
Genome-Wide Identification and Expression Analysis of Wall-Associated Kinase (WAK) Gene Family in Cannabis sativa L.




Abstract
1. Introduction
2. Results
