
Genetic Aspects and Molecular Causes of Seed Longevity in Plants—A Review



Abstract
1. Introduction
2. Mechanisms of Seed Ageing
3. Classical Genetic Studies
4. Genetic Studies in the 21st Century
4.1. Arabidopsis thaliana L.
4.2. Rice (Oryza sativa L.)
4.3. Barley (Hordeum vulgare L.)
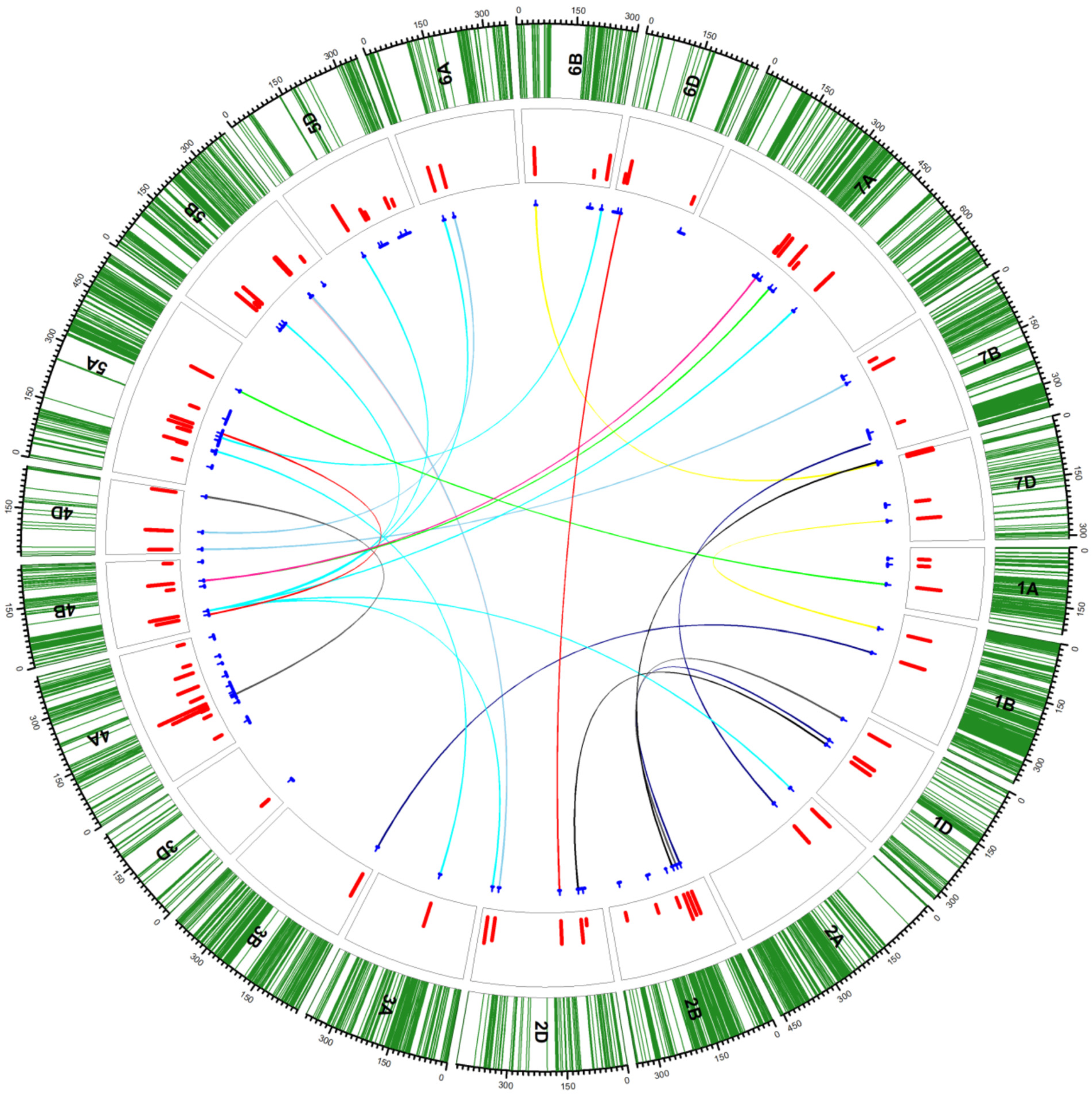
4.4. Wheat (Triticum aestivum L.)
4.5. Maize (Zea mays L.)
4.6. Soybean (Glycine max (L.) Merr.)
4.7. Lettuce (Lactuca sativa L.)
4.8. Tobacco (Nicotiana tabacum L.)
4.9. Tomato (Solanum lycoperscum L.)
5. Future Directions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 1-Cys Prx | 1-cys peroxiredoxin and/or PER1 |
| AA | artificial ageing |
| ABA | abscisic acid |
| AKR1 | aldo-ketoreductases |
| Ale | Aleurain |
| AM | association mapping |
| AS | alternative splicing |
| ASPG1 | ASPARTIC PROTEASE IN GUARD CELL 1 |
| ATM | ATAXIA TELANGIECTASIA MUTATED (ATM) |
| ATR | RAD3-RELATED |
| BIL | backcross inbred lines |
| BR | brassinosteroid |
| BSA-seq | bulked segregant analysis through whole genome sequencing |
| Cas-9 | CRISPR-associated protein-9 |
| CDT | controlled deterioration test |
| CRISPR | clustered regularly interspaced short palindromic repeats |
| CSSL | chromosome segment substitution lines |
| DArT | diversity array technology |
| DEG | differentially expressed genes |
| DH | doubled haploid |
| DREB | dehydration responsive element binding protein |
| EIN2 | ETHYLENE-INSENSITIVE 2-like |
| GA | gibberellins |
| ERE | ethylene-responsive element |
| GAAS | germination ability after storage |
| GBS | genotyping by sequencing (GBS |
| GO | gene ontology |
| GWAS | genome wide association study |
| HSP | heat shock proteins |
| IF2 | immortalized F2 |
| IAA | indole-3-acetic acid [IAA] |
| ITMI/MP | International Triticeae Mapping Initiative” mapping population |
| LACS2 | long-chain acyl-CoA synthetase 2 |
| LEA | late embryogenesis abundant |
| LG | linkage groups |
| LOX | lipoxygenase |
| MACE | massive analysis of cDNA 3′-ends |
| miRNA | microRNA |
| mQTL | meta-QTL |
| NADP-ME | NADP-dependent malic enzyme |
| NAM | nested association mapping |
| NIL | near isogenic line |
| NS | null segregant |
| OS | oligosaccharides |
| OsALDH7 | rice aldehyde dehydrogenase |
| Per2 | peroxidase |
| PIMT1 | protein L-isoaspartyle methyltransferase |
| PPR | pentatricopeptide repeat |
| QTL | quantitative trait loci |
| RAB18 | responsive to abscisic acid 18 |
| RFO | raffinose family oligosaccharide |
| Rluc | Rennila LUCIFERASE reporter |
| RFO | raffinose series oligosaccharides |
| RIL | recombinant inbred population |
| RNA-seq | RNA sequence |
| SNP | single nucleotide polymorphism |
| SSP | seed storage proteins |
| SSR | simple sequence repeats |
| TIP | tonoplast intrinsic proteins |
| TPP | trehalose-6-phosphate phosphatase |
| Vps29 | vacuolar protein sorting 29 |
| Wip | wound-induced protein |
References
- Padulosi, S. Final Report: Conservation and Use of Underutilized Mediterranean Species; International Plant Genetic Resources Institute, Regional Office for Central and West Asia and North Africa (CWANA): Aleppo, Syria Panayotou, 1999. [Google Scholar]
- Levin, S.A. Encyclopedia of Biodiversity; Elsevier Inc.: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Arif, M.A.R.; Börner, A. Mapping of QTL associated with seed longevity in durum wheat (Triticum durum Desf.). J. Appl. Genet. 2019, 60, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Börner, A.; Khlestkina, E.K.; Chebotar, S.; Nagel, M.; Arif, M.A.R.; Neumann, K.; Kobiljski, B.; Lohwasser, U.; Röder, M.S. Molecular markers in management of ex situ PGR–A case study. J. Biosci. 2012, 37, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Sano, N.; Rajjou, L.; North, H.M.; Debeaujon, I.; Marion-Poll, A.; Seo, M. Staying alive: Molecular aspects of seed longevity. Plant Cell Physiol. 2016, 57, 660–674. [Google Scholar] [CrossRef] [PubMed]
- Barton, L.V. Seed Preservation and Longevity; Leornard Hill: London, UK; Inerscience: Hoboken, NY, USA, 1961. [Google Scholar]
- Rehman Arif, M.A.; Börner, A. An SNP based GWAS analysis of seed longevity in wheat. Cereal Res. Commun. 2020, 48, 149–156. [Google Scholar] [CrossRef]
- Roberts, E. Viability of cereal seed for brief and extended periods. Ann. Bot. 1961, 25, 373–380. [Google Scholar] [CrossRef]
- McDonald, M. Seed deterioration: Physiology, repair and assessment. Seed Sci. Technol. 1999, 27, 177–237. [Google Scholar]
- Coolbear, P. Mechanisms of seed deterioration. In Seed Quality; CRC Press: Boca Raton, FL, USA, 2020; pp. 223–277. [Google Scholar]
- McGee, D.C. Pathology of seed deterioration. Genet. Improv. Seed Qual. 2000, 31, 11–19. [Google Scholar]
- Clerkx, E.J.; El-Lithy, M.E.; Vierling, E.; Ruys, G.J.; Blankestijn-De Vries, H.; Groot, S.P.; Vreugdenhil, D.; Koornneef, M. Analysis of natural allelic variation of Arabidopsis seed germination and seed longevity traits between the accessions Landsberg erecta and Shakdara, using a new recombinant inbred line population. Plant Physiol. 2004, 135, 432–443. [Google Scholar] [CrossRef]
- Wettlaufer, S.H.; Leopold, A.C. Relevance of Amadori and Maillard products to seed deterioration. Plant Physiol. 1991, 97, 165–169. [Google Scholar] [CrossRef]
- Priestley, D.A. Seed Aging: Implications for Seed Storage and Persistence in the Soil; Comstock Associates: Ithaca, NY, USA, 1986. [Google Scholar]
- Bartosz, G. Non-specific reactions: Molecular basis for ageing. J. Theor. Biol. 1981, 91, 233–235. [Google Scholar] [CrossRef]
- McDonald, M., Jr. Physical Seed Quality of Soybean. Seed Sci. Technol. 1985, 13, 601–628. [Google Scholar]
