
Both Adaptability and Endophytic Bacteria Are Linked to the Functional Traits in the Invasive Clonal Plant Wedelia trilobata

 , and
, and 
Abstract
1. Introduction
2. Results
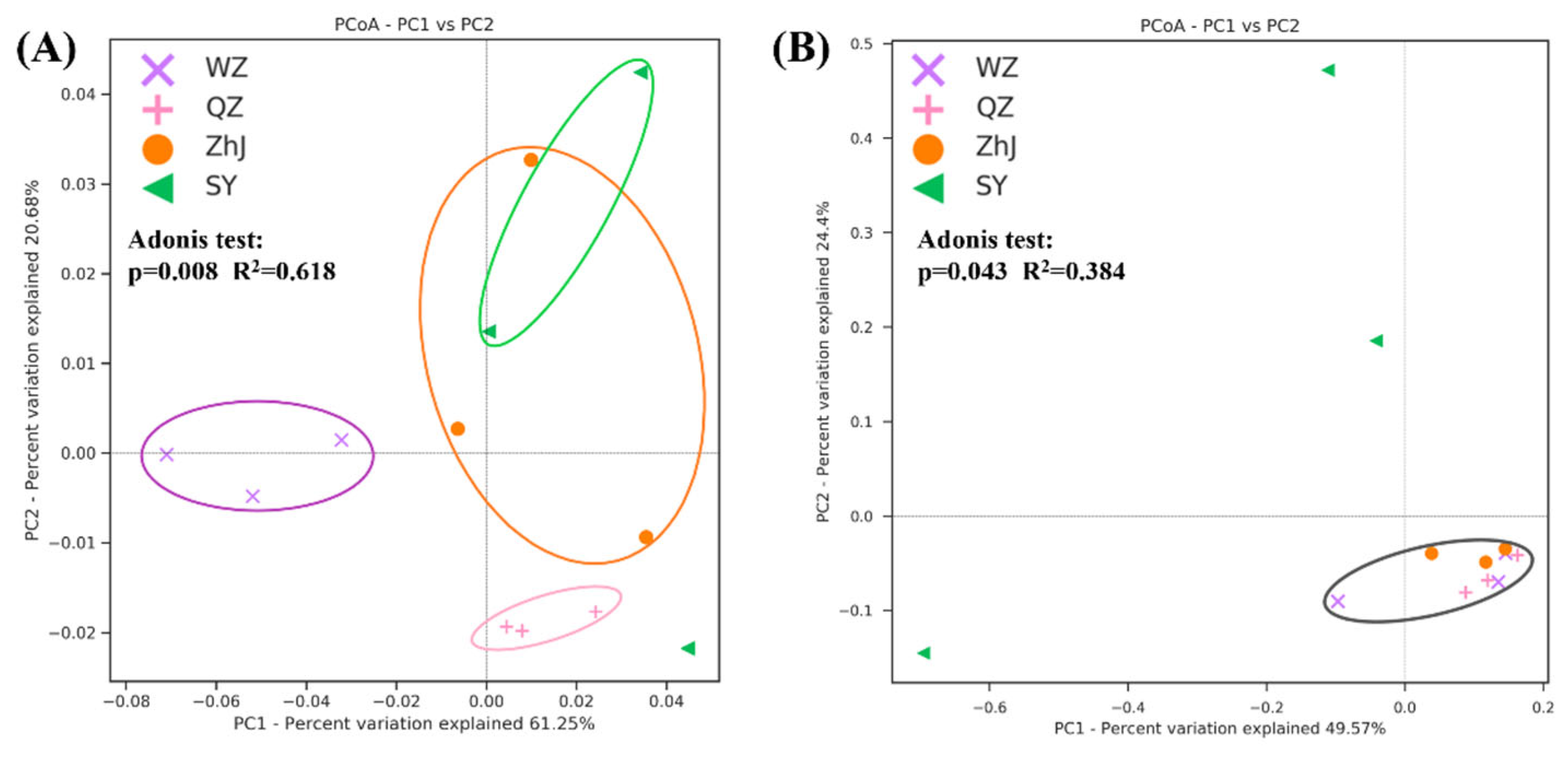
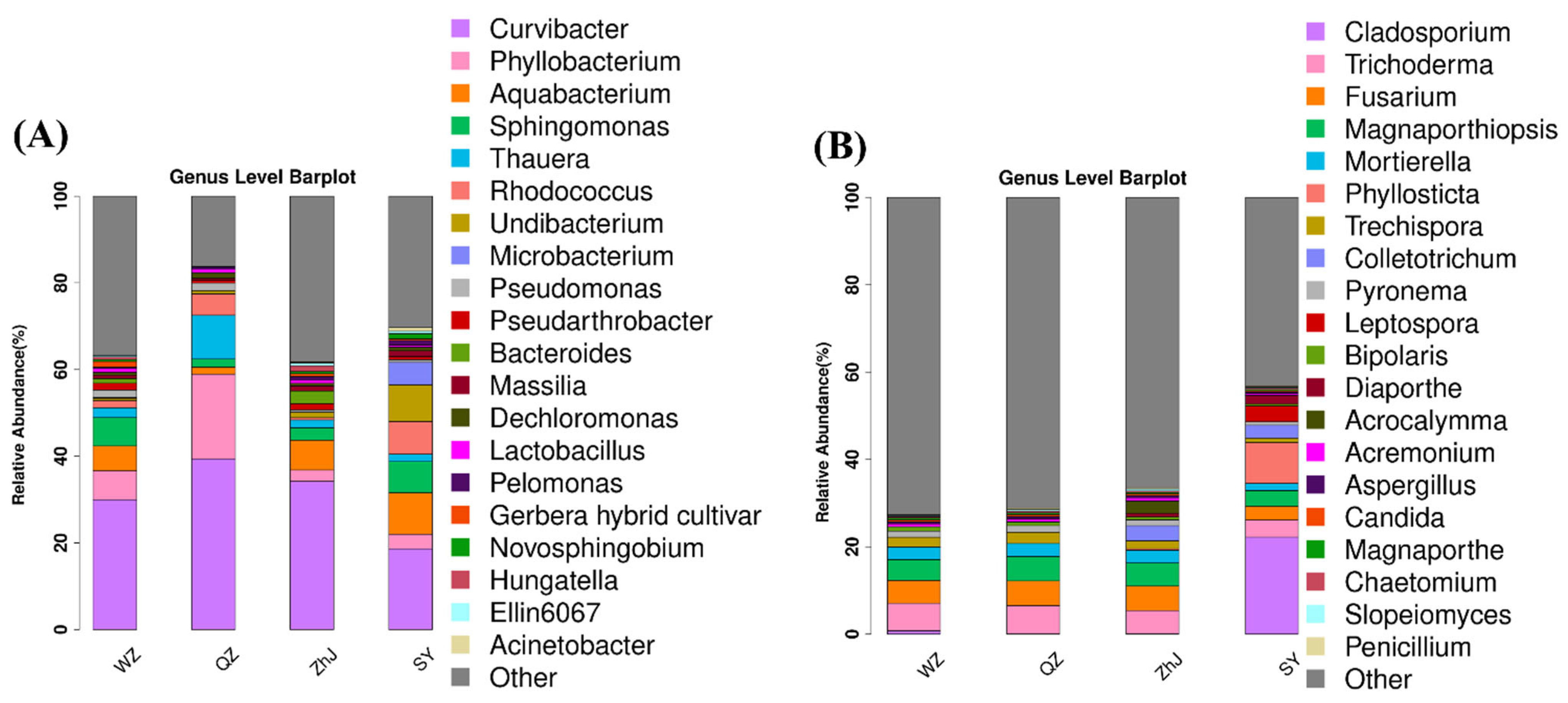
2.1. Endophytic Microbial Diversity and Composition
2.2. Plant Phenotypic Growth
2.3. Correlation Patterns between Microbial Diversity and Growth Indices of W. trilobata
3. Discussion
3.1. Geographical Changes of Endophytic Microbe and Clonal Growth Performances in Different Populations of W. trilobata
3.2. The Endophytic Microbial Composition May Contribute to the Clonal Phenotypic Growth of Invasive W. trilobata
4. Materials and Methods
4.1. Plant Materials and Pretreatment
4.2. Common Garden Experiment
4.3. Microbial Community Sequencing and Analysis
4.4. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hardoim, P.R.; van Overbeek, L.S.; van Elsas, J.D. Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol. 2008, 16, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.C.; Fu, W.; Wan, L.Y.; Cai, H.H.; Wang, N.; Qi, S.S.; Du, D.L. Different Growth Promoting Effects of En-dophytic Bacteria on Invasive and Native Clonal Plants. Front. Plant Sci. 2016, 7, 706. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Zhang, C.; Ye, Y.; Wen, J.; Wu, Y.; Wang, H.; Li, H.; Cai, S.; Cai, W.; Cheng, Z.; et al. Beneficial traits of bacterial endophytes belonging to the core communities of the tomato root microbiome. Agric. Ecosyst. Environ. 2017, 247, 149–156. [Google Scholar] [CrossRef]
- Verma, S.C.; Ladha, J.; Tripathi, A.K. Evaluation of plant growth promoting and colonization ability of endophytic diazotrophs from deep water rice. J. Biotechnol. 2001, 91, 127–141. [Google Scholar] [CrossRef]
- Egamberdieva, D. Alleviation of salt stress by plant growth regulators and IAA producing bacteria in wheat. Acta Physiol. Plant. 2009, 31, 861–864. [Google Scholar] [CrossRef]
- Ait Barka, E.; Nowak, J.; Clément, C. Enhancement of Chilling Resistance of Inoculated Grapevine Plantlets with a Plant Growth-Promoting Rhizobacterium, Burkholderia phytofirmans Strain PsJN. Appl. Environ. Microbiol. 2006, 72, 7246–7252. [Google Scholar] [CrossRef]
- Ding, S.; Huang, C.-L.; Sheng, H.-M.; Song, C.-L.; Li, Y.-B.; An, L.-Z. Effect of inoculation with the endophyte Clavibacter sp. strain Enf12 on chilling tolerance in Chorispora bungeana. Physiol. Plant. 2011, 141, 141–151. [Google Scholar] [CrossRef]
