
Production of Nematicidal Pinosylvin Stilbenes in Cell Suspension Cultures of Pinus koraiensis by Fungal Elicitation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
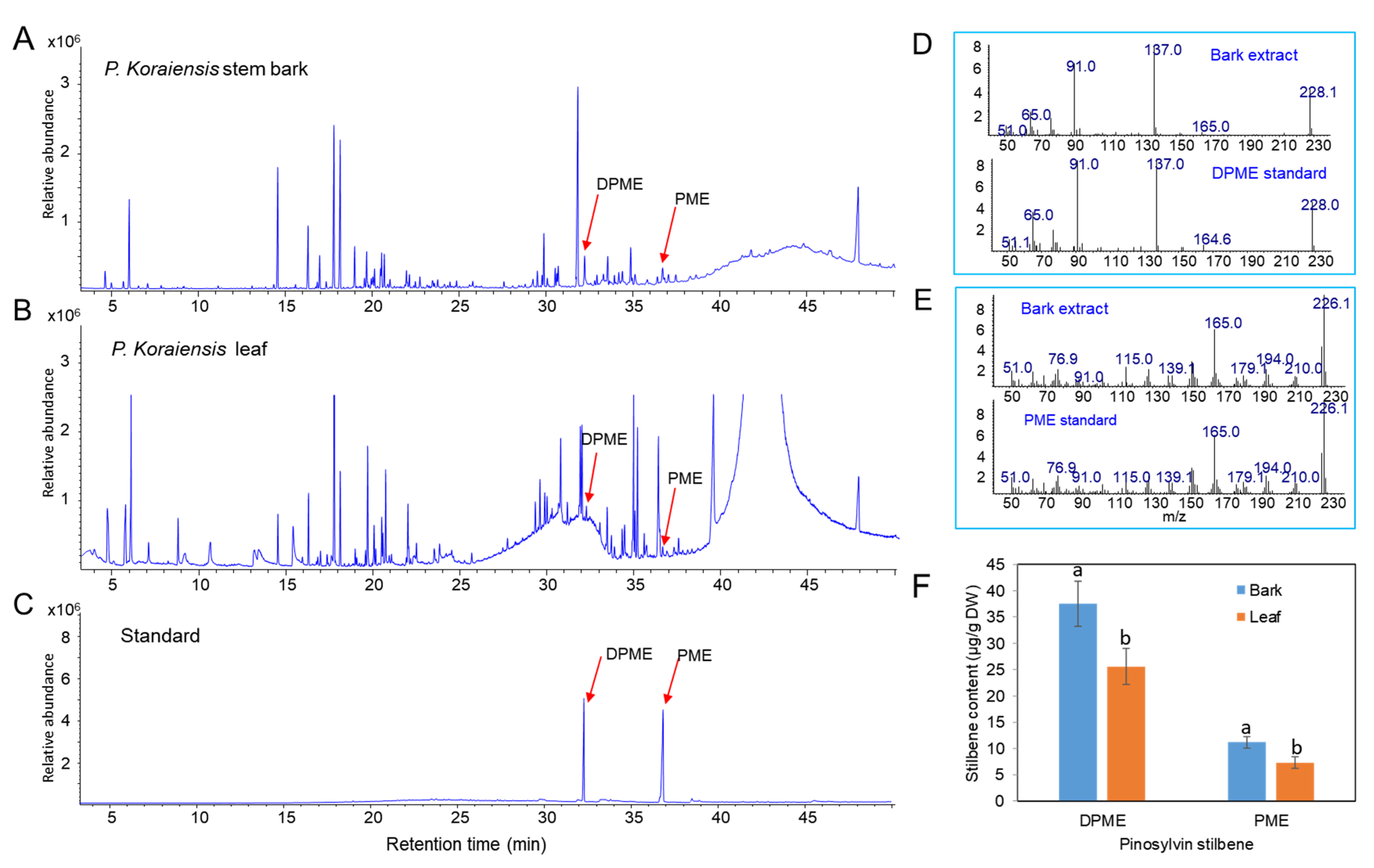
2.1. Content of Pinosylvin Stilbenes (DPME and PME) in Stem Bark in P. koraiensis
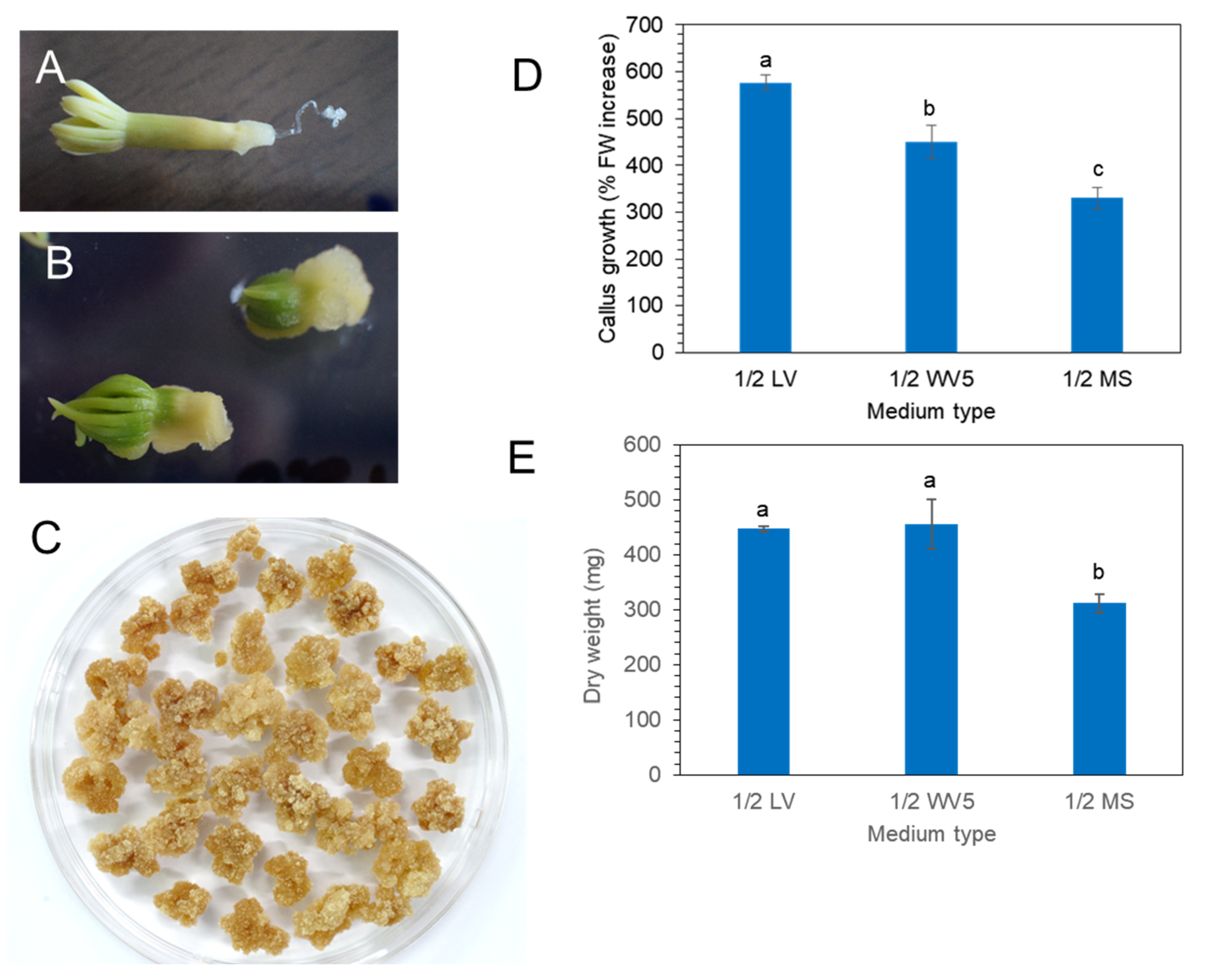
2.2. Induction and Proliferation of P. koraiensis Calli
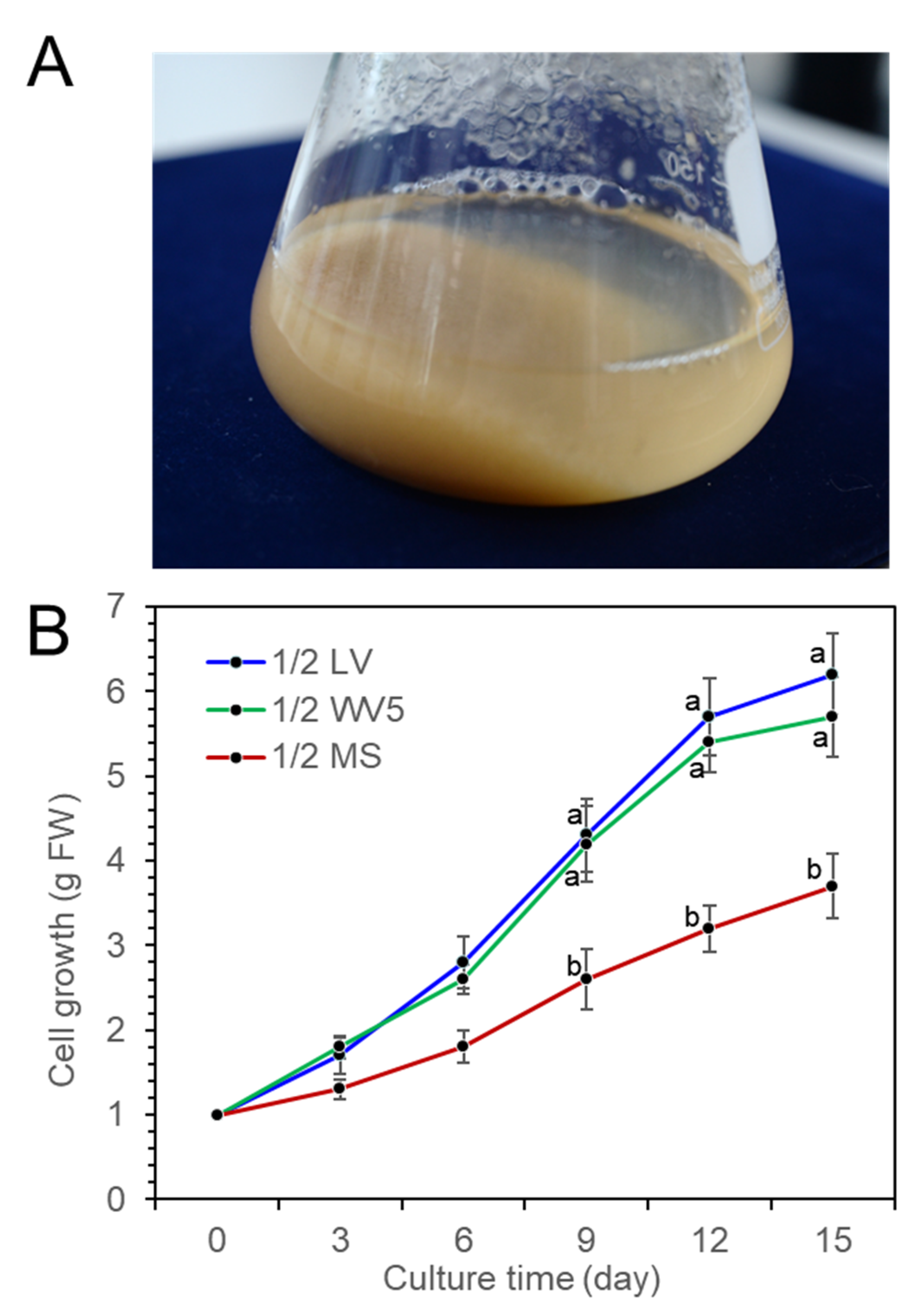
2.3. Cell Suspension Culture
2.4. Enhanced Production of DPME and PME in Cell Suspension by Fungal Elicitor Treatment
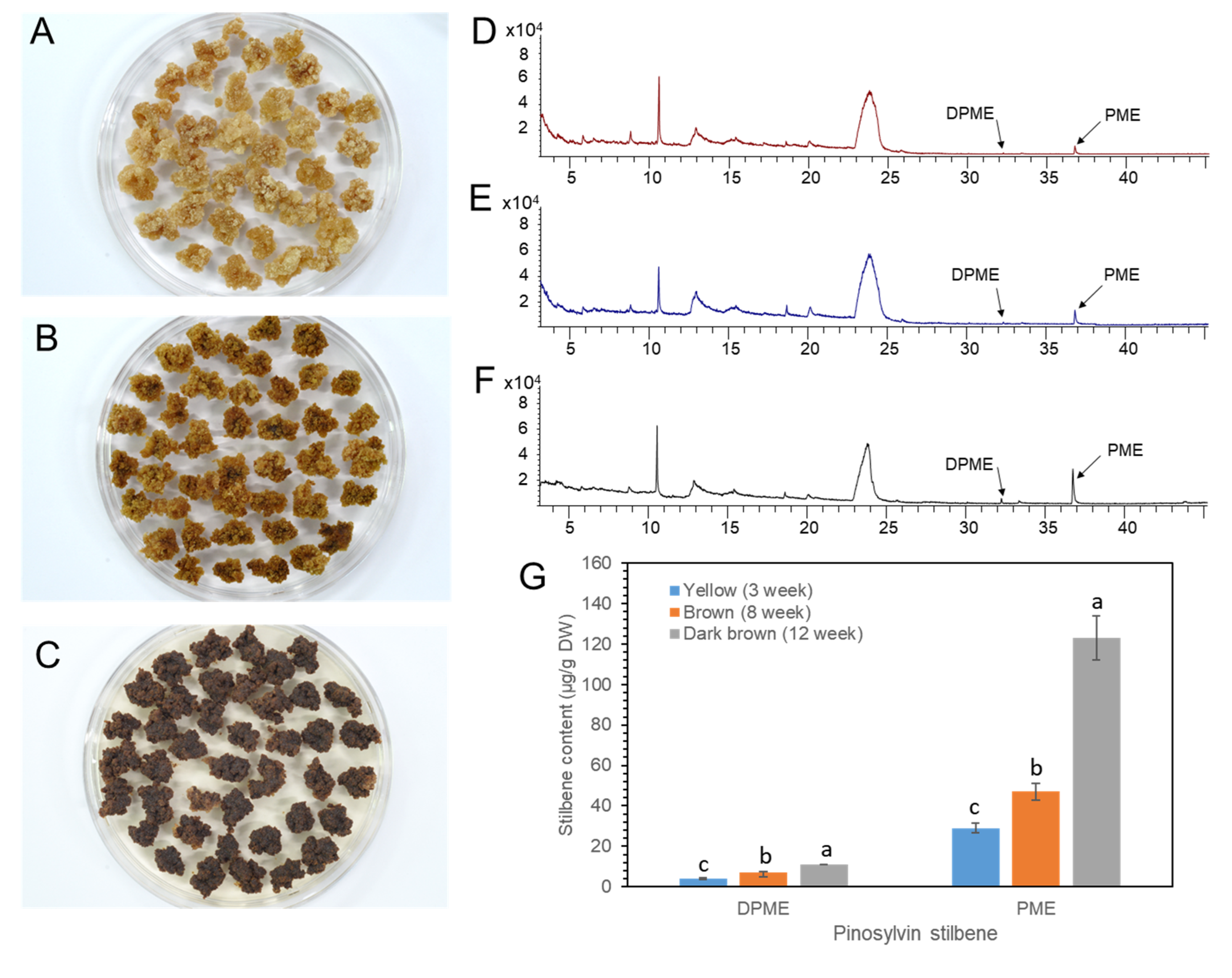
2.5. DPME and PME Accumulation by Prolonged Culture of Calli
2.6. qPCR Analysis of PsSTS and PsPMT Genes
2.7. Nematicidal Activity of Crude Extracts from P. koraiensis Calli
3. Discussion
4. Materials and Methods
4.1. Callus Induction from Mature Zygotic Embryos of P. koraiensis
4.2. Callus Proliferation
4.3. Cell Suspension Culture
4.4. Preparation of Fungal Elicitors
4.5. Fungal Elicitor Treatment during Cell Suspension Culture of P. koraiensis
4.6. qRT-PCR Analysis in Fungal Elicitor-Treated Calli
4.7. GC–MS Analysis
4.8. Nematicidal Activity of Callus Extracts
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mamiya, Y. Pathology of the pine wilt disease caused by Bursaphelenchus xylophilus. Ann. Rev. Phytopathol. 1983, 21, 201–220. [Google Scholar] [CrossRef] [PubMed]
