
LC–DAD–MS Phenolic Characterisation of Six Invasive Plant Species in Croatia and Determination of Their Antimicrobial and Cytotoxic Activity

 ,
,  ,
,  ,
, 
 , , , and
, , , and 
Abstract
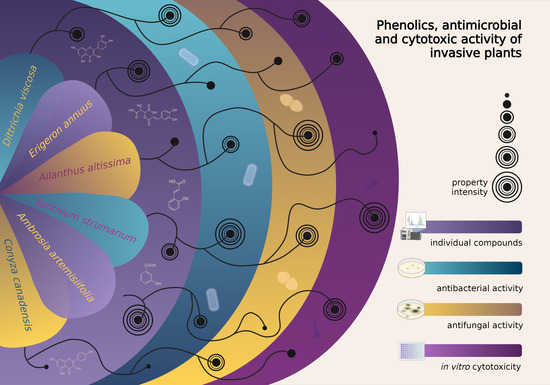
:
1. Introduction
2. Results and Discussion
2.1. Extraction Yield and Phenolic Concentration
2.2. LC–DAD–MS Identification and Quantification of Phenolic Compounds
2.3. Antimicrobial Activity and Bioautography
3. Materials and Methods
3.1. Plant Material
3.2. Extraction Procedures
3.3. Determination of Individual Phenolic Compounds Using LC–DAD–MS Analysis
3.4. Antimicrobial Testing
3.4.1. Bioautography
3.4.2. Antibacterial and Antifungal Activity
3.5. Cytotoxicity
3.6. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bennett, A.E.; Thomsen, M.; Strauss, S.Y. Multiple mechanisms enable invasive species to suppress native species. Am. J. Bot. 2011, 98, 1086–1094. [Google Scholar] [CrossRef] [PubMed]
- Lind, E.M.; Parker, J.D. Novel weapons testing: Are invasive pants more chemically defended than native plants? PLoS ONE 2010, 5, e10429. [Google Scholar] [CrossRef] [PubMed]
- Macel, M.; de Vos, R.C.H.; Jansen, J.J.; van der Putten, W.H.; van Dam, N.M. Novel chemistry of invasive plants: Exotic species have more unique metabolomic profiles than native congeners. Ecol. Evol. 2014, 4, 2777–2786. [Google Scholar] [CrossRef] [PubMed]
- Callaway, R.M.; Aschehoug, E.T. Invasive plants versus their new and old neighbors: A mechanism for exotic invasion. Science 2000, 290, 521–523. [Google Scholar] [CrossRef]
- Cappuccino, N.; Arnason, J.T. Novel chemistry of invasive exotic plants. Biol. Lett. 2006, 2, 189–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.O.; Lee, E.J. Comparison of phenolic compounds and the effects of invasive and native species in East Asia: Support for the novel weapons hypothesis. Ecol. Res. 2011, 26, 87–94. [Google Scholar] [CrossRef]
- Sladonja, B.; Poljuha, D.; Uzelac, M. Non-Native Invasive Species as Ecosystem Service Providers. In Ecosystem Services and Global Ecology; Hufnagel, L., Ed.; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Sahu, A.; Devkota, A. Antimicrobial activity of leaf extracts of some invasive alien plant species of Aster family against clinical bacteria. Ecoprint Int. J. Ecol. 2016, 23, 1–12. [Google Scholar] [CrossRef]
- Poljuha, D.; Sladonja, B.; Šola, I.; Dudaš, S.; Bilić, J.; Rusak, G.; Motlhatlego, K.E.; Eloff, J.N. Phenolic composition of leaf extracts of Ailanthus altissima (Simaroubaceae) with antibacterial and antifungal activity equivalent to standard antibiotics. Nat. Prod. Commun. 2017, 12, 1609–1612. [Google Scholar] [CrossRef] [Green Version]
- Meela, M.M.; Mdee, L.K.; Masoko, P.; Eloff, J.N. Acetone leaf extracts of seven invasive weeds have promising activity against eight important plant fungal pathogens. S. Afr. J. Bot. 2019, 123, 93–97. [Google Scholar] [CrossRef]
- Eloff, J.; Angeh, I.; Mcgaw, L. Solvent-solvent fractionation can increase the antifungal activity of a Melianthus comosus (Melianthaceae) acetone extract to yield a potentially useful commercial antifungal product. Ind. Crop. Prod. 2018, 111, 69–77. [Google Scholar] [CrossRef]
- Sladonja, B.; Poljuha, D.; Sušek, M.; Dudaš, S. Herbicidal effect of Ailanthus altissima leaves water extracts on Medicago sativa seeds germination. In Proceedings of the Conference VIVUS: Transmission of Innovations, Knowledge and Practical Experience into Everyday Practice, Naklo, Slovenia, 14–15 November 2014; Maček, M.A., Maček Jerala, M., Kolenc Artiček, M., Eds.; Biotehniški Center: Naklo, Slovenia, 2014; pp. 476–481. [Google Scholar]