- Ryan, R.P.; Germaine, K.; Franks, A.; Ryan, D.J.; Dowling, D.N. Bacterial endophytes: Recent developments and applications. FEMS Microbiol. Lett. 2008, 278, 1–9. [Google Scholar] [CrossRef]
- Pyšek, P.; Richardson, D.M. Invasive Species, Environmental Change and Management, and Health. Annu. Rev. Environ. Resour. 2010, 35, 25–55. [Google Scholar] [CrossRef]
- Bellard, C.; Cassey, P.; Blackburn, T.M. Alien species as a driver of recent extinctions. Biol. Lett. 2016, 12, 20150623. [Google Scholar] [CrossRef]
- Paini, D.R.; Sheppard, A.W.; Cook, D.C.; De Barro, P.J.; Worner, S.P.; Thomas, M.B. Global threat to agriculture from invasive species. Proc. Natl. Acad. Sci. USA 2016, 113, 7575–7579. [Google Scholar] [CrossRef] [PubMed]
- Matthews, J.; van derVelde, G.; Collas, F.P.L.; de Hoop, L.; Koopman, K.R.; Hendriks, A.J.; Leuven, R.S.E.W. Inconsistencies in the risk classification of alien species and implications for risk assessment in the European Union. Ecosphere 2017, 8, e01832. [Google Scholar] [CrossRef]
- Aschehoug, E.T.; Metlen, K.L.; Callaway, R.M.; Newcombe, G. Fungal endophytes directly increase the competitive effects of an invasive forb. Ecology 2012, 93, 3–8. [Google Scholar] [CrossRef]
- Rout, M.E.; Chrzanowski, T.H.; Westlie, T.K.; DeLuca, T.H.; Callaway, R.M.; Holben, W.E. Bacterial endophytes enhance competition by invasive plants. Am. J. Bot. 2013, 100, 1726–1737. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhang, X.; Zheng, R.; Li, X.; Elmer, W.H.; Wolfe, L.M.; Li, B. Indirect effects of non-native Spartina alterniflora and its fungal pathogen (Fusarium palustre) on native saltmarsh plants in China. J. Ecol. 2014, 102, 1112–1119. [Google Scholar] [CrossRef]
- Afzal, I.; Shinwari, Z.K.; Sikandar, S.; Shahzad, S. Plant beneficial endophytic bacteria: Mechanisms, diversity, host range and genetic determinants. Microbiol. Res. 2019, 221, 36–49. [Google Scholar] [CrossRef]
- Liu, J.H.; Yong, X.H.; Han, Q.; Ali, A.; Wang, Y.J. Response of plant functional traits to species origin and adaptive reproduction in weeds. Plant Biosyst. 2017, 151, 323–330. [Google Scholar] [CrossRef]
- Naik, B.S. Functional roles of fungal endophytes in host fitness during stress conditions. Symbiosis 2019, 79, 99–115. [Google Scholar] [CrossRef]
- Mei, L.; Zhu, M.; Zhang, D.-Z.; Wang, Y.-Z.; Guo, J.; Zhang, H.-B. Geographical and Temporal Changes of Foliar Fungal Endophytes Associated with the Invasive Plant Ageratina adenophora. Microb. Ecol. 2014, 67, 402–409. [Google Scholar] [CrossRef]
- Koyama, A.; Maherali, H.; Antunes, P.M. Plant geographic origin and phylogeny as potential drivers of community structure in root-inhabiting fungi. J. Ecol. 2019, 107, 1720–1736. [Google Scholar] [CrossRef]