- Hirata, A.; Nakamura, K.; Nakao, K.; Kominami, Y.; Tanaka, N.; Ohashi, H.; Takano, K.T.; Takeuchi, W.; Matsui, T. Potential distribution of pine wilt disease under future climate change scenarios. PLoS ONE 2017, 12, e0182837. [Google Scholar] [CrossRef]
- Futai, K. Pine Wood Nematode, Bursaphelenchus Xylophilus. Annu. Rev. Phytopathol. 2013, 51, 61–83. [Google Scholar] [CrossRef] [PubMed]
- Linit, M.J. Nematode-vector relationships in the pine wilt disease system. J. Nematol. 1988, 20, 227–235. [Google Scholar]
- Kurinobu, S. Current status of resistance breeding of Japanese pine species to pine wilt disease. For. Sci. Technol. 2008, 4, 51–57. [Google Scholar] [CrossRef]
- James, R.; Tisserat, N.; Todd, T. Prevention of pine wilt of scots pine (Pinus sylvestris) with systemic abamectin injections. Arboric. Urban For. 2006, 32, 195–201. [Google Scholar] [CrossRef]
- Sousa, E.; Naves, P.; Vieira, M. Prevention of pine wilt disease induced by Bursaphelenchus xylophilus and Monochamus galloprovincialis by trunk injection of emamectin benzoate. Phytoparasitica 2013, 41, 143–148. [Google Scholar] [CrossRef]
- Andrés, M.F.; González-Coloma, A.; Sanz, J.; Burillo, J.; Sainz, P. Nematicidal activity of essential oils: A review. Phytochem. Rev. 2012, 11, 371–390. [Google Scholar] [CrossRef]
- Faria, J.M.S.; Barbosa, P.; Vieira, P.; Vicente, C.S.L.; Figueiredo, A.C.; Mota, M. Phytochemicals as biopesticides against the pinewood nematode Bursaphelenchus xylophilus: A review on essential oils and their volatiles. Plants 2021, 10, 2614. [Google Scholar] [CrossRef]
