
Tracing the Evolution of the SEPALLATA Subfamily across Angiosperms Associated with Neo- and Sub-Functionalization for Reproductive and Agronomically Relevant Traits


Abstract
1. Introduction
2. Results and Discussion
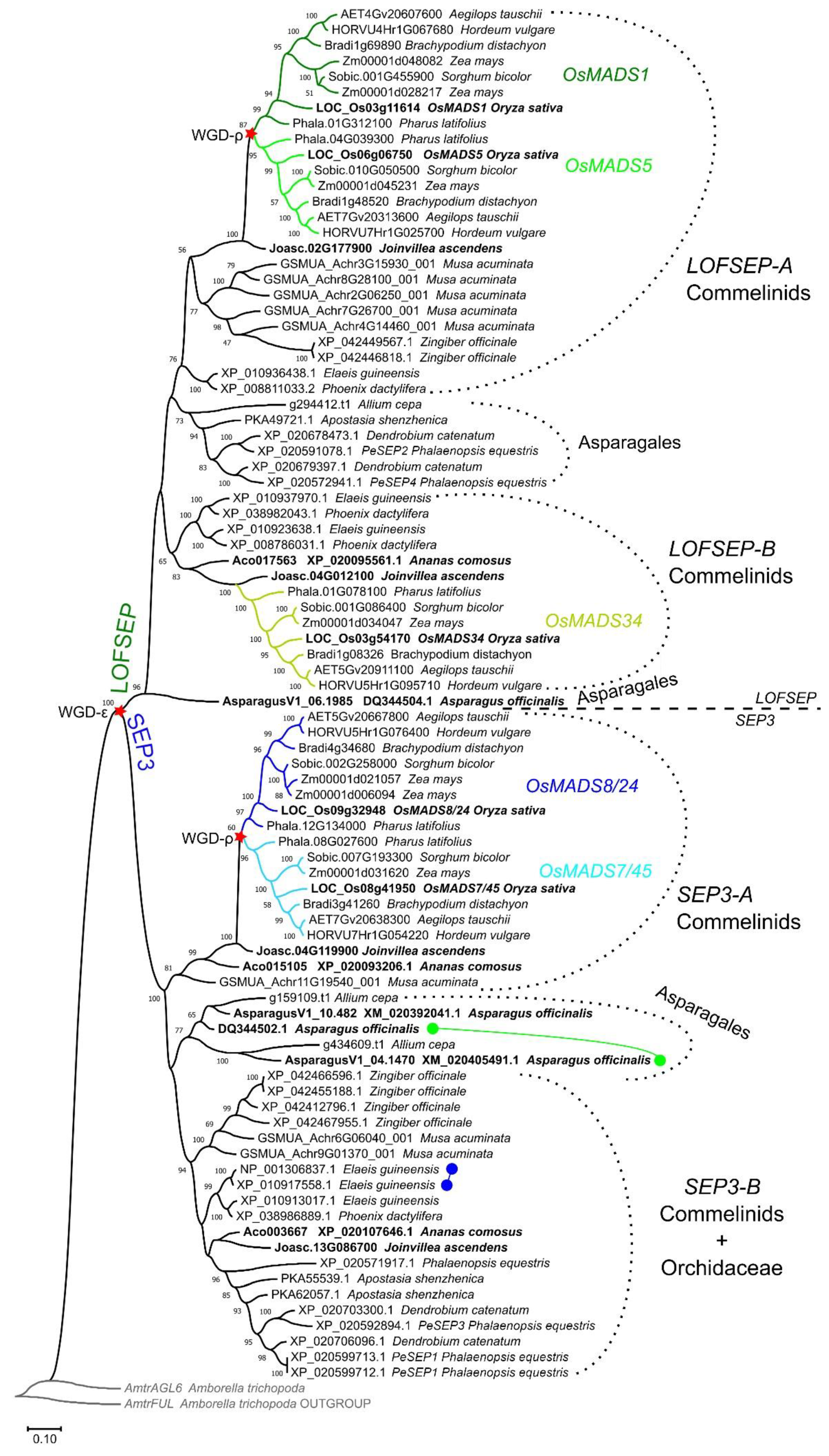
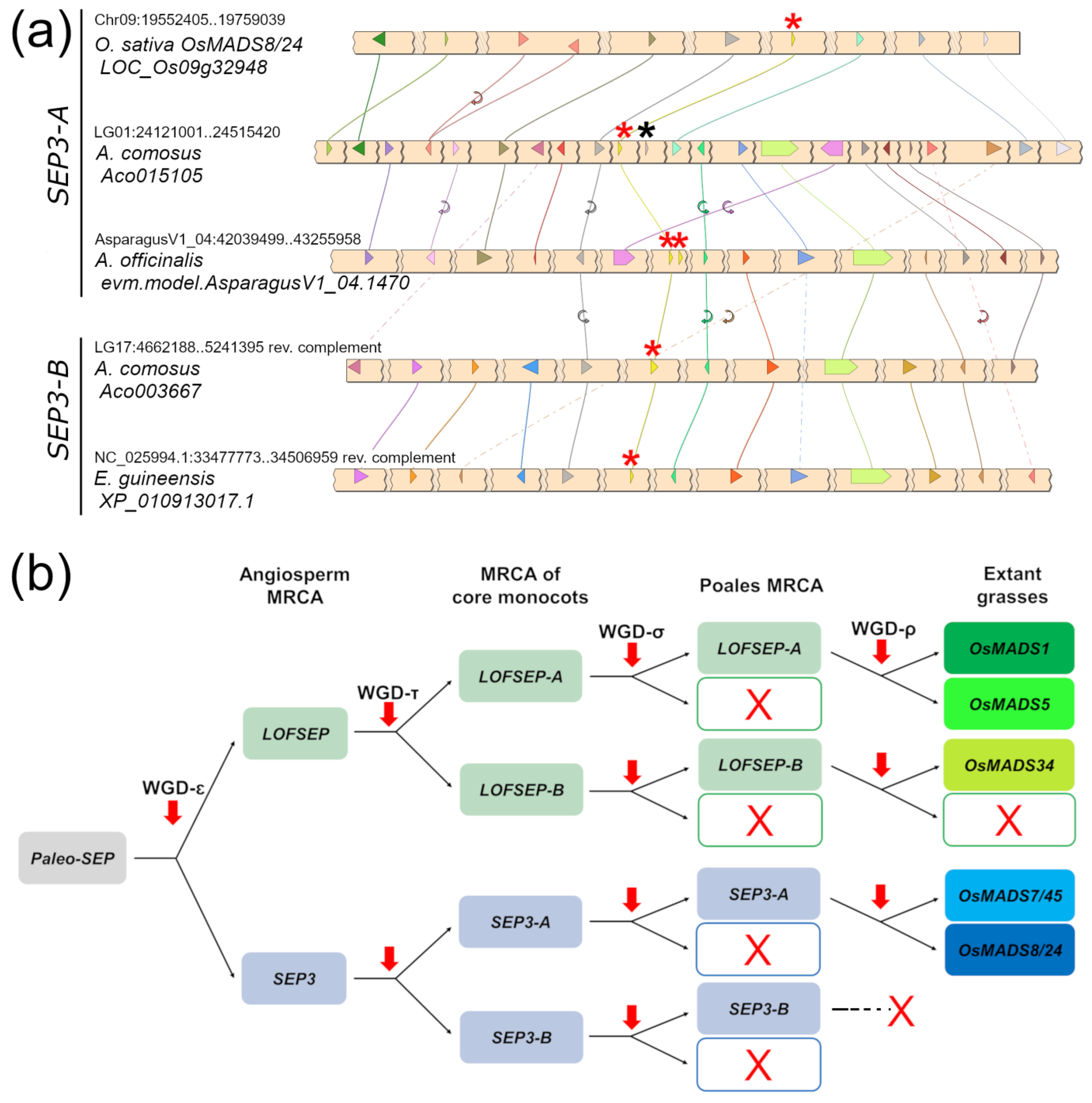
2.1. Evolution of the SEPALLATA Subfamily in Core Monocots
Deciphering the Evolution of SEPALLATA Genes along the Lineage That Led to Grasses
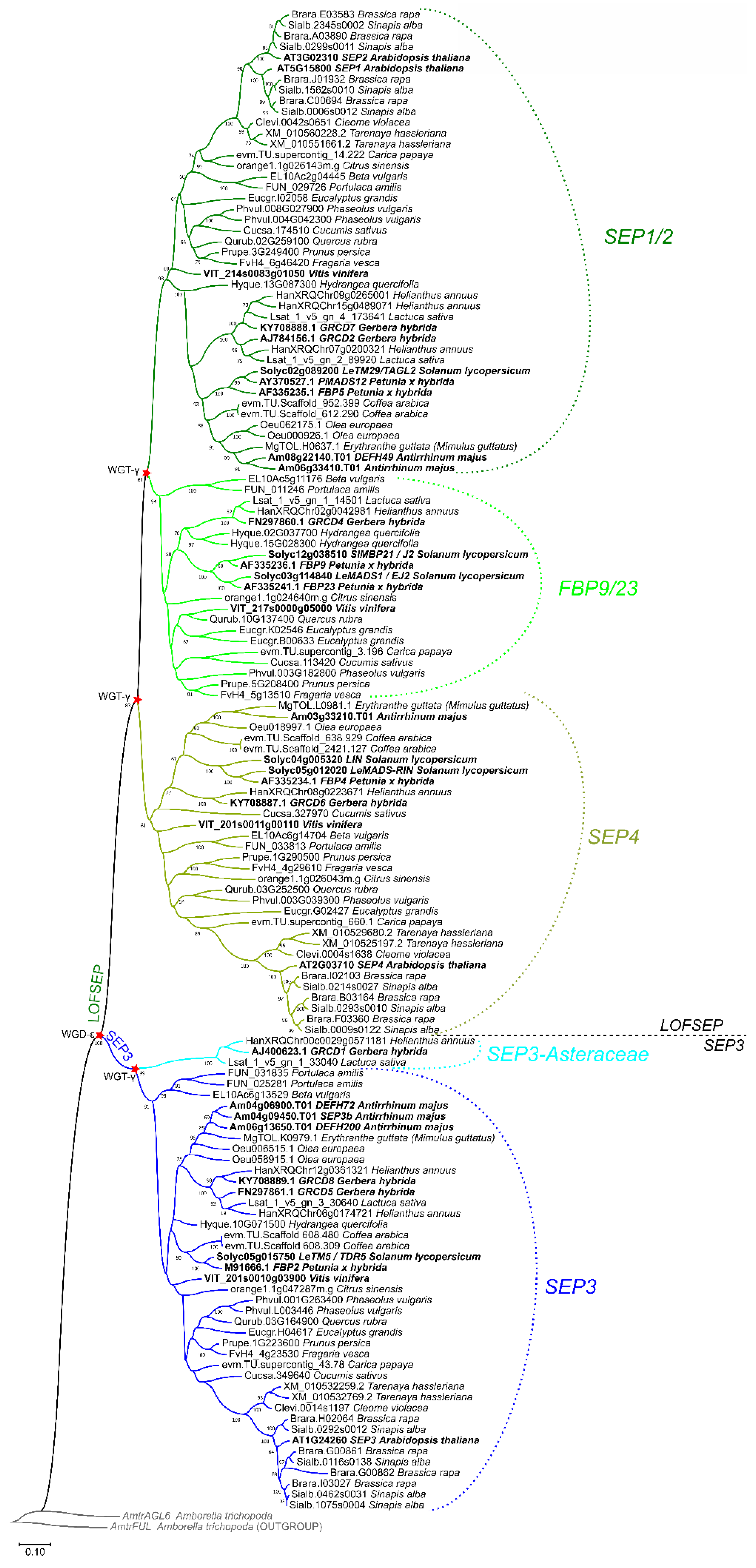
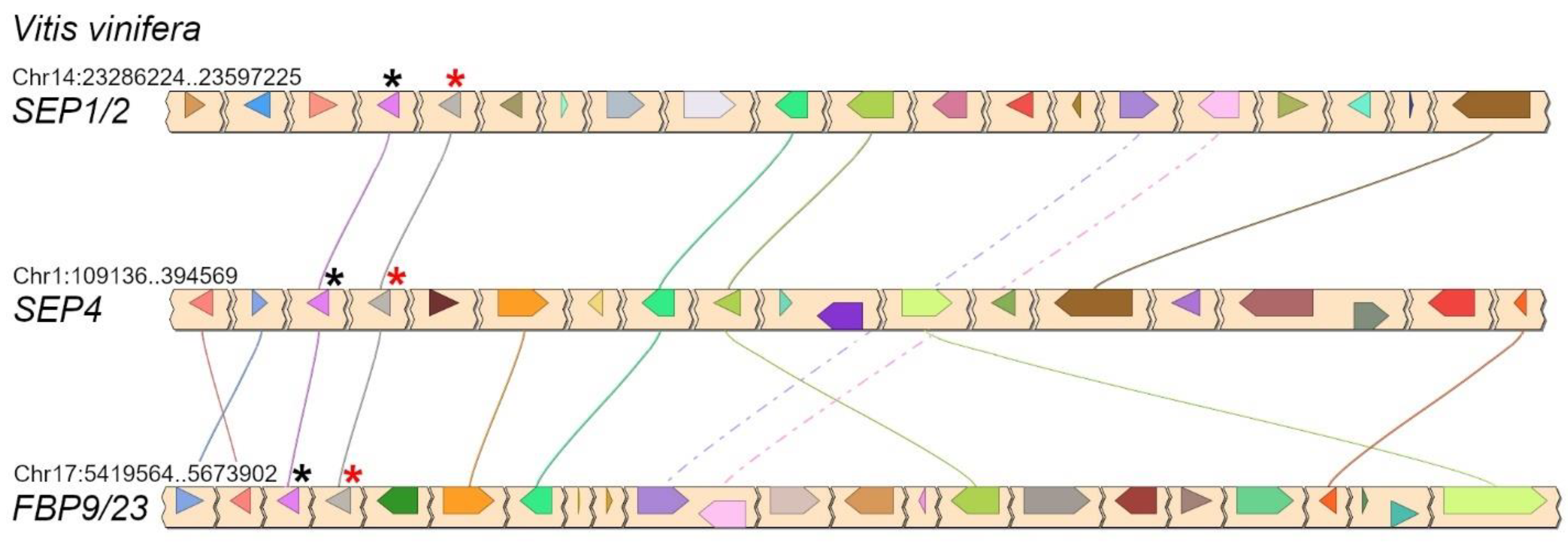
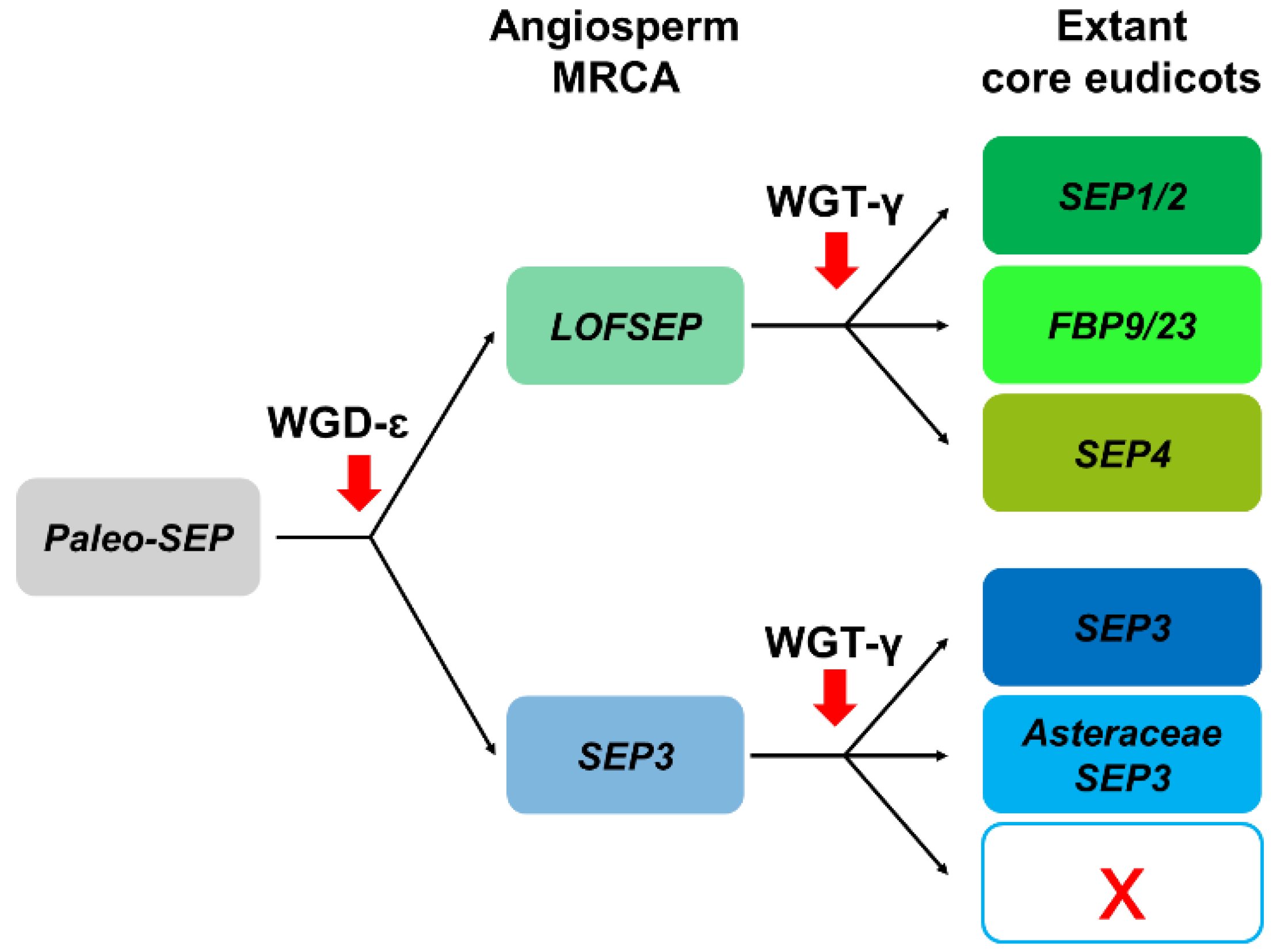
2.2. Three LOFSEP Sister Clades and a Single SEP3 Clade Evolved in Core Eudicots
2.3. Conserved Genetic Linkage between SEPALLATA, SQUAMOSA and FLOWERING LOCUS C Subfamilies
2.4. Patterns of Sub- and neo-Functionalization Associated with Diverged SEPALLATA Lineages
3. Conclusions
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Coen, E.; Meyerowitz, E. The War of the Whorls: Genetic Interactions Controlling Flower Development. Nature 1991, 353, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Theißen, G.; Saedler, H. Floral Quartets. Nature 2001, 409, 469–471. [Google Scholar] [CrossRef] [PubMed]
- Puranik, S.; Acajjaoui, S.; Conn, S.; Costa, L.; Conn, V.; Vial, A.; Marcellin, R.; Melzer, R.; Brown, E.; Hart, D.; et al. Structural Basis for the Oligomerization of the MADS Domain Transcription Factor SEPALLATA3 in Arabidopsis. Plant Cell 2014, 26, 3603–3615. [Google Scholar] [CrossRef] [PubMed]
- Smaczniak, C.; Immink, R.G.H.; Muiño, J.M.; Blanvillain, R.; Busscher, M.; Busscher-Lange, J.; Dinh, Q.D.; Liu, S.; Westphal, A.H.; Boeren, S.; et al. Characterization of MADS-Domain Transcription Factor Complexes in Arabidopsis Flower Development. Proc. Natl. Acad. Sci. USA 2012, 109, 1560–1565. [Google Scholar] [CrossRef]
- Hugouvieux, V.; Silva, C.S.; Jourdain, A.; Stigliani, A.; Charras, Q.; Conn, V.; Conn, S.J.; Carles, C.C.; Parcy, F.; Zubieta, C. Tetramerization of MADS Family Transcription Factors SEPALLATA3 and AGAMOUS Is Required for Floral Meristem Determinacy in Arabidopsis. Nucleic Acids Res. 2018, 46, 4966–4977. [Google Scholar] [CrossRef] [PubMed]
- Theißen, G.; Melzer, R.; Ruümpler, F. MADS-Domain Transcription Factors and the Floral Quartet Model of Flower Development: Linking Plant Development and Evolution. Development 2016, 143, 3259–3271. [Google Scholar] [CrossRef]
- Pelaz, S.; Ditta, G.S.; Baumann, E.; Wisman, E.; Yanofsky, M.F. B and C Floral Organ Identity Functions Require SEPALLATA MADS-Box Genes. Nature 2000, 405, 200–203. [Google Scholar] [CrossRef]
- Pelaz, S.; Tapia-López, R.; Alvarez-Buylla, E.R.; Yanofsky, M.F. Conversion of Leaves into Petals in Arabidopsis. Curr. Biol. 2001, 11, 182–184. [Google Scholar] [CrossRef]
- Favaro, R.; Pinyopich, A.; Battaglia, R.; Kooiker, M.; Borghi, L.; Ditta, G.; Yanofsky, M.F.; Kater, M.M.; Colombo, L. MADS-Box Protein Complexes Control Carpel and Ovule Development in Arabidopsis. Plant Cell 2003, 15, 2603–2611. [Google Scholar] [CrossRef]