- Harrington, J.F. Germination of seeds from carrot, lettuce, and pepper plants grown under severe nutrient deficiencies. Hilgardia 1960, 30, 219–235. [Google Scholar] [CrossRef]
- Haferkamp, M.E.; Smith, L.; Nilan, R. Studies on Aged Seeds I Relation of Age of Seed to Germination and Longevity 1. Agron. J. 1953, 45, 434–437. [Google Scholar] [CrossRef]
- Justice, O.L.; Bass, L.N. Principles and Practices of Seed Storage; US Department of Agriculture, U.S. Gov. Printing Office: Washington, DC, USA, 1978.
- Kranner, I.; Minibayeva, F.V.; Beckett, R.P.; Seal, C.E. What is stress? Concepts, definitions and applications in seed science. New Phytol. 2010, 188, 655–673. [Google Scholar] [CrossRef]
- Davies, M.J. The oxidative environment and protein damage. Biochim. Et Biophys. Acta (BBA)-Proteins Proteom. 2005, 1703, 93–109. [Google Scholar] [CrossRef]
- Bailly, C.; El-Maarouf-Bouteau, H.; Corbineau, F. From intracellular signaling networks to cell death: The dual role of reactive oxygen species in seed physiology. Comptes Rendus Biol. 2008, 331, 806–814. [Google Scholar] [CrossRef]
- Rajjou, L.; Lovigny, Y.; Groot, S.P.; Belghazi, M.; Job, C.; Job, D. Proteome-wide characterization of seed aging in Arabidopsis: A comparison between artificial and natural aging protocols. Plant Physiol. 2008, 148, 620–641. [Google Scholar] [CrossRef]
- Almoguera, C.; Prieto-Dapena, P.; Díaz-Martín, J.; Espinosa, J.M.; Carranco, R.; Jordano, J. The HaDREB2 transcription factor enhances basal thermotolerance and longevity of seeds through functional interaction with HaHSFA9. BMC Plant Biol. 2009, 9, 75. [Google Scholar] [CrossRef]
- Kibinza, S.; Vinel, D.; Côme, D.; Bailly, C.; Corbineau, F. Sunflower seed deterioration as related to moisture content during ageing, energy metabolism and active oxygen species scavenging. Physiol. Plant. 2006, 128, 496–506. [Google Scholar] [CrossRef]
- Sivori, E.; Nakayama, F.; Cigliano, E. Germination of Achira seed (Canna sp.) approximately 550 years old. Nature 1968, 219, 1269–1270. [Google Scholar] [CrossRef]
- Shen-Miller, J.; Mudgett, M.B.; Schopf, J.W.; Clarke, S.; Berger, R. Exceptional seed longevity and robust growth: Ancient sacred lotus from China. Am. J. Bot. 1995, 82, 1367–1380. [Google Scholar] [CrossRef]
- Harrington, J.F.; Kozlowski, T. Seed storage and longevity. Seed Biol. 1972, 3, 145–245. [Google Scholar]
- Nagel, M.; Börner, A. The longevity of crop seeds stored under ambient conditions. Seed Sci. Res. 2010, 20, 1–12. [Google Scholar] [CrossRef]
- Copeland, L.O.; McDonald, M.F. Principles of Seed Science and Technology; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Roberts, E.; Ellis, R. Prediction of seed longevity at sub-zero temperatures and genetic resources conservation. Nature 1977, 268, 431–433. [Google Scholar] [CrossRef]
- Clerkx, E.J.; Vries, H.B.-D.; Ruys, G.J.; Groot, S.P.; Koornneef, M. Characterization of green seed, an enhancer of abi3-1 in Arabidopsis that affects seed longevity. Plant Physiol. 2003, 132, 1077–1084. [Google Scholar] [CrossRef] [PubMed]
- Contreras, S.; Bennett, M.A.; Metzger, J.D.; Tay, D.; Nerson, H. Red to far-red ratio during seed development affects lettuce seed germinability and longevity. HortScience 2009, 44, 130–134. [Google Scholar] [CrossRef]
- Contreras, S.; Bennett, M.A.; Metzger, J.D.; Tay, D. Maternal light environment during seed development affects lettuce seed weight, germinability, and storability. HortScience 2008, 43, 845–852. [Google Scholar] [CrossRef]
- Bewley, J.D.; Black, M. Seeds: Physiology of Development and Germination; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Arif, M.A.R. Seed Longevity and Dormancy in Wheat (Triticum aestivum L.)-Phenotypic Variation and Genetic Mapping. Ph.D. Thesis, Martin-Luther-Universität Halle-Wittenberg, Halle (Saale), Germany, November 2012. [Google Scholar]
- Fan, S. On “Fan Shengzhi Shu”: An Agriculturistic Book of China Written in the First Century BC; Peking Science Press: Beijing, China, 1963. [Google Scholar]
- Weiss, M.G.; Wentz, J.B. Effect of luteus genes on longevity of seed in maize. J. Am. Soc. Agron. 1937, 29, 63–75. [Google Scholar] [CrossRef]
- Lindstrom, E. Genetic relations of inbred lines of corn. Rep. Agrie. Res. Part 1943, 2, 41–45. [Google Scholar]
- Lindstrom, E. Inheritance of seed longevity in maize inbreds and hybrids. Genetics 1942, 27, 1943. [Google Scholar]
- RAO, A.P.; Fleming, A. Cytoplasmic–Genotypic Influences on Seed Viability in a Maize Inbred. Can. J. Plant Sci. 1979, 59, 241–242. [Google Scholar] [CrossRef]
- Haber, E. Longevity of the seed of sweet corn inbreds and hybrids. Proc. Am. Soc. Hortic.Sci. 1950, 55, 410–412. [Google Scholar]
- Scott, G.E. Improvement for Accelerated Aging Response of Seed in Maize Populations 1. Crop Sci. 1981, 21, 41–43. [Google Scholar] [CrossRef]
- Van der Mey, J.; Kilpatrick, R.; Smit, I. The germination of wheat and oat seed stored at Bethlehem, Republic of South Africa, 1956–1981. Cereal Res. Commun. 1982, 10, 159–163. [Google Scholar]
- Arif, M.A.R.; Nagel, M.; Lohwasser, U.; Börner, A. Genetic architecture of seed longevity in bread wheat (Triticum aestivum L.). J. Biosci. 2017, 42, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Mackay, D.; Tonkin, J. Investigations in crop seed longevity. I. An analysis of long-term experiments, with special reference to the influence of species, cultivar, provenance and season. J. Natl. Inst. Agric. Bot. 1967, 11, 209–225. [Google Scholar]
- Flood, R. Contribution of impermeable seed to longevity in Trifolium subterraneum (subterranean clover). Seed Sci. Technol. 1978, 6, 647–654. [Google Scholar]