- Rivière, C.; Pawlus, A.D.; Mérillon, J.-M. Natural stilbenoids: Distribution in the plant kingdom and chemotaxonomic interest in Vitaceae. Nat. Prod. Rep. 2012, 29, 1317–1333. [Google Scholar] [CrossRef]
- Hart, J.H. Role of phytostilbenes in decay and disease resistance. Annu. Rev. Phytopathol. 1981, 19, 437–458. [Google Scholar] [CrossRef]
- Harju, A.M.; Venäläinen, M. Measuring the decay resistance of Scots pine heartwood indirectly by the Folin-Ciocalteu assay. Can. J. For. Res. 2006, 36, 1797–1804. [Google Scholar] [CrossRef]
- Chong, J.; Poutaraud, A.; Hugueney, P. Metabolism and roles of stilbenes in plants. Plant Sci. 2009, 117, 143–155. [Google Scholar] [CrossRef]
- Valletta, A.; Iozia, L.M.; Leonelli, F. Impact of environmental factors on stilbene biosynthesis. Plants 2021, 10, 90. [Google Scholar] [CrossRef]
- Suga, T.; Ohta, S.; Munesada, K.; Ide, N.; Kurokawa, M.; Shimizu, M.; Ohta, E. Endogenous pine wood nematicidal substances in Pines, Pinus Massoniana, P. Strobus and P. Palustris. Phytochtmistry 1993, 33, 1395–1401. [Google Scholar]
- Hwang, H.S.; Han, J.Y.; Choi, Y.E. Enhanced accumulation of pinosylvin stilbenes and related gene expression in Pinus strobus after infection of pine wood nematode. Tree Physiol. 2021, 41, 1972–1987. [Google Scholar] [CrossRef]
- Hwang, H.S.; Kim, Y.R.; Han, J.Y.; Choi, Y.E. Nematicidal properties and chemical composition of Pinus rigida Mill. resin against pinewood nematodes. Forests 2022, 13, 1131. [Google Scholar] [CrossRef]