- Fang, K.; Miao, Y.-F.; Chen, L.; Zhou, J.; Yang, Z.-P.; Dong, X.-F.; Zhang, H.-B. Tissue-Specific and Geographical Variation in Endophytic Fungi of Ageratina adenophora and Fungal Associations with the Environment. Front. Microbiol. 2019, 10, 2919. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.D.; Tian, Z.S.; Feng, L.; Xu, L.; Wang, H.M. Diversity analysis of the rhizospheric and endophytic bacterial communities of Senecio vulgaris L. (Asteraceae) in an invasive range. PeerJ 2019, 6, e6162. [Google Scholar] [CrossRef] [PubMed]
- Lowe, S.J.; Browne, M.; Boudjelas, S. 100 of The World’s Worst Invasive Alien Species. A selection from the global invasive species database; Invasive Species Specialist Group: Auckland, New Zealand, 2000. [Google Scholar]
- Si, C.-C.; Dai, Z.-C.; Lin, Y.; Qi, S.-S.; Huang, P.; Miao, S.-L.; Du, D.-L. Local adaptation and phenotypic plasticity both occurred in Wedelia trilobata invasion across a tropical island. Biol. Invasions 2014, 16, 2323–2337. [Google Scholar] [CrossRef]
- Chen, S.; Jin, W.; Liu, A.; Zhang, S.; Liu, D.; Wang, F.; Lin, X.; He, C. Arbuscular mycorrhizal fungi (AMF) increase growth and secondary metabolism in cucumber subjected to low temperature stress. Sci. Hortic. 2013, 160, 222–229. [Google Scholar] [CrossRef]
- Dai, Z.-C.; Fu, W.; Qi, S.-S.; Zhai, D.-L.; Chen, S.-C.; Wan, L.-Y.; Huang, P.; Du, D.-L. Different Responses of an Invasive Clonal Plant Wedelia trilobata and its Native Congener to Gibberellin: Implications for Biological Invasion. J. Chem. Ecol. 2016, 42, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Javed, Q.; Azeem, A.; Ullah, I.; Saifullah, M.; Kama, R.; Du, D. Fluctuated water depth with high nutrient concentrations promote the invasiveness of Wedelia trilobata in Wetland. Ecol. Evol. 2020, 10, 832–842. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Hu, B.Q.; Hu, G. Assessment of allelopathic potential of Wedelia trilobata on the germination, seedling growth and chlorophyll content of rape. Adv. Mater. Res. 2013, 807, 719–722. [Google Scholar]
- Dai, Z.-C.; Wang, X.-Y.; Qi, S.-S.; Cai, H.-H.; Sun, J.-F.; Huang, P.; Du, D.-L. Effects of leaf litter on inter-specific competitive ability of the invasive plant Wedelia trilobata. Ecol. Res. 2016, 31, 367–374. [Google Scholar] [CrossRef]
- Azizan, K.A.; Ibrahim, S.; Ghani, N.H.A.; Nawawi, M.F. Metabolomics approach to investigate phytotoxic effects of Wedelia trilobata leaves, litter and soil. Plant Biosyst. 2019, 153, 691–699. [Google Scholar] [CrossRef]