- Van der Maesen, L. Seed storage, viability and rejuvenation. In Genetic Resources and Their Exploitation—Chickpeas, Faba Beans and Lentils; Springer: Berlin/Heidelberg, Germany, 1984; pp. 13–22. [Google Scholar]
- Starzinger, E.; EK, S.; SH, W. An observation on the relationship of soybean seed coat colour to viability maintenance. Seed Sci. Technol. 1982, 10, 301–305. [Google Scholar]
- Nakayama, R.; Saito, K. Diallel analysis of the longevity of seeds in kidney beans [Phaseolus vulgaris]. Bull. Fac. Agric. Hirosaki Univ. 1980, 34, 47–50. [Google Scholar]
- Kueneman, E. Genetic Control of Seed Longevity in Soybeans 1. Crop Sci. 1983, 23, 5–8. [Google Scholar] [CrossRef]
- Nagel, M.; Vogel, H.; Landjeva, S.; Buck-Sorlin, G.; Lohwasser, U.; Scholz, U.; Börner, A. Seed conservation in ex situ genebanks—Genetic studies on longevity in barley. Euphytica 2009, 170, 5–14. [Google Scholar] [CrossRef]
- Nagel, M.; Rosenhauer, M.; Willner, E.; Snowdon, R.J.; Friedt, W.; Börner, A. Seed longevity in oilseed rape (Brassica napus L.)–genetic variation and QTL mapping. Plant Genet. Resour. 2011, 9, 260–263. [Google Scholar] [CrossRef]
- Nagel, M.; Rehman-Arif, M.; Rosenhauer, M.; Börner, A. Longevity of seeds-intraspecific differences in the Gatersleben genebank collections. Tagungsband 2010, 60, 179–181. [Google Scholar]
- Bentsink, L.; Alonso-Blanco, C.; Vreugdenhil, D.; Tesnier, K.; Groot, S.P.; Koornneef, M. Genetic analysis of seed-soluble oligosaccharides in relation to seed storability of Arabidopsis. Plant Physiol. 2000, 124, 1595–1604. [Google Scholar] [CrossRef] [PubMed]
- de Souza Vidigal, D.; Willems, L.; van Arkel, J.; Dekkers, B.J.; Hilhorst, H.W.; Bentsink, L. Galactinol as marker for seed longevity. Plant Sci. 2016, 246, 112–118. [Google Scholar] [CrossRef]
- Clerkx, E.J.; Blankestijn-De Vries, H.; Ruys, G.J.; Groot, S.P.; Koornneef, M. Genetic differences in seed longevity of various Arabidopsis mutants. Physiol. Plant. 2004, 121, 448–461. [Google Scholar] [CrossRef]
- Sattler, S.E.; Gilliland, L.U.; Magallanes-Lundback, M.; Pollard, M.; DellaPenna, D. Vitamin E is essential for seed longevity and for preventing lipid peroxidation during germination. Plant Cell 2004, 16, 1419–1432. [Google Scholar] [CrossRef]
- Gerna, D.; Arc, E.; Holzknecht, M.; Roach, T.; Jansen-Dürr, P.; Weiss, A.K.; Kranner, I. AtFAHD1a: A New Player Influencing Seed Longevity and Dormancy in Arabidopsis? Int. J. Mol. Sci. 2021, 22, 2997. [Google Scholar] [CrossRef]
- Ogé, L.; Bourdais, G.; Bove, J.; Collet, B.; Godin, B.; Granier, F.; Boutin, J.-P.; Job, D.; Jullien, M.; Grappin, P. Protein repair L-isoaspartyl methyltransferase1 is involved in both seed longevity and germination vigor in Arabidopsis. Plant Cell 2008, 20, 3022–3037. [Google Scholar] [CrossRef]
- Hundertmark, M.; Buitink, J.; Leprince, O.; Hincha, D.K. The reduction of seed-specific dehydrins reduces seed longevity in Arabidopsis thaliana. Seed Sci. Res. 2011, 21, 165–173. [Google Scholar] [CrossRef]
- Nguyen, T.-P.; Keizer, P.; van Eeuwijk, F.; Smeekens, S.; Bentsink, L. Natural variation for seed longevity and seed dormancy are negatively correlated in Arabidopsis. Plant Physiol. 2012, 160, 2083–2092. [Google Scholar] [CrossRef] [PubMed]
- Bueso, E.; Ibañez, C.; Sayas, E.; Muñoz-Bertomeu, J.; Gonzalez-Guzmán, M.; Rodriguez, P.L.; Serrano, R. A forward genetic approach in Arabidopsis thaliana identifies a RING-type ubiquitin ligase as a novel determinant of seed longevity. Plant Sci. 2014, 215, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Bueso, E.; Muñoz-Bertomeu, J.; Campos, F.; Brunaud, V.; Martínez, L.; Sayas, E.; Ballester, P.; Yenush, L.; Serrano, R. ARABIDOPSIS THALIANA HOMEOBOX25 uncovers a role for Gibberellins in seed longevity. Plant Physiol. 2014, 164, 999–1010. [Google Scholar] [CrossRef] [PubMed]
- He, H.; de Souza Vidigal, D.; Snoek, L.B.; Schnabel, S.; Nijveen, H.; Hilhorst, H.; Bentsink, L. Interaction between parental environment and genotype affects plant and seed performance in Arabidopsis. J. Exp. Bot. 2014, 65, 6603–6615. [Google Scholar] [CrossRef]
- Mao, Z.; Sun, W. Arabidopsis seed-specific vacuolar aquaporins are involved in maintaining seed longevity under the control of ABSCISIC ACID INSENSITIVE 3. J. Exp. Bot. 2015, 66, 4781–4794. [Google Scholar] [CrossRef]
- Nguyen, T.-P.; Cueff, G.; Hegedus, D.D.; Rajjou, L.; Bentsink, L. A role for seed storage proteins in Arabidopsis seed longevity. J. Exp. Bot. 2015, 66, 6399–6413. [Google Scholar] [CrossRef]
- Waterworth, W.M.; Footitt, S.; Bray, C.M.; Finch-Savage, W.E.; West, C.E. DNA damage checkpoint kinase ATM regulates germination and maintains genome stability in seeds. Proc. Natl. Acad. Sci. USA 2016, 113, 9647–9652. [Google Scholar] [CrossRef]
- Chen, H.H.; Chu, P.; Zhou, Y.L.; Ding, Y.; Li, Y.; Liu, J.; Jiang, L.w.; Huang, S.Z. Ectopic expression of NnPER1, a Nelumbo nucifera 1-cysteine peroxiredoxin antioxidant, enhances seed longevity and stress tolerance in Arabidopsis. Plant J. 2016, 88, 608–619. [Google Scholar] [CrossRef]
- Sano, N.; Kim, J.-S.; Onda, Y.; Nomura, T.; Mochida, K.; Okamoto, M.; Seo, M. RNA-Seq using bulked recombinant inbred line populations uncovers the importance of brassinosteroid for seed longevity after priming treatments. Sci. Rep. 2017, 7, 8095. [Google Scholar] [CrossRef]
- Shen, W.; Yao, X.; Ye, T.; Ma, S.; Liu, X.; Yin, X.; Wu, Y. Arabidopsis aspartic protease ASPG1 affects seed dormancy, seed longevity and seed germination. Plant Cell Physiol. 2018, 59, 1415–1431. [Google Scholar] [CrossRef]
- Boussardon, C.; Martin-Magniette, M.-L.; Godin, B.; Benamar, A.; Vittrant, B.; Citerne, S.; Mary-Huard, T.; Macherel, D.; Rajjou, L.; Budar, F. Novel cytonuclear combinations modify Arabidopsis thaliana seed physiology and vigor. Front. Plant Sci. 2019, 10, 32. [Google Scholar] [CrossRef] [PubMed]
