
Innovating the Synergistic Assets of β-Amino Butyric Acid (BABA) and Selenium Nanoparticles (SeNPs) in Improving the Growth, Nitrogen Metabolism, Biological Activities, and Nutritive Value of Medicago interexta Sprouts

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
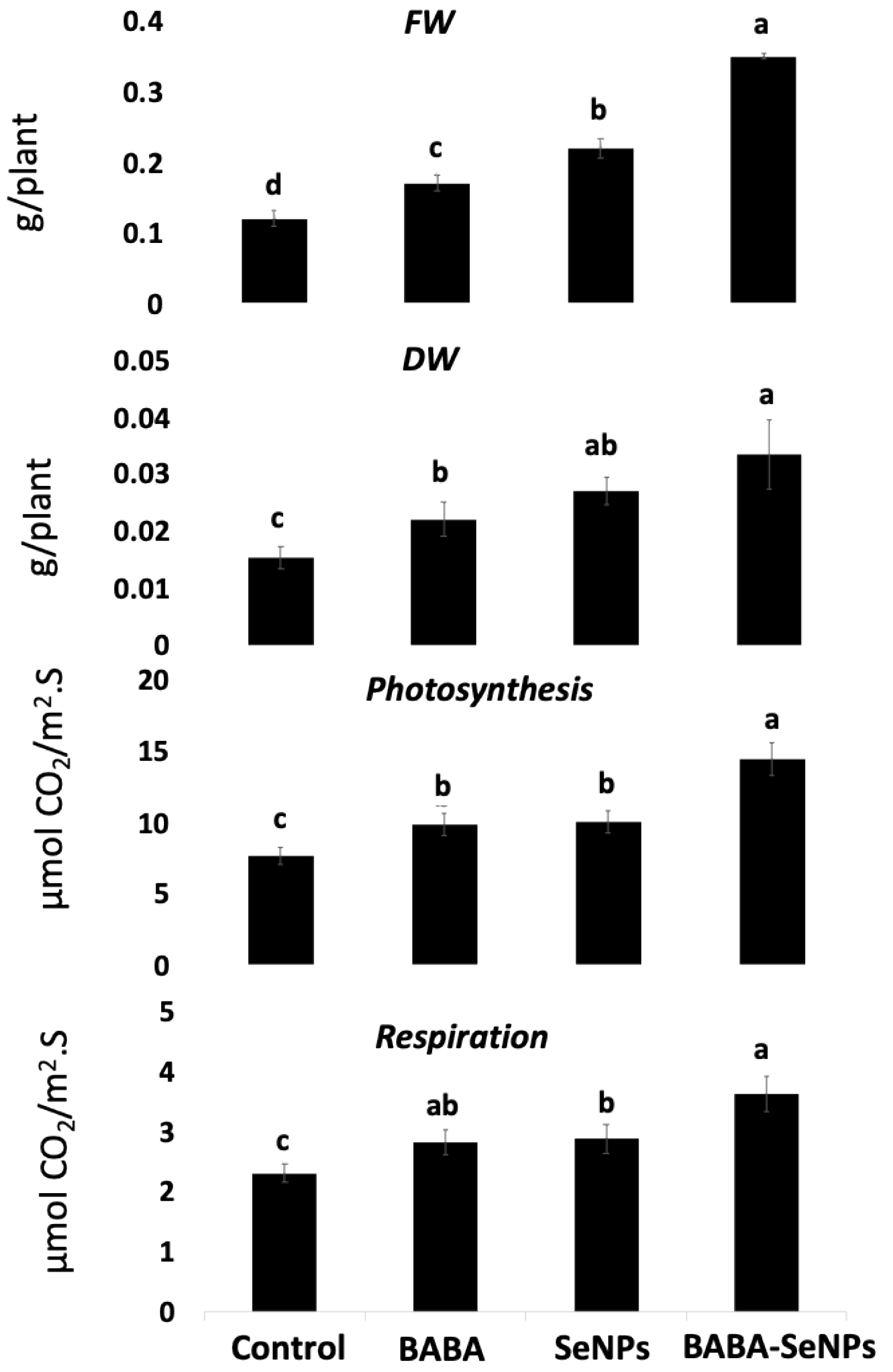
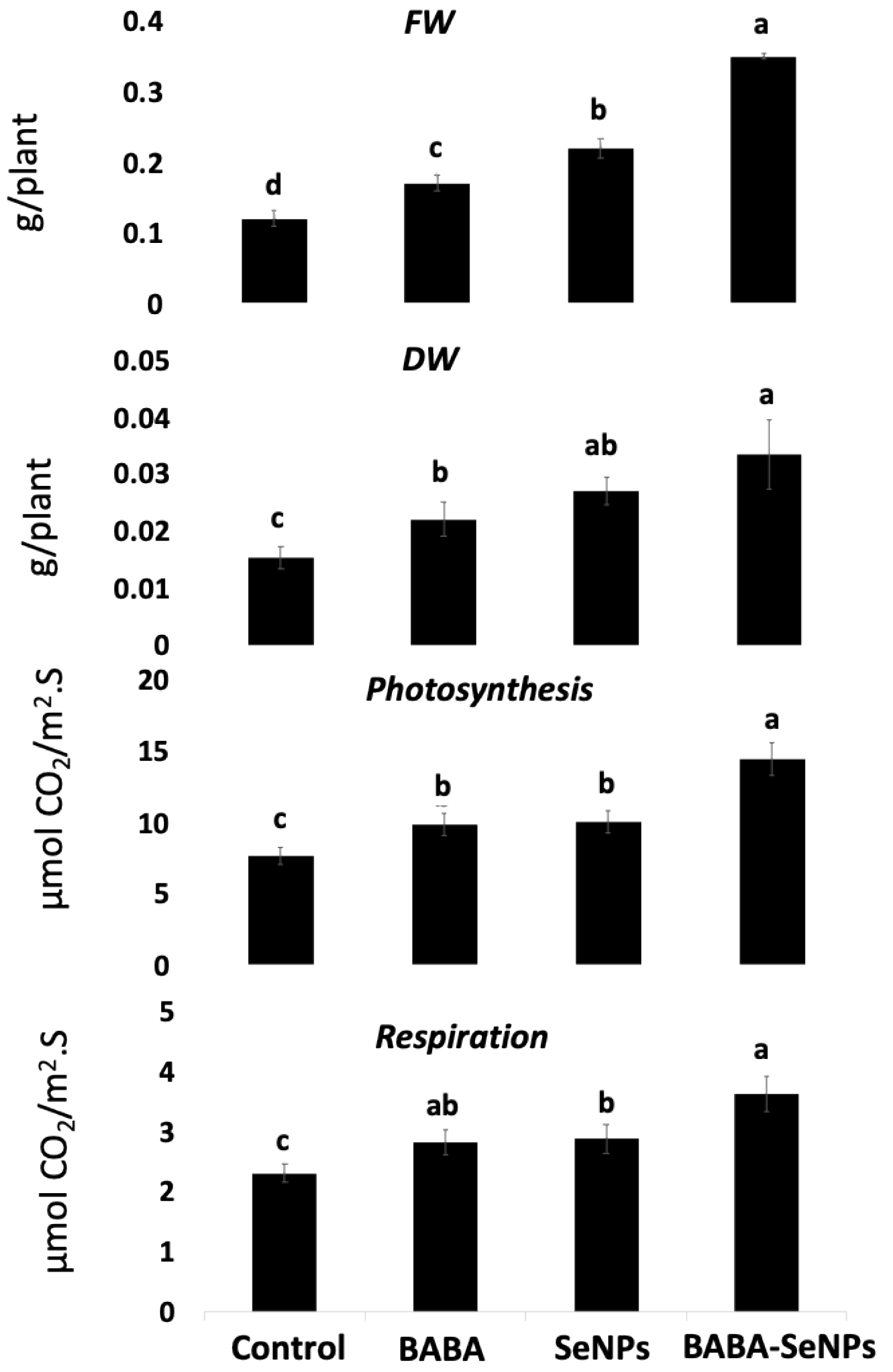
2.1. Enhanced Growth of M. interexta Sprouts under Sole and Combined Treatments with BABA and/or SeNPs
2.2. Combined Treatment of M. interexta Sprouts with BABA and SeNPs Induced a More Pronounced Effect on Mineral and Vitamin Profiles than That of a Sole Treatment
2.3. M. interexta Sprouts Were More Responsive to the Combined Effect of BABA and SeNPs on Nitrogen Metabolism than to Individual Treatments
2.4. Antioxidants of M. interexta Sprouts Were Improved by the Sole and Combined Treatments with BABA and/or SeNPs
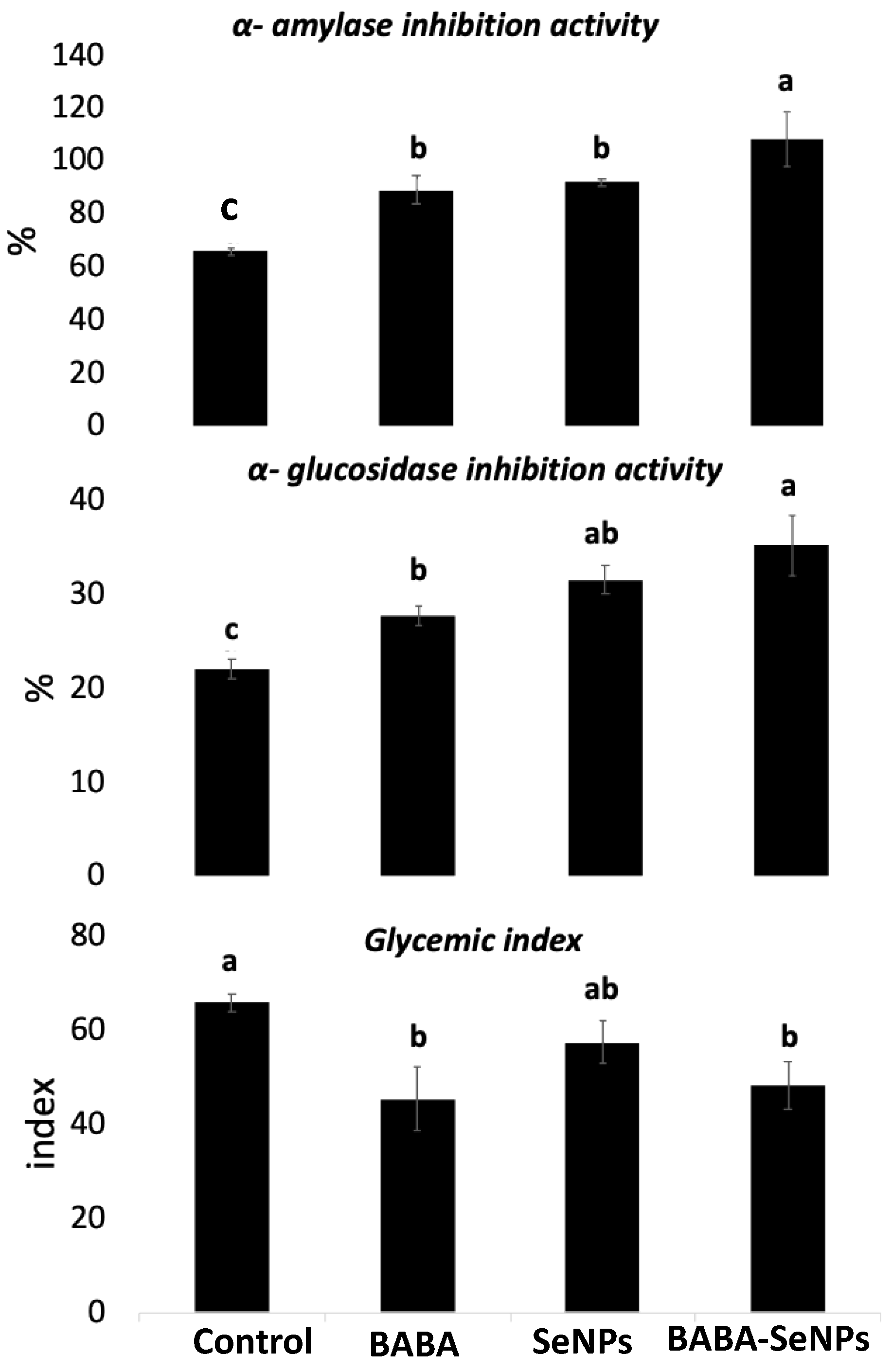
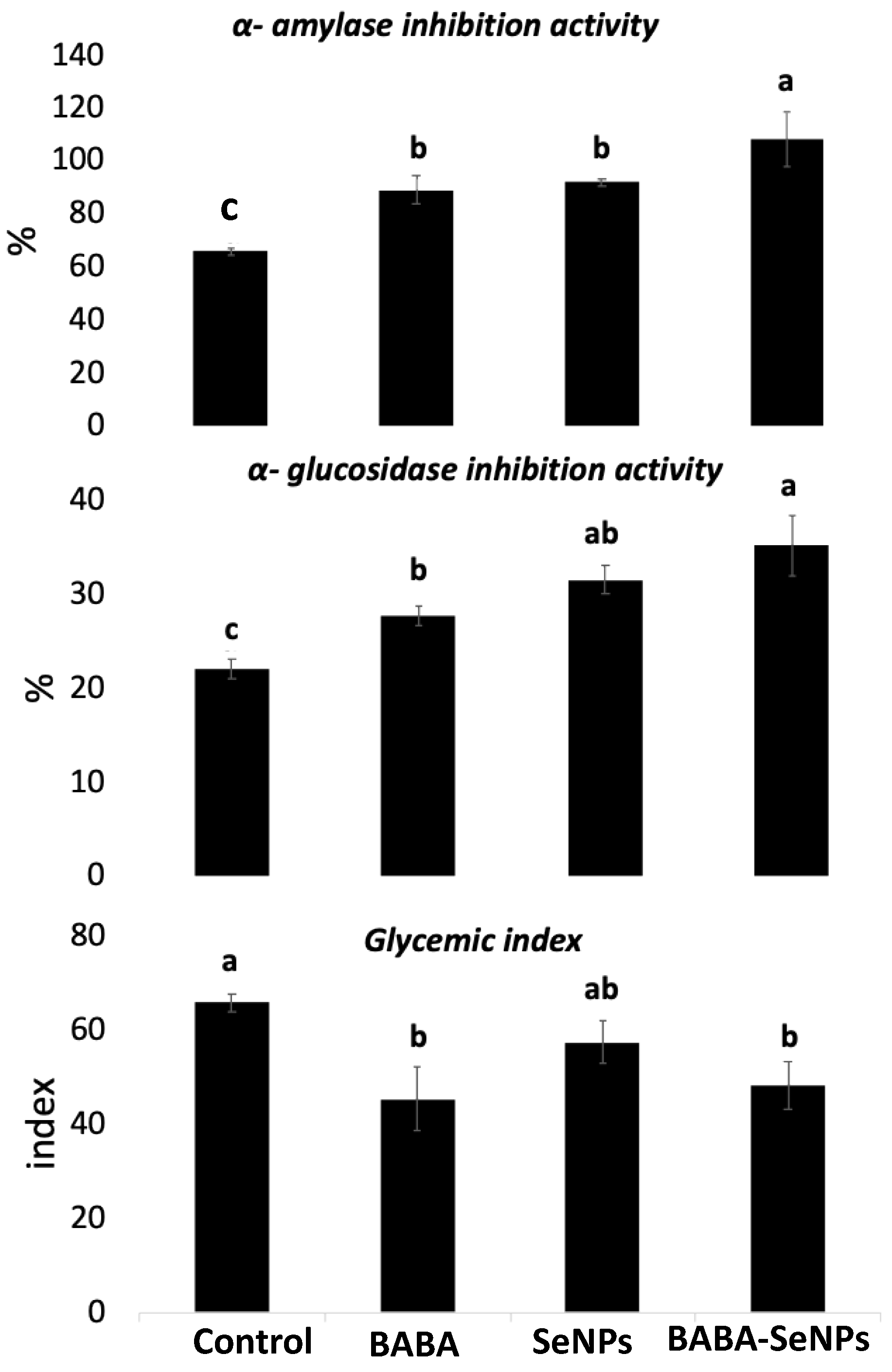
2.5. Anti-Diabetic Activity of M. interexta Sprouts Was More Improved by the Combined Treatment with BABA and Se NPs than by Individual Treatments
3. Discussion
3.1. Improved Growth of M. Interexta Sprouts
3.2. Improved Pigment Content of M. interexta Sprouts
3.3. Improved Mineral Content and Vitamin Profile of M. interexta Sprouts
3.4. Improved Nitrogen Metabolism of M. interexta Sprouts
3.5. Improved Antioxidants of M. interexta Sprouts
3.6. Improved Antidiabetic Activity of M. interexta Sprouts
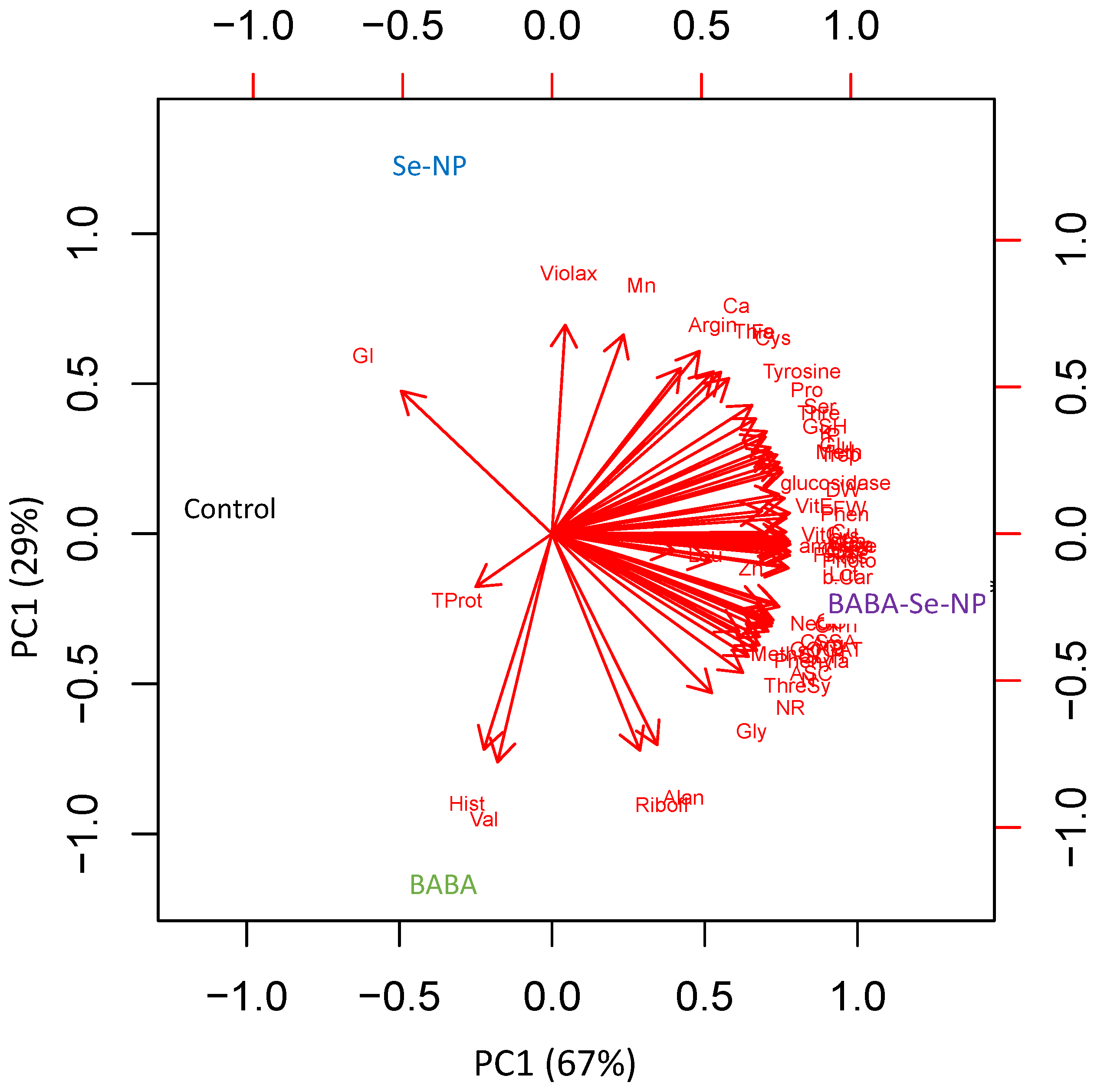
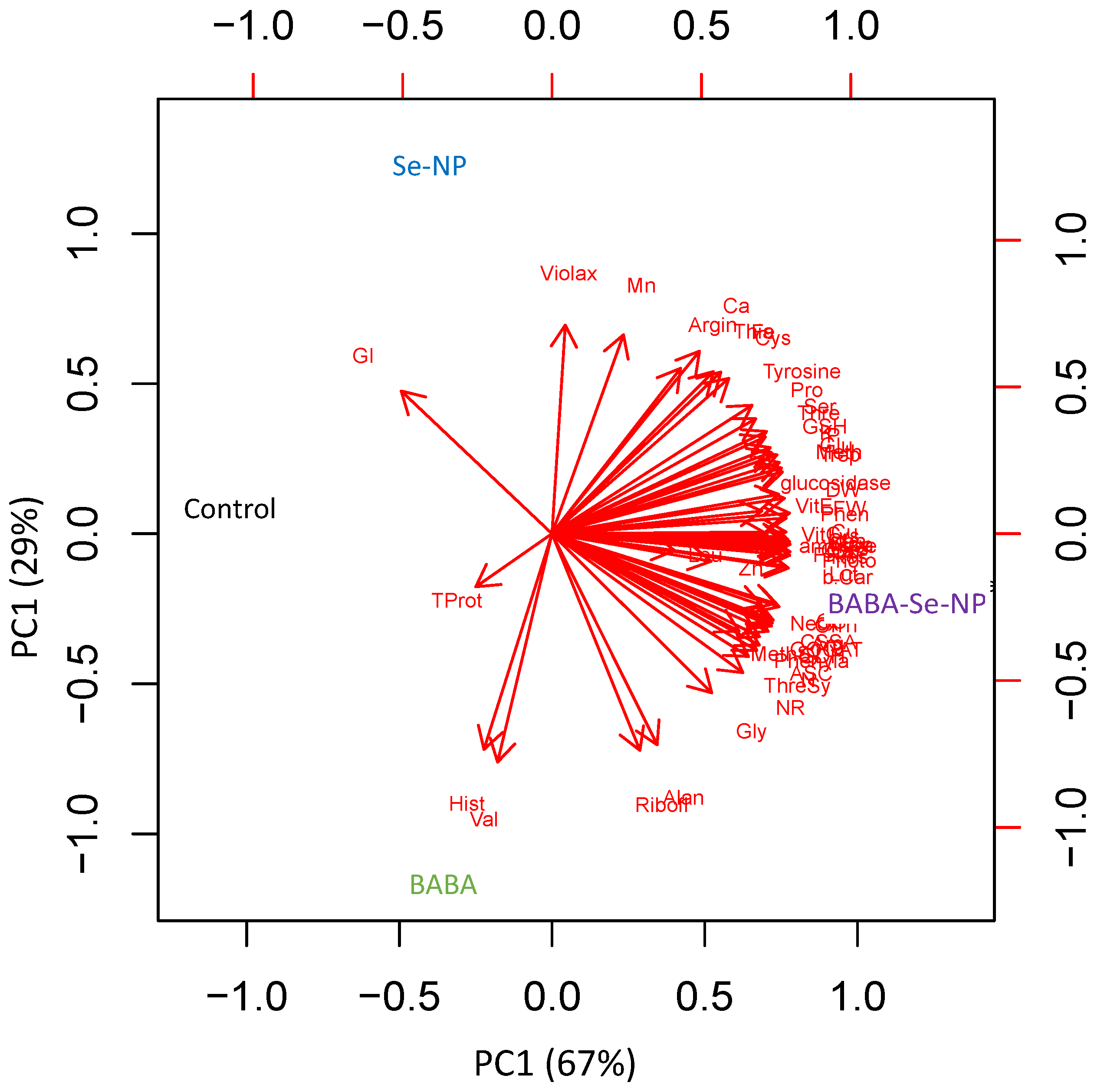
3.7. Species-Specific Response to BABA and/or SeNPs
4. Materials and Methods
4.1. Experimental Setup
4.2. Selenium Nanoparticles Characterization
4.3. Determination of Photosynthetic Rate
4.4. Pigment Analysis
4.5. Analysis of Mineral Contents
4.6. Determination of Phenolic, Flavonoid Contents, and Vitamins Levels
4.6.1. Total Antioxidant Capacity (FRAP)
4.6.2. Amino Acid Analysis
4.7. Determination of Nitrogen Content and Metabolism
4.8. Determination of Antidiabetic Activity
4.9. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haminiuk, C.W.I.; Maciel, G.M.; Plata-Oviedo, M.S.V.; Peralta, R.M. Phenolic compounds in fruits–an overview. Int. J. Food Sci. Technol. 2012, 47, 2023–2044. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Kostov, R.V. Glucosinolates and isothiocyanates in health and disease. Trends Mol. Med. 2012, 18, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Björkman, M.; Klingen, I.; Birch, A.N.E.; Bones, A.M.; Bruce, T.J.A.; Johansen, T.J.; Meadow, R.; Mølmann, J.; Seljåsen, R.; Smart, L.E. Phytochemicals of Brassicaceae in plant protection and human health–Influences of climate, environment and agronomic practice. Phytochemistry 2011, 72, 538–556. [Google Scholar] [CrossRef] [PubMed]
- Poulev, A.; O’Neal, J.M.; Logendra, S.; Pouleva, R.B.; Timeva, V.; Garvey, A.S.; Gleba, D.; Jenkins, I.S.; Halpern, B.T.; Kneer, R. Elicitation, a new window into plant chemodiversity and phytochemical drug discovery. J. Med. Chem. 2003, 46, 2542–2547. [Google Scholar] [CrossRef]