- Sladonja, B.; Poljuha, D.; Uzelac, M. Tree of Heaven leaf extract: A new generation of bioherbicides. In Proceedings of the International Conference on Biological Invasions: The Human Role in Biological Invasions—A case of Dr Jekyll and Mr Hyde? NEOBIOTA, Vodice, Hrvatsko ekološko društvo, Zagreb, Croatia, 15–18 September 2020; p. 107. [Google Scholar]
- Peter, A.; Dujmović Purgar, D.; Voća, N.; Filipčić, P. Use of invasive plant species Japanese knotweed (Reynoutria japonica Houtt.) biomass in energy production. In Proceedings of the 3rd Croatian Symposium on Invasive Species with International Participation, Zagreb, Croatia, 26–27 November 2018; Jelaska, S.D., Ed.; Croatian Ecological Society: Zagreb, Croatia, 2018; p. 79. [Google Scholar]
- Nikolić, T.; Mitić, B.; Boršić, I. Flora Hrvatske—Invazivne biljke; Izdavač ALFA dd.: Zagreb, Croatia, 2014. [Google Scholar]
- Kožuharova, E.; Lebanova, H.; Getov, I.; Benbassat, N.; Kochmarov, V. Ailanthus altissima (Mill.) Swingle—A terrible invasive pest in Bulgaria or potential useful plant? Bothalia J. 2014, 44, 213–230. [Google Scholar]
- Ding, W.; Huang, R.; Zhou, Z.; Li, Y. New Sesquiterpenoids from Ambrosia artemisiifolia L. Molecules 2015, 20, 4450–4459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saluk-Juszczak, J.; Olas, B.; Nowak, P.; Wachowicz, B.; Bald, E.; Głowacki, R.; Pawlaczyk, I.; Gancarz, R. Extract from Conyza canadensis as a modulator of plasma protein oxidation induced by peroxynitrite in vitro. Cent. Eur. J. Biol. 2010, 5, 800–807. [Google Scholar] [CrossRef]
- Brahmi-Chendouh, N.; Piccolella, S.; Crescente, G.; Pacifico, F.; Boulekbache-Makhlouf, L.; Hamri-Zeghichi, S.; Akkal, S.; Madani, K.; Pacifico, S. A nutraceutical extract from Inula viscosa leaves: UHPLC-HR-MS/MS based polyphenol profile, and antioxidant and cytotoxic activities. J. Food Drug Anal. 2019, 27, 692–702. [Google Scholar] [CrossRef] [PubMed]
- Kamboj, A.; Saluja, A.K. Phytopharmacological review of Xanthium strumarium L. (Cocklebur). Int. J. Green Pharm. 2010, 4, 129–139. [Google Scholar] [CrossRef]
- Jeong, C.H.; Nam, E.K.; Shim, K.H. Chemical Components in Different Parts of Erigeron annuus. J. Korean Soc. Food Sci. Nutr. 2005, 34, 857–861. [Google Scholar] [CrossRef]
- Chalchat, J.C.; Maksimović, Z.A.; Petrović, S.D.; Gorunović, M.S.; Đordević, S.; Mraović, M. Chemical composition and antimicrobial activity of Ambrosia artemisiifolia L. essential oil. J. Essent. Oil Res. 2004, 16, 270–273. [Google Scholar] [CrossRef]
- Kowarik, I.; Säumel, I. Biological flora of Central Europe: Ailanthus altissima (Mill.) Swingle. Perspect. Plant Ecol. Evol. Syst. 2007, 8, 207–237. [Google Scholar] [CrossRef]
- Talib, W.; Abu Zarga, M.; Mahasneh, A. Antiproliferative, antimicrobial and apoptosis inducing effects of compounds isolated from Inula viscosa. Molecules 2012, 17, 3291–3303. [Google Scholar] [CrossRef]
- Biswas, K.; Sinha, S.N. Evaluation of antibacterial activity of Conyza canadensis (L.) Cronquist collected from Nainital, Uttarakhand, India. Int. J. Univ. Pharm. Biol. Sci. 2014, 3, 46–53. [Google Scholar]
- Sharma, R.K.; Verma, N.; Jha, K.K.; Singh, N.K.; Kumar, B. Phytochemistry, pharmacological activity, traditional and medicinal uses of Erigeron species: A review. IJARI 2014, 2, 379–383. [Google Scholar]
- Al-Mekhlafi, F.A.; Abutaha, N.; Mashaly, A.M.A.; Nasr, F.A.; Ibrahim, K.E.; Wadaan, M.A. Biological activity of Xanthium strumarium seed extracts on different cancer cell lines and Aedes caspius, Culex pipiens (Diptera: Culicidae). Saudi J. Biol. Sci. 2017, 24, 817–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietta, P.; Minoggio, M.; Bramati, L. Plant Polyphenols: Structure, Occurrence and Bioactivity. Stud. Nat. Prod. Chem. 2003, 28, 257–312. [Google Scholar] [CrossRef]
- Eloff, J.N.; Picard, J.; Masoko, P. Resistance of animal fungal pathogens to solvents used in bioassays. S. Afr. J. Bot. 2007, 73, 667–669. [Google Scholar] [CrossRef] [Green Version]