- Tsujimura, M.; Terachi, T. Cytoplasmic Genome. In The Allium Genomes; Springer: Berlin/Heidelberg, Germany, 2018; pp. 89–98. [Google Scholar]
- Yazdanpanah, F.; Maurino, V.G.; Mettler-Altmann, T.; Buijs, G.; Bailly, M.; Karimi Jashni, M.; Willems, L.; Sergeeva, L.I.; Rajjou, L.; Hilhorst, H.W. NADP-MALIC ENZYME 1 affects germination after seed storage in Arabidopsis thaliana. Plant Cell Physiol. 2019, 60, 318–328. [Google Scholar] [CrossRef] [PubMed]
- Durand, T.C.; Cueff, G.; Godin, B.; Valot, B.; Clément, G.; Gaude, T.; Rajjou, L. Combined proteomic and metabolomic profiling of the Arabidopsis thaliana vps29 mutant reveals pleiotropic functions of the retromer in seed development. Int. J. Mol. Sci. 2019, 20, 362. [Google Scholar] [CrossRef] [PubMed]
- Renard, J.; Niñoles, R.; Martínez-Almonacid, I.; Gayubas, B.; Mateos-Fernández, R.; Bissoli, G.; Bueso, E.; Serrano, R.; Gadea, J. Identification of novel seed longevity genes related to oxidative stress and seed coat by genome-wide association studies and reverse genetics. Plant Cell Environ. 2020, 43, 2523–2539. [Google Scholar] [CrossRef] [PubMed]
- Renard, J.; Martínez-Almonacid, I.; Sonntag, A.; Molina, I.; Moya-Cuevas, J.; Bissoli, G.; Muñoz-Bertomeu, J.; Faus, I.; Niñoles, R.; Shigeto, J. PRX2 and PRX25, peroxidases regulated by COG1, are involved in seed longevity in Arabidopsis. Plant Cell Environ. 2020, 43, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Renard, J.; Martínez-Almonacid, I.; Queralta Castillo, I.; Sonntag, A.; Hashim, A.; Bissoli, G.; Campos, L.; Muñoz-Bertomeu, J.; Niñoles, R.; Roach, T. Apoplastic lipid barriers regulated by conserved homeobox transcription factors extend seed longevity in multiple plant species. New Phytol. 2021, 231, 679–694. [Google Scholar] [CrossRef] [PubMed]
- Miura, K.; Lin, S.; Yano, M.; Nagamine, T. Mapping quantitative trait loci controlling seed longevity in rice (Oryza sativa L.). Theor. Appl. Genet. 2002, 104, 981–986. [Google Scholar] [CrossRef]
- Sasaki, K.; Fukuta, Y.; Sato, T. Mapping of quantitative trait loci controlling seed longevity of rice (Oryza sativa L.) after various periods of seed storage. Plant Breed. 2005, 124, 361–366. [Google Scholar] [CrossRef]
- Zeng, D.; Guo, L.; Xu, Y.; Yasukumi, K.; Zhu, L.; Qian, Q. QTL analysis of seed storability in rice. Plant Breed. 2006, 125, 57–60. [Google Scholar] [CrossRef]
- Shigemune, A.; Miura, K.; Sasahara, H.; Goto, A.; Yoshida, T. Role of maternal tissues in qLG-9 control of seed longevity in rice (Oryza sativa L.). Breed. Sci. 2008, 58, 1–5. [Google Scholar] [CrossRef][Green Version]
- Xue, Y.; Zhang, S.; Yao, Q.; Peng, R.; Xiong, A.; Li, X.; Zhu, W.; Zhu, Y.; Zha, D. Identification of quantitative trait loci for seed storability in rice (Oryza sativa L.). Euphytica 2008, 164, 739–744. [Google Scholar] [CrossRef]
- Sasaki, K.; Takeuchi, Y.; Miura, K.; Yamaguchi, T.; Ando, T.; Ebitani, T.; Higashitani, A.; Yamaya, T.; Yano, M.; Sato, T. Fine mapping of a major quantitative trait locus, qLG-9, that controls seed longevity in rice (Oryza sativa L.). Theor. Appl. Genet. 2015, 128, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.-H.; Kim, S.-R.; An, G. Rice Aldehyde Dehydrogenase7 Is Needed for Seed Maturation and Viability. Plant Physiol. 2008, 149, 905–915. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Lee, J.; Jin, Y.-M.; Qiao, Y.; Piao, R.; Jang, S.M.; Woo, M.-O.; Kwon, S.-W.; Liu, X.; Pan, H.-Y. Identification of QTLs for seed germination capability after various storage periods using two RIL populations in rice. Mol. Cells 2011, 31, 385–392. [Google Scholar] [CrossRef]
- Li, L.; Lin, Q.; Liu, S.; Liu, X.; Wang, W.; Hang, N.T.; Liu, F.; Zhao, Z.; Jiang, L.; Wan, J. Identification of quantitative trait loci for seed storability in rice (Oryza sativa L.). Plant Breed. 2012, 131, 739–743. [Google Scholar] [CrossRef]
- Long, Q.; Zhang, W.; Wang, P.; Shen, W.; Zhou, T.; Liu, N.; Wang, R.; Jiang, L.; Huang, J.; Wang, Y. Molecular genetic characterization of rice seed lipoxygenase 3 and assessment of its effects on seed longevity. J. Plant Biol. 2013, 56, 232–242. [Google Scholar] [CrossRef]
- Xu, H.; Wei, Y.; Zhu, Y.; Lian, L.; Xie, H.; Cai, Q.; Chen, Q.; Lin, Z.; Wang, Z.; Xie, H. Antisense suppression of LOX3 gene expression in rice endosperm enhances seed longevity. Plant Biotechnol. J. 2015, 13, 526–539. [Google Scholar] [CrossRef]
- Huang, J.; Cai, M.; Long, Q.; Liu, L.; Lin, Q.; Jiang, L.; Chen, S.; Wan, J. OsLOX2, a rice type I lipoxygenase, confers opposite effects on seed germination and longevity. Transgenic Res. 2014, 23, 643–655. [Google Scholar] [CrossRef]
- Hang, N.T.; Lin, Q.; Liu, L.; Liu, X.; Liu, S.; Wang, W.; Li, L.; He, N.; Liu, Z.; Jiang, L. Mapping QTLs related to rice seed storability under natural and artificial aging storage conditions. Euphytica 2015, 203, 673–681. [Google Scholar] [CrossRef]
- Wei, Y.; Xu, H.; Diao, L.; Zhu, Y.; Xie, H.; Cai, Q.; Wu, F.; Wang, Z.; Zhang, J.; Xie, H. Protein repair L-isoaspartyl methyltransferase 1 (PIMT1) in rice improves seed longevity by preserving embryo vigor and viability. Plant Mol. Biol. 2015, 89, 475–492. [Google Scholar] [CrossRef]
- Li, G.; Na, Y.-W.; Kwon, S.-W.; Park, Y.-J. Association analysis of seed longevity in rice under conventional and high-temperature germination conditions. Plant Syst. Evol. 2014, 300, 389–402. [Google Scholar] [CrossRef]
- Lee, J.-S.; Kwak, J.; Yoon, M.-R.; Lee, J.-S.; Hay, F.R. Contrasting tocol ratios associated with seed longevity in rice variety groups. Seed Sci. Res. 2017, 27, 273–280. [Google Scholar] [CrossRef]