- Ditta, G.; Pinyopich, A.; Robles, P.; Pelaz, S.; Yanofsky, M.F. The SEP4 Gene of Arabidopsis Thaliana Functions in Floral Organ and Meristem Identity. Curr. Biol. 2004, 14, 1935–1940. [Google Scholar] [CrossRef]
- Goethe, J.W. Versuch Die Metamorphose Der Pflanzen Zu Erklären; Ettingersche Buchhandlung: Gotha, Germany, 1790. [Google Scholar]
- Irish, V.F. Variations on a Theme: Flower Development and Evolution. Genome Biol. 2000, 1, reviews1015.1. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ronse De Craene, L.P.; Brockington, S.F. Origin and Evolution of Petals in Angiosperms. Plant Ecol. Evol. 2013, 146, 5–25. [Google Scholar] [CrossRef]
- Kellogg, E. A Evolutionary History of the Grasses. Plant Physiol. 2001, 125, 1198–1205. [Google Scholar] [CrossRef]
- Causier, B.; Schwarz-Sommer, Z.; Davies, B. Floral Organ Identity: 20 Years of ABCs. Semin. Cell Dev. Biol. 2010, 21, 73–79. [Google Scholar] [CrossRef]
- Chanderbali, A.S.; Berger, B.A.; Howarth, D.G.; Soltis, P.S.; Soltis, D.E. Evolving Ideas on the Origin and Evolution of Flowers: New Perspectives in the Genomic Era. Genetics 2016, 202, 1255–1265. [Google Scholar] [CrossRef]
- Endress, P.K. Disentangling Confusions in Inflorescence Morphology: Patterns and Diversity of Reproductive Shoot Ramification in Angiosperms. J. Syst. Evol. 2010, 48, 225–239. [Google Scholar] [CrossRef]
- Claßen-Bockhoff, R.; Bull-Hereñu, K. Towards an Ontogenetic Understanding of Inflorescence Diversity. Ann. Bot. 2013, 112, 1523–1542. [Google Scholar] [CrossRef]
- Kellogg, E.A. Genetic Control of Branching Patterns in Grass Inflorescences. Plant Cell 2022, 34, 2518–2533. [Google Scholar] [CrossRef]
- Bartlett, M.E.; Thompson, B. Meristem Identity and Phyllotaxis in Inflorescence Development. Front. Plant Sci. 2014, 2014, 508. [Google Scholar] [CrossRef]
- Bommert, P.; Whipple, C. Grass Inflorescence Architecture and Meristem Determinacy. Semin. Cell Dev. Biol. 2018, 79, 37–47. [Google Scholar]
- Kyozuka, J. Chapter Seven—Grass Inflorescence: Basic Structure and Diversity. In Advances in Botanical Research; Academic Press; Elsevier: Oxford, UK, 2014; Volume 72, pp. 191–219. [Google Scholar]
- Zhang, D.; Yuan, Z. Molecular Control of Grass Inflorescence Development. Annu. Rev. Plant Biol. 2014, 65, 553–578. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, K.; Hibrand-Saint Oyant, L.; Crespel, L.; Thouroude, T.; Lalanne, D.; Foucher, F. Quantitative Trait Loci for Flowering Time and Inflorescence Architecture in Rose. Theor. Appl. Genet. 2011, 122, 661–675. [Google Scholar] [CrossRef]
- Benlloch, R.; Berbel, A.; Ali, L.; Gohari, G.; Millán, T.; Madueño, F. Genetic Control of Inflorescence Architecture in Legumes. Front. Plant Sci. 2015, 6, 543. [Google Scholar] [CrossRef]
- Soyk, S.; Lemmon, Z.H.; Oved, M.; Fisher, J.; Liberatore, K.L.; Park, S.J.; Goren, A.; Jiang, K.; Ramos, A.; van der Knaap, E.; et al. Bypassing Negative Epistasis on Yield in Tomato Imposed by a Domestication Gene. Cell 2017, 169, 1142–1155. [Google Scholar] [CrossRef]
