
Tobacco Rattle Virus as a Tool for Rapid Reverse-Genetics Screens and Analysis of Gene Function in Cannabis sativa L.



Abstract
:1. Introduction
2. Results and Discussion
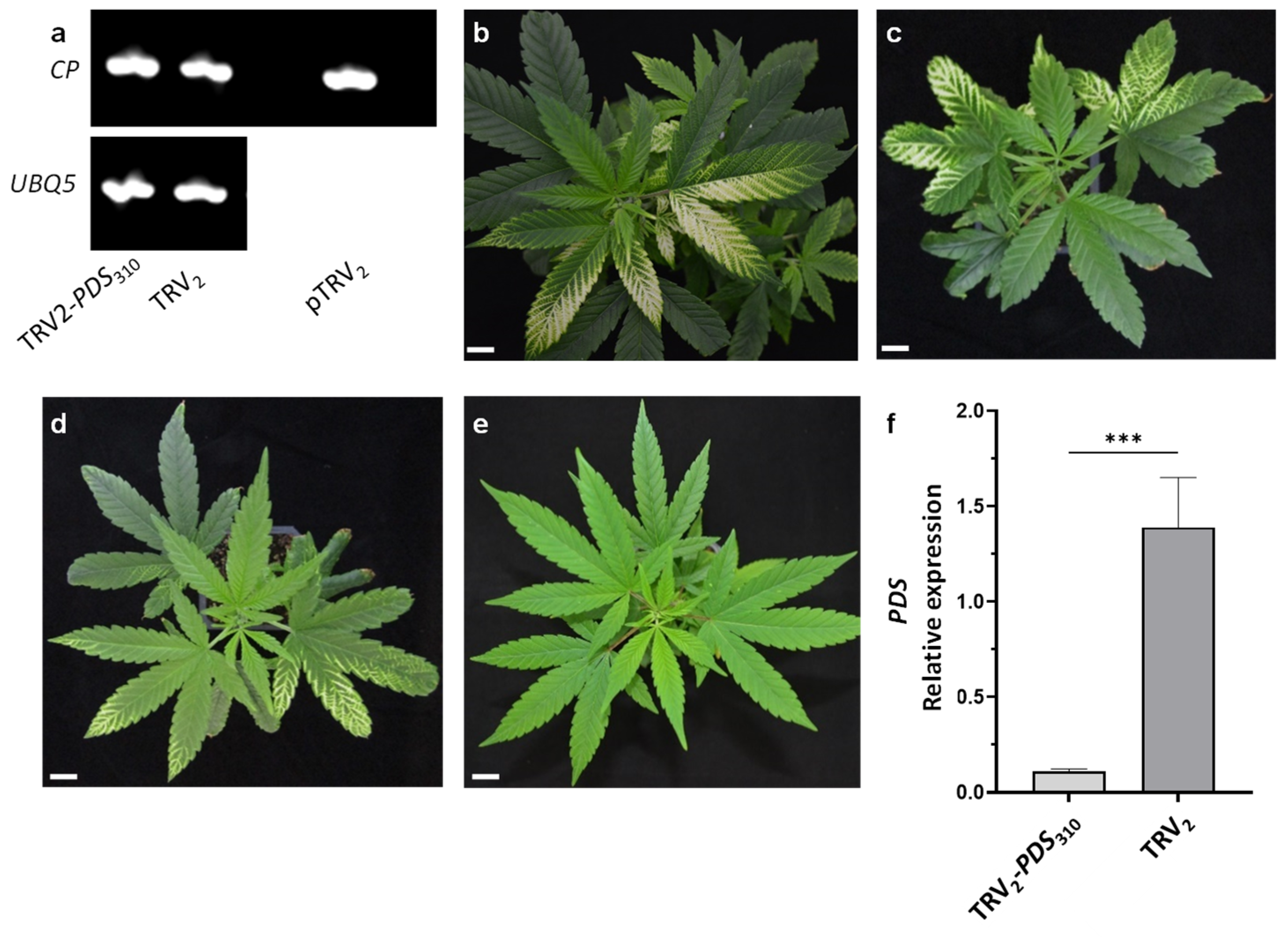
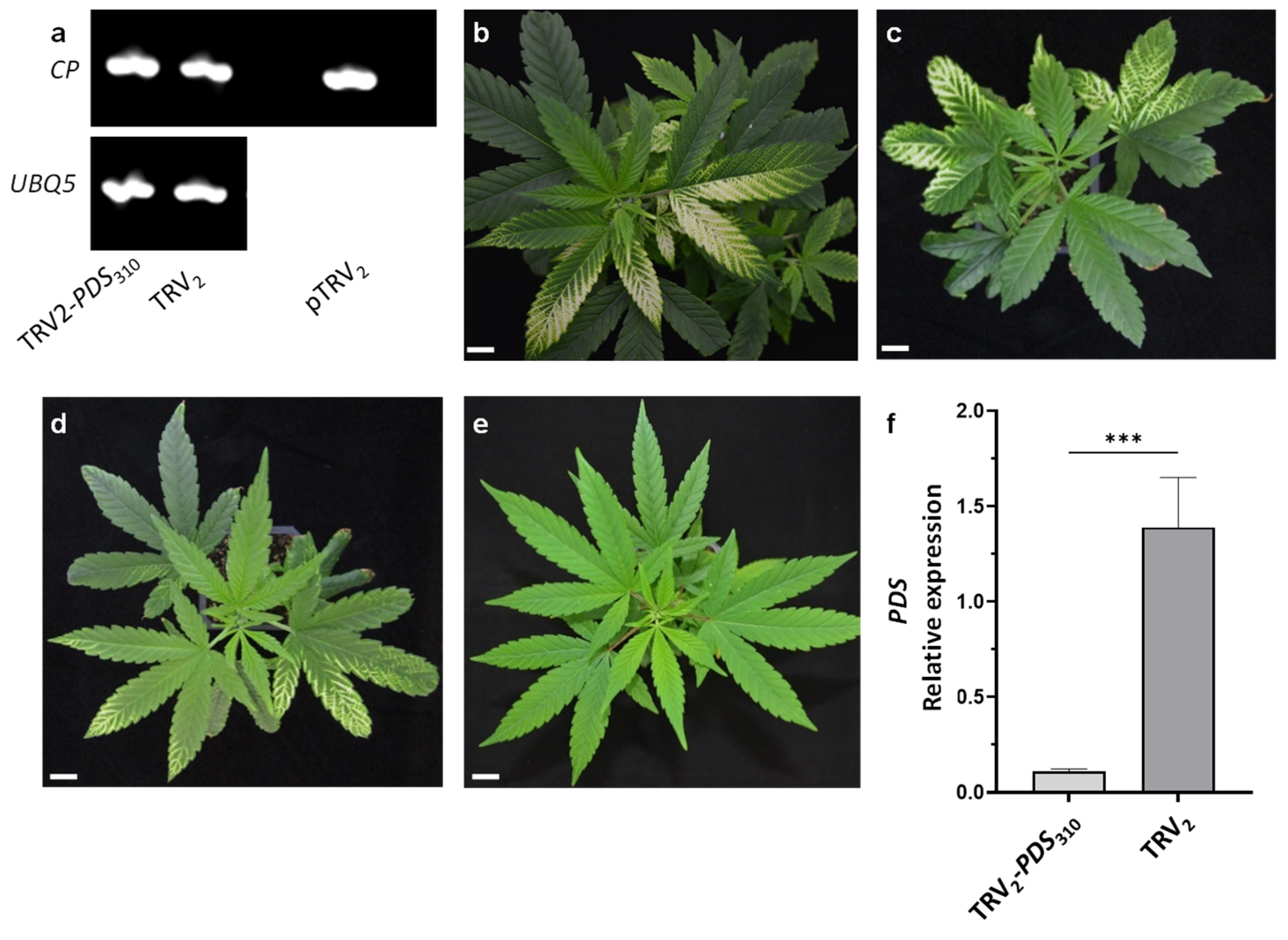
2.1. VIGS in Cannabis Using a TRV Vector
2.2. VAGE in Cannabis Using a TRV Vector
3. Materials and Methods
3.1. Plant Material
3.2. Construction of pTRV2 Vectors
3.3. Agro-Inoculation of pTRV Vectors
3.4. RNA Extraction, cDNA Synthesis, and Quantitative Real-Time PCR (qRT)
3.5. Imaging
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chandra, S.; Lata, H.; Elsohly, M.A. Propagation of Cannabis for Clinical Research: An Approach Towards a Modern Herbal Medicinal Products Development. Front. Plant Sci. 2020, 11, 958. [Google Scholar] [CrossRef] [PubMed]
- McPartland, J.M. Cannabis Systematics at the Levels of Family, Genus, and Species. Cannabis Cannabinoid Res. 2018, 3, 203–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, R.; Merlin, M. Cannabis: Evolution and Ethnobotany; University of California Press: Berkeley, CA, USA, 2016; ISBN 978-0-520-29248-2. [Google Scholar]
- Small, E. Evolution and Classification of Cannabis Sativa (Marijuana, Hemp) in Relation to Human Utilization. Bot. Rev. 2015, 81, 189–294. [Google Scholar] [CrossRef]
- Chandra, S.; Lata, H.; Khan, I.A.; ElSohly, M.A. Cannabis Sativa L.: Botany and Horticulture. In Cannabis sativa L.—Botany and Biotechnology; Chandra, S., Lata, H., ElSohly, M.A., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 79–100. ISBN 978-3-319-54564-6. [Google Scholar]
- Andre, C.M.; Hausman, J.-F.; Guerriero, G. Cannabis Sativa: The Plant of the Thousand and One Molecules. Front. Plant Sci. 2016, 7, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurgobin, B.; Tamiru-Oli, M.; Welling, M.T.; Doblin, M.S.; Bacic, A.; Whelan, J.; Lewsey, M.G. Recent Advances in Cannabis sativa Genomics Research. New Phytol. 2021, 230, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Prentout, D.; Razumova, O.; Rhoné, B.; Badouin, H.; Henri, H.; Feng, C.; Käfer, J.; Karlov, G.; Marais, G.A.B. An Efficient RNA-Seq-Based Segregation Analysis Identifies the Sex Chromosomes of Cannabis sativa. Genome Res. 2020, 30, 164–172. [Google Scholar] [CrossRef]