- Lee, J.-S.; Kwak, J.; Cho, J.-H.; Chebotarov, D.; Yoon, M.-R.; Lee, J.-S.; Hamilton, N.R.S.; Hay, F.R. A high proportion of beta-tocopherol in vitamin E is associated with poor seed longevity in rice produced under temperate conditions. Plant Genet. Resour. 2019, 17, 375–378. [Google Scholar] [CrossRef]
- Nisarga, K.N.; Vemanna, R.S.; Chandrashekar, B.K.; Rao, H.; Vennapusa, A.R.; Narasimaha, A.; Makarla, U.; Basavaiah, M.R. Aldo-ketoreductase 1 (AKR1) improves seed longevity in tobacco and rice by detoxifying reactive cytotoxic compounds generated during ageing. Rice 2017, 10, 11. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, S.; Wang, L.; Wu, D.; Cheng, H.; Du, X.; Mao, D.; Zhang, C.; Jiang, X. miR164c and miR168a regulate seed vigor in rice. J. Integr. Plant Biol. 2020, 62, 470–486. [Google Scholar] [CrossRef]
- Liu, W.; Pan, X.; Li, Y.; Duan, Y.; Min, J.; Liu, S.; Sheng, X.; Li, X. Detection and validation of QTL s associated with seed longevity in rice (Oryza sativa L.). Plant Breed. 2018, 137, 546–552. [Google Scholar] [CrossRef]
- Yan, S.; Huang, W.; Gao, J.; Fu, H.; Liu, J. Comparative metabolomic analysis of seed metabolites associated with seed storability in rice (Oryza sativa L.) during natural aging. Plant Physiol. Biochem. 2018, 127, 590–598. [Google Scholar] [CrossRef]
- Lee, J.-S.; Velasco-Punzalan, M.; Pacleb, M.; Valdez, R.; Kretzschmar, T.; McNally, K.L.; Ismail, A.M.; Cruz, P.C.S.; Sackville Hamilton, N.R.; Hay, F.R. Variation in seed longevity among diverse Indica rice varieties. Ann. Bot. 2019, 124, 447–460. [Google Scholar] [CrossRef]
- Raquid, R.; Kohli, A.; Reinke, R.; Dionisio-Sese, M.; Kwak, J.; Chebotarov, D.; Mo, Y.; Lee, J.-S. Genetic factors enhancing seed longevity in tropical japonica rice. Curr. Plant Biol. 2021, 26, 100196. [Google Scholar] [CrossRef]
- Wu, F.; Luo, X.; Wang, L.; Wei, Y.; Li, J.; Xie, H.; Zhang, J.; Xie, G. Genome-Wide Association Study Reveals the QTLs for Seed Storability in World Rice Core Collections. Plants 2021, 10, 812. [Google Scholar] [CrossRef]
- Zhao, M.; Hu, B.; Fan, Y.; Ding, G.; Yang, W.; Chen, Y.; Chen, Y.; Xie, J.; Zhang, F. Identification, Analysis, and Confirmation of Seed Storability-Related Loci in Dongxiang Wild Rice (Oryza rufipogon Griff.). Genes 2021, 12, 1831. [Google Scholar] [CrossRef] [PubMed]
- Lundqvist, U. New and revised descriptions of barley genes. Barley Genet. Newslett. 1997, 26, 22–516. [Google Scholar]
- Sun, S.; Yu, J.-P.; Chen, F.; Zhao, T.-J.; Fang, X.-H.; Li, Y.-Q.; Sui, S.-F. TINY, a dehydration-responsive element (DRE)-binding protein-like transcription factor connecting the DRE-and ethylene-responsive element-mediated signaling pathways in Arabidopsis. J. Biol. Chem. 2008, 283, 6261–6271. [Google Scholar] [CrossRef] [PubMed]
- Zareie, R.; Melanson, D.L.; Murphy, P.J. Isolation of fungal cell wall degrading proteins from barley (Hordeum vulgare L.) leaves infected with Rhynchosporium secalis. Mol. Plant-Microbe Interact. 2002, 15, 1031–1039. [Google Scholar] [CrossRef] [PubMed]
- Nagel, M.; Kranner, I.; Neumann, K.; Rolletschek, H.; Seal, C.E.; Colville, L.; Fernández-Marín, B.; Börner, A. Genome-wide association mapping and biochemical markers reveal that seed ageing and longevity are intricately affected by genetic background and developmental and environmental conditions in barley. Plant Cell Environ. 2015, 38, 1011–1022. [Google Scholar] [CrossRef] [PubMed]
- Wozny, D.; Kramer, K.; Finkemeier, I.; Acosta, I.F.; Koornneef, M. Genes for seed longevity in barley identified by genomic analysis on near isogenic lines. Plant Cell Environ. 2018, 41, 1895–1911. [Google Scholar] [CrossRef] [PubMed]
- Puchta, M.; Groszyk, J.; Małecka, M.; Koter, M.D.; Niedzielski, M.; Rakoczy-Trojanowska, M.; Boczkowska, M. Barley Seeds miRNome Stability during Long-Term Storage and Aging. Int. J. Mol. Sci. 2021, 22, 4315. [Google Scholar] [CrossRef]
- Landjeva, S.; Lohwasser, U.; Börner, A. Genetic mapping within the wheat D genome reveals QTL for germination, seed vigour and longevity, and early seedling growth. Euphytica 2010, 171, 129. [Google Scholar] [CrossRef]
- Arif, M.R.; Nagel, M.; Neumann, K.; Kobiljski, B.; Lohwasser, U.; Börner, A. Genetic studies of seed longevity in hexaploid wheat using segregation and association mapping approaches. Euphytica 2012, 186, 1–13. [Google Scholar] [CrossRef]
- Li, W.; Faris, J.; Chittoor, J.; Leach, J.; Hulbert, S.; Liu, D.; Chen, P.; Gill, B. Genomic mapping of defense response genes in wheat. Theor. Appl. Genet. 1999, 98, 226–233. [Google Scholar] [CrossRef]
- Quarrie, S.; Steed, A.; Calestani, C.; Semikhodskii, A.; Lebreton, C.; Chinoy, C.; Steele, N.; Pljevljakusić, D.; Waterman, E.; Weyen, J. A high-density genetic map of hexaploid wheat (Triticum aestivum L.) from the cross Chinese Spring× SQ1 and its use to compare QTLs for grain yield across a range of environments. Theor. Appl. Genet. 2005, 110, 865–880. [Google Scholar] [CrossRef] [PubMed]
- Sourdille, P.; Tixier, M.; Charmet, G.; Gay, G.; Cadalen, T.; Bernard, S.; Bernard, M. Location of genes involved in ear compactness in wheat (Triticum aestivum) by means of molecular markers. Mol. Breed. 2000, 6, 247–255. [Google Scholar] [CrossRef]
- Agacka-Mołdoch, M.; Arif, M.A.R.; Lohwasser, U.; Doroszewska, T.; Qualset, C.O.; Börner, A. The inheritance of wheat grain longevity: A comparison between induced and natural ageing. J. Appl. Genet. 2016, 57, 477–481. [Google Scholar] [CrossRef] [PubMed]
- Arif, M.A.R.; Shokat, S.; Plieske, J.; Ganal, M.; Lohwasser, U.; Chesnokov, Y.V.; Kocherina, N.V.; Kulwal, P.; Kumar, N.; McGuire, P.E. A SNP-based genetic dissection of versatile traits in bread wheat (Triticum aestivum L.). Plant J. 2021, 108, 960–976. [Google Scholar] [CrossRef]