- Javed, Q.; Sun, J.F.; Azeem, A.; Jabran, K.; Du, D.L. Competitive ability and plasticity of Wedelia trilobata (L.) under wetland hydrological variations. Sci. Rep. 2020, 10, 9431. [Google Scholar] [CrossRef]
- Song, L.; Chow, W.S.; Sun, L.; Li, C.; Peng, C. Acclimation of photosystem II to high temperature in two Wedelia species from different geographical origins: Implications for biological invasions upon global warming. J. Exp. Bot. 2010, 61, 4087–4096. [Google Scholar] [CrossRef] [PubMed]
- Song, L.-Y.; Li, C.-H.; Peng, S.-L. Elevated CO2 increases energy-use efficiency of invasive Wedelia trilobata over its indigenous congener. Biol. Invasions 2010, 12, 1221–1230. [Google Scholar] [CrossRef]
- Qi, S.S.; Dai, Z.C.; Miao, S.L.; Zhai, D.L.; Si, C.C.; Huang, P.; Wang, R.P.; Du, D.L. Light limitation and litter of an invasive clonal plant, Wedelia trilobata, inhibit its seedling recruitment. Ann. Bot. 2014, 114, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Hallmann, J.; Berg, G. Spectrum and Population Dynamics of Bacterial Root Endophytes. Microb. Root Endophytes 2006, 9, 15–31. [Google Scholar] [CrossRef]
- Pérez-Izquierdo, L.; Zabal-Aguirre, M.; González-Martínez, S.C.; Buée, M.; Verdú, M.; Rincón, A.; Goberna, M. Plant intraspecific variation modulates nutrient cycling through its below ground rhizospheric microbiome. J. Ecol. 2019, 107, 1594–1605. [Google Scholar] [CrossRef]
- Rodríguez-Caballero, G.; Caravaca, F.; Díaz, G.; Torres, P.; Roldán, A. The invader Carpobrotus edulis promotes a specific rhizosphere microbiome across globally distributed coastal ecosystems. Sci. Total Environ. 2020, 719, 137347. [Google Scholar] [CrossRef]
- Werner, J.J.; Zhou, D.; Caporaso, J.G.; Knight, R.; Angenent, L.T. Comparison of Illumina paired-end and single-direction sequencing for microbial 16S rRNA gene amplicon surveys. ISME J. 2012, 6, 1273–1276. [Google Scholar] [CrossRef]
- Coleman-Derr, D.; Desgarennes, D.; Fonseca-Garcia, C.; Gross, S.; Clingenpeel, S.; Woyke, T.; North, G.; Visel, A.; Partida-Martinez, L.P.; Tringe, S.G. Plant compartment and biogeography affect microbiome composition in cultivated and native Agave species. New Phytol. 2016, 209, 798–811. [Google Scholar] [CrossRef]
- Douhovnikoff, V.; Hazelton, E.L.G. Clonal growth: Invasion or stability? A comparative study of clonal architecture and diversity in native and introduced lineages of Phragmites australis (Poaceae). Am. J. Bot. 2014, 101, 1577–1584. [Google Scholar] [CrossRef]
