cineole Inhibits Root Growth and DNA Synthesis in the Root Apical Meristem of Brassica campestris L. J. Plant Res. 1997, 110, 1–6. [Google Scholar] [CrossRef]
- Gouda, N.A.A.; Saad, M.M.G.; Abdelgaleil, S.A.M. PRE and POST Herbicidal Activity of Monoterpenes against Barnyard Grass (Echinochloa crus-galli). Weed Sci. 2016, 64, 191–200. [Google Scholar] [CrossRef]
- Labbé, C.; Faini, F.; Calderón, D.; Molina, J.; Arredondo, S. Variations of Carnosic Acid and Carnosol Concentrations in Ethanol Extracts of Wild Lepechinia salviae in Spring (2008–2011). Nat. Prod. Commun. 2014, 9, 1413–1416. [Google Scholar] [CrossRef]
- Soltanabad, M.H.; Bagherieh-Najjar, M.B.; Mianabadi, M. Seasonal Variations in Carnosic Acid Content of Rosemary Correlates with Anthocyanins and Soluble Sugars. J. Med. Plants By-Prod. 2018, 7, 163–171. [Google Scholar]
- Celiktas, O.Y.; Kocabas, E.E.H.; Bedir, E.; Sukan, F.V.; Ozek, T.; Baser, K.H.C. Antimicrobial Activities of Methanol Extracts and Essential Oils of Rosmarinus officinalis, Depending on Location and Seasonal Variations. Food Chem. 2007, 100, 553–559. [Google Scholar] [CrossRef]
- Ben-Hammouda, M.; Kremer, R.J.; Minor, H.C.; Sarwar, M. A Chemical Basis for Differential Allelopathic Potential of Sorghum Hybrids on Wheat. J. Chem. Ecol. 1995, 21, 6. [Google Scholar] [CrossRef] [PubMed]
- Reberg-Horton, S.C.; Burton, J.D.; Danehower, D.A.; Ma, G.; Monks, D.W.; Murphy, J.P.; Ranells, N.N.; Williamson, J.D.; Creamer, N.G. Changes over Time in the Allelochemical Content of Ten Cultivars of Rye (Secale cereale L.). J. Chem. Ecol. 2005, 31, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Haig, T.; Wu, H.; An, M.; Pratley, J. Correlation between Phytotoxicity on Annual Ryegrass (Lolium rigidum) and Production Dynamics of Allelochemicals within Root Exudates of an Allelopathic Wheat. J. Chem. Ecol. 2003, 29, 2263–2279. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Collection Date | Geographical Area | Elevation (m) | Sample Codes |
|---|---|---|---|---|
| 1 | June 2011 | Fahs | 430 | UT-ARENA 00327 |
| 2 | June 2011 | Fahs | 320 | UT-ARENA 00334 |
| 3 | June 2011 | Fahs | 300 | UT-ARENA 00340 |
| 4 | June 2011 | Fahs | 430 | UT-ARENA 00349 |
| 5 | June 2011 | Fahs | 420 | UT-ARENA 00357 |
| 6 | June 2011 | Matmata | 620 | UT-ARENA 00364 |
| 7 | June 2011 | Matmata | 585 | UT-ARENA 00371 |
| 8 | June 2011 | Matmata | 575 | UT-ARENA 00379 |
| 9 | June 2011 | Matmata | 535 | UT-ARENA 00387 |
| 10 | June 2011 | Matmata | 555 | UT-ARENA 00395 |
| 11 | September 2011 | Fahs | 430 | UT-ARENA 00402 |
| 12 | September 2011 | Fahs | 320 | UT-ARENA 00411 |
| 13 | September 2011 | Fahs | 300 | UT-ARENA 00417 |
| 14 | September 2011 | Fahs | 430 | UT-ARENA 00426 |
| 15 | September 2011 | Fahs | 420 | UT-ARENA 00434 |
| 16 | September 2011 | Matmata | 620 | UT-ARENA 00442 |
| 17 | September 2011 | Matmata | 585 | UT-ARENA 00453 |
| 18 | September 2011 | Matmata | 575 | UT-ARENA 00460 |
| 19 | September 2011 | Matmata | 535 | UT-ARENA 00469 |
| 20 | September 2011 | Matmata | 555 | UT-ARENA 00478 |
| 21 | November 2011 | Fahs | 430 | UT-ARENA 00515 |
