
Light Quality Impacts Vertical Growth Rate, Phytochemical Yield and Cannabinoid Production Efficiency in Cannabis sativa
 ,
,
Abstract
1. Introduction
2. Results
2.1. Morphology
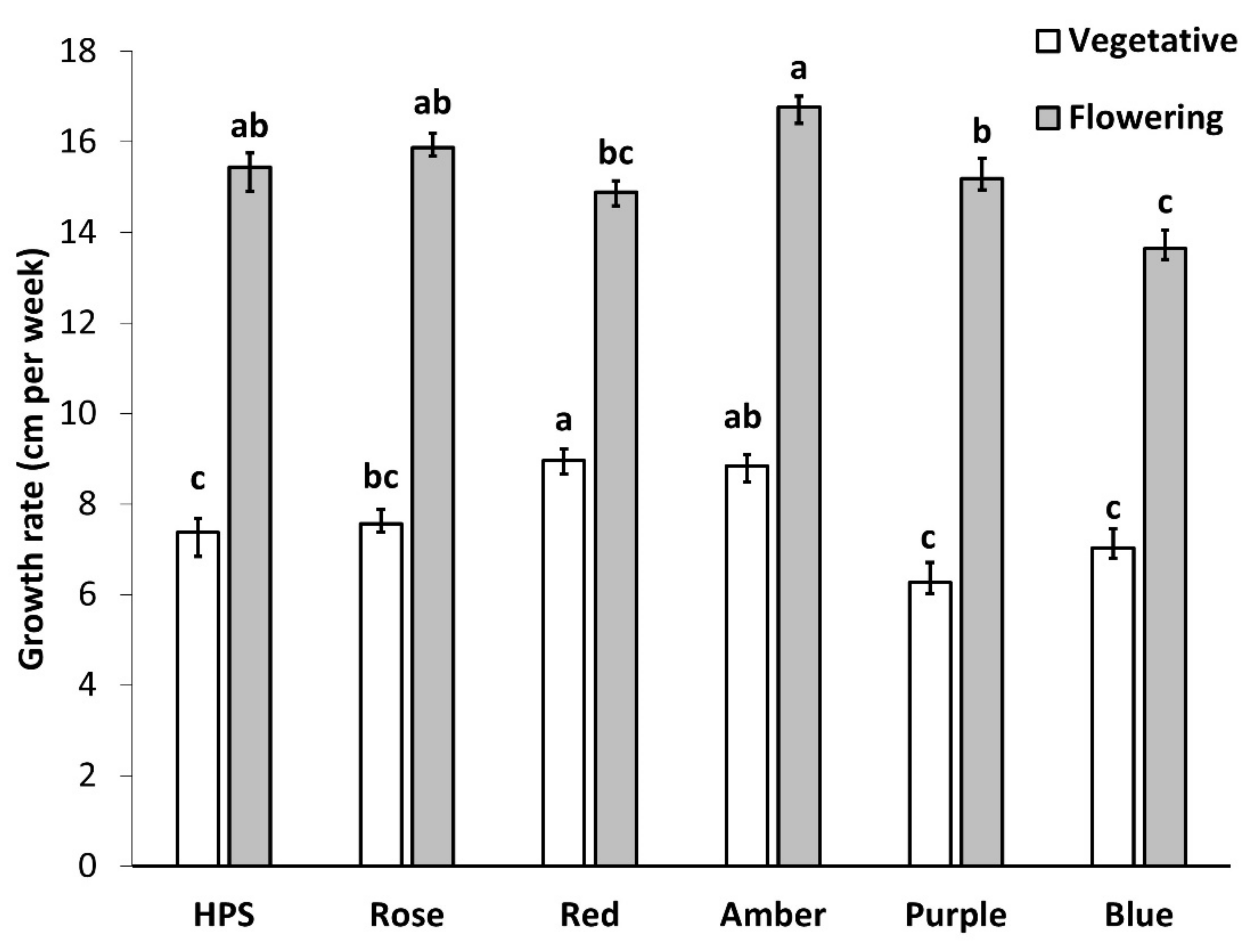
2.1.1. Plant Height and Vertical Growth Rate
2.1.2. Tissue Coloration
2.2. Inflorescence Yield
2.3. Phytochemical Concentration
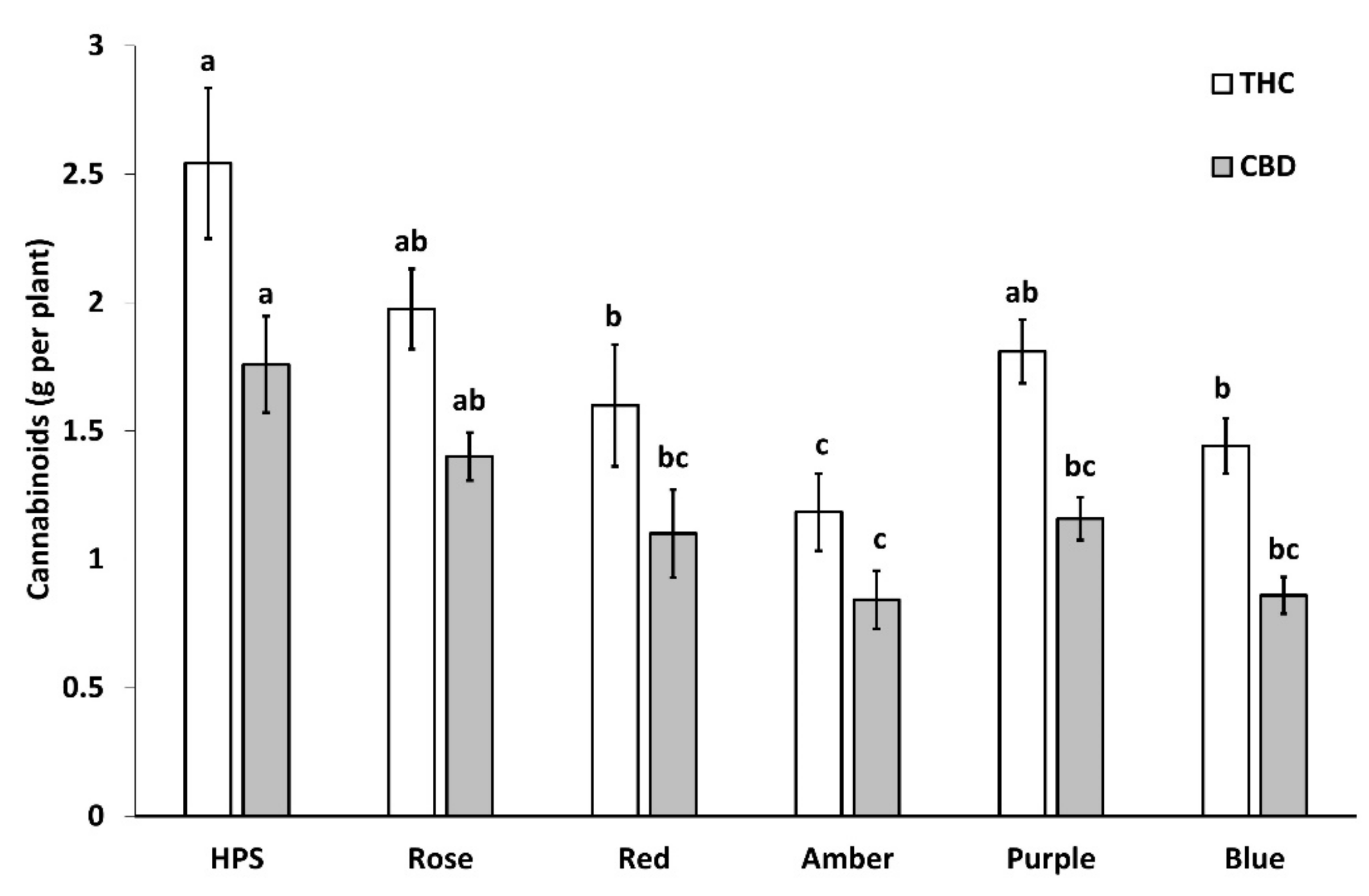
2.3.1. Total Cannabinoid Concentration
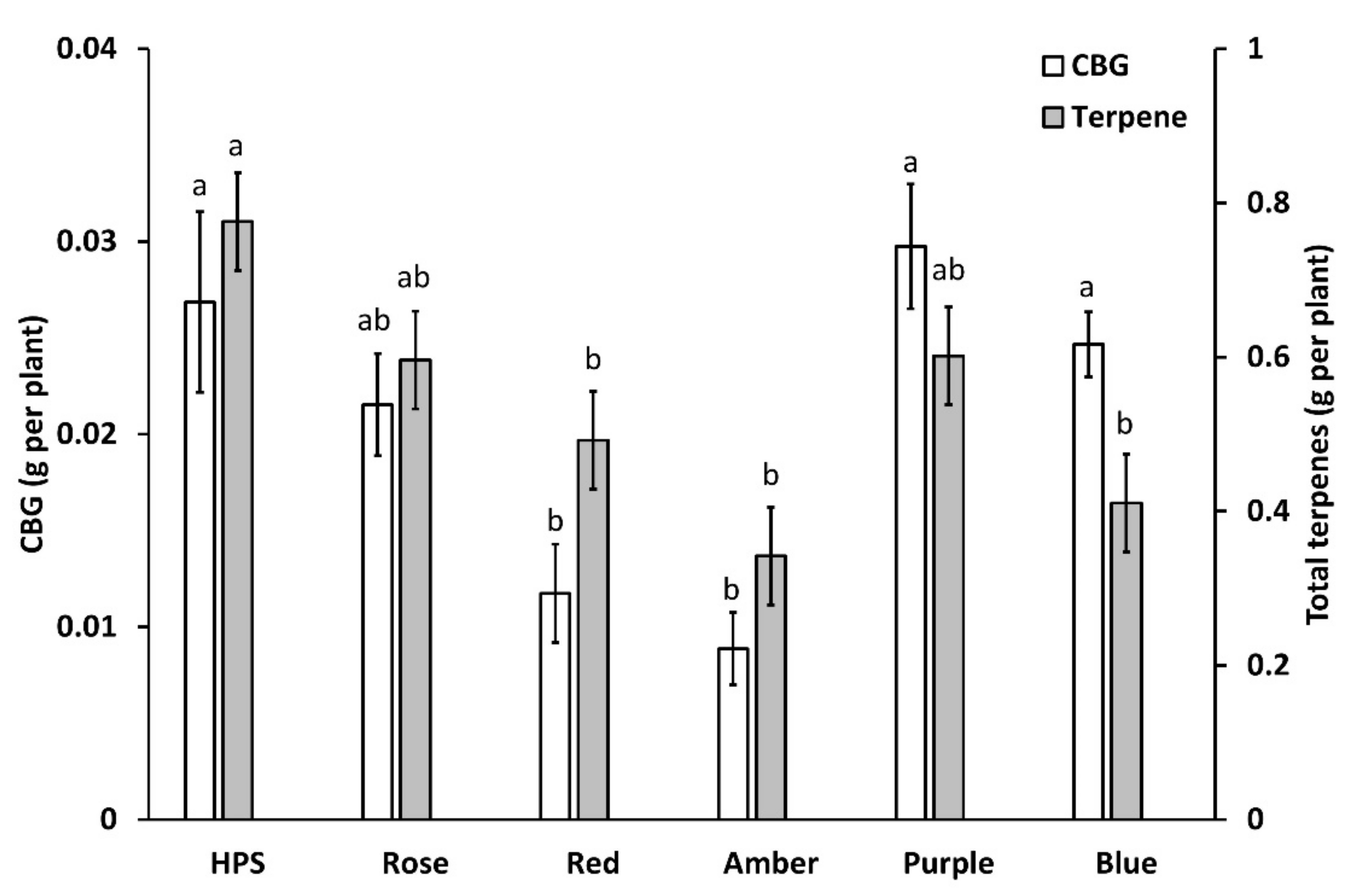
2.3.2. Total Terpene Concentration
2.3.3. Light Effect on Terpene Profile
2.4. Total Phytochemical Yield
2.4.1. Light Treatment Impacts Total THC and CBD Yield per Plant
2.4.2. Light Treatment Impacts Total THC and CBD Yield per Plant
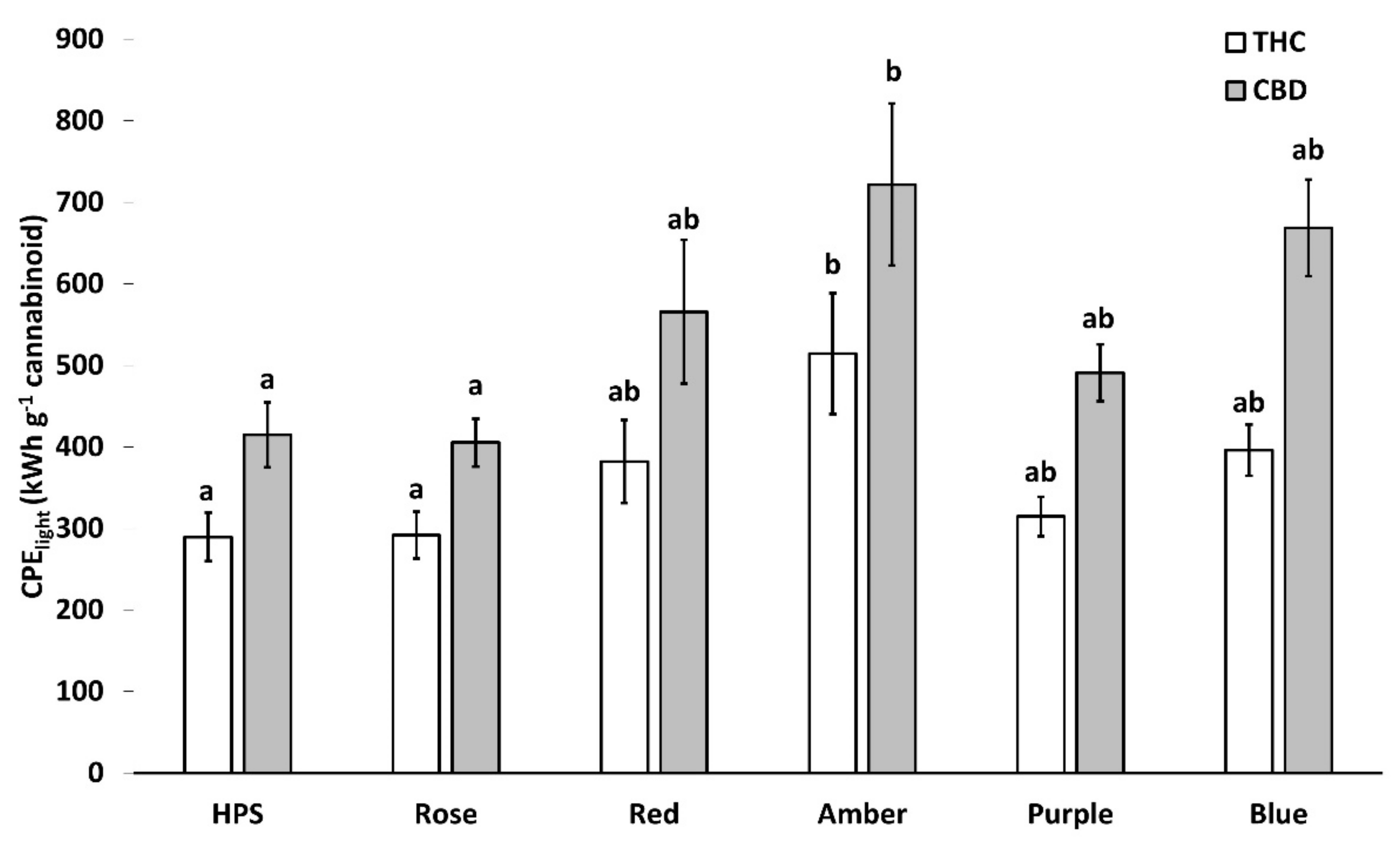
2.5. Light-Associated Cannabinoid Production Efficiency
3. Discussion
3.1. Morphology
3.2. Inflorescence Mass
3.3. Cannabinoids Concentration, Yield, and Profitability
3.4. Terpenes
3.5. Limitations (Differences between Replicates)
3.6. Implication for Growers
4. Materials and Methods
4.1. Cultivation Environment
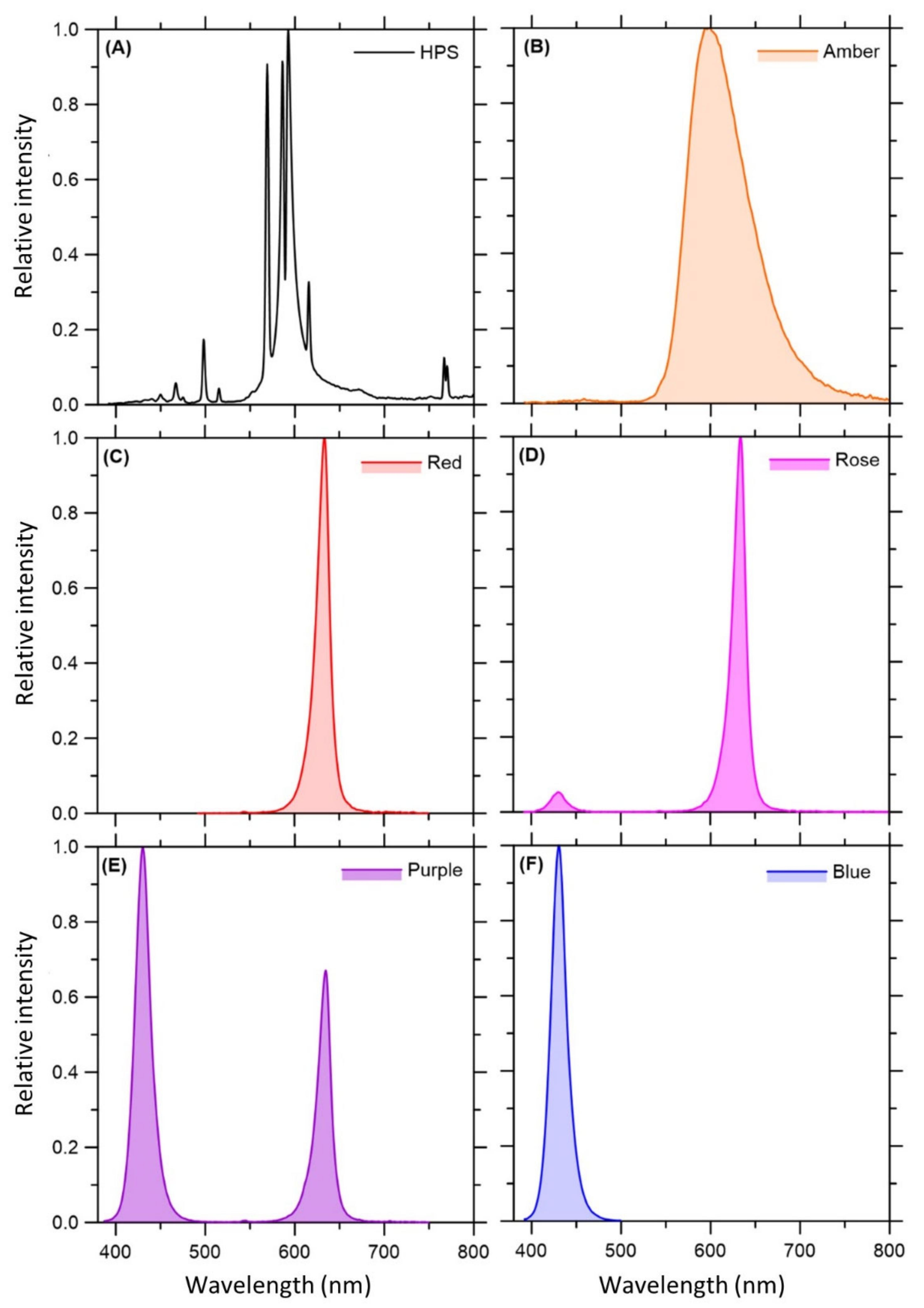
4.2. Light Treatments
4.3. Plant Materials and Cultivation
4.4. Vertical Growth Rate
4.5. Plant Images
4.6. Plant Harvest and Phytochemical Measurement
4.7. Light Associated Cannabinoid Production Efficacy
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Petrovska, B.B. Historical Review of Medicinal Plants’ Usage. Pharmacogn. Rev. 2012, 6, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Bilodeau, S.E.; Wu, B.-S.; Rufyikiri, A.-S.; MacPherson, S.; Lefsrud, M. An Update on Plant Photobiology and Implications for Cannabis Production. Front. Plant Sci. 2019, 10, 296. [Google Scholar] [CrossRef] [PubMed]
- Bahji, A.; Stephenson, C. International Perspectives on the Implications of Cannabis Legalization: A Systematic Review & Thematic Analysis. Int. J. Environ. Res. Public Health 2019, 16, 3095. [Google Scholar] [CrossRef]
- ElSohly, M.A.; Radwan, M.M.; Gul, W.; Chandra, S.; Galal, A. Phytochemistry of Cannabis sativa L. Prog. Chem. Org. Nat. Prod. 2017, 103, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.; Dai, K.; Xie, Z.; Chen, J. Secondary Metabolites Profiled in Cannabis Inflorescences, Leaves, Stem Barks, and Roots for Medicinal Purposes. Sci. Rep. 2020, 10, 3309–3314. [Google Scholar] [CrossRef] [PubMed]
- Livingston, S.J.; Quilichini, T.; Booth, J.K.; Wong, D.C.J.; Rensing, K.H.; Laflamme-Yonkman, J.; Castellarin, S.D.; Bohlmann, J.; Page, J.E.; Samuels, A.L. Cannabis glandular trichomes alter morphology and metabolite content during flower maturation. Plant J. 2019, 101, 37–56. [Google Scholar] [CrossRef]