- Soyk, S.; Lemmon, Z.H.; Sedlazeck, F.J.; Jiménez-Gómez, J.M.; Alonge, M.; Hutton, S.F.; Van Eck, J.; Schatz, M.C.; Lippman, Z.B. Duplication of a Domestication Locus Neutralized a Cryptic Variant That Caused a Breeding Barrier in Tomato. Nat. Plants 2019, 5, 471–479. [Google Scholar] [CrossRef]
- Kobayashi, K.; Maekawa, M.; Miyao, A.; Hirochika, H.; Kyozuka, J. PANICLE PHYTOMER2 (PAP2), Encoding a SEPALLATA Subfamily MADS-Box Protein, Positively Controls Spikelet Meristem Identity in Rice. Plant Cell Physiol. 2010, 51, 41–51. [Google Scholar] [CrossRef]
- Gao, X.; Liang, W.; Yin, C.; Ji, S.; Wang, H.; Su, X.; Guo, C.; Kong, H.; Xue, H.; Zhang, D. The SEPALLATA-like Gene OsMADS34 Is Required for Rice Inflorescence and Spikelet Development. Plant Physiol. 2010, 110, 728–740. [Google Scholar] [CrossRef]
- Zhu, W.; Yang, L.; Wu, D.; Meng, Q.; Deng, X.; Huang, G.; Zhang, J.; Chen, X.; Ferrándiz, C.; Liang, W.; et al. Rice SEPALLATA Genes OsMADS5 and OsMADS34 Cooperate to Limit Inflorescence Branching by Repressing the TERMINAL FLOWER1-like Gene RCN4. New Phytol. 2022, 233, 1682–1700. [Google Scholar] [CrossRef]
- Zhang, T.; Zhao, Y.; Juntheikki, I.; Mouhu, K.; Broholm, S.K.; Rijpkema, A.S.; Kins, L.; Lan, T.; Albert, V.A.; Teeri, T.H.; et al. Dissecting Functions of SEPALLATA-like MADS Box Genes in Patterning of the Pseudanthial Inflorescence of Gerbera Hybrida. New Phytol. 2017, 216, 939–954. [Google Scholar] [CrossRef]
- Malcomber, S.T.; Kellogg, E.A. SEPALLATA Gene Diversification: Brave New Whorls. Trends Plant Sci. 2005, 10, 427–435. [Google Scholar] [CrossRef]
- Zahn, L.M.; Kong, H.; Leebens-Mack, J.H.; Kim, S.; Soltis, P.S.; Landherr, L.L.; Soltis, D.E.; DePamphilis, C.W.; Ma, H. The Evolution of the SEPALLATA Subfamily of MADS-Box Genes: A Preangiosperm Origin with Multiple Duplications throughout Angiosperm History. Genetics 2005, 169, 2209–2223. [Google Scholar] [CrossRef] [PubMed]
- DePamphilis, C.W.; Palmer, J.D.; Rounsley, S.; Sankoff, D.; Schuster, S.C.; Ammiraju, J.S.S.; Barbazuk, W.B.; Chamala, S.; Chanderbali, A.S.; Determann, R.; et al. The Amborella Genome and the Evolution of Flowering Plants. Science 2013, 342, 1241089. [Google Scholar] [CrossRef]
- Ruelens, P.; De Maagd, R.A.; Proost, S.; Theißen, G.; Geuten, K.; Kaufmann, K. FLOWERING LOCUS C in Monocots and the Tandem Origin of Angiosperm-Specific MADS-Box Genes. Nat. Commun. 2013, 4, 2280. [Google Scholar] [CrossRef]
- Airoldi, C.A.; Davies, B. Gene Duplication and the Evolution of Plant MADS-Box Transcription Factors. J. Genet. Genomics 2012, 39, 157–165. [Google Scholar] [CrossRef]
- Causier, B.; Castillo, R.; Xue, Y.; Schwarz-Sommer, Z.; Davies, B. Tracing the Evolution of the Floral Homeotic B-and C-Function Genes through Genome Synteny. Mol. Biol. Evol. 2010, 27, 2651–2664. [Google Scholar] [CrossRef]