- Eloff, J.N. Which extractant should be used for the screening and isolation of antimicrobial components from plants? J. Ethnopharmacol. 1998, 60, 1–8. [Google Scholar] [CrossRef]
- Kotze, M.; Eloff, J.N. Extraction of antibacterial compounds from Combretum microphyllum (Combretaceae). S. Afr. J. Bot. 2002, 68, 62–67. [Google Scholar] [CrossRef] [Green Version]
- Panda, S.K.; Luyten, W. Antiparasitic activity in Asteraceae with special attention to ethnobotanical use by the tribes of Odisha, India. Parasite 2018, 25, 10. [Google Scholar] [CrossRef] [Green Version]
- Sytar, O.; Hemmerich, I.; Zivcak, M.; Rauh, C.; Brestic, M. Comparative analysis of bioactive phenolic compounds composition from 26 medicinal plants. Saudi J. Biol. Sci. 2018, 25, 631–641. [Google Scholar] [CrossRef] [Green Version]
- Sladonja, B.; Sušek, M.; Guillermic, J. Review on Invasive Tree of Heaven (Ailanthus altissima (Mill.) Swingle) Conflicting Values: Assessment of Its Ecosystem Services and Potential Biological Threat. Environ. Manag. 2015, 56, 1009–1034. [Google Scholar] [CrossRef]
- Kurita, S.; Kashiwagi, T.; Ebisu, T.; Shimamura, T.; Ukeda, H. Identification of neochlorogenic acid as the predominant antioxidant in Polygonum cuspidatum leaves. Ital. J. Food Sci. 2016, 28, 25–31. [Google Scholar] [CrossRef]
- Cipollini, D.; Stevenson, R.; Enright, S.; Eyles, A.; Bonello, P. Phenolic metabolites in leaves of the invasive shrub, Lonicera maackii, and their potential phytotoxic and anti-herbivore effects. J. Chem. Ecol. 2008, 34, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Omokhua, A.G.; Abdalla, M.A.; Van Staden, J.A. comprehensive study of the potential phytomedicinal use and toxicity of invasive Tithonia species in South Africa. BMC Complement. Altern. Med. 2018, 18, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitalini, S.; Orlando, F.; Palmioli, A.; Alali, S.; Airoldi, C.; De Noni, I.; Vaglia, V.; Bocchi, S.; Iriti, M. Different phytotoxic effect of Lolium multiflorum Lam. leaves against Echinochloa oryzoides (Ard.) Fritsch and Oriza sativa L. Environ. Sci. Pollut. Res. 2020, 27, 33204–33214. [Google Scholar] [CrossRef] [PubMed]
- Omezzine, F.; Bouaziz, M.; Simmonds, M.S.J.; Haouala, R. Variation in chemical composition and allelopathic potential of mixoploid Trigonella foenum graecum L. with developmental stage. Food Chem. 2014, 148, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Liu, W.-X.; Zheng, M.-F.; Xu, Q.-L.; Wan, F.-H.; Wang, J.; Lei, T.; Zhou, Z.-Y.; Tan, J.-W. Bioactive quinic acid derivatives from Ageratina adenophora. Molecules 2013, 18, 14096–14104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefanescu, B.E.; Szabo, K.; Mocan, A.; Crisan, G. Phenolic compounds from five Ericaceae species leaves and their related bioavailability and health benefits. Molecules 2019, 24, 2046. [Google Scholar] [CrossRef] [Green Version]
- Dias, M.I.; Barros, L.; Morales, P.; Camara, M.; Alves, M.J.; Oliveira, M.B.P.P.; Santos-Buelga, C.; Ferreira, I.C.F.R. Wild Fragaria vesca L. fruits: A rich source of bioactive phytochemicals. Food Funct. 2016, 7, 4523. [Google Scholar] [CrossRef]
- Duckstein, S.M.; Lotter, E.M.; Meyer, U.; Lindequist, U.; Stintzing, F.C. Phenolic constituents from Alchemilla vulgaris L. and Alchemilla mollis (Buser) Rothm. at different dates of harvest. Z. Für Nat. C 2012, 67, 529–540. [Google Scholar] [CrossRef] [Green Version]
- Jones, G.L.; Tomlinson, M.; Owen, R.; Scullion, J.; Winters, A.; Jenkins, T.; Ratcliffe, J.; Gwynn-Jones, D. Shrub establishment favoured and grass dominance reduced in acid heath grassland systems cleared of invasive Rhododendron ponticum. Sci. Rep. 2019, 9, 2239. [Google Scholar] [CrossRef]