- Arif, M.A.R.; Agacka-Mołdoch, M.; Qualset, C.O.; Börner, A. Mapping of additive and epistatic QTLs linked to seed longevity in bread wheat (Triticum aestivum L.). Cereal Res. Commun 2022. Available online: https://link.springer.com/article/10.1007/s42976-021-00240-3 (accessed on 12 January 2022). [CrossRef]
- Meng, L.; Li, H.; Zhang, L.; Wang, J. QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop J. 2015, 3, 269–283. [Google Scholar] [CrossRef]
- Kolodziej, M.C.; Singla, J.; Sánchez-Martín, J.; Zbinden, H.; Šimková, H.; Karafiátová, M.; Doležel, J.; Gronnier, J.; Poretti, M.; Glauser, G. A membrane-bound ankyrin repeat protein confers race-specific leaf rust disease resistance in wheat. Nat. Commun. 2021, 12, 956. [Google Scholar] [CrossRef]
- Yan, J.; Yao, Y.; Hong, S.; Yang, Y.; Shen, C.; Zhang, Q.; Zhang, D.; Zou, T.; Yin, P. Delineation of pentatricopeptide repeat codes for target RNA prediction. Nucleic Acids Res. 2019, 47, 3728–3738. [Google Scholar] [CrossRef]
- Figueiredo, J.; Sousa Silva, M.; Figueiredo, A. Subtilisin-like proteases in plant defence: The past, the present and beyond. Mol. Plant Pathol. 2018, 19, 1017–1028. [Google Scholar] [CrossRef]
- Reinbothe, C.; Springer, A.; Samol, I.; Reinbothe, S. Plant oxylipins: Role of jasmonic acid during programmed cell death, defence and leaf senescence. FEBS J. 2009, 276, 4666–4681. [Google Scholar] [CrossRef]
- Ohmiya, A. Carotenoid cleavage dioxygenases and their apocarotenoid products in plants. Plant Biotechnol. 2009, 26, 351–358. [Google Scholar] [CrossRef]
- Saibi, W.; Zouari, N.; Masmoudi, K.; Brini, F. Role of the durum wheat dehydrin in the function of proteases conferring salinity tolerance in Arabidopsis thaliana transgenic lines. Int. J. Biol. Macromol. 2016, 85, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Sgarbi, C.; Malbrán, I.; Saldúa, L.; Lori, G.A.; Lohwasser, U.; Arif, M.A.R.; Börner, A.; Yanniccari, M.; Castro, A.M. Mapping Resistance to Argentinean Fusarium (Graminearum) Head Blight Isolates in Wheat. Int. J. Mol. Sci. 2021, 22, 13653. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.; Liu, J.; Gao, F.; Yin, G.; Wang, Z.; Chen, F.; Li, X.; Xu, J.; Chen, T.; Li, L. Genome-wide linkage mapping reveals qtls for seed vigor-related traits under artificial aging in common wheat (Triticum aestivum ). Front. Plant Sci. 2018, 9, 1101. [Google Scholar] [CrossRef]
- Zuo, J.H.; Chen, F.Y.; Li, X.Y.; Xia, X.C.; Cao, H.; Liu, J.D.; Liu, Y.X. Genome-wide association study reveals loci associated with seed longevity in common wheat (Triticum aestivum L.). Plant Breed. 2020, 139, 295–303. [Google Scholar] [CrossRef]
- Wang, S.; Wong, D.; Forrest, K.; Allen, A.; Chao, S.; Huang, B.E.; Maccaferri, M.; Salvi, S.; Milner, S.G.; Cattivelli, L. Characterization of polyploid wheat genomic diversity using a high-density 90 000 single nucleotide polymorphism array. Plant Biotechnol. J. 2014, 12, 787–796. [Google Scholar] [CrossRef]
- Wu, X.; Ning, F.; Hu, X.; Wang, W. Genetic modification for improving seed vigor is transitioning from model plants to crop plants. Front. Plant Sci. 2017, 8, 8. [Google Scholar] [CrossRef]
- Shi, H.; Guan, W.; Shi, Y.; Wang, S.; Fan, H.; Yang, J.; Chen, W.; Zhang, W.; Sun, D.; Jing, R. QTL mapping and candidate gene analysis of seed vigor-related traits during artificial aging in wheat (Triticum aestivum). Sci. Rep. 2020, 10, 22060. [Google Scholar] [CrossRef]
- Revilla, P.; Velasco, P.; Malvar, R.A.; Cartea, M.E.; Ordás, A. Variability among maize (Zea mays L.) inbred lines for seed longevity. Genet. Resour. Crop Evol. 2006, 53, 771–777. [Google Scholar] [CrossRef][Green Version]
- Revilla, P.; Butrón, A.; Rodríguez, V.; Malvar, R.; Ordás, A. Identification of genes related to germination in aged maize seed by screening natural variability. J. Exp. Bot. 2009, 60, 4151–4157. [Google Scholar] [CrossRef]
- Liu, J.-B.; Fu, Z.-Y.; Xie, H.-L.; Hu, Y.-M.; Liu, Z.-H.; Duan, L.-J.; Xu, S.-Z.; Tang, J.-H. Identification of QTLs for maize seed vigor at three stages of seed maturity using a RIL population. Euphytica 2011, 178, 127–135. [Google Scholar] [CrossRef]
- Han, Z.; Ku, L.; Zhang, Z.; Zhang, J.; Guo, S.; Liu, H.; Zhao, R.; Ren, Z.; Zhang, L.; Su, H. QTLs for seed vigor-related traits identified in maize seeds germinated under artificial aging conditions. PLoS ONE 2014, 9, e92535. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, Z.; Fu, Z.; Liu, Z.; Hu, Y.; Tang, J. Comparative QTL analysis of maize seed artificial aging between an immortalized F2 population and its corresponding RILs. Crop J. 2016, 4, 30–39. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, H.; Li, X.; Wang, F.; Lyle, D.; Sun, L.; Wang, G.; Wang, J.; Li, L.; Gu, R. Quantitative trait locus mapping for seed artificial aging traits using an F2: 3 population and a recombinant inbred line population crossed from two highly related maize inbreds. Plant Breed. 2019, 138, 29–37. [Google Scholar]
- Li, L.; Wang, F.; Li, X.; Peng, Y.; Zhang, H.; Hey, S.; Wang, G.; Wang, J.; Gu, R. Comparative analysis of the accelerated aged seed transcriptome profiles of two maize chromosome segment substitution lines. PLoS ONE 2019, 14, e0216977. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Chen, K.; Yan, D.; Hao, G.; Qi, J.; Wang, C.; Dirk, L.M.; Bruce Downie, A.; Gong, J.; Wang, J. ZmDREB2A regulates ZmGH3. 2 and ZmRAFS, shifting metabolism towards seed aging tolerance over seedling growth. Plant J. 2020, 104, 268–282. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Sun, X.; Liu, S.; Gong, C.; Feng, C.; Han, X.; Lv, T.; Zhou, Y.; Wang, Z.; Di, H. Screening and application of SSR markers related to seed storability traits in maize (Zea mays L.). Genet. Resour. Crop Evol. 2021, 68, 2521–2535. [Google Scholar] [CrossRef]