- Zhao, T.; Holmer, R.; De Bruijn, S.; Angenent, G.C.; Van Den Burg, H.A.; Eric Schranz, M. Phylogenomic Synteny Network Analysis of MADS-Box Transcription Factor Genes Reveals Lineage-Specific Transpositions, Ancient Tandem Duplications, and Deep Positional Conservation. Plant Cell 2017, 29, 1278–1292. [Google Scholar] [CrossRef]
- Cantino, P.D.; Doyle, J.A.; Graham, S.W.; Judd, W.S.; Olmstead, R.G.; Soltis, D.E.; Soltis, P.S.; Donoghue, M.J. Towards a Phylogenetic Nomenclature of Tracheophyta. Taxon 2007, 56, E1–E44. [Google Scholar] [CrossRef]
- Coiffard, C.; Kardjilov, N.; Manke, I.; Bernardes-de-Oliveira, M.E.C. Fossil Evidence of Core Monocots in the Early Cretaceous. Nat. Plants 2019, 5, 691–696. [Google Scholar] [CrossRef]
- Ming, R.; VanBuren, R.; Wai, C.M.; Tang, H.; Schatz, M.C.; Bowers, J.E.; Lyons, E.; Wang, M.L.; Chen, J.; Biggers, E.; et al. The Pineapple Genome and the Evolution of CAM Photosynthesis. Nat. Genet. 2015, 47, 1435–1442. [Google Scholar] [CrossRef]
- Xu, Q.; Jin, L.; Zhang, Y.; Zhang, X.; Zheng, C.; Leebens-Mack, J.H.; Sankoff, D. Ancestral Flowering Plant Chromosomes and Gene Orders Based on Generalized Adjacencies and Chromosomal Gene Co-Occurrences. J. Comput. Biol. 2021, 28, 1156–1179. [Google Scholar] [CrossRef]
- Tang, H.; Bowers, J.E.; Wang, X.; Paterson, A.H. Angiosperm Genome Comparisons Reveal Early Polyploidy in the Monocot Lineage. Proc. Natl. Acad. Sci. USA 2010, 107, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Arora, R.; Agarwal, P.; Ray, S.; Singh, A.K.; Singh, V.P.; Tyagi, A.K.; Kapoor, S. MADS-Box Gene Family in Rice: Genome-Wide Identification, Organization and Expression Profiling during Reproductive Development and Stress. BMC Genomics 2007, 8, 242. [Google Scholar] [CrossRef] [PubMed]
- Messing, J. The Polyploid Origin of Maize. In Handbook of Maize: Genetics and Genomics; Springer: New York, NY, USA, 2009; pp. 221–238. ISBN 978-0-387-77863-1. [Google Scholar]
- Swigoňová, Z.; Lai, J.; Ma, J.; Ramakrishna, W.; Llaca, V.; Bennetzen, J.L.; Messing, J. Close Split of Sorghum and Maize Genome Progenitors. Genome Res. 2004, 14, 1916–1923. [Google Scholar] [CrossRef] [PubMed]
- Schnable, P.S.; Ware, D.; Fulton, R.S.; Stein, J.C.; Wei, F.; Pasternak, S.; Liang, C.; Zhang, J.; Fulton, L.; Graves, T.A.; et al. The B73 Maize Genome: Complexity, Diversity, and Dynamics. Science 2009, 326, 1112–1115. [Google Scholar] [CrossRef]
- Matsuoka, Y. Evolution of Polyploid Triticum Wheats under Cultivation: The Role of Domestication, Natural Hybridization and Allopolyploid Speciation in Their Diversification. Plant Cell Physiol. 2011, 52, 750–764. [Google Scholar] [CrossRef] [PubMed]
- Shitsukawa, N.; Tahira, C.; Kassai, K.I.; Hirabayashi, C.; Shimizu, T.; Takumi, S.; Mochida, K.; Kawaura, K.; Ogihara, Y.; Muraia, K. Genetic and Epigenetic Alteration among Three Homoeologous Genes of a Class E MADS Box Gene in Hexaploid Wheat. Plant Cell 2007, 19, 1723–1737. [Google Scholar] [CrossRef]