- Muzaffar, S.; Ali, B.; Wani, N.A. Effect of catechol, gallic acid and pyrogallic acid on the germination, seedling growth and the level of endogenous phenolics in cucumber (Cucumis sativus L.). Inter. J. Life Sci. Biotechnol. Pharma. Res. 2012, 1, 50–55. [Google Scholar]
- Weidenhamer, J.D.; Romeo, J.T. Allelochemicals of Polygonella myriophylla: Chemistry and soil degradation. J. Chem. Ecol. 2004, 30, 1061–1078. [Google Scholar] [CrossRef] [PubMed]
- Weidenhamer, J.D.; Li, M.; Allman, J.; Bergosh, R.G.; Posner, M. Evidence does not support a role for gallic acid in Phragmites australis invasion success. J. Chem. Ecol. 2013, 39, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Yoo, N.H.; Jang, D.S.; Yoo, J.L.; Lee, Y.M.; Kim, Y.S.; Cho, J.H.; Kim, J.S. Erigeroflavanone, a flavanone derivative from the flowers of Erigeron annuus with protein glycation and aldose reductase inhibitory activity. J. Nat. Prod. 2008, 71, 713–715. [Google Scholar] [CrossRef] [PubMed]
- Rhimi, W.; Salem, I.B.; Immediato, D.; Saidi, M.; Boulila, A.; Cafarchia, C. Chemical composition, antibacterial and antifungal activities of crude Dittrichia viscosa (L.) Greuter leaf extracts. Molecules 2017, 22, 942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Queiroz, S.; Cantrell, C.; Duke, S.; Wedge, D.; Nandula, V.; Moraes, R.; Cerdeira, A. Bioassay-Directed Isolation and Identification of Phytotoxic and Fungitoxic Acetylenes from Conyza canadensis. J. Agric. Food Chem. 2012, 60, 5893–5898. [Google Scholar] [CrossRef] [PubMed]
- Martinez, E.J.L.; Barrales, P.O.; Zengin, G.; Uysal, S.; Ceylan, R.; Guler, G.O.; Mocan, A.; Aktumsek, A. Lathyrus aureus and Lathyrus pratensis: Characterization of phytochemical profiles by liquid chromatography-mass spectrometry, and evaluation of their enzyme inhibitory and antioxidant activities. RSC Adv. 2016, 6, 88996–89006. [Google Scholar] [CrossRef]
- Yisimayili, Z.; Abdulla, R.; Tian, Q.; Wang, Y.; Chen, M.; Sun, Z.; Li, Z.; Liu, F.; Aisa, H.A.; Huang, C. A comprehensive study of pomegranate flowers polyphenols and metabolites in rat biological samples by high-performance liquid chromatography quadrupole time-of-flight mass spectrometry. J. Chromatogr. A 2019, 1604, 460472. [Google Scholar] [CrossRef]
- Fontanel, D.; Galtier, C.; Viel, C.; Gueiffier, A. Caffeoyl quinic and tartaric acids and flavonoids from Lapsana communis L. subsp. communis (Asteraceae). Z. Für Nat. C 1998, 53, 1090–1092. [Google Scholar] [CrossRef]
- Duke, S.O.; Blair, A.C.; Dayan, F.E.; Johnson, R.D.; Meepagala, K.M.; Cook, D.; Bajsa, J. Is (−)-catechin a novel weapon of spotted knapweed (Centaurea stoebe)? J. Chem. Ecol. 2009, 35, 141–153. [Google Scholar] [CrossRef]
- Eloff, J.N. A Proposal towards a Rational Classification of the Antimicrobial Activity of Acetone Tree Leaf Extracts in a Search for New Antimicrobials. Planta Med. 2021, 87, 836–840. [Google Scholar] [CrossRef]
- Eloff, J.N. Quantifying the bioactivity of plant extracts during screening and bioassay-guided fractionation. Phytomedicine 2004, 11, 370–371. [Google Scholar] [CrossRef] [PubMed]
- Kuete, V.; Krusche, B.; Youns, M.; Voukeng, I.; Fankam, A.G.; Tankeo, S.; Lacmata, S.; Efferth, T. Cytotoxicity of some Cameroonian spices and selected medicinal plant extracts. J. Ethnopharmacol. 2011, 134, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.S.; Elgorashi, E.E.; Moodley, N.; McGaw, L.J.; Naidoo, V.; Eloff, J.N. The antimicrobial, antioxidative, anti-inflammatory activity and cytotoxicity of different fractions of four South African Bauhinia species used traditionally to treat diarrhoea. J. Ethnopharmacol. 2012, 143, 826–839. [Google Scholar] [CrossRef]
- Invazivne Vrste. Available online: https://invazivnevrste.haop.hr/crna-i-bijela-lista/2 (accessed on 17 October 2021).