- Singh, R.; Raipuria, R.; Bhatia, V.; Rani, A.; Husain, S.; Chauhan, D.; Chauhan, G.; Mohapatra, T. SSR markers associated with seed longevity in soybean. Seed Sci. Technol. 2008, 36, 162–167. [Google Scholar] [CrossRef]
- Cregan, P.; Jarvik, T.; Bush, A.; Shoemaker, R.; Lark, K.; Kahler, A.; Kaya, N.; VanToai, T.; Lohnes, D.; Chung, J. An integrated genetic linkage map of the soybean genome. Crop Sci. 1999, 39, 1464–1490. [Google Scholar] [CrossRef]
- Singh, R.; Raipuria, R.; Bhatia, V.; Rani, A.; Husain, S.; Satyavathi, C.T.; Chauhan, G.; Mohapatra, T. Identification of SSR markers associated with seed coat permeability and electrolyte leaching in soybean. Physiol. Mol. Biol. Plants 2008, 14, 173–177. [Google Scholar] [CrossRef]
- Naik, S.; Madhusudan, K.; Motagi, B.; Mugali, S.; Nadaf, H. Molecular characterization of seed longevity and associated characters using SSR markers in soybean [Glycine max (L.) Merrill]. J. Pharmacogn. Phytochem. 2019, 8, 2357–2360. [Google Scholar]
- Hosamani, J.; Kumar, M.A.; Talukdar, A.; Lal, S.; Dadlani, M. Molecular characterization and identification of candidate markers for seed longevity in soybean [Glycine max (L.) Merill]. Indian J. Genet. 2013, 73, 64–71. [Google Scholar] [CrossRef]
- Dargahi, H.; Tanya, P.; Srinives, P. Mapping of the genomic regions controlling seed storability in soybean (Glycine max L.). J. Genet. 2014, 93, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Pereira Lima, J.J.; Buitink, J.; Lalanne, D.; Rossi, R.F.; Pelletier, S.; Da Silva, E.A.A.; Leprince, O. Molecular characterization of the acquisition of longevity during seed maturation in soybean. PLoS ONE 2017, 12, e0180282. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Hina, A.; Song, S.; Kong, J.; Bhat, J.A.; Zhao, T. Whole-genome mapping identified novel “QTL hotspots regions” for seed storability in soybean (Glycine max L.). BMC Genom. 2019, 20, 499. [Google Scholar] [CrossRef]
- Pawar, P.; Naik, R.; Deshmukh, M.; Satbhai, R.; Mohite, S. Biochemical and molecuar marker based screening of seed longevity in soybean [Glycine max (L.) Merill]. Legume Res. Int. J. 2019, 42, 575–584. [Google Scholar] [CrossRef]
- Wang, R.; Wu, F.; Xie, X.; Yang, C. Quantitative Trait Locus Mapping of Seed Vigor in Soybean under −20 °C Storage and Accelerated Aging Conditions via RAD Sequencing. Curr. Issues Mol. Biol. 2021, 43, 1977–1996. [Google Scholar] [CrossRef]
- Schwember, A.R.; Bradford, K.J. Quantitative trait loci associated with longevity of lettuce seeds under conventional and controlled deterioration storage conditions. J. Exp. Bot. 2010, 61, 4423–4436. [Google Scholar] [CrossRef]
- Agacka-Mołdoch, M.; Nagel, M.; Doroszewska, T.; Lewis, R.; Börner, A. Mapping quantitative trait loci determining seed longevity in tobacco (Nicotiana tabacum L.). Euphytica 2015, 202, 479–486. [Google Scholar] [CrossRef]
- Xiao, B.; Drake, K.; Vontimitta, V.; Tong, Z.; Zhang, X.; Li, M.Y.; Leng, X.D.; Li, Y.; Lewis, R.S. Location of genomic regions contributing to Phytophthora nicotianae resistance in tobacco cultivar Florida 301. Crop Sci. 2013, 53, 473–481. [Google Scholar] [CrossRef]
- Agacka-Mołdoch, M.; Arif, M.A.R.; Lohwasser, U.; Doroszewska, T.; Lewis, R.S.; Börner, A. QTL analysis of seed germination traits in tobacco (Nicotiana tabacum L.). J. Appl. Genet. 2021, 62, 441–444. [Google Scholar] [CrossRef] [PubMed]
- Barbosa Batista, T.; Javier Fernandez, G.; Alexandre da Silva, T.; Maia, J.; Amaral da Silva, E.A. Transcriptome analysis in osmo-primed tomato seeds with enhanced longevity by heat shock treatment. AoB Plants 2020, 12, plaa041. [Google Scholar] [CrossRef] [PubMed]
- Bizouerne, E.; Buitink, J.; Vu, B.L.; Vu, J.L.; Esteban, E.; Pasha, A.; Provart, N.; Verdier, J.; Leprince, O. Gene co-expression analysis of tomato seed maturation reveals tissue-specific regulatory networks and hubs associated with the acquisition of desiccation tolerance and seed vigour. BMC Plant Biol. 2021, 21, 124. [Google Scholar] [CrossRef] [PubMed]
- Graner, A.; Börner, A. Quest for seed immortality is mission impossible. Nature 2006, 442, 353. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Gore, M.; Buckler, E.S.; Yu, J. Status and prospects of association mapping in plants. Plant Genome 2008, 1, 5–20. [Google Scholar] [CrossRef]
- Jannink, J.-L.; Bink, M.C.; Jansen, R.C. Using complex plant pedigrees to map valuable genes. Trends Plant Sci. 2001, 6, 337–342. [Google Scholar] [CrossRef]
- Röder, M.S.; Korzun, V.; Wendehake, K.; Plaschke, J.; Tixier, M.-H.; Leroy, P.; Ganal, M.W. A microsatellite map of wheat. Genetics 1998, 149, 2007–2023. [Google Scholar] [CrossRef]
- Arif, M.R.; Neumann, K.; Nagel, M.; Kobiljski, B.; Lohwasser, U.; Börner, A. An association mapping analysis of dormancy and pre-harvest sprouting in wheat. Euphytica 2012, 188, 409–417. [Google Scholar] [CrossRef]
- Joosen, R.V.; Kodde, J.; Willems, L.A.; Ligterink, W.; van der Plas, L.H.; Hilhorst, H.W. GERMINATOR: A software package for high-throughput scoring and curve fitting of Arabidopsis seed germination. Plant J. 2010, 62, 148–159. [Google Scholar] [CrossRef]
- Nelson, J.C. QGENE: Software for marker-based genomic analysis and breeding. Mol. Breed. 1997, 3, 239–245. [Google Scholar] [CrossRef]
- Wang, S.; Basten, C.; Zeng, Z. Windows QTL Cartographer 2.5; Raleigh, N.C., Ed.; Department of Statistics, North Carolina State University: Raleigh, NC, USA, 2012; Available online: https://brcwebportal.cos.ncsu.edu/qtlcart/WQTLCart.htm (accessed on 23 November 2021).