- Thiel, T.; Graner, A.; Waugh, R.; Grosse, I.; Close, T.J.; Stein, N. Evidence and Evolutionary Analysis of Ancient Whole-Genome Duplication in Barley Predating the Divergence from Rice. BMC Evol. Biol. 2009, 9, 209. [Google Scholar] [CrossRef]
- Meng, Q.; Li, X.; Zhu, W.; Yang, L.; Liang, W.; Dreni, L.; Zhang, D. Regulatory Network and Genetic Interactions Established by OsMADS34 in Rice Inflorescence and Spikelet Morphogenesis. J. Integr. Plant Biol. 2017, 59, 693–707. [Google Scholar] [CrossRef]
- Vekemans, D.; Proost, S.; Vanneste, K.; Coenen, H.; Viaene, T.; Ruelens, P.; Maere, S.; Van De Peer, Y.; Geuten, K. Gamma Paleohexaploidy in the Stem Lineage of Core Eudicots: Significance for MADS-BOX Gene and Species Diversification. Mol. Biol. Evol. 2012, 29, 3793–3806. [Google Scholar] [CrossRef]
- Jiao, Y.; Leebens-Mack, J.; Ayyampalayam, S.; Bowers, J.E.; McKain, M.R.; McNeal, J.; Rolf, M.; Ruzicka, D.R.; Wafula, E.; Wickett, N.J.; et al. A Genome Triplication Associated with Early Diversification of the Core Eudicots. Genome Biol. 2012, 13, R3. [Google Scholar] [CrossRef]
- Chanderbali, A.S.; Jin, L.; Xu, Q.; Zhang, Y.; Zhang, J.; Jian, S.; Carroll, E.; Sankoff, D.; Albert, V.A.; Howarth, D.G.; et al. Buxus and Tetracentron Genomes Help Resolve Eudicot Genome History. Nat. Commun. 2022, 13, 643. [Google Scholar] [CrossRef] [PubMed]
- Jaillon, O.; Aury, J.M.; Noel, B.; Policriti, A.; Clepet, C.; Casagrande, A.; Choisne, N.; Aubourg, S.; Vitulo, N.; Jubin, C.; et al. The Grapevine Genome Sequence Suggests Ancestral Hexaploidization in Major Angiosperm Phyla. Nature 2007, 449, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Chanderbali, A.S.; Berger, B.A.; Howarth, D.G.; Soltis, D.E.; Soltis, P.S. Evolution of Floral Diversity: Genomics, Genes and Gamma. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20150509. [Google Scholar] [CrossRef]
- Vrebalov, J.; Ruezinsky, D.; Padmanabhan, V.; White, R.; Medrano, D.; Drake, R.; Schuch, W.; Giovannoni, J. A MADS-Box Gene Necessary for Fruit Ripening at the Tomato Ripening-Inhibitor (Rin) Locus. Science 2002, 296, 343–346. [Google Scholar] [CrossRef]
- Kobayashi, K.; Yasuno, N.; Sato, Y.; Yoda, M.; Yamazaki, R.; Kimizu, M.; Yoshida, H.; Nagamura, Y.; Kyozukaa, J. Inflorescence Meristem Identity in Rice Is Specified by Overlapping Functions of Three AP1/FUL-Like MADS Box Genes and PAP2, a SEPALLATA MADS Box Gene. Plant Cell 2012, 24, 1848–1859. [Google Scholar] [CrossRef] [PubMed]
- Trevaskis, B.; Bagnall, D.J.; Ellis, M.H.; Peacock, W.J.; Dennis, E.S. MADS Box Genes Control Vernalization-Induced Flowering in Cereals. Proc. Natl. Acad. Sci. USA 2003, 100, 13099–13104. [Google Scholar] [CrossRef]
- Robinson, R.; Tomes, M. Ripening Inhibitor: A Gene with Multiple Effects on Ripening. Rep. Tomato Genet. Coop. 1968, 18, 36–37. [Google Scholar]
- Ito, Y.; Nishizawa-Yokoi, A.; Endo, M.; Mikami, M.; Shima, Y.; Nakamura, N.; Kotake-Nara, E.; Kawasaki, S.; Toki, S. Re-Evaluation of the Rin Mutation and the Role of RIN in the Induction of Tomato Ripening. Nat. Plants 2017, 3, 866–874. [Google Scholar] [CrossRef]