- Global Register of Introduced and Invasive Species—GRIIS. Available online: http://www.griis.org (accessed on 20 December 2021).
- Mikulic-Petkovsek, M.; Schmitzer, V.; Slatnar, A.; Weber, N.; Veberic, R.; Stampar, F.; Munda, A.; Koron, D. Alteration of the content of primary and secondary metabolites in strawberry fruit by Colletotrichum nymphaeae infection. J. Agric. Food Chem. 2013, 61, 5987–5995. [Google Scholar] [CrossRef] [PubMed]
- Eloff, J.N. A sensitive and quick microplate method to determine the minimal inhibitory concentration of plant extracts for bacteria. Planta Med. 1998, 64, 711–714. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.Y.; Zheng, W.; Galletta, G.J. Cultural system affects fruit quality and antioxidant capacity in strawberries. J. Agric. Food Chem. 2002, 50, 6534–6542. [Google Scholar] [CrossRef]
- Mikulic-Petkovsek, M.; Ivancic, A.; Todorovic, B.; Veberic, R.; Stampar, F. Fruit phenolic composition of different elderberry species and hybrids. J. Food Sci. 2015, 80, C2180–C2190. [Google Scholar] [CrossRef]
- Masoko, P.; Picard, J.; Eloff, J.N. Antifungal activities of six South African Terminalia species (Combretaceae). J. Ethnopharmacol. 2005, 99, 301–308. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Method 1983, 65, 55–63. [Google Scholar] [CrossRef]
- McGaw, L.J.; Van Der Merwe, D.; Eloff, J.N. In vitro anthelmintic, antibacterial and cytotoxic effects of extracts from plants used in South African ethnoveterinary medicine. Vet. J. 2007, 173, 366–372. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
| Ailanthus altissima | Ambrosia artemisiifolia | Conyza canadensis | Dittrichia viscosa | Erigeron annuus | Xanthium strumarium | Stat. Significance | |
|---|---|---|---|---|---|---|---|
| Dicaffeoylquinic acids | n.d. | 3.78 ± 0.06 | 4.76 ± 0.52 | 22.36 ± 0.14 | 3.99 ± 0.33 | 11.46 ± 0.18 | |
| 3-caffeoylquinic acid | 2.44 ± 0.20 | 0.37 ± 0.00 | 0.26 ± 0.02 | 0.24 ± 0.01 | 0.18 ± 0.01 | 0.61 ± 0.06 | |
| 4-caffeoylquinic acid | 2.83 ± 0.28 | 0.28 ± 0.02 | 0.28 ± 0.03 | n.d. | 0.13 ± 0.02 | 0.20 ± 0.02 | |
| 5-caffeoylquinic acid | 7.42 ± 1.11 | 5.04 ± 0.15 | 2.72 ± 0.10 | 8.42 ± 0.08 | 2.86 ± 0.21 | 11.25 ± 0.09 | |
| triCQA | n.d. | n.d. | n.d. | n.d. | 1.06 ± 0.04 | 0.83 ± 0.04 | |
| Caffeic acid | n.d. | 0.43 ± 0.00 | n.d. | n.d. | n.d. | n.d. | |
| Other caffeic acid derivatives | n.d. | 0.21 ± 0.01 | n.d. | n.d. | 0.15 ± 0.00 | n.d. | |
| Caffeic acid derivatives total | 12.68 ± 1.62 b | 10.11 ± 0.28 b | 8.02 ± 0.77 b | 31.02 ± 0.01 a | 8.37 ± 0.72 b | 24.35 ± 0.81 a | *** |
| Caftaric acids | n.d. | 3.72 ± 0.09 | n.d. | n.d. | n.d. | n.d. | |
| 3-p-coumaroylquinic acid | 0.09 ± 0.02 | 0.03 ± 0.01 | 0.04 ± 0.01 | 0.03 ± 0.01 | 0.05 ± 0.00 | n.d. | |
| 4-p-coumaroylquinic acid | 0.24 ± 0.05 | n.d. | n.d. | n.d. | n.d. | n.d. | |