- Smetanska, I. Production of secondary metabolites using plant cell cultures. Food Biotechnol. 2008, 111, 187–228. [Google Scholar]
- Zhao, J.; Davis, L.C.; Verpoorte, R. Elicitor signal transduction leading to production of plant secondary metabolites. Biotechnol. Adv. 2005, 23, 283–333. [Google Scholar] [CrossRef]
- Ferrari, S. Biological elicitors of plant secondary metabolites: Mode of action and use in the production of nutraceutics. In Bio-Farms for Nutraceuticals; Springer: Berlin/Heidelberg, Germany, 2010; pp. 152–166. [Google Scholar]
- Thevenet, D.; Pastor, V.; Baccelli, I.; Balmer, A.; Vallat, A.; Neier, R.; Glauser, G.; Mauch-Mani, B. The priming molecule β-aminobutyric acid is naturally present in plants and is induced by stress. New Phytol. 2017, 213, 552–559. [Google Scholar] [CrossRef]
- Jakab, G.; Cottier, V.; Toquin, V.; Rigoli, G.; Zimmerli, L.; Métraux, J.-P.; Mauch-Mani, B. β-Aminobutyric acid-induced resistance in plants. Eur. J. Plant Pathol. 2001, 107, 29–37. [Google Scholar] [CrossRef]
- Cohen, Y.R. β-aminobutyric acid-induced resistance against plant pathogens. Plant Dis. 2002, 86, 448–457. [Google Scholar] [CrossRef] [Green Version]
- Shaw, A.K.; Bhardwaj, P.K.; Ghosh, S.; Roy, S.; Saha, S.; Sherpa, A.R.; Saha, S.K.; Hossain, Z. β-aminobutyric acid mediated drought stress alleviation in maize (Zea mays L.). Environ. Sci. Pollut. Res. 2016, 23, 2437–2453. [Google Scholar] [CrossRef]
- Mátai, A.; Jakab, G.; Hideg, É. Single-dose β-aminobutyric acid treatment modifies tobacco (Nicotiana tabacum L.) leaf acclimation to consecutive UV-B treatment. Photochem. Photobiol. Sci. 2019, 18, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Giraldo, J.P.; Landry, M.P.; Faltermeier, S.M.; McNicholas, T.P.; Iverson, N.M.; Boghossian, A.A.; Reuel, N.F.; Hilmer, A.J.; Sen, F.; Brew, J.A.; et al. Plant nanobionics approach to augment photosynthesis and biochemical sensing. Nat. Mater. 2014, 13, 400–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majumder, D.D.; Ulrichs, C.; Majumder, D.; Mewis, I.; Thakur, A.R.; Brahmachary, R.L.; Banerjee, R.; Rahman, A.; Debnath, N.; Seth, D. Current status and future trends of nanoscale technology and its impact on modern computing, biology, medicine and agricultural biotechnology. In Proceedings of the 2007 International Conference on Computing: Theory and Applications (ICCTA’07), Kolkata, India, 5–7 March 2007; pp. 563–573. [Google Scholar]
- Siddiqui, M.H.; Al-Whaibi, M.H. Role of nano-SiO2 in germination of tomato (Lycopersicum esculentum seeds Mill.). Saudi J. Biol. Sci. 2014, 21, 13–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terry, N.; Zayed, A.M.; De Souza, M.P.; Tarun, A.S. Selenium in higher plants. Annu. Rev. Plant Biol. 2000, 51, 401–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Germ, M.; Stibilj, V. Selenium and plants. Acta Agric. Slov. 2007, 89, 65–71. [Google Scholar] [CrossRef]
- Kuznetsov, V.V.; Kholodova, V.P.; Kuznetsov, V.V.; Yagodin, B.A. Selenium regulates the water status of plants exposed to drought. In Doklady Biological Sciences; Consultants Bureau: New York, NY, USA, 2003; pp. 266–268. [Google Scholar]