- Ramachandra, R.; Ravishankar, G.A. Plant cell cultures: Chemical factories of secondary metabolites. Biotechnol. Adv. 2002, 20, 101–153. [Google Scholar] [CrossRef]
- Mulabagal, V.; Tsay, H.S. Plant cell cultures—An alternative and efficient source for the production of biologically important secondary metabolites. Int. J. Eng. Sci. 2004, 2, 29–48. [Google Scholar]
- Sotheeswaran, S.; Pasupathy, V. Distribution of resveratrol oligomers in plants. Phtyochemistry 1993, 32, 1083–1092. [Google Scholar] [CrossRef]
- Jeandet, P.; Vannozzi, A.; Sobarzo-Sánchez, E.; Uddin, M.S.; Bru, R.; Martínez-Márquez, A.; Clement, C.; Cordelier, S.; Manayi, A.; Nabavi, S.F.; et al. Phytostilbenes as agrochemicals: Biosynthesis, bioactivity, metabolic engineering and biotechnology. Nat. Prod. Rep. 2021, 38, 1282–1329. [Google Scholar] [CrossRef]
- Lange, B.M.; Trost, M.; Heller, W.; Langebartels, C.; Sandermann, H. Elicitor-Induced formation of free and cell-wall-bound stilbenes in cell-suspension cultures of Scots pine (Pinus sylvestris L.). Planta 1994, 194, 143–148. [Google Scholar] [CrossRef]
- Koo, H.B.; Hwang, H.; Han, J.Y.; Cheong, E.J.; Kwon, Y.; Choi, Y.E. Enhanced production of pinosylvin stilbene with aging of Pinus strobus callus and nematicidal activity of callus extracts against pinewood nematodes. Sci. Rep. 2022, 12, 770. [Google Scholar] [CrossRef]