| 5-p-coumaroylquinic acid | 0.68 ± 0.14 | 0.11 ± 0.01 | 0.04 ± 0.00 | 0.03 ± 0.01 | n.d. | 0.04 ± 0.00 | |
| p-coumaric acid derivatives | 1.02 ± 0.19 a | 0.14 ± 0.01 b | 0.09 ± 0.01 b | 0.07 ± 0.01 b | 0.05 ± 0.00 b | 0.04 ± 0.00 b | ** |
| 3-feruloylquinic acid | 0.09 ± 0.01 | 0.03 ± 0.00 | 0.03 ± 0.01 | n.d. | 0.01 ± 0.00 | 0.01 ± 0.00 | |
| 5-feruloylquinic acid | n.d. | 0.31 ± 0.02 | n.d. | n.d. | n.d. | 0.34 ± 0.04 | |
| Ferulic acid derivatives | 0.09 ± 0.01 b | 0.35 ± 0.01 a | 0.03 ± 0.00 c | n.d. | 0.01 ± 0.00 c | 0.35 ± 0.04 a | *** |
| Hydroxycinnamic acid derivatives | 13.79 ± 1.82 b | 10.60 ± 0.29 b | 8.14 ± 0.78 b | 31.08 ± 0.11 a | 8.21 ± 0.73 b | 24.39 ± 0.42 a | *** |
| Ellagic acid | n.d. | n.d. | 3.24 ± 1.61 | n.d. | n.d. | n.d. | |
| Ellagic acid pentoside | 12.13 ± 2.31 | n.d. | n.d. | n.d. | n.d. | n.d. | |
| Ellagic acid hexosides | n.d. | n.d. | n.d. | 31.63 ± 0.43 | 1.07 ± 0.18 | n.d. | |
| Ellagic acid rhamnoside | n.d. | n.d. | n.d. | n.d. | 0.07 ± 0.01 | n.d. | |
| Hydroxybenzoic acid derivatives | 12.13 ± 2.39 b | n.d. | 3.24 ± 1.97 bc | 31.63 ± 0.53 a | 1.14 ± 0.23 c | n.d. | *** |
| Apigenin | n.d. | n.d. | n.d. | n.d. | n.d. | 0.05 ± 0.01 | |
| Apigenin hexoside | 0.87 ± 0.07 | n.d. | n.d. | n.d. | n.d. | n.d. | |
| Flavones | 0.87 ± 0.07 a | n.d. | n.d. | n.d. | n.d. | 0.05 ± 0.01 b | *** |
| Procyanidin dimer | n.d. | n.d. | n.d. | n.d. | n.d. | 1.19 ± 0.09 | |
| Epicatechin | n.d. | n.d. | n.d. | n.d. | n.d. | 0.02 ± 0.00 | |
| Flavanols | n.d. | n.d. | n.d. | n.d. | n.d. | 1.21 ± 0.11 | |
| Quercetin-dihexoside | n.d. | 1.19 ± 0.06 | 0.04 ± 0.01 | n.d. | 0.15 ± 0.02 | n.d. | |
| Quercetin-galloyl-hexosides | 0.69 ± 0.07 | n.d. | n.d. | n.d. | n.d. | n.d. | |
| Quercetin-3-galactoside | 2.81 ± 0.28 | 0.45 ± 0.01 | n.d. | 0.25 ± 0.10 | n.d. | n.d. | |
| Quercetin-3-glucoside | 0.56 ± 0.05 | 0.10 ± 0.00 | 0.05 ± 0.01 | 0.11 ± 0.01 | 0.01 ± 0.00 | 0.34 ± 0.00 | *** |
| Quercetin-3-rutinoside | n.d. | 0.24 ± 0.00 | n.d. | 0.25 ± 0.01 | n.d. | 0.10 ± 0.02 | |
| Quercetin-3-xyloside | n.d. | 0.01 ± 0.00 | 0.06 ± 0.01 | 0.08 ± 0.00 | n.d. | 0.05 ± 0.01 | |
| Quercetin-3-rhamnoside | n.d. | n.d. | n.d. | n.d. | 0.27 ± 0.02 | n.d. | |
| Quercetin-3-glucuronide | n.d. | 0.43 ± 0.01 | 0.07 ± 0.02 | 0.24 ± 0.00 | 0.08 ± 0.01 | 0.75 ± 0.02 | |
| Quercetin-3-arabinopyranoside | n.d. | 0.10 ± 0.00 | 0.10 ± 0.01 | 1.32 ± 0.09 | n.d. | n.d. | |
| Quercetin-acetyl hexoside | 0.13 ± 0.02 | n.d. | n.d. | n.d. | n.d. | n.d. | |
| Kaempferol-hexoside | 1.04 ± 0.09 | 0.06 ± 0.00 | 0.07 ± 0.02 | 0.05 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 | |
| Kaempferol-acetyl-hexosides | 0.43 ± 0.05 | 0.19 ± 0.00 | n.d. | n.d. | n.d. | n.d. | |
| Kaempferol-3-rutinoside | n.d. | n.d. | n.d. | n.d. | 0.31 ± 0.04 | n.d. | |