- Ferrarese, M.; Sourestani, M.M.; Quattrini, E.; Schiavi, M.; Ferrante, A. Biofortification of spinach plants applying selenium in the nutrient solution of floating system. Veg. Crop. Res. Bull. 2012, 2012, 127–136. [Google Scholar] [CrossRef]
- Guevara Moreno, O.D.; Acevedo Aguilar, F.J.; Yanez Barrientos, E. Selenium uptake and biotransformation and effect of selenium exposure on the essential and trace elements status: Comparative evaluation of four edible plants. J. Mex. Chem. Soc. 2018, 62, 247–258. [Google Scholar] [CrossRef]
- Li, X.; Xu, H.; Chen, Z.-S.; Chen, G. Biosynthesis of nanoparticles by microorganisms and their applications. J. Nanomater. 2011, 2011, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Zohra, E.; Ikram, M.; Omar, A.A.; Hussain, M.; Satti, S.H.; Raja, N.I.; Ehsan, M. Potential applications of biogenic selenium nanoparticles in alleviating biotic and abiotic stresses in plants: A comprehensive insight on the mechanistic approach and future perspectives. Green Process. Synth. 2021, 10, 456–475. [Google Scholar] [CrossRef]
- Rady, M.O.A.; Semida, W.M.; Abd El-Mageed, T.A.; Howladar, S.M.; Shaaban, A. Foliage applied selenium improves photosynthetic efficiency, antioxidant potential and wheat productivity under drought stress. Int. J. Agric. Biol 2020, 24, 1293–1300. [Google Scholar]
- Chang, K.C.; Satterlee, L.D. Chemical, nutritional and microbiological quality of a protein concentrate from culled dry beans. J. Food Sci. 1979, 44, 1589–1593. [Google Scholar] [CrossRef]
- Bakoğlu, A.; Bağcı, E.; Koçak, A.; Yüce, E. Fatty acid composition of some Medicago L. (Fabaceae) species from Turkey. Asian J. Chem. 2010, 2, 651–656. [Google Scholar]
- Kokten, K.; Bakoglu, A.; Kocak, A.; Bagci, E.; Akcura, M.; Kaplan, M. Chemical composition of the seeds of some Medicago species. Chem. Nat. Compd. 2011, 47, 619–621. [Google Scholar] [CrossRef]
- Jisha, K.C.; Puthur, J.T. Seed priming with beta-amino butyric acid improves abiotic stress tolerance in rice seedlings. Rice Sci. 2016, 23, 242–254. [Google Scholar] [CrossRef] [Green Version]
- El Lateef Gharib, F.A.; Zeid, I.M.; Ghazi, S.M.; Ahmed, E.Z. The response of cowpea (Vigna unguiculata L) plants to foliar application of sodium selenate and selenium nanoparticles (SeNPs). J. Nanomater. Mol. Nanotechnol. 2019, 8, 1000272. [Google Scholar]
- Wang, H.; Zhang, J.; Yu, H. Elemental selenium at nano size possesses lower toxicity without compromising the fundamental effect on selenoenzymes: Comparison with selenomethionine in mice. Free Radic. Biol. Med. 2007, 42, 1524–1533. [Google Scholar] [CrossRef]
- Morales-Espinoza, M.C.; Cadenas-Pliego, G.; Pérez-Alvarez, M.; Hernández-Fuentes, A.D.; Cabrera de la Fuente, M.; Benavides-Mendoza, A.; Valdés-Reyna, J.; Juárez-Maldonado, A. Se nanoparticles induce changes in the growth, antioxidant responses, and fruit quality of tomato developed under NaCl Stress. Molecules 2019, 24, 3030. [Google Scholar] [CrossRef] [Green Version]