- McKernan, K.J.; Helbert, Y.; Kane, L.T.; Ebling, H.; Zhang, L.; Liu, B.; Eaton, Z.; McLaughlin, S.; Kingan, S.; Baybayan, P.; et al. Sequence and Annotation of 42 Cannabis Genomes Reveals Extensive Copy Number Variation in Cannabinoid Synthesis and Pathogen Resistance Genes. bioRxiv 2020. [Google Scholar] [CrossRef]
- Adal, A.M.; Doshi, K.; Holbrook, L.; Mahmoud, S.S. Comparative RNA-Seq Analysis Reveals Genes Associated with Masculinization in Female Cannabis Sativa. Planta 2021, 253, 17. [Google Scholar] [CrossRef]
- Livingston, S.J.; Quilichini, T.D.; Booth, J.K.; Wong, D.C.J.; Rensing, K.H.; Laflamme-Yonkman, J.; Castellarin, S.D.; Bohlmann, J.; Page, J.E.; Samuels, A.L. Cannabis Glandular Trichomes Alter Morphology and Metabolite Content during Flower Maturation. Plant J. 2020, 101, 37–56. [Google Scholar] [CrossRef]
- Lipson Feder, C.; Cohen, O.; Shapira, A.; Katzir, I.; Peer, R.; Guberman, O.; Procaccia, S.; Berman, P.; Flaishman, M.; Meiri, D. Fertilization Following Pollination Predominantly Decreases Phytocannabinoids Accumulation and Alters the Accumulation of Terpenoids in Cannabis Inflorescences. Front. Plant Sci. 2021, 12, 2426. [Google Scholar] [CrossRef]
- Koltai, H.; Namdar, D. Cannabis Phytomolecule “Entourage”: From Domestication to Medical Use. Trends Plant Sci. 2020, 25, 976–984. [Google Scholar] [CrossRef] [PubMed]
- Russo, E.B. The Case for the Entourage Effect and Conventional Breeding of Clinical Cannabis: No “Strain,” No Gain. Front. Plant Sci. 2019, 9, 1969. [Google Scholar] [CrossRef] [PubMed]
- Hesami, M.; Pepe, M.; Alizadeh, M.; Rakei, A.; Baiton, A.; Phineas Jones, A.M. Recent Advances in Cannabis Biotechnology. Ind. Crops Prod. 2020, 158, 113026. [Google Scholar] [CrossRef]
- Romero, P.; Peris, A.; Vergara, K.; Matus, J.T. Comprehending and Improving Cannabis Specialized Metabolism in the Systems Biology Era. Plant Sci. 2020, 298, 110571. [Google Scholar] [CrossRef]
- Braich, S.; Baillie, R.C.; Jewell, L.S.; Spangenberg, G.C.; Cogan, N.O.I. Generation of a Comprehensive Transcriptome Atlas and Transcriptome Dynamics in Medicinal Cannabis. Sci. Rep. 2019, 9, 16583. [Google Scholar] [CrossRef]
- van Bakel, H.; Stout, J.M.; Cote, A.G.; Tallon, C.M.; Sharpe, A.G.; Hughes, T.R.; Page, J.E. The Draft Genome and Transcriptome of Cannabis sativa. Genome Biol. 2011, 12, R102. [Google Scholar] [CrossRef] [Green Version]