| Kaempferol-3-glucuronide | n.d. | 0.19 ± 0.01 | 1.05 ± 0.07 | 0.56 ± 0.02 | 2.95 ± 0.28 | 0.38 ± 0.01 | |
| Kaempferol-rhamnoside-hexoside | 0.10 ± 0.01 | n.d. | n.d. | n.d. | 0.09 ± 0.01 | n.d. | |
| Kaempferol-galloyl-hexoside | 0.22 ± 0.02 | n.d. | n.d. | n.d. | 0.02 ± 0.00 | n.d. | |
| Kaempferol-glucuronyl-hexoside | n.d. | n.d. | n.d. | n.d. | 0.02 ± 0.00 | n.d. | *** |
| Isorhamnetin hexoside | n.d. | 0.19 ± 0.01 | n.d. | 1.34 ± 0.07 | n.d. | n.d. | |
| Isorhamnetin acetyl hexoside | n.d. | 0.31 ± 0.01 | n.d. | n.d. | n.d. | n.d. | |
| Isorhamnetin-3-rutinoside | n.d. | 0.03 ± 0.00 | n.d. | n.d. | n.d. | n.d. | *** |
| Myricetin-3-glucuronide | n.d. | n.d. | n.d. | n.d. | 0.17 ± 0.03 | n.d. | |
| Myricetin hexoside | n.d. | n.d. | n.d. | 0.64 ± 0.02 | n.d. | n.d. | |
| Laricitrin-3-glucuronide | n.d. | 0.57 ± 0.01 | n.d. | 0.23 ± 0.01 | n.d. | n.d. | |
| Laricitrin | n.d. | n.d. | n.d. | n.d. | n.d. | 0.65 ± 0.03 | |
| Syringetin | n.d. | n.d. | n.d. | n.d. | n.d. | 0.44 ± 0.03 | |
| Flavonols | 5.97 ± 0.58 a | 4.05 ± 0.14 abc | 1.45 ± 0.14 c | 5.07 ± 0.28 ab | 4.07 ± 0.49 abc | 3.23 ± 0.10 bc | *** |
| Naringenin-hexosides | n.d. | n.d. | 0.17 ± 0.05 b | n.d. | 32.07 ± 2.05 | n.d. | *** |
| Flavanones | n.d. | n.d. | 0.17 ± 0.05 b | n.d. | 32.07 ± 2.05 | n.d. | *** |
| Vescalagin | 6.55 ± 1.27 | n.d. | n.d. | n.d. | n.d. | n.d. | *** |
| Ellagitannins | 6.55 ± 1.27 | n.d. | n.d. | n.d. | n.d. | n.d. | |
| Gallic acid | n.d. | 0.35 ± 0.01 b | n.d. | n.d. | n.d. | n.d. | |
| Digalloyl-HHDP-hexoside isomer | 2.39 ± 0.52 a | n.d. | n.d. | n.d. | n.d. | n.d. | *** |
| Gallotannins | 2.39 ± 0.52 a | 0.35 ± 0.01 b | n.d. | n.d. | n.d. | n.d. | *** |
| TOTAL | 41.70 ± 6.59 ab | 15.00 ± 0.42 b | 13.00 ± 2.90 b | 67.78 ± 0.57 a | 45.50 ± 4.32 ab | 29.02 ± 0.65 ab |
| Plant Species | Yield | LC50 | Escherichia coli | Enterococcus faecalis | Staphylococcus aureus | Pseudomonas aeruginosa | Average | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MIC | TAA | SI | MIC | TAA | SI | MIC | TAA | SI | MIC | TAA | SI | MIC | TAA | SI | |||
| A. altissima | 42 | 0.17 | 0.23 | 183 | 0.74 | 0.04 | 1050 | 4.25 | 0.31 | 135 | 0.55 | 0.31 | 135 | 0.55 | 0.22 | 376 | 1.52 |
| A. artemisiifolia | 101 | 0.02 | 0.31 | 326 | 0.06 | 0.06 | 1683 | 0.33 | 0.31 | 326 | 0.06 | 0.63 | 160 | 0.03 | 0.32 | 624 | 0.12 |
| C. canadensis | 47 | 0.17 | 0.31 | 152 | 0.55 | 0.04 | 1175 | 4.25 | 1.88 | 25 | 0.09 | 1.25 | 38 | 0.14 | 0.87 | 348 | 1.29 |
| D. viscosa | 63 | 0.03 | 0.16 | 394 | 0.19 | 0.04 | 1575 | 0.75 | 0.31 | 203 | 0.10 | 0.31 | 203 | 0.10 | 0.21 | 594 | 0.33 |
| E. annuus | 66 | 0.15 | 0.31 | 213 | 0.48 | 0.04 | 1650 | 3.75 | 1.25 | 53 | 0.12 | 1.25 | 53 | 0.12 | 0.71 | 532 | 1.17 |
| X. strumarium | 57 | 0.001 | 0.23 | 248 | 0.004 | 0.04 | 1425 | 0.03 | 0.08 | 713 | 0.01 | 0.08 | 713 | 0.01 | 0.11 | 775 | 0.01 |