- Haghighi, M.; Abolghasemi, R.; Teixeira da Silva, J.A. Low and high temperature stress affect the growth characteristics of tomato in hydroponic culture with Se and nano-Se amendment. Sci. Hortic. 2014, 178, 231–240. [Google Scholar] [CrossRef]
- Ragavan, P.; Ananth, A.; Rajan, M.R. Impact of selenium nanoparticles on growth, biochemical characteristics and yield of cluster bean Cyamopsis tetragonoloba. Int. J. Environ. Agric. Biotechnol. 2017, 2, 238983. [Google Scholar] [CrossRef]
- Wu, C.C.; Singh, P.; Chen, M.C.; Zimmerli, L. L-Glutamine inhibits beta-aminobutyric acid-induced stress resistance and priming in Arabidopsis. J. Exp. Bot. 2010, 61, 995–1002. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, A.; Kumar, A.; Kaur, H.; Balamurugan, A.; Padhy, A.K.; Mehta, S. Plant Performance and Defensive Role of β-Amino Butyric Acid Under Environmental Stress. In Plant Performance under Environmental Stress; Springer: Berlin/Heidelberg, Germany, 2021; pp. 249–275. [Google Scholar]
- Shafique, H.; Jamil, Y.; ul Haq, Z.; Mujahid, T.; Khan, A.U.; Iqbal, M.; Abbas, M. Low power continuous wave-laser seed irradiation effect on Moringa oleifera germination, seedling growth and biochemical attributes. J. Photochem. Photobiol. B Biol. 2017, 170, 314–323. [Google Scholar]
- Cao, S.; Jiang, L.; Yuan, H.; Jian, H.; Ren, G.; Bian, X.; Zou, J.; Chen, Z. β-Amino-butyric acid protects Arabidopsis against low potassium stress. Acta Physiol. Plant. 2008, 30, 309–314. [Google Scholar] [CrossRef]
- Jiang, L.; Yang, R.Z.; Lu, Y.F.; Cao, S.Q.; Ci, L.K.; Zhang, J.J. β-aminobutyric acid-mediated tobacco tolerance to potassium deficiency. Russ. J. Plant Physiol. 2012, 59, 781–787. [Google Scholar] [CrossRef]
- Erenoglu, E.B.; Kutman, U.B.; Ceylan, Y.; Yildiz, B.; Cakmak, I. Improved nitrogen nutrition enhances root uptake, root-to-shoot translocation and remobilization of zinc (65Zn) in wheat. New Phytol. 2011, 189, 438–448. [Google Scholar] [CrossRef] [PubMed]
- Broadley, M.R.; White, P.J.; Hammond, J.P.; Zelko, I.; Lux, A. Zinc in plants. New Phytol. 2007, 173, 677–702. [Google Scholar] [CrossRef] [PubMed]
- Pandey, C.; Gupta, M. Selenium amelioration of arsenic toxicity in rice shows genotypic variation: A transcriptomic and biochemical analysis. J. Plant Physiol. 2018, 231, 168–181. [Google Scholar] [CrossRef]
- Poldma, P.; Tonutare, T.; Viitak, A.; Luik, A.; Moor, U. Effect of selenium treatment on mineral nutrition, bulb size, and antioxidant properties of garlic (Allium sativum L.). J. Agric. Food Chem. 2011, 59, 5498–5503. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liang, D.; Song, W.; Lei, L.; Yu, D.; Miao, S. Effects of nitrogen application on selenium accumulation, translocation and distribution of winter wheat at different growth periods. J. Plant Nutr. Fert. 2012, 22, 395–402. [Google Scholar]
- Helali, S.M.; Nebli, H.; Kaddour, R.; Mahmoudi, H.; Lachaâl, M.; Ouerghi, Z. Influence of nitrate—Ammonium ratio on growth and nutrition of Arabidopsis thaliana. Plant Soil 2010, 336, 65–74. [Google Scholar] [CrossRef]