- Zhang, Y.-W.; Wen, Y.-J.; Dunwell, J.M.; Zhang, Y.-M. QTL. gCIMapping. GUI v2. 0: An R software for detecting small-effect and linked QTLs for quantitative traits in bi-parental segregation populations. Comput. Struct. Biotechnol. J. 2020, 18, 59–65. [Google Scholar] [CrossRef]
- Singh, S.; Sehgal, D.; Kumar, S.; Arif, M.; Vikram, P.; Sansaloni, C.; Fuentes-Dávila, G.; Ortiz, C. GWAS revealed a novel resistance locus on chromosome 4D for the quarantine disease Karnal bunt in diverse wheat pre-breeding germplasm. Sci. Rep. 2020, 10, 5999. [Google Scholar] [CrossRef] [PubMed]
- Sehgal, D.; Autrique, E.; Singh, R.; Ellis, M.; Singh, S.; Dreisigacker, S. Identification of genomic regions for grain yield and yield stability and their epistatic interactions. Sci. Rep. 2017, 7, 41578. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Jighly, A.; Sehgal, D.; Burgueño, J.; Joukhadar, R.; Singh, S.; Sharma, A.; Vikram, P.; Sansaloni, C.; Govindan, V. Direct introgression of untapped diversity into elite wheat lines. Nat. Food 2021, 2, 819–827. [Google Scholar] [CrossRef]
- Singh, S.; Vikram, P.; Sehgal, D.; Burgueño, J.; Sharma, A.; Singh, S.K.; Sansaloni, C.P.; Joynson, R.; Brabbs, T.; Ortiz, C. Harnessing genetic potential of wheat germplasm banks through impact-oriented-prebreeding for future food and nutritional security. Sci. Rep. 2018, 8, 12527. [Google Scholar] [CrossRef]
- Kidane, Y.G.; Gesesse, C.A.; Hailemariam, B.N.; Desta, E.A.; Mengistu, D.K.; Fadda, C.; Pè, M.E.; Dell’Acqua, M. A large nested association mapping population for breeding and quantitative trait locus mapping in Ethiopian durum wheat. Plant Biotechnol. J. 2019, 17, 1380–1393. [Google Scholar] [CrossRef]
- Saade, S.; Maurer, A.; Shahid, M.; Oakey, H.; Schmöckel, S.M.; Negrão, S.; Pillen, K.; Tester, M. Yield-related salinity tolerance traits identified in a nested association mapping (NAM) population of wild barley. Sci. Rep. 2016, 6, 32586. [Google Scholar] [CrossRef]
- Morris, G.P.; Ramu, P.; Deshpande, S.P.; Hash, C.T.; Shah, T.; Upadhyaya, H.D.; Riera-Lizarazu, O.; Brown, P.J.; Acharya, C.B.; Mitchell, S.E. Population genomic and genome-wide association studies of agroclimatic traits in sorghum. Proc. Natl. Acad. Sci. USA 2013, 110, 453–458. [Google Scholar] [CrossRef]
- McCouch, S.R.; Wright, M.H.; Tung, C.-W.; Maron, L.G.; McNally, K.L.; Fitzgerald, M.; Singh, N.; DeClerck, G.; Agosto-Perez, F.; Korniliev, P. Open access resources for genome-wide association mapping in rice. Nat. Commun. 2016, 7, 10532. [Google Scholar] [CrossRef]
- Zhao, J.; Sauvage, C.; Zhao, J.; Bitton, F.; Bauchet, G.; Liu, D.; Huang, S.; Tieman, D.M.; Klee, H.J.; Causse, M. Meta-analysis of genome-wide association studies provides insights into genetic control of tomato flavor. Nat. Commun. 2019, 10, 1534. [Google Scholar] [CrossRef]
- Tripodi, P.; Rabanus-Wallace, M.T.; Barchi, L.; Kale, S.; Esposito, S.; Acquadro, A.; Schafleitner, R.; van Zonneveld, M.; Prohens, J.; Diez, M.J. Global range expansion history of pepper (Capsicum spp.) revealed by over 10,000 genebank accessions. Proc. Natl. Acad. Sci. USA 2021, 118, e2104315118. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-H.; Reif, J.C.; Hong, H.-L.; Li, H.-H.; Liu, Z.-X.; Ma, Y.-S.; Li, J.; Tian, Y.; Li, Y.-F.; Li, W.-B. Genome-wide association mapping of QTL underlying seed oil and protein contents of a diverse panel of soybean accessions. Plant Sci. 2018, 266, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Do, T.D.; Vuong, T.D.; Dunn, D.; Clubb, M.; Valliyodan, B.; Patil, G.; Chen, P.; Xu, D.; Nguyen, H.T.; Shannon, J.G. Identification of new loci for salt tolerance in soybean by high-resolution genome-wide association mapping. BMC Genom. 2019, 20, 318. [Google Scholar] [CrossRef] [PubMed]
- Bhatta, B.P.; Malla, S. Improving horticultural crops via CRISPR/Cas9: Current successes and prospects. Plants 2020, 9, 1360. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.; Vaidya, E.; Rani, R.; Yadav, N.K.; Singh, B.; Rai, P.; Singh, D. Recent perspective of next generation sequencing: Applications in molecular plant biology and crop improvement. Proc. Natl. Acad.Sci. India Sect. B Biol. Sci. 2018, 88, 435–449. [Google Scholar] [CrossRef]
- Duarte-Delgado, D.; Dadshani, S.; Schoof, H.; Oyiga, B.C.; Schneider, M.; Mathew, B.; Léon, J.; Ballvora, A. Transcriptome profiling at osmotic and ionic phases of salt stress response in bread wheat uncovers trait-specific candidate genes. BMC Plant Biol. 2020, 20, 428. [Google Scholar] [CrossRef]
- Hrdlickova, R.; Toloue, M.; Tian, B. RNA-Seq methods for transcriptome analysis. Wiley Interdiscip.Rev. RNA 2017, 8, e1364. [Google Scholar] [CrossRef]
- Asmann, Y.W.; Klee, E.W.; Thompson, E.A.; Perez, E.A.; Middha, S.; Oberg, A.L.; Therneau, T.M.; Smith, D.I.; Poland, G.A.; Wieben, E.D. 3’tag digital gene expression profiling of human brain and universal reference RNA using Illumina Genome Analyzer. BMC Genom. 2009, 10, 531. [Google Scholar] [CrossRef]


{kind=link}
| Sr. No. | Plant Species | Number of Accessions | Genotyping Platform | References |
|---|---|---|---|---|
| 1 | Hexaploid wheat (Triticum aestivum L.) | >2500 accession in SeeDs of Discovery project | 7180 genotyping by sequencing (GBS) SNPs | [167,168] |
| 2 | Durum wheat (Triticum durum Desf.) |
|
| |
| 3 | Sorghum (Sorghum bicolor (L.) Moench.) | 971 world wide accessions | GBS SNPs | [171] |
| 4 | Rice (Oryza sativa L.) | 1568 inbred varieties | 700,000 high density rice array SNPs | [172] |
| 5 | Tomato (Solanum lycopersicum L.) | 163 accesions, 291 accesions and 402 accesions | 5995 SNPs, 9013 SNPs and 2014,488 SNPs, respectively | [173] |
| 6 | Pepper (Capsicum spp.) | 10,038 genebank accession | GBS SNPs | [174] |
| 7 | Soybean (Glycine max L.) |
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arif, M.A.R.; Afzal, I.; Börner, A. Genetic Aspects and Molecular Causes of Seed Longevity in Plants—A Review. Plants 2022, 11, 598. https://doi.org/10.3390/plants11050598
Arif MAR, Afzal I, Börner A. Genetic Aspects and Molecular Causes of Seed Longevity in Plants—A Review. Plants. 2022; 11(5):598. https://doi.org/10.3390/plants11050598
Chicago/Turabian StyleArif, Mian Abdur Rehman, Irfan Afzal, and Andreas Börner. 2022. "Genetic Aspects and Molecular Causes of Seed Longevity in Plants—A Review" Plants 11, no. 5: 598. https://doi.org/10.3390/plants11050598
APA StyleArif, M. A. R., Afzal, I., & Börner, A. (2022). Genetic Aspects and Molecular Causes of Seed Longevity in Plants—A Review. Plants, 11(5), 598. https://doi.org/10.3390/plants11050598