- Grassa, C.J.; Wenger, J.P.; Dabney, C.; Poplawski, S.G.; Motley, S.T.; Michael, T.P.; Schwartz, C.J.; Weiblen, G.D. A Complete Cannabis Chromosome Assembly and Adaptive Admixture for Elevated Cannabidiol (CBD) Content. bioRxiv 2018, 458083. [Google Scholar] [CrossRef] [Green Version]
- Laverty, K.U.; Stout, J.M.; Sullivan, M.J.; Shah, H.; Gill, N.; Holbrook, L.; Deikus, G.; Sebra, R.; Hughes, T.R.; Page, J.E.; et al. A Physical and Genetic Map of Cannabis sativa Identifies Extensive Rearrangements at the THC/CBD Acid Synthase Loci. Genome Res. 2019, 29, 146–156. [Google Scholar] [CrossRef] [Green Version]
- Kovalchuk, I.; Pellino, M.; Rigault, P.; van Velzen, R.; Ebersbach, J.; Ashnest, J.R.; Mau, M.; Schranz, M.E.; Alcorn, J.; Laprairie, R.B.; et al. The Genomics of Cannabis and Its Close Relatives. Annu. Rev. Plant Biol. 2020, 71, 713–739. [Google Scholar] [CrossRef] [Green Version]
- de Meijer, E.P.M.; Bagatta, M.; Carboni, A.; Crucitti, P.; Moliterni, V.M.C.; Ranalli, P.; Mandolino, G. The Inheritance of Chemical Phenotype in Cannabis sativa L. Genetics 2003, 163, 335–346. [Google Scholar] [CrossRef]
- Bielecka, M.; Kaminski, F.; Adams, I.; Poulson, H.; Sloan, R.; Li, Y.; Larson, T.R.; Winzer, T.; Graham, I.A. Targeted Mutation of Δ12 and Δ15 Desaturase Genes in Hemp Produce Major Alterations in Seed Fatty Acid Composition Including a High Oleic Hemp Oil. Plant Biotechnol. J. 2014, 12, 613–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galán-Ávila, A.; Gramazio, P.; Ron, M.; Prohens, J.; Herraiz, F.J. A Novel and Rapid Method for Agrobacterium-Mediated Production of Stably Transformed Cannabis Sativa L. Plants. Ind. Crops Prod. 2021, 170, 113691. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, G.; Cheng, C.; Lei, L.; Sun, J.; Xu, Y.; Deng, C.; Dai, Z.; Yang, Z.; Chen, X.; et al. Establishment of an Agrobacterium-Mediated Genetic Transformation and CRISPR/Cas9-Mediated Targeted Mutagenesis in Hemp (Cannabis sativa L.). Plant Biotechnol. J. 2021, 19, 1979–1987. [Google Scholar] [CrossRef] [PubMed]
- Senthil-Kumar, M.; Mysore, K.S. New Dimensions for VIGS in Plant Functional Genomics. Trends Plant Sci. 2011, 16, 656–665. [Google Scholar] [CrossRef]