- Gonçalves, J.; Rosado, T.; Soares, S.; Simão, A.Y.; Caramelo, D.; Luís, Â.; Fernández, N.; Barroso, M.; Gallardo, E.; Duarte, A.P. Cannabis and Its Secondary Metabolites: Their Use as Therapeutic Drugs, Toxicological Aspects, and Analytical Determination. Medicines 2019, 6, 31. [Google Scholar] [CrossRef]
- Mao, J.; Price, D.; Lu, J.; Keniston, L.; Mayer, D. Two distinctive antinociceptive systems in rats with pathological pain. Neurosci. Lett. 2000, 280, 13–16. [Google Scholar] [CrossRef]
- Van Klingeren, B.; Ham, M.T. Antibacterial activity of Δ9-tetrahydrocannabinol and cannabidiol. Antonie Van Leeuwenhoek 1976, 42, 9–12. [Google Scholar] [CrossRef]
- Russo, E.B.; Marcu, J. Cannabis Pharmacology: The Usual Suspects and a Few Promising Leads. In Advances in Pharmacology; Academic Press: Cambridge, MA, USA, 2017; Volume 80, pp. 67–134. [Google Scholar] [CrossRef]
- Gallily, R.; Yekhtin, Z.; Hanuš, L.O. Overcoming the Bell-Shaped Dose-Response of Cannabidiol by Using Cannabis Extract Enriched in Cannabidiol. Pharmacol. Pharm. 2015, 06, 75–85. [Google Scholar] [CrossRef]
- Johnson, J.R.; Burnell-Nugent, M.; Lossignol, D.; Ganae-Motan, E.D.; Potts, R.; Fallon, M.T. Multicenter, Double-Blind, Randomized, Placebo-Controlled, Parallel-Group Study of the Efficacy, Safety, and Tolerability of THC:CBD Extract and THC Extract in Patients with Intractable Cancer-Related Pain. J. Pain Symptom Manag. 2010, 39, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Russo, E.B. Taming THC: Potential cannabis synergy and phytocannabinoid-terpenoid entourage effects. Br. J. Pharmacol. 2011, 163, 1344–1364. [Google Scholar] [CrossRef] [PubMed]
- Mechoulam, R.; Ben-Shabat, S. From gan-zi-gun-nu to anandamide and 2-arachidonoylglycerol: The ongoing story of cannabis. Nat. Prod. Rep. 1999, 16, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Russo, E.B. The Case for the Entourage Effect and Conventional Breeding of Clinical Cannabis: No “Strain,” No Gain. Front. Plant Sci. 2019, 9, 1969. [Google Scholar] [CrossRef] [PubMed]
- Brousseau, V.D.; Wu, B.-S.; MacPherson, S.; Morello, V.; Lefsrud, M. Cannabinoids and Terpenes: How Production of Photo-Protectants Can Be Manipulated to Enhance Cannabis sativa L. Phytochemistry. Front. Plant Sci. 2021, 12, 620021. [Google Scholar] [CrossRef]
- Hawley, D.; Graham, T.; Stasiak, M.; Dixon, M. Improving Cannabis Bud Quality and Yield with Subcanopy Lighting. HortScience 2018, 53, 1593–1599. [Google Scholar] [CrossRef]