- Alpert, P.; Simms, E.L. The relative advantages of plasticity and fixity in different environments: When is it good for a plant to adjust? Evol. Ecol. 2002, 16, 285–297. [Google Scholar] [CrossRef]
- Goulas, E.; Le Dily, F.; Teissedre, L.; Corbel, G.; Robin, C.; Ourry, A. Vegetative storage proteins in white clover (Trifolium repens L.): Quantitative and qualitative features. Ann. Bot. 2001, 88, 789–795. [Google Scholar]
- Stuefer, J.F. The role of stolon internodes for ramet survival after clone fragmentation in Potentilla anserina. Ecol. Lett. 2010, 2, 135–139. [Google Scholar] [CrossRef]
- Suzuki, J.-I.; Stuefer, J. On the ecological and evolutionary significance of storage in clonal plants. Plant Species Biol. 1999, 14, 11–17. [Google Scholar] [CrossRef]
- Bittebiere, A.K.; Benot, M.L.; Mony, C. Clonality as a key but overlooked driver of biotic interactions in plants. Perspect. Plant Ecol. Evol. Syst. 2020, 43, 125510. [Google Scholar] [CrossRef]
- Dai, Z.-C.; Wan, L.-Y.; Qi, S.-S.; Rutherford, S.; Ren, G.-Q.; Wan, J.S.; Du, D.-L. Synergy among hypotheses in the invasion process of alien plants: A road map within a timeline. Perspect. Plant Ecol. Evol. Syst. 2020, 47, 125575. [Google Scholar] [CrossRef]
- Roiloa, S.R. Clonal traits and plant invasiveness: The case of Carpobrotus NE.Br. (Aizoaceae). Perspect. Plant Ecol. Evol. Syst. 2019, 40, 125479. [Google Scholar] [CrossRef]
- Dahal, K.; Kane, K.; Gadapati, W.; Webb, E.; Savitch, L.V.; Singh, J.; Sharma, P.; Sarhan, F.; Longstaffe, F.J.; Grodzinski, B. The effects of phenotypic plasticity on photosynthetic performance in winter rye, winter wheat and Brassica napus. Physiol. Plant 2012, 144, 169–188. [Google Scholar] [CrossRef]
- Ott, J.P.; Klimešová, J.; Hartnett, D.C. The ecology and significance of below-ground bud banks in plants. Ann. Bot. 2019, 123, 1099–1118. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.D.; Knapp, A.K. Physiological and morphological traits of exotic, invasive exotic, and native plant species in tallgrass prairie. Int. J. Plant Sci. 2001, 162, 785–792. [Google Scholar] [CrossRef]
- Van Kleunen, M.; Weber, E.; Fischer, M. A meta-analysis of trait differences between invasive and non-invasive plant species. Ecol. Lett. 2009, 13, 235–245. [Google Scholar] [CrossRef]
- Ma, Y.; Rajkumar, M.; Zhang, C.; Freitas, H. Beneficial role of bacterial endophytes in heavy metal phytoremedia-tion. J. Environ. Manag. 2016, 174, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Miliute, I.; Buzaite, O.; Baniulis, D.; Stanys, V. Bacterial endophytes in agricultural crops and their role in stress tolerance: A review. Zemdirb. Agric. 2015, 102, 465–478. [Google Scholar] [CrossRef]