- Spitzer, B.; Zvi, M.M.B.; Ovadis, M.; Marhevka, E.; Barkai, O.; Edelbaum, O.; Marton, I.; Masci, T.; Alon, M.; Morin, S.; et al. Reverse Genetics of Floral Scent: Application of Tobacco Rattle Virus-Based Gene Silencing in Petunia. Plant Physiol. 2007, 145, 1241–1250. [Google Scholar] [CrossRef] [Green Version]
- Spitzer-Rimon, B.; Marhevka, E.; Barkai, O.; Marton, I.; Edelbaum, O.; Masci, T.; Prathapani, N.-K.; Shklarman, E.; Ovadis, M.; Vainstein, A. EOBII, a Gene Encoding a Flower-Specific Regulator of Phenylpropanoid Volatiles’ Biosynthesis in Petunia. Plant Cell 2010, 22, 1961–1976. [Google Scholar] [CrossRef] [Green Version]
- Spitzer-Rimon, B.; Cna’ani, A.; Vainstein, A. Virus-Aided Gene Expression and Silencing Using TRV for Functional Analysis of Floral Scent-Related Genes. Methods Mol. Biol. 2013, 975, 139–148. [Google Scholar] [CrossRef]
- Lu, R.; Martin-Hernandez, A.M.; Peart, J.R.; Malcuit, I.; Baulcombe, D.C. Virus-Induced Gene Silencing in Plants. Methods 2003, 30, 296–303. [Google Scholar] [CrossRef]
- Burch-Smith, T.M.; Anderson, J.C.; Martin, G.B.; Dinesh-Kumar, S.P. Applications and Advantages of Virus-Induced Gene Silencing for Gene Function Studies in Plants. Plant J. 2004, 39, 734–746. [Google Scholar] [CrossRef]
- Wangdi, T.; Uppalapati, S.R.; Nagaraj, S.; Ryu, C.-M.; Bender, C.L.; Mysore, K.S. A Virus-Induced Gene Silencing Screen Identifies a Role for Thylakoid Formation in Pseudomonas syringae Pv Tomato Symptom Development in Tomato and Arabidopsis. Plant Physiol. 2010, 152, 281–292. [Google Scholar] [CrossRef] [Green Version]
- Schachtsiek, J.; Hussain, T.; Azzouhri, K.; Kayser, O.; Stehle, F. Virus-Induced Gene Silencing (VIGS) in Cannabis Sativa L. Plant Methods 2019, 15, 157. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-C.; Jiang, C.-Z.; Gookin, T.; Hunter, D.; Clark, D.; Reid, M. Chalcone Synthase as a Reporter in Virus-Induced Gene Silencing Studies of Flower Senescence. Plant Mol. Biol. 2004, 55, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Cai, X.; Wang, X.; Zheng, Z.; Wang, C.; Cai, X.; Wang, X.; Zheng, Z. Optimisation of Tobacco Rattle Virus-Induced Gene Silencing in Arabidopsis. Funct. Plant Biol. 2006, 33, 347–355. [Google Scholar] [CrossRef]