- Magagnini, G.; Grassi, G.; Kotiranta, S. The Effect of Light Spectrum on the Morphology and Cannabinoid Content of Cannabis sativa L. Med. Cannabis Cannabinoids 2018, 1, 19–27. [Google Scholar] [CrossRef]
- Reichel, P.; Munz, S.; Hartung, J.; Präger, A.; Kotiranta, S.; Burgel, L.; Schober, T.; Graeff-Hönninger, S. Impact of Three Different Light Spectra on the Yield, Morphology and Growth Trajectory of Three Different Cannabis sativa L. Strains. Plants 2021, 10, 1866. [Google Scholar] [CrossRef]
- Singh, D.; Basu, C.; Meinhardt-Wollweber, M.; Roth, B. LEDs for energy efficient greenhouse lighting. Renew. Sustain. Energy Rev. 2015, 49, 139–147. [Google Scholar] [CrossRef]
- Wu, B.-S.; Hitti, Y.; MacPherson, S.; Orsat, V.; Lefsrud, M.G. Comparison and perspective of conventional and LED lighting for photobiology and industry applications. Environ. Exp. Bot. 2019, 171, 103953. [Google Scholar] [CrossRef]
- Mills, E. The carbon footprint of indoor Cannabis production. Energy Policy 2012, 46, 58–67. [Google Scholar] [CrossRef]
- Potter, D.J.; Duncombe, P. The Effect of Electrical Lighting Power and Irradiance on Indoor-Grown Cannabis Potency and Yield. J. Forensic Sci. 2011, 57, 618–622. [Google Scholar] [CrossRef] [PubMed]
- Vanhove, W.; Van Damme, P.; Meert, N. Factors determining yield and quality of illicit indoor cannabis (Cannabis spp.) production. Forensic Sci. Int. 2011, 212, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Amrein, P.; Rinner, S.; Pittorino, T.; Espel, J.; Schmidmayr, D. Influence of Light Spectra on the Production of Cannabinoids. Med. Cannabis Cannabinoids 2020, 3, 103–110. [Google Scholar] [CrossRef]
- Danziger, N.; Bernstein, N. Light matters: Effect of light spectra on cannabinoid profile and plant development of medical cannabis (Cannabis sativa L.). Ind. Crops Prod. 2021, 164, 113351. [Google Scholar] [CrossRef]
- Namdar, D.; Charuvi, D.; Ajjampura, V.; Mazuz, M.; Ion, A.; Kamara, I.; Koltai, H. LED lighting affects the composition and biological activity of Cannabis sativa secondary metabolites. Ind. Crops Prod. 2019, 132, 177–185. [Google Scholar] [CrossRef]
- Hogewoning, S.; Trouwborst, G.; Maljaars, H.; Poorter, H.; Van Ieperen, W.; Harbinson, J. Blue light dose-responses of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef]
- Liu, X.; Guo, S.; Chang, T.; Xu, Z.; Tezuka, T. Regulation of the growth and photosynthesis of cherry tomato seedlings by different light irradiations of light emitting diodes (LED). Afr. J. Biotechnol. 2012, 11, 6169–6177. [Google Scholar] [CrossRef]