| 22 | November 2011 | Fahs | 320 | UT-ARENA 00523 |
| 23 | November 2011 | Fahs | 300 | UT-ARENA 00535 |
| 24 | November 2011 | Fahs | 430 | UT-ARENA 00543 |
| 25 | November 2011 | Fahs | 420 | UT-ARENA 00550 |
| 26 | November 2011 | Matmata | 620 | UT-ARENA 00559 |
| 27 | November 2011 | Matmata | 585 | UT-ARENA 00565 |
| 28 | November 2011 | Matmata | 575 | UT-ARENA 00574 |
| 29 | November 2011 | Matmata | 535 | UT-ARENA 00583 |
| 30 | November 2011 | Matmata | 555 | UT-ARENA 00587 |
| 31 | February 2012 | Fahs | 430 | UT-ARENA 00617 |
| 32 | February 2012 | Fahs | 320 | UT-ARENA 00622 |
| 33 | February 2012 | Fahs | 300 | UT-ARENA 00628 |
| 34 | February 2012 | Fahs | 430 | UT-ARENA 00633 |
| 35 | February 2012 | Fahs | 420 | UT-ARENA 00638 |
| 36 | February 2012 | Matmata | 620 | UT-ARENA 00647 |
| 37 | February 2012 | Matmata | 585 | UT-ARENA 00652 |
| 38 | February 2012 | Matmata | 575 | UT-ARENA 00657 |
| 39 | February 2012 | Matmata | 535 | UT-ARENA 00662 |
| 40 | February 2012 | Matmata | 555 | UT-ARENA 00667 |
| Attribute | Elevation | Precipitation | Temperature | CA Concentration |
|---|---|---|---|---|
| Elevation | 1.00 | |||
| Precipitation | −0.61 ** | 1.00 | ||
| Temperature | 0.19 | −0.71 ** | 1.00 | |
| CA amt | 0.33 * | −0.49 * | 0.30 * | 1.00 |
| Source of Variation | DF | CA Concentration | Hypocotyl Growth | Radicle Growth | |||
|---|---|---|---|---|---|---|---|
| MS | p-level | MS | p-level | MS | p-level | ||
| Location | 1 | 167.4 | <0.001 ** | 0.02 | >0.05 | 0.52 | >0.05 |
| Month | 3 | 61.9 | <0.001 ** | 12.7 | <0.001 ** | 1.6 | <0.05 * |
| Location × Month | 3 | 8.9 | <0.05 * | 0.9 | <0.05 * | 4.5 | <0.001 ** |
| Error | 24 | 2.2 | 0.3 | 0.5 | |||
| Total | 31 | ||||||
| R2 | 0.88 | 0.85 | 0.63 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Appiah, K.S.; Omari, R.A.; Onwona-Agyeman, S.; Amoatey, C.A.; Ofosu-Anim, J.; Smaoui, A.; Arfa, A.B.; Suzuki, Y.; Oikawa, Y.; Okazaki, S.; et al. Seasonal Changes in the Plant Growth-Inhibitory Effects of Rosemary Leaves on Lettuce Seedlings. Plants 2022, 11, 673. https://doi.org/10.3390/plants11050673
Appiah KS, Omari RA, Onwona-Agyeman S, Amoatey CA, Ofosu-Anim J, Smaoui A, Arfa AB, Suzuki Y, Oikawa Y, Okazaki S, et al. Seasonal Changes in the Plant Growth-Inhibitory Effects of Rosemary Leaves on Lettuce Seedlings. Plants. 2022; 11(5):673. https://doi.org/10.3390/plants11050673
Chicago/Turabian StyleAppiah, Kwame Sarpong, Richard Ansong Omari, Siaw Onwona-Agyeman, Christiana Adukwei Amoatey, John Ofosu-Anim, Abderrazak Smaoui, Abdelkarim Ben Arfa, Yoko Suzuki, Yosei Oikawa, Shin Okazaki, and et al. 2022. "Seasonal Changes in the Plant Growth-Inhibitory Effects of Rosemary Leaves on Lettuce Seedlings" Plants 11, no. 5: 673. https://doi.org/10.3390/plants11050673
APA StyleAppiah, K. S., Omari, R. A., Onwona-Agyeman, S., Amoatey, C. A., Ofosu-Anim, J., Smaoui, A., Arfa, A. B., Suzuki, Y., Oikawa, Y., Okazaki, S., Katsura, K., Isoda, H., Kawada, K., & Fujii, Y. (2022). Seasonal Changes in the Plant Growth-Inhibitory Effects of Rosemary Leaves on Lettuce Seedlings. Plants, 11(5), 673. https://doi.org/10.3390/plants11050673