- Liu, E.; Page, J.E. Optimized cDNA Libraries for Virus-Induced Gene Silencing (VIGS) Using Tobacco Rattle Virus. Plant Methods 2008, 4, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Park, M.; Jeong, E.S.; Lee, J.M.; Choi, D. Harnessing Anthocyanin-Rich Fruit: A Visible Reporter for Tracing Virus-Induced Gene Silencing in Pepper Fruit. Plant Methods 2017, 13, 3. [Google Scholar] [CrossRef] [Green Version]
- Quadrana, L.; Rodriguez, M.C.; López, M.; Bermúdez, L.; Nunes-Nesi, A.; Fernie, A.R.; Descalzo, A.; Asis, R.; Rossi, M.; Asurmendi, S.; et al. Coupling Virus-Induced Gene Silencing to Exogenous Green Fluorescence Protein Expression Provides a Highly Efficient System for Functional Genomics in Arabidopsis and across All Stages of Tomato Fruit Development. Plant Physiol. 2011, 156, 1278–1291. [Google Scholar] [CrossRef] [Green Version]
- Orzaez, D.; Medina, A.; Torre, S.; Fernández-Moreno, J.P.; Rambla, J.L.; Fernández-del-Carmen, A.; Butelli, E.; Martin, C.; Granell, A. A Visual Reporter System for Virus-Induced Gene Silencing in Tomato Fruit Based on Anthocyanin Accumulation. Plant Physiol. 2009, 150, 1122–1134. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Li, H.; Fu, D. Applications of Virus-Induced Gene Silencing for Identification of Gene Function in Fruit. Food Qual. Saf. 2021, 5, fyab018. [Google Scholar] [CrossRef]
- Marton, I.; Zuker, A.; Shklarman, E.; Zeevi, V.; Tovkach, A.; Roffe, S.; Ovadis, M.; Tzfira, T.; Vainstein, A. Nontransgenic Genome Modification in Plant Cells. Plant Physiol. 2010, 154, 1079–1087. [Google Scholar] [CrossRef] [Green Version]
- Ali, Z.; Abul-faraj, A.; Piatek, M.; Mahfouz, M.M. Activity and Specificity of TRV-Mediated Gene Editing in Plants. Plant Signal. Behav. 2015, 10, e1044191. [Google Scholar] [CrossRef]
- Ellison, E.E.; Nagalakshmi, U.; Gamo, M.E.; Huang, P.; Dinesh-Kumar, S.; Voytas, D.F. Multiplexed Heritable Gene Editing Using RNA Viruses and Mobile Single Guide RNAs. Nat. Plants 2020, 6, 620–624. [Google Scholar] [CrossRef] [PubMed]
- Honig, A.; Marton, I.; Rosenthal, M.; Smith, J.J.; Nicholson, M.G.; Jantz, D.; Zuker, A.; Vainstein, A. Transient Expression of Virally Delivered Meganuclease In Planta Generates Inherited Genomic Deletions. Mol. Plant 2015, 8, 1292–1294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaidi, S.S.-A.; Mansoor, S. Viral Vectors for Plant Genome Engineering. Front. Plant Sci. 2017, 8, 539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awan, M.J.A.; Amin, I.; Mansoor, S. Mini CRISPR-Cas12f1: A New Genome Editing Tool. Trends Plant Sci. 2021, 27, 110–112. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.Y.; Lee, J.M.; Moon, S.B.; Chin, H.J.; Park, S.; Lim, Y.; Kim, D.; Koo, T.; Ko, J.-H.; Kim, Y.-S. Efficient CRISPR Editing with a Hypercompact Cas12f1 and Engineered Guide RNAs Delivered by Adeno-Associated Virus. Nat. Biotechnol. 2021, 49, 94–102. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, Y.; Yu, H.; Pan, D.; Wang, Y.; Wang, Y.; Li, F.; Liu, C.; Nan, H.; Chen, W.; et al. Programmed Genome Editing by a Miniature CRISPR-Cas12f Nuclease. Nat. Chem. Biol. 2021, 17, 1132–1138. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Primer Number | Product | Forward Primer | Reverse Primer |
|---|---|---|---|
| 1 | PDS424 | 5′-AATTCTCGAGCTTCAGCTCCCACCAGAGTC-3’ | 5’-ATTCTAGATCACCGTCATCATCTTTCCA-3’ |
| 2 | PDS486 | 5’-AATTCTCGAGACTGGAAAGAGATTCCGTATTTCA-3’ | 5’-ATTCTAGAACAAAACCGCACCTTCCAT-3’ |
| 3 | TRV2-CP | 5′-ACGATTCTTGGGTGGAATCA-3′ | 5′-TCGTAACCGTTGTGTTTGGA-3′ |
| 4 | TRV2-DsRed | 5′-ACGATTCTTGGGTGGAATCA-3′ | 5′-CCCATGGTCTTCTTCTGCAT-3′ |
| 5 | PDS | 5′-ACTGTTCCTGATTGCGAACC-3′ | 5′-CTCGGCCAAAATTCTCTGAC-3′ |
| 6 | UBQ5 | 5′-AAGCTCGCTCTTCTCCAGTTC-3′ | 5′-CACACTTGCCGCAGTAATGTC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alter, H.; Peer, R.; Dombrovsky, A.; Flaishman, M.; Spitzer-Rimon, B. Tobacco Rattle Virus as a Tool for Rapid Reverse-Genetics Screens and Analysis of Gene Function in Cannabis sativa L. Plants 2022, 11, 327. https://doi.org/10.3390/plants11030327
Alter H, Peer R, Dombrovsky A, Flaishman M, Spitzer-Rimon B. Tobacco Rattle Virus as a Tool for Rapid Reverse-Genetics Screens and Analysis of Gene Function in Cannabis sativa L. Plants. 2022; 11(3):327. https://doi.org/10.3390/plants11030327
Chicago/Turabian StyleAlter, Hanan, Reut Peer, Aviv Dombrovsky, Moshe Flaishman, and Ben Spitzer-Rimon. 2022. "Tobacco Rattle Virus as a Tool for Rapid Reverse-Genetics Screens and Analysis of Gene Function in Cannabis sativa L." Plants 11, no. 3: 327. https://doi.org/10.3390/plants11030327
APA StyleAlter, H., Peer, R., Dombrovsky, A., Flaishman, M., & Spitzer-Rimon, B. (2022). Tobacco Rattle Virus as a Tool for Rapid Reverse-Genetics Screens and Analysis of Gene Function in Cannabis sativa L. Plants, 11(3), 327. https://doi.org/10.3390/plants11030327