- Sullivan, J.; Deng, X.W. From seed to seed: The role of photoreceptors in Arabidopsis development. Dev. Biol. 2003, 260, 289–297. [Google Scholar] [CrossRef]
- Meng, Q.; Boldt, J.; Runkle, E.S. Blue Radiation Interacts with Green Radiation to Influence Growth and Predominantly Controls Quality Attributes of Lettuce. J. Am. Soc. Hortic. Sci. 2020, 145, 75–87. [Google Scholar] [CrossRef]
- Naznin, M.T.; Lefsrud, M.; Gravel, V.; Azad, O.K. Blue Light added with Red LEDs Enhance Growth Characteristics, Pigments Content, and Antioxidant Capacity in Lettuce, Spinach, Kale, Basil, and Sweet Pepper in a Controlled Environment. Plants 2019, 8, 93. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Zhao, X.; Long, S.; Xiao, Q.; Guo, Y.; Qiu, C.; Qiu, H.; Wang, Y. Wavelengths of LED light affect the growth and cannabidiol content in Cannabis sativa L. Ind. Crops Prod. 2021, 165, 113433. [Google Scholar] [CrossRef]
- Westmoreland, F.M.; Kusuma, P.; Bugbee, B. Cannabis lighting: Decreasing blue photon fraction increases yield but efficacy is more important for cost effective production of cannabinoids. PLoS ONE 2021, 16, e0248988. [Google Scholar] [CrossRef]
- Giupponi, L.; Leoni, V.; Pavlovic, R.; Giorgi, A. Influence of Altitude on Phytochemical Composition of Hemp Inflorescence: A Metabolomic Approach. Molecules 2020, 25, 1381. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.J.; Ryu, B.R.; Azad, M.O.K.; Rahman, M.H.; Cheong, E.J.; Lim, J.-D.; Lim, Y.-S. Cannabinoids Accumulation in Hemp (Cannabis sativa L.) Plants under LED Light Spectra and Their Discrete Role as a Stress Marker. Biology 2021, 10, 710. [Google Scholar] [CrossRef] [PubMed]
- Tohidi, B.; Rahimmalek, M.; Arzani, A.; Sabzalian, M.R. Thymol, carvacrol, and antioxidant accumulation in Thymus species in response to different light spectra emitted by light-emitting diodes. Food Chem. 2019, 307, 125521. [Google Scholar] [CrossRef]
- Ghaffari, Z.; Rahimmalek, M.; Sabzalian, M.R. Variation in the primary and secondary metabolites derived from the isoprenoid pathway in the Perovskia species in response to different wavelengths generated by light emitting diodes (LEDs). Ind. Crops Prod. 2019, 140, 111592. [Google Scholar] [CrossRef]
- Ueda, T.; Murata, M.; Yokawa, K. Single Wavelengths of LED Light Supplement Promote the Biosynthesis of Major Cyclic Monoterpenes in Japanese Mint. Plants 2021, 10, 1420. [Google Scholar] [CrossRef]
- Appolloni, E.; Pennisi, G.; Zauli, I.; Carotti, L.; Paucek, I.; Quaini, S.; Orsini, F.; Gianquinto, G. Beyond vegetables: Effects of indoor LED light on specialized metabolite biosynthesis in medicinal and aromatic plants, edible flowers, and microgreens. J. Sci. Food Agric. 2021, 102, 472–487. [Google Scholar] [CrossRef]