- Okla, M.K.; Akhtar, N.; Alamri, S.A.; Al-Qahtani, S.M.; Ismail, A.; Abbas, Z.K.; Al-Ghamdi, A.A.; Qahtan, A.A.; Soufan, W.H.; Alaraidh, I.A. Potential importance of molybdenum priming to metabolism and nutritive value of Canavalia spp. sprouts. Plants 2021, 10, 2387. [Google Scholar] [CrossRef]
- Janse van Rensburg, H.C.; Van den Ende, W. Priming with γ-aminobutyric acid against Botrytis cinerea reshuffles metabolism and reactive oxygen species: Dissecting signalling and metabolism. Antioxidants 2020, 9, 1174. [Google Scholar] [CrossRef] [PubMed]
- Winter, G.; Todd, C.D.; Trovato, M.; Forlani, G.; Funck, D. Physiological implications of arginine metabolism in plants. Front. Plant Sci. 2015, 6, 534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddappa, S.; Marathe, G.K. What we know about plant arginases? Plant Physiol. Biochem. 2020, 156, 600–610. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Wang, B.; Yan, J.; Cheng, L.; Yao, L.; Xiao, L.; Wu, T. DL-β-aminobutyric acid-induced resistance in soybean against Aphis glycines Matsumura (Hemiptera: Aphididae). PLoS ONE 2014, 9, e85142. [Google Scholar] [CrossRef] [Green Version]
- Baccelli, I.; Mauch-Mani, B. Beta-aminobutyric acid priming of plant defense: The role of ABA and other hormones. Plant Mol. Biol. 2016, 91, 703–711. [Google Scholar] [CrossRef]
- Jóźwiak, W.; Politycka, B. Effect of selenium on alleviating oxidative stress caused by a water deficit in cucumber roots. Plants 2019, 8, 217. [Google Scholar] [CrossRef] [Green Version]
- Menon, S.; Devi, K.S.S.; Agarwal, H.; Shanmugam, V.K. Efficacy of biogenic selenium nanoparticles from an extract of ginger towards evaluation on anti-microbial and anti-oxidant activities. Colloid Interface Sci. Commun. 2019, 29, 1–8. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Abdelrahman, M.; Hosseini, M.S.; Hoveizeh, N.F.; Tran, L.-S.P. Alleviation of the effect of salinity on growth and yield of strawberry by foliar spray of selenium-nanoparticles. Environ. Pollut. 2019, 253, 246–258. [Google Scholar] [CrossRef]
- Baysal, Ö.; Gürsoy, Y.Z.; Örnek, H.; Duru, A. Induction of oxidants in tomato leaves treated with DL-β-Amino butyric acid (BABA) and infected with Clavibacter michiganensis ssp. michiganensis. Eur. J. Plant Pathol. 2005, 112, 361–369. [Google Scholar] [CrossRef]
- Astaneh, R.K.; Bolandnazar, S.; Nahandi, F.Z.; Oustan, S. Effect of selenium application on phenylalanine ammonia-lyase (PAL) activity, phenol leakage and total phenolic content in garlic (Allium sativum L.) under NaCl stress. Inf. Process. Agric. 2018, 5, 339–344. [Google Scholar] [CrossRef]
- Altamiranda, E.A.G.; Andreu, A.B.; Daleo, G.R.; Olivieri, F.P. Effect of β-aminobutyric acid (BABA) on protection against Phytophthora infestans throughout the potato crop cycle. Australas. Plant Pathol. 2008, 37, 421–427. [Google Scholar] [CrossRef]
- Habeeb, T.H.; Abdel-Mawgoud, M.; Yehia, R.S.; Khalil, A.M.A.; Saleh, A.M.; AbdElgawad, H. Interactive Impact of Arbuscular Mycorrhizal Fungi and Elevated CO2 on Growth and Functional Food Value of Thymus vulgare. J. Fungi 2020, 6, 168. [Google Scholar] [CrossRef]