- Hyun, M.W.; Kim, J.H.; Suh, D.Y.; Lee, S.K.; Kim, S.H. Fungi isolated from pine wood nematode, its vector Japanese pine sawyer, and the nematode-infected Japanese black pine wood in Korea. Mycobiology 2007, 35, 159–161. [Google Scholar] [CrossRef]
- Gehlert, R.; Schöppner, A.; Kindl, H. Stilbene synthase from seedlings of Pinus sylvestris: Purification and induction in response to fungal infection. Mol. Plant Microbe Interact. 1990, 3, 444–449. [Google Scholar] [CrossRef]
- Chiron, H.; Drouet, A.; Claudot, A.C.; Eckerskorn, C.; Trost, M.; Heller, W.; Ernst, D.; Sandermann, H., Jr. Molecular cloning and functional expression of a stress-induced multifunctional O-methyltransferase with pinosylvin methyltransferase activity from Scots pine (Pinus sylvestris L.). Plant Mol. Biol. 2000, 446, 733–745. [Google Scholar] [CrossRef]
- Bakrim, S.; Machate, H.; Benali, T.; Sahib, N.; Jaouadi, I.; Omari, N.E.; Aboulaghras, S.; Bangar, S.P.; Lorenzo, J.M.; Zengin, G.; et al. Natural sources and pharmacological properties of pinosylvin. Plants 2022, 11, 1541. [Google Scholar] [CrossRef]
- Sriwati, R.; Takemoto, S.; Futai, K. Cohabitation of the pine wood nematode, Bursaphelenchus xylophilus, and fungal species in pine trees inoculated with B. xylophilus. Nematology 2007, 9, 77–86. [Google Scholar]
- Dou, G.; Yan, D.-H. Research progress on biocontrol of pine wilt disease by microorganisms. Forests 2022, 13, 1047. [Google Scholar] [CrossRef]