- Wu, B.-S.; MacPherson, S.; Lefsrud, M. Filtering Light-Emitting Diodes to Investigate Amber and Red Spectral Effects on Lettuce Growth. Plants 2021, 10, 1075. [Google Scholar] [CrossRef]
- Martineau, V.; Lefsrud, M.; Naznin, M.T.; Kopsell, D. Comparison of Light-emitting Diode and High-pressure Sodium Light Treatments for Hydroponics Growth of Boston Lettuce. HortScience 2012, 47, 477–482. [Google Scholar] [CrossRef]
- Gajc-Wolska, J.; Kowalczyk, K.; Metera, A.; Mazur, K.; Bujalski, D.; Hemka, L. Effect of supplementary lighting on selected physiological parameters and yielding of tomato plants. Folia Hortic. 2013, 25, 153–159. [Google Scholar] [CrossRef]
- Son, K.-H.; Oh, M.-M. Leaf Shape, Growth, and Antioxidant Phenolic Compounds of Two Lettuce Cultivars Grown under Various Combinations of Blue and Red Light-emitting Diodes. HortScience 2013, 48, 988–995. [Google Scholar] [CrossRef]
- Wollaeger, H.M.; Runkle, E.S. Growth and Acclimation of Impatiens, Salvia, Petunia, and Tomato Seedlings to Blue and Red Light. HortScience 2015, 50, 522–529. [Google Scholar] [CrossRef]
- Kong, Y.; Stasiak, M.; Dixon, M.A.; Zheng, Y. Blue light associated with low phytochrome activity can promote elongation growth as shade-avoidance response: A comparison with red light in four bedding plant species. Environ. Exp. Bot. 2018, 155, 345–359. [Google Scholar] [CrossRef]
- Franklin, K.A.; Whitelam, G.C. Phytochromes and Shade-avoidance Responses in Plants. Ann. Bot. 2005, 96, 169–175. [Google Scholar] [CrossRef]
- Britz, S.J.; Sager, J.C. Photomorphogenesis and photoassimilation in soybean and sorghum grown under broad spectrum or blue-deficient light sources. Plant Physiol. 1990, 94, 448–454. [Google Scholar] [CrossRef]
- Grimstad, S. The effect of supplemental irradiation with different light sources on growth and flowering of gloxinia (Sinningia speciosa (Lodd.) Hiern). Sci. Hortic. 1987, 32, 297–305. [Google Scholar] [CrossRef]
- Bergstrand, K.-J.; Mortensen, L.M.; Suthaparan, A.; Gislerød, H.R. Acclimatisation of greenhouse crops to differing light quality. Sci. Hortic. 2016, 204, 1–7. [Google Scholar] [CrossRef]
- Yavari, N.; Lefsrud, M.G. Proteomic Analysis Provides Insights into Arabidopsis Thaliana Response Under Narrow-Wavelength LED of 595 Nm Light. J. Proteom. Bioinform. 2019, 12, 134–143. [Google Scholar] [CrossRef]