- Saleh, A.M.; Abdel-Mawgoud, M.; Hassan, A.R.; Habeeb, T.H.; Yehia, R.S.; AbdElgawad, H. Global metabolic changes induced by arbuscular mycorrhizal fungi in oregano plants grown under ambient and elevated levels of atmospheric CO2. Plant Physiol. Biochem. 2020, 151, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Almuhayawi, M.S.; Hassan, A.H.A.; Abdel-Mawgoud, M.; Khamis, G.; Selim, S.; Al Jaouni, S.K.; AbdElgawad, H. Laser light as a promising approach to improve the nutritional value, antioxidant capacity and anti-inflammatory activity of flavonoid-rich buckwheat sprouts. Food Chem. 2020, 345, 128788. [Google Scholar] [CrossRef] [PubMed]
- Crawford, N.M.; Brian, G.F. Molecular and developmental biology of inorganic nitrogen nutrition. In The Arabidopsis Book/American Society of Plant Biologists 1; SSPA Sweden AB: Gothenburg, Sweden, 2002; p. e0011. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, P.; Das, C.; Biswas, S.; Nag, S.K.; Dutta, A.; Biswas, M.; Sil, S.; Hazra, L.; Ghosh, C.; Das, S.; et al. Phytochemical composition analysis and evaluation of in vitro medicinal properties and cytotoxicity of five wild weeds: A comparative study. F1000Research 2020, 9, 493. [Google Scholar] [CrossRef] [PubMed]
- Thayer, S.S.; Björkman, O. Leaf Xanthophyll content and composition in sun and shade determined by HPLC. Photosynth. Res. 1990, 23, 331–343. [Google Scholar] [CrossRef] [PubMed]
- AbdElgawad, H.; Okla, M.K.; Al-Amri, S.S.; Al-Hashimi, A.; Al-Qahtani, W.H.; Al-Qahtani, S.M.; Abbas, Z.K.; Al-Harbi, N.A.; Abd Algafar, A.; Almuhayawi, M.S. Effect of elevated CO2 on biomolecules’ accumulation in caraway (Carum carvi L.) plants at different developmental stages. Plants 2021, 10, 2434. [Google Scholar] [CrossRef]
- Almuhayawi, M.S.; Abdel-Mawgoud, M.; Al Jaouni, S.K.; Almuhayawi, S.M.; Alruhaili, M.H.; Selim, S.; AbdElgawad, H. Bacterial endophytes as a promising approach to enhance the growth and accumulation of bioactive metabolites of three species of Chenopodium Sprouts. Plants 2021, 10, 2745. [Google Scholar] [CrossRef]
- Almuhayawi, M.S.; Mohamed, M.S.M.; Abdel-Mawgoud, M.; Selim, S.; Al Jaouni, S.K.; AbdElgawad, H. Bioactive potential of several actinobacteria isolated from microbiologically barely explored desert habitat, Saudi Arabia. Biology 2021, 10, 235. [Google Scholar] [CrossRef]
- Almuhayawi, M.S.; Al Jaouni, S.K.; Almuhayawi, S.M.; Selim, S.; Abdel-Mawgoud, M. Elevated CO2 improves the nutritive value, antibacterial, anti-inflammatory, antioxidant and hypocholestecolemic activities of lemongrass sprouts. Food Chem. 2021, 357, 129730. [Google Scholar] [CrossRef]
- Sinha, A.K.; Giblen, T.; AbdElgawad, H.; De Rop, M.; Asard, H.; Blust, R.; De Boeck, G. Regulation of amino acid metabolism as a defensive strategy in the brain of three freshwater teleosts in response to high environmental ammonia exposure. Aquat. Toxicol. 2013, 130–131, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.H.; Cho, Y.D. Purification and properties of arginase from soybean, Glycine max, axes. Plant Physiol. 1990, 93, 1230–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Zinta, G.; AbdElgawad, H.; Peshev, D.; Weedon, J.T.; Van den Ende, W.; Nijs, I.; Janssens, I.A.; Beemster, G.T.S.; Asard, H. Dynamics of metabolic responses to periods of combined heat and drought in Arabidopsis thaliana under ambient and elevated atmospheric CO2. J. Exp. Bot. 2018, 69, 2159–2170. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Control | BABA | SeNPs | BABA-SeNPs | |