- Gravel, V.; Antoun, H.; Tweddell, R.J. Growth stimulation and fruit yield improvement of greenhouse tomato plants by inoculation with Pseudomonas putida or Trichoderma atroviride: Possible role of indole acetic acid (IAA). Soil Biol. Biochem. 2008, 39, 1968–1977. [Google Scholar] [CrossRef]
- Phetcharat, P.; Duangpaeng, A. Screening of Endophytic Bacteria from Organic Rice Tissue for Indole Acetic Acid Production. Procedia Eng. 2012, 32, 177–183. [Google Scholar] [CrossRef]
- Ali, A.; Mohanta, T.K.; Asaf, S.; Rehman, N.; Al-Housni, S.; Al-Harrasi, A.; Khan, A.L.; Al-Rawahi, A. Biotransformation of benzoin by Sphingomonas sp. LK11 and ameliorative effects on growth of Cucumis sativus. Arch. Microbiol. 2019, 201, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Ham, S.; Yoon, H.; Park, J.-M.; Park, Y.G. Optimization of Fermentation Medium for Indole Acetic Acid Production by Pseudarthrobacter sp. NIBRBAC000502770. Appl. Biochem. Biotechnol. 2021, 193, 2567–2579. [Google Scholar] [CrossRef]
- Vives-Peris, V.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Salt stress alleviation in citrus plants by plant growth-promoting rhizobacteria Pseudomonas putida and Novosphingobium sp. Plant Cell Rep. 2018, 37, 1557–1569. [Google Scholar] [CrossRef]
- Pang, F.; Xia, W.K.; He, M.; Qi, S.S.; Dai, Z.C.; Du, D.L. Nitrogen fixing bacteria alleviate the nutritional competition of arbuscular mycorrhizal fungi against Solidago canadensis in a nitrogen limited environment. Chin. J. Plant Ecol. 2020, 44, 782–790. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Growth Performance | Geographical Latitude | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pre-Cultivate | Post-Cultivate | |||||||||||
| Shoot Length | Stem Diameter | Spacer Length | Stem Number | Shoot Length | Stem Diameter | Spacer Length | Stem Number | Leaf Area | ||||
| Bacterial | Chao1 index | r | −0.151 | 0.1418 | −0.195 | −0.0411 | 0.3882 | 0.1187 | 0.3078 | −0.012 | 0.372 | −0.512 |
| p | 0.6384 | 0.6602 | 0.5432 | 0.8990 | 0.2124 | 0.7133 | 0.3304 | 0.97 | 0.234 | 0.089 | ||
| Observed species | r | −0.048 | 0.2336 | −0.064 | −0.0417 | 0.3847 | 0.2136 | 0.4645 | −0.202 | 0.417 | −0.478 | |
| p | 0.8823 | 0.4649 | 0.8438 | 0.8975 | 0.2169 | 0.5051 | 0.1282 | 0.53 | 0.178 | 0.116 | ||
| Shannon diversity | r | 0.0521 | 0.4535 | 0.2475 | −0.1538 | 0.336 | 0.3786 | 0.4867 | −0.282 | 0.446 | −0.237 | |