- Yigal, E.; Shtienberg, D. Botrytis Cinerea in Greenhouse Vegetables: Chemical, Cultural, Physiological and Biological Controls and Their Integration. Integr. Pest Manag. Rev. 1995, 29, 15–29. [Google Scholar] [CrossRef]
- Punja, Z.K. Epidemiology of Fusarium oxysporum causing root and crown rot of cannabis (Cannabis sativa L., marijuana) plants in commercial greenhouse production. Can. J. Plant Pathol. 2020, 43, 216–235. [Google Scholar] [CrossRef]
- Benke, K.; Tomkins, B. Future food-production systems: Vertical farming and controlled-environment agriculture. Sustain. Sci. Pract. Policy 2017, 13, 13–26. [Google Scholar] [CrossRef]
- Broséus, J.; Anglada, F.; Esseiva, P. The differentiation of fibre- and drug type Cannabis seedlings by gas chromatography/mass spectrometry and chemometric tools. Forensic Sci. Int. 2010, 200, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Smalls, E.; Beckstead, H.D. Common Cannabinoid Phenotypes in 350 Stocks of Cannabis. Lloydia 1973, 36, 144–165. [Google Scholar]
- Deiana, S. Potential Medical Uses of Cannabigerol: A Brief Overview; Elsevier Inc.: Amsterdam, The Netherlands, 2017; ISBN 9780128008270. [Google Scholar]
- Pacifico, D.; Miselli, F.; Carboni, A.; Moschella, A.; Mandolino, G. Time course of cannabinoid accumulation and chemotype development during the growth of Cannabis sativa L. Euphytica 2007, 160, 231–240. [Google Scholar] [CrossRef]
- Booth, J.K.; Page, J.E.; Bohlmann, J. Terpene synthases from Cannabis sativa. PLoS ONE 2017, 12, e0173911. [Google Scholar] [CrossRef]
- Rodriguez-Morrison, V.; Llewellyn, D.; Zheng, Y. Cannabis Yield, Potency, and Leaf Photosynthesis Respond Differently to Increasing Light Levels in an Indoor Environment. Front. Plant Sci. 2021, 12, 646020. [Google Scholar] [CrossRef]
- Balasus, J.; Blank, J.; Babilon, S.; Hegemann, T.; Khanh, T.Q. Energy Efficient Lighting in Plant Factories: Addressing Utilance. Agronomy 2021, 11, 2570. [Google Scholar] [CrossRef]
- De Backer, B.; Debrus, B.; Lebrun, P.; Theunis, L.; Dubois, N.; Decock, L.; Verstraete, A.; Hubert, P.; Charlier, C. Innovative development and validation of an HPLC/DAD method for the qualitative and quantitative determination of major cannabinoids in cannabis plant material. J. Chromatogr. B 2009, 877, 4115–4124. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publication: Carol Stream, IL, USA, 2006. [Google Scholar]
- Mondello, L. Mass Spectra of Flavors and Fragrances of Natural and Synthetic Compounds, 3rd ed.; Wiley: Hoboken, NJ, USA, 2011; ISBN 978-1-119-06984-3. [Google Scholar]
- Tkachyov, A.V. Analysis of Volatile Plant Compounds; Ofset: Novosibirsk, Russia, 2008. [Google Scholar]