|---|---|---|---|---|
| Chl a | 0.65 ± 0.06 c | 0.92 ± 0.02 b | 1.05 ± 0.2 b | 1.97 ± 0.17 a |
| Chl b | 0.43 ± 0.069 c | 0.53 ± 0.08 bc | 0.59 ± 0.116 b | 1.18 ± 0.19 a |
| β-Carotene | 0.04 ± 0.01 c | 0.07 ± 0.004 b | 0.07 ± 0.017 b | 0.11 ± 0.01 a |
| Lutein | 0.14 ± 0.03 c | 0.24 ± 0.02 b | 0.23 ± 0.02 b | 0.53 ± 0.03 a |
| Neoxanthin | 0.02 ± 0.01 c | 0.02 ± 0.003 b | 0.01 ± 0.001 c | 0.05 ± 0.007 a |
| Violaxanthin | 0.05 ± 0.01 c | 0.04 ± 0.003 b | 0.07 ± 0.009 a | 0.05 ± 0.001 b |
| Parameters | Control | BABA | SeNPs | BABA-SeNPs |
|---|---|---|---|---|
| Elements | ||||
| Ca | 17.57 ± 2.3 b | 15.79 ± 3.5 b | 27.79 ± 6.7 a | 25.17 ± 0.47 a |
| Cu | 2.26 ± 0.71 b | 2.57 ± 1.07 b | 2.87 ± 0.28 b | 4.38 ± 1.1 a |
| Fe | 3.99 ± 0.23 b | 3.15 ± 0.78 b | 5.48 ± 1.02 a | 5.76 ± 0.44 a |
| Zn | 22.62 ± 2.0 b | 36.62 ± 3.3 a | 35.88 ± 3.2 a | 35.72 ± 3.2 a |
| Mn | 0.25 ± 0.03 a | 0.13 ± 0.1 b | 0.28 ± 0.13 a | 0.27 ± 0.1 a |
| K | 15.60 ± 1.3 c | 11.95 ± 3 c | 40.60 ± 3.6 b | 67.29 ± 6 a |
| P | 5.81 ± 0.6 c | 6.48 ± 0.5 c | 10.44 ± 0.8 b | 13.56 ± 1.1 a |
| Vitamins | ||||
| Vit C | 7.81 ± 1.3 b | 7.31 ± 1.2 b | 8.15 ± 2.4 b | 13.92 ± 0.7 a |
| Vit E | 47.47 ± 1.2 b | 44.57 ± 1.6 cb | 48.47 ± 4.4 b | 61.92 ± 3.9 a |
| Thiamin | 0.10 ± 0 b | 0.07 ± 0 b | 0.13 ± 0.02 a | 0.14 ± 0.06 a |
| Riboflavin | 0.35 ± 0.3 b | 0.51 ± 0.75 a | 0.24 ± 0.47 b | 0.49 ± 0.96 a |
| Amino Acids | Control | BABA | SeNPs | BABA-SeNPs |
|---|---|---|---|---|
| Asparagine | 1.53 ± 0.1 b | 1.71 ± 0.06 b | 1.76 ± 0.02 b | 2.17 ± 0.01 a |
| Glutamine | 1.89 ± 0.19 c | 2.15 ± 0.25 c | 3.39 ± 0.08 b | 4.53 ± 0.12 a |
| Serine | 1.18 ± 0.07 c | 1.36 ± 0.13 ab | 2.31 ± 0.13 b | 2.66 ± 0.3 a |
| Glycine | 1.40 ± 0.01 c | 1.69 ± 0.07 b | 1.16 ± 0.1 c | 2.01 ± 0.01 a |
| Arginine | 0.30 ± 0.05 c | 0.38 ± 0.08 c | 0.77 ± 0.05 a | 0.57 ± 0.08 b |
| Alanine | 0.54 ± 0.03 b | 0.62 ± 0.03 a | 0.51 ± 0 b | 0.61 ± 0.02 a |
| Proline | 0.93 ± 0.01 c | 1.25 ± 0.03 b | 2.54 ± 0.06 a | 2.71 ± 0.18 a |
| Histidine | 0.75 ± 0.05 b | 0.90 ± 0.05 a | 0.67 ± 0.04 b | 0.72 ± 0.09 b |
| Valine | 0.76 ± 0.15 b | 0.91 ± 0.2 a | 0.61 ± 0.09 b | 0.73 ± 0.11 b |
| Methionine | 0.66 ± 0.09 c | 0.75 ± 0.05 c | 0.97 ± 0.01 b | 1.17 ± 0.1 a |
| Cystine | 0.99 ± 0.14 b | 0.79 ± 0.15 b | 1.47 ± 0.08 a | 1.56 ± 0.04 a |
| Ornithine | 1.17 ± 0.18 c | 2.10 ± 0.21 b | 1.72 ± 0.04 b | 3.06 ± 0.1 a |
| Leucine | 0.98 ± 0.06 a | 0.86 ± 0.18 a | 0.86 ± 0.07 a | 1.07 ± 0.12 a |
| Phenylalanine | 1.42 ± 0.22 b | 1.87 ± 0.23 a | 1.65 ± 0.11 b | 2.04 ± 0.11 a |
| Tyrosine | 0.31 ± 0.04 a | 0.30 ± 0 a | 0.42 ± 0.01 ab | 0.45 ± 0.01 ab |
| Lysine | 0.70 ± 0.02 b | 0.88 ± 0.02 b | 1.02 ± 0.03 b | 1.91 ± 0.03 a |