- Mandel, M.A.; Yanofsky, M.F. The Arabidopsis AGL9 MADS Box Gene Is Expressed in Young Flower Primordia. Sex Plant Reprod. 1998, 11, 22–28. [Google Scholar] [CrossRef]
- Honma, T.; Goto, K. Complexes of MADS-Box Proteins Are Sufficient to Convert Leaves into Floral Organs. Nature 2001, 409, 525–529. [Google Scholar] [CrossRef]
- Ferrario, S.; Immink, R.G.H.; Shchennikova, A.; Busscher-Lange, J.; Angenent, G.C. The MADS Box Gene FBP2 Is Required for SEPALLATA Function in Petunia. Plant Cell 2003, 15, 914–925. [Google Scholar] [CrossRef] [PubMed]
- Pnueli, L.; Hareven, D.; Broday, L.; Hurwitz, C.; Lifschitz, E. The TM5 MADS Box Gene Mediates Organ Differentiation in the Three Inner Whorls of Tomato Flowers. Plant Cell 1994, 6, 175–186. [Google Scholar] [CrossRef] [PubMed]
- Davies, B.; Egea-Cortines, M.; de Andrade Silva, E.; Saedler, H.; Sommer, H. Multiple Interactions amongst Floral Homeotic MADS Box Proteins. EMBO J. 1996, 15, 4330–4343. [Google Scholar] [CrossRef] [PubMed]
- Cui, R.; Han, J.; Zhao, S.; Su, K.; Wu, F.; Du, X.; Xu, Q.; Chong, K.; Theißen, G.; Meng, Z. Functional Conservation and Diversification of Class e Floral Homeotic Genes in Rice (Oryza sativa). Plant J. 2010, 61, 767–781. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.J.; Chen, Y.Y.; Du, J.S.; Chen, Y.Y.; Chung, M.C.; Tsai, W.C.; Wang, C.N.; Chen, H.H. Flower Development of Phalaenopsis Orchid Involves Functionally Divergent SEPALLATA-like Genes. New Phytol. 2014, 202, 1024–1042. [Google Scholar] [CrossRef] [PubMed]
- Vandenbussche, M.; Zethof, J.; Souer, E.; Koes, R.; Tornielli, G.B.; Pezzotti, M.; Ferrario, S.; Angenent, G.C.; Gerats, T. Toward the Analysis of the Petunia MADS Box Gene Family by Reverse and Forward Transposon Insertion Mutagenesis Approaches: B, C, and D Floral Organ Identity Functions Require SEPALLATA-like MADS Box Genes in Petunia. Plant Cell 2003, 15, 2680–2693. [Google Scholar] [CrossRef]
- Angenent, G.C.; Franken, J.; Busscher, M.; Weiss, D.; Van Tunen, A.J. Co-suppression of the Petunia Homeotic Gene Fbp2 Affects the Identity of the Generative Meristem. Plant J. 1994, 5, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Mitoma, M.; Kanno, A. The Greenish Flower Phenotype of Habenaria Radiata (Orchidaceae) Is Caused by a Mutation in the SEPALLATA-like MADS-Box Gene HrSEP-1. Front. Plant Sci. 2018, 9, 831. [Google Scholar] [CrossRef]
- Morel, P.; Chambrier, P.; Boltz, V.; Chamot, S.; Rozier, F.; Bento, S.R.; Trehin, C.; Monniaux, M.; Zethof, J.; Vandenbussche, M. Divergent Functional Diversification Patterns in the SEP/AGL6/AP1 MADS-Box Transcription Factor Superclade. Plant Cell 2019, 31, 3033–3056. [Google Scholar] [CrossRef]
- Wu, D.; Liang, W.; Zhu, W.; Chen, M.; Ferrándiz, C.; Burton, R.A.; Dreni, L.; Zhang, D. Loss of LOFSEP Transcription Factor Function Converts Spikelet to Leaf-like Structures in Rice. Plant Physiol. 2018, 176, 1646–1664. [Google Scholar] [CrossRef]