| p | 0.8723 | 0.1387 | 0.4381 | 0.6331 | 0.2856 | 0.2249 | 0.1086 | 0.374 | 0.147 | 0.458 | ||
| Simpson diversity | r | −0.185 | 0.776 | 0.702 | −0.542 | 0.3396 | 0.713 | 0.596 | −0.407 | 0.636 | 0.406 | |
| p | 0.565 | 0.003 | 0.011 | 0.0687 | 0.2801 | 0.0093 | 0.041 | 0.19 | 0.026 | 0.191 | ||
| Fungal | Chao1 index | r | −0.304 | −0.0951 | −0.139 | −0.0851 | −0.053 | −0.0198 | 0.0321 | −0.081 | 0.0689 | 0.248 |
| p | 0.3371 | 0.7688 | 0.6664 | 0.7926 | 0.8704 | 0.9512 | 0.9211 | 0.803 | 0.8314 | 0.436 | ||
| Observed species | r | −0.495 | −0.0517 | −0.101 | −0.2352 | −0.186 | 0.0044 | 0.0069 | −0.143 | 0.0714 | 0.418 | |
| p | 0.1018 | 0.8732 | 0.7551 | 0.4618 | 0.5625 | 0.9891 | 0.983 | 0.657 | 0.8255 | 0.176 | ||
| Shannon diversity | r | −0.452 | 0.0807 | 0.0735 | −0.3209 | −0.026 | 0.1604 | 0.1581 | −0.187 | 0.1718 | 0.472 | |
| p | 0.1399 | 0.8031 | 0.8204 | 0.3091 | 0.9355 | 0.6186 | 0.6236 | 0.561 | 0.5934 | 0.121 | ||
| Simpson diversity | r | −0.369 | 0.0875 | 0.1297 | −0.3018 | 0.0319 | 0.173 | 0.2058 | −0.21 | 0.1606 | 0.420 | |
| p | 0.2385 | 0.7868 | 0.6879 | 0.3404 | 0.9215 | 0.5908 | 0.5211 | 0.511 | 0.6181 | 0.174 | ||
| Geographical Latitude | r | r | 0.536 | 0.64 | −0.737 | −0.203 | 0.523 | 0.328 | −0.51 | 0.285 | - | |
| p | p | 0.73 | 0.025 | 0.006 | 0.528 | 0.081 | 0.299 | 0.091 | 0.368 | - | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mei, Y.-H.; Li, X.; Zhou, J.-Y.; Kong, F.-L.; Qi, S.-S.; Zhu, B.; Naz, M.; Dai, Z.-C.; Du, D.-L. Both Adaptability and Endophytic Bacteria Are Linked to the Functional Traits in the Invasive Clonal Plant Wedelia trilobata. Plants 2022, 11, 3369. https://doi.org/10.3390/plants11233369
Mei Y-H, Li X, Zhou J-Y, Kong F-L, Qi S-S, Zhu B, Naz M, Dai Z-C, Du D-L. Both Adaptability and Endophytic Bacteria Are Linked to the Functional Traits in the Invasive Clonal Plant Wedelia trilobata. Plants. 2022; 11(23):3369. https://doi.org/10.3390/plants11233369
Chicago/Turabian StyleMei, Ying-Hao, Xu Li, Jian-Yu Zhou, Fang-Li Kong, Shan-Shan Qi, Bin Zhu, Misbah Naz, Zhi-Cong Dai, and Dao-Lin Du. 2022. "Both Adaptability and Endophytic Bacteria Are Linked to the Functional Traits in the Invasive Clonal Plant Wedelia trilobata" Plants 11, no. 23: 3369. https://doi.org/10.3390/plants11233369
APA StyleMei, Y.-H., Li, X., Zhou, J.-Y., Kong, F.-L., Qi, S.-S., Zhu, B., Naz, M., Dai, Z.-C., & Du, D.-L. (2022). Both Adaptability and Endophytic Bacteria Are Linked to the Functional Traits in the Invasive Clonal Plant Wedelia trilobata. Plants, 11(23), 3369. https://doi.org/10.3390/plants11233369