| Threonine | 1.18 ± 0.05 b | 1.32 ± 0.03 b | 1.68 ± 0.08 a | 1.78 ± 0.09 a |
| Treptophane | 0.72 ± 0.08 b | 0.83 ± 0.1 b | 1.06 ± 0.02 ab | 1.28 ± 0.04 a |
| Control | BABA | SeNPs | BABA-SeNPs | |
|---|---|---|---|---|
| Nitrogen | 23.3 ± 0.8 b | 35.7 ± 0.5 a | 28.1 ± 1.2 b | 41.2 ± 0.8 a |
| Total Protein | 169.5 ± 1.9 a | 118.0 ± 3.1 d | 99.6 ± 2.2 c | 136 ± 2.8 b |
| Nitrate reductase | 45.2 ± 0.03 c | 86.1 ± 5.4 b | 43.1 ± 2.2 c | 118 ± 11 a |
| GDH | 4.14 ± 0.2 c | 6.99 ± 0.48 b | 4.9 ± 0.21 c | 10 ± 0.48 a |
| GOGAT | 7.8 ± 0.28 d | 14.35 ± 0.4 b | 10.3± 0.2 c | 21 ± 1.8 a |
| GS | 16.12 ± 0.9 d | 26.10 ± 0.4 c | 23.0 ± 1 b | 32 ± 0.8 a |
| Cyst syn ser acetyltransferase | 6.7 ± 0.28 d | 11.05 ± 0.0 b | 9.0 ± 0.4 c | 14.2 ± 0.38 a |
| Arginase | 4.01 ± 0.02 d | 7.7 ± 0.46 b | 5.9 ± 0.2 cd | 10.7 ± 0.9 a |
| Threonine synthase | 1.0 ± 0.02 c | 1.70 ± 0.1 b | 0.9 ± 0.04 c | 2.6 ± 0.17 a |
| Methionine synthase | 2.0 ± 0.01 c | 4.30 ± 0.05 a | 3.40 ± 0.1 b | 4.4 ± 0.2 a |
| Control | BABA | SeNPs | BABA-SeNPs | |
|---|---|---|---|---|
| FRAP | 11.9 ± 1.19 c | 18.3 ± 2.5 b | 18.8 ± 3.6 b | 24.0 ± 5.6 a |
| Phenolics | 3.54 ± 0.01 c | 5.7 ± 0.02 b | 6.7 ± 0.04 b | 8.9 ± 0.04 a |
| Flavonoids | 0.58 ± 0.01 c | 0.81 ± 0.01 b | 0.89 ± 0 b | 1.47 ± 0.02 a |
| Reduced GSH | 0.85 ± 0.11 b | 1.03 ± 0.3 b | 1.1 ± 0.24 a | 1.56 ± 0.19 a |
| Reduced ASC | 4.22 ± 0.47 b | 7.19 ± 0.69 a | 5.4 ± 0.56 b | 8.56 ± 0.38 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Selim, S.; Akhtar, N.; El Azab, E.; Warrad, M.; Alhassan, H.H.; Abdel-Mawgoud, M.; Al Jaouni, S.K.; Abdelgawad, H. Innovating the Synergistic Assets of β-Amino Butyric Acid (BABA) and Selenium Nanoparticles (SeNPs) in Improving the Growth, Nitrogen Metabolism, Biological Activities, and Nutritive Value of Medicago interexta Sprouts. Plants 2022, 11, 306. https://doi.org/10.3390/plants11030306
Selim S, Akhtar N, El Azab E, Warrad M, Alhassan HH, Abdel-Mawgoud M, Al Jaouni SK, Abdelgawad H. Innovating the Synergistic Assets of β-Amino Butyric Acid (BABA) and Selenium Nanoparticles (SeNPs) in Improving the Growth, Nitrogen Metabolism, Biological Activities, and Nutritive Value of Medicago interexta Sprouts. Plants. 2022; 11(3):306. https://doi.org/10.3390/plants11030306
Chicago/Turabian StyleSelim, Samy, Nosheen Akhtar, Eman El Azab, Mona Warrad, Hassan H. Alhassan, Mohamed Abdel-Mawgoud, Soad K. Al Jaouni, and Hamada Abdelgawad. 2022. "Innovating the Synergistic Assets of β-Amino Butyric Acid (BABA) and Selenium Nanoparticles (SeNPs) in Improving the Growth, Nitrogen Metabolism, Biological Activities, and Nutritive Value of Medicago interexta Sprouts" Plants 11, no. 3: 306. https://doi.org/10.3390/plants11030306
APA StyleSelim, S., Akhtar, N., El Azab, E., Warrad, M., Alhassan, H. H., Abdel-Mawgoud, M., Al Jaouni, S. K., & Abdelgawad, H. (2022). Innovating the Synergistic Assets of β-Amino Butyric Acid (BABA) and Selenium Nanoparticles (SeNPs) in Improving the Growth, Nitrogen Metabolism, Biological Activities, and Nutritive Value of Medicago interexta Sprouts. Plants, 11(3), 306. https://doi.org/10.3390/plants11030306