- Jeon, J.S.; Jang, S.; Lee, S.; Nam, J.; Kim, C.; Lee, S.H.; Chung, Y.Y.; Kim, S.R.; Lee, Y.H.; Cho, Y.G.; et al. Leafy Hull Sterile 1 Is a Homeotic Mutation in a Rice MADS Box Gene Affecting Rice Flower Development. Plant Cell 2000, 12, 871–884. [Google Scholar] [CrossRef] [PubMed]
- Prasad, K.; Parameswaran, S.; Vijayraghavan, U. OsMADS1, a Rice MADS-Box Factor, Controls Differentiation of Specific Cell Types in the Lemma and Palea and Is an Early-Acting Regulator of Inner Floral Organs. Plant J. 2005, 43, 915–928. [Google Scholar] [CrossRef] [PubMed]
- Prasad, K.; Sriram, P.; Santhosh Kumar, C.; Kushalappa, K.; Vijayraghavan, U. Ectopic Expression of Rice OsMADS1 Reveals a Role in Specifying the Lemma and Palea, Grass Floral Organs Analogous to Sepals. Dev. Genes Evol. 2001, 211, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, G.K.; Abe, K.; Yamazaki, M.; Miyao, A.; Hirochika, H. Conservation of the E-Function for Floral Organ Identity in Rice Revealed by the Analysis of Tissue Culture-Induced Loss-of-Function Mutants of the OsMADS1 Gene. Plant Mol. Biol. 2005, 59, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Liang, W.; Yin, C.; Yang, X.; Ping, B.; Li, A.; Jia, R.; Chen, M.; Luo, Z.; Cai, Q.; et al. Interactions of OsMADS1 with Floral Homeotic Genes in Rice Flower Development. Mol. Plant 2015, 8, 1366–1384. [Google Scholar] [CrossRef]
- Lin, X.; Wu, F.; Du, X.; Shi, X.; Liu, Y.; Liu, S.; Hu, Y.; Theißen, G.; Meng, Z. The Pleiotropic SEPALLATA-like Gene OsMADS34 Reveals That the “empty Glumes” of Rice (Oryza Sativa) Spikelets Are in Fact Rudimentary Lemmas. New Phytol. 2014, 202, 689–702. [Google Scholar] [CrossRef]
- Uimari, A.; Kotilainen, M.; Elomaa, P.; Yu, D.; Albert, V.A.; Teeri, T.H. Integration of Reproductive Meristem Fates by a SEPALLATA-like MADS-Box Gene. Proc. Natl. Acad. Sci. USA 2004, 101, 15817–15822. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, H.; Tian, C.; Sajjad, M.; Gao, C.; Tong, Y.; Wang, X.; Jiao, Y. Transcriptome Association Identifies Regulators of Wheat Spike Architecture. Plant Physiol. 2017, 175, 746–757. [Google Scholar] [CrossRef]
- Hussin, S.H.; Wang, H.; Tang, S.; Zhi, H.; Tang, C.; Zhang, W.; Jia, G.; Diao, X. SiMADS34, an E-Class MADS-Box Transcription Factor, Regulates Inflorescence Architecture and Grain Yield in Setaria Italica. Plant Mol. Biol. 2021, 105, 419–434. [Google Scholar] [CrossRef]
- Kater, M.M.; Franken, J.; Inggamer, H.; Gretenkort, M.; Van Tunen, A.J.; Mollema, C.; Angenent, G.C. The Use of Floral Homeotic Mutants as a Novel Way to Obtain Durable Resistance to Insect Pests. Plant Biotechnol. J. 2003, 1, 123–127. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the Number of Nucleotide Substitutions in the Control Region of Mitochondrial DNA in Humans and Chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Bomhoff, M.D.; Briones, E.; Zhang, L.; Schnable, J.C.; Lyons, E. SynFind: Compiling Syntenic Regions across Any Set of Genomes on Demand. Genome Biol. Evol. 2015, 7, 3286–3298. [Google Scholar] [CrossRef]