- Willför, S.; Hemming, J.; Reunanen, M.; Holmbom, B. Phenolic and lipophilic extractives in Scots pine knots and stemwood. Holzforschung 2003, 57, 359–372. [Google Scholar] [CrossRef]
- Jorgensen, E. The formation of pinosylvin and its monomethyl ether in the sapwood of Pinus resinosa Ait. Can. J. Bot. 1996, 39, 1765–1772. [Google Scholar] [CrossRef]
- Yamada, T.; Ito, S.I. Chemical defense responses of wilt- resistant pine species, Pinus strobus and P. taeda, against Bursaphelenchus xylophilus infection. Nippon Shokubutsu Byori Gakkaiho 1993, 59, 666–672. [Google Scholar] [CrossRef]
- Litvay, J.D.; Verma, D.C.; Johnson, M.A. Influence of loblolly pine (Pinus taeda L.). Culture medium and its components on growth and somatic embryogenesis of the wild carrot (Daucus carota L.). Plant Cell Rep. 1985, 4, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Coke, J.E. Basal Nutrient Medium for In Vitro Cultures of Loblolly Pines. U.S. Patent 5534434, 9 July 1996. Available online: http://www.freepatentsonline.com/55344345.pdf (accessed on 20 June 2011).
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Baldi, A.; Srivastava, A.K.; Bisaria, V.S. Fungal elicitors for enhanced production of secondary metabolites in plant cell suspension cultures. Soil Biol. 2009, 18, 373–380. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real time quantitative PCR and the 2−∆∆CT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Hooper, D.J. Extraction of nematodes from plant material. In Laboratory Methods for Work with Plant and Soil Nematodes; Reference Book No., 402; Southey, J.F., Ed.; Ministry of Agriculture, Fisheries and Food, Her Majesty’s Stationery Office: London, UK, 1986; pp. 51–58. [Google Scholar]
- He, J.; Zheng, Z.P.; Zhu, Q.; Guo, F.; Chen, J. Encapsulation mechanism of oxyresveratrol by β-cyclodextrin and hydroxypropyl-β-cyclodextrin and computational analysis. Molecules 2017, 22, 1801. [Google Scholar] [CrossRef] [PubMed]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.R.; Han, J.Y.; Choi, Y.E. Production of Nematicidal Pinosylvin Stilbenes in Cell Suspension Cultures of Pinus koraiensis by Fungal Elicitation. Plants 2022, 11, 2933. https://doi.org/10.3390/plants11212933
Kim YR, Han JY, Choi YE. Production of Nematicidal Pinosylvin Stilbenes in Cell Suspension Cultures of Pinus koraiensis by Fungal Elicitation. Plants. 2022; 11(21):2933. https://doi.org/10.3390/plants11212933
Chicago/Turabian StyleKim, Yi Rae, Jung Yeon Han, and Yong Eui Choi. 2022. "Production of Nematicidal Pinosylvin Stilbenes in Cell Suspension Cultures of Pinus koraiensis by Fungal Elicitation" Plants 11, no. 21: 2933. https://doi.org/10.3390/plants11212933
APA StyleKim, Y. R., Han, J. Y., & Choi, Y. E. (2022). Production of Nematicidal Pinosylvin Stilbenes in Cell Suspension Cultures of Pinus koraiensis by Fungal Elicitation. Plants, 11(21), 2933. https://doi.org/10.3390/plants11212933