- Hochmuth, D.H. MassFinder 4; 4.25.; Dr. Hochmuth Scientific Consulting: Hamburg, Germany, 2004. [Google Scholar]
- National Institute of Standards and Technology. NIST Mass Spectral Database for NIST/EPA/NIH and Mass Spectral Search Program (Version 2.3). Available online: https://chemdata.nist.gov/dokuwiki/doku.php?id=chemdata:nist17 (accessed on 10 January 2017).
- Cachet, T.; Brevard, H.; Chaintreau, A.; Demyttenaere, J.; French, L.; Gassenmeier, K.; Joulain, D.; Koenig, T.; Leijs, H.; Liddle, P.; et al. IOFI recommended practice for the use of predicted relative-response factors for the rapid quantification of volatile flavouring compounds by GC-FID. Flavour Fragr. J. 2016, 31, 191–194. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Average Plant Height at Harvest (cm) | ||
|---|---|---|
| Light Treatment | Replicate 1 | Replicate 2 |
| HPS | 78.8 ± 1.6 | 67.4 ± 2.1 |
| Rose | 78.1 ± 1.2 | 71.2 ± 0.9 |
| Red | 76.5 ± 1.2 | 73.2 ± 1.0 |
| Amber | 84.4 ± 1.2 | 74.8 ± 1.6 |
| Purple | 72.0 ± 2.1 | 69.4 ± 1.2 |
| Blue | 68.0 ± 2.1 | 66.6 ± 1.0 |
| Light Treatment | Dry Inflorescence (g per Plant) | Fresh Inflorescence (g per Plant) | ||
|---|---|---|---|---|
| Replicate 1 | Replicate 2 | Replicate 1 | Replicate 2 | |
| HPS | 28.3 ± 3.5 | 24.2 ± 7.8 | 139.3 ± 16.9 | 127.9 ± 38.7 |
| Rose | 27.7 ± 4.1 | 22.9 ± 3.5 | 134.8 ± 16.7 | 123.1 ± 13.6 |
| Red | 24.7 ± 3.7 | 23.0 ± 3.8 | 126.3 ± 18.1 | 119.7 ± 17.7 |
| Amber | 21.4 ± 3.3 | 15.3 ± 2.8 | 112.8 ± 15.9 | 93.5 ± 20.6 |
| Purple | 20.2 ± 4.1 | 17.3 ± 2.1 | 101.9 ± 17.4 | 92.3 ± 9.8 |
| Blue | 17.4 ± 3.3 | 13.5 ± 2.3 | 82.3 ± 14.9 | 70.5 ± 9.6 |
| Light Treatment | Total THC (%) | Total CBD (%) | Total CBG (%) | Total Terpenes (mg g−1) | ||
|---|---|---|---|---|---|---|
| Replicate 1 | Replicate 2 | Replicate 1 | Replicate 2 | Both | Both | |
| HPS | 7.7 ± 0.7 | 9.4 ± 0.8 | 5.5 ± 0.5 | 6.3 ± 0.6 | 0.09 ± 0.03 b | 25.8 ± 2.4 ab |
| Rose | 7.0 ± 1.0 | 9.4 ± 0.2 | 5.2 ± 0.4 | 6.4 ± 0.4 | 0.09 ± 0.03 b | 24.5 ± 3.4 ab |
| Red | 5.0 ± 0.5 | 8.3 ± 1.0 | 3.6 ± 0.7 | 5.5 ± 0.9 | 0.05 ± 0.02 b | 20.3 ± 5.3 bc |
| Amber | 5.1 ± 1.8 | 7.7 ± 0.6 | 4.0 ± 1.4 | 5.0 ± 0.5 | 0.05 ± 0.02 b | 18.1 ± 3.2 c |
| Purple | 8.0 ± 0.8 | 10.0 ± 0.2 | 5.4 ± 0.5 | 6.0 ± 0.2 | 0.15 ± 0.03 a | 29.4 ± 2.8 a |
| Blue | 10.0 ± 0.1 | 10.4 ± 0.3 | 6.0 ± 0.1 | 6.1 ± 0.1 | 0.18 ± 0.03 a | 28.5 ± 2.8 a |
| Terpene Compound (mg g−1) | Light Treatment | |||||
|---|---|---|---|---|---|---|
| HPS | Amber | Red | Rose | Purple | Blue | |