- Veltri, D.; Wight, M.M.; Crouch, J.A. SimpleSynteny: A Web-Based Tool for Visualization of Microsynteny across Multiple Species. Nucleic Acids Res. 2016, 44, W41–W45. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oryza sativa | P. latifolius | B. distachyon | H. vulgare | A. tauschii | T. aestivum | S. bicolor | Z. mays | ||
|---|---|---|---|---|---|---|---|---|---|
| LOFSEP | OsMADS1 | TraesCS4A02G028100 TraesCS4A02G058900 TraesCS4A02G078700 | Zm00001d028217 Zm00001d048082 | ||||||
| LOC_Os03g11614 | Phala.01G312100 | Bradi1g69890 | HORVU4Hr1G067680 | AET4Gv20607600 AET4Gv20611300 AET4Gv20678000 | TraesCS4B02G245700 TraesCS4B02G245800 TraesCS4B02G277800 | Sobic.001G455900 | |||
| TraesCS4D02G243700 TraesCS4D02G245200 TraesCS4D02G276100 | |||||||||
| OsMADS5 | TraesCS7A02G122000 TraesCS7A02G122100 | ||||||||
| LOC_Os06g06750 | Phala.04G039300 | Bradi1g48520 | HORVU7Hr1G025700 | AET7Gv20313600 AET7Gv20313900 | TraesCS7B02G020800 TraesCS7B02G020900 TraesCS7B02G021000 | Sobic.010G050500 | Zm00001d045231 | ||
| TraesCS7D02G120500 TraesCS7D02G120600 | |||||||||
| OsMADS34 | TraesCS5A02G391800 | ||||||||
| LOC_Os03g54170 | Phala.01G078100 | Bradi1g08326 | HORVU5Hr1G095710 | AET5Gv20911100 | TraesCS5B02G396700 | Sobic.001G086400 | Zm00001d034047 | ||
| TraesCS5D02G401700 | |||||||||
| SEP3 | OsMADS7/45 | TraesCS7A02G260600 | |||||||
| LOC_Os08g41950 | Phala.08G027600 | Bradi3g41260 | HORVU7Hr1G054220 | AET7Gv20638300 | TraesCS7B02G158600 | Sobic.007G193300 | Zm00001d031620 | ||
| TraesCS7D02G261600 | |||||||||
| OsMADS8/24 | TraesCS5A02G286800 | Zm00001d021057 Zm00001d006094 | |||||||
| LOC_Os09g32948 | Phala.12G134000 | Bradi4g34680 | HORVU5Hr1G076400 | AET5Gv20667800 | TraesCS5B02G286100 | Sobic.002G258000 | |||
| TraesCS5D02G294500 | |||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dreni, L.; Ferrándiz, C. Tracing the Evolution of the SEPALLATA Subfamily across Angiosperms Associated with Neo- and Sub-Functionalization for Reproductive and Agronomically Relevant Traits. Plants 2022, 11, 2934. https://doi.org/10.3390/plants11212934
Dreni L, Ferrándiz C. Tracing the Evolution of the SEPALLATA Subfamily across Angiosperms Associated with Neo- and Sub-Functionalization for Reproductive and Agronomically Relevant Traits. Plants. 2022; 11(21):2934. https://doi.org/10.3390/plants11212934
Chicago/Turabian StyleDreni, Ludovico, and Cristina Ferrándiz. 2022. "Tracing the Evolution of the SEPALLATA Subfamily across Angiosperms Associated with Neo- and Sub-Functionalization for Reproductive and Agronomically Relevant Traits" Plants 11, no. 21: 2934. https://doi.org/10.3390/plants11212934
APA StyleDreni, L., & Ferrándiz, C. (2022). Tracing the Evolution of the SEPALLATA Subfamily across Angiosperms Associated with Neo- and Sub-Functionalization for Reproductive and Agronomically Relevant Traits. Plants, 11(21), 2934. https://doi.org/10.3390/plants11212934