| Average | 0.26 | 253 | 0.34 | 0.04 | 1426 | 2.23 | 0.69 | 243 | 0.16 | 0.64 | 217 | 0.16 | |||||
| Gentamicin | 8 × 10−4 | 2 × 10−4 | 1.3 × 10−4 | 3 × 10−4 | 3.6 × 10−4 | ||||||||||||
| Doxorubicin | 0.012 | ||||||||||||||||
| Plant Species | Yield | LC50 | Candida albicans | Cryptococcus neoformans | Aspergillus fumigatus | Average | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MIC | TAA | SI | MIC | TAA | SI | MIC | TAA | SI | MIC | TAA | SI | |||
| A. altissima | 42 | 0.17 | 0.16 | 269 | 1.12 | 0.31 | 134 | 0.56 | 0.16 | 269 | 1.12 | 0.21 | 224 | 0.93 |
| A. artemisiifolia | 101 | 0.02 | 0.16 | 646 | 0.11 | 0.63 | 162 | 0.03 | 1.25 | 81 | 0.01 | 0.68 | 296 | 0.05 |
| C. canadensis | 47 | 0.17 | 0.16 | 301 | 1.09 | 0.31 | 150 | 0.55 | 0.63 | 75 | 0.27 | 0.37 | 175 | 0.64 |
| D. viscosa | 63 | 0.03 | 0.16 | 403 | 0.22 | 0.31 | 202 | 0.11 | 0.63 | 101 | 0.05 | 0.37 | 235 | 0.13 |
| E. annuus | 66 | 0.15 | 0.31 | 211 | 0.46 | 0.31 | 211 | 0.46 | 1.25 | 53 | 0.12 | 0.62 | 158 | 0.35 |
| X. strumarium | 57 | 0.001 | 0.23 | 243 | 0.003 | 0.16 | 365 | 0.01 | 0.23 | 243 | 0.003 | 0.21 | 284 | 0.003 |
| Average | 0.20 | 346 | 0.50 | 0.34 | 204 | 0.29 | 0.69 | 137 | 0.26 | |||||
| Amphotericin B | 8 × 10−3 | 8 × 10−3 | 8 × 10−3 | |||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Poljuha, D.; Sladonja, B.; Šola, I.; Šenica, M.; Uzelac, M.; Veberič, R.; Hudina, M.; Famuyide, I.M.; Eloff, J.N.; Mikulic-Petkovsek, M. LC–DAD–MS Phenolic Characterisation of Six Invasive Plant Species in Croatia and Determination of Their Antimicrobial and Cytotoxic Activity. Plants 2022, 11, 596. https://doi.org/10.3390/plants11050596
Poljuha D, Sladonja B, Šola I, Šenica M, Uzelac M, Veberič R, Hudina M, Famuyide IM, Eloff JN, Mikulic-Petkovsek M. LC–DAD–MS Phenolic Characterisation of Six Invasive Plant Species in Croatia and Determination of Their Antimicrobial and Cytotoxic Activity. Plants. 2022; 11(5):596. https://doi.org/10.3390/plants11050596
Chicago/Turabian StylePoljuha, Danijela, Barbara Sladonja, Ivana Šola, Mateja Šenica, Mirela Uzelac, Robert Veberič, Metka Hudina, Ibukun Michael Famuyide, Jacobus N. Eloff, and Maja Mikulic-Petkovsek. 2022. "LC–DAD–MS Phenolic Characterisation of Six Invasive Plant Species in Croatia and Determination of Their Antimicrobial and Cytotoxic Activity" Plants 11, no. 5: 596. https://doi.org/10.3390/plants11050596
APA StylePoljuha, D., Sladonja, B., Šola, I., Šenica, M., Uzelac, M., Veberič, R., Hudina, M., Famuyide, I. M., Eloff, J. N., & Mikulic-Petkovsek, M. (2022). LC–DAD–MS Phenolic Characterisation of Six Invasive Plant Species in Croatia and Determination of Their Antimicrobial and Cytotoxic Activity. Plants, 11(5), 596. https://doi.org/10.3390/plants11050596