| Myrcene | 7.50 ± 0.43 ab | 4.80 ± 0.44 c | 5.66 ± 0.75 bc | 7.34 ± 0.57 ab | 8.91 ± 0.53 a | 7.89 ± 0.65 a |
| α-Pinene | 5.33 ± 0.45 | 4.48 ± 0.25 | 4.73 ± 0.12 | 5.13 ± 0.31 | 5.71 ± 0.36 | 5.73 ± 0.38 |
| Limonene | 2.66 ± 0.12 ab | 1.85 ± 0.16 c | 2.17 ± 0.24 bc | 2.59 ± 0.17 abc | 3.05 ± 0.11 a | 2.65 ± 0.23 ab |
| β-Pinene | 2.47 ± 0.16 ab | 1.92 ± 0.10 b | 2.03 ± 0.11 b | 2.30 ± 0.08 ab | 2.70 ± 0.13 a | 2.68 ± 0.16 a |
| Linalool | 0.717 ± 0.06 ab | 0.428 ± 0.06 b | 0.467 ± 0.09 ab | 0.59 ± 0.06 ab | 0.84 ± 0.04 a | 0.84 ± 0.02 a |
| (E)-β-Ocimene | 0.59 ± 0.05 abc | 0.347 ± 0.039 c | 0.437 ± 0.06 bc | 0.625 ± 0.06 ab | 0.75 ± 0.06 a | 0.631 ± 0.05 ab |
| Light Fraction (nm) | Light Treatments (μmol m−2 s−1) | |||||
|---|---|---|---|---|---|---|
| HPS | Amber (595 nm) | Red (630 nm) | Rose (430 nm + 630 nm; 1:10) | Purple (430 nm + 630 nm; 2:1) | Blue (430 nm) | |
| 300–399 | 0.2 | 0.5 | 0.0 | 4.1 | 29.4 | 44.6 |
| 400–499 | 17.8 | 0.4 | 0.2 | 32.1 | 236.8 | 352.6 |
| 500–599 | 164.6 | 258.5 | 40.9 | 37.4 | 15.2 | 2.6 |
| 600–699 | 137.5 | 131.9 | 355.3 | 323 | 117.3 | 0.0 |
| 700–799 | 15.1 | 6.8 | 3.1 | 2.8 | 1.0 | 0.0 |
| >800 | 65.3 | 0.8 | - | - | - | - |
| sum | 398.8 | 398.9 | 399.6 | 399.4 | 399.7 | 399.8 |
| Electrical parameters | ||||||
| Voltage (VAC) | 120 | 120 | 120 | 120 | 120 | 120 |
| Current (A) | 6.15 | 4.85 | 4.92 | 4.88 | 4.98 | 4.90 |
| Power (W) | 738 | 582 | 590.4 | 585.6 | 597.6 | 588 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morello, V.; Brousseau, V.D.; Wu, N.; Wu, B.-S.; MacPherson, S.; Lefsrud, M. Light Quality Impacts Vertical Growth Rate, Phytochemical Yield and Cannabinoid Production Efficiency in Cannabis sativa. Plants 2022, 11, 2982. https://doi.org/10.3390/plants11212982
Morello V, Brousseau VD, Wu N, Wu B-S, MacPherson S, Lefsrud M. Light Quality Impacts Vertical Growth Rate, Phytochemical Yield and Cannabinoid Production Efficiency in Cannabis sativa. Plants. 2022; 11(21):2982. https://doi.org/10.3390/plants11212982
Chicago/Turabian StyleMorello, Victorio, Vincent Desaulniers Brousseau, Natalie Wu, Bo-Sen Wu, Sarah MacPherson, and Mark Lefsrud. 2022. "Light Quality Impacts Vertical Growth Rate, Phytochemical Yield and Cannabinoid Production Efficiency in Cannabis sativa" Plants 11, no. 21: 2982. https://doi.org/10.3390/plants11212982
APA StyleMorello, V., Brousseau, V. D., Wu, N., Wu, B.-S., MacPherson, S., & Lefsrud, M. (2022). Light Quality Impacts Vertical Growth Rate, Phytochemical Yield and Cannabinoid Production Efficiency in Cannabis sativa. Plants, 11(21), 2982. https://doi.org/10.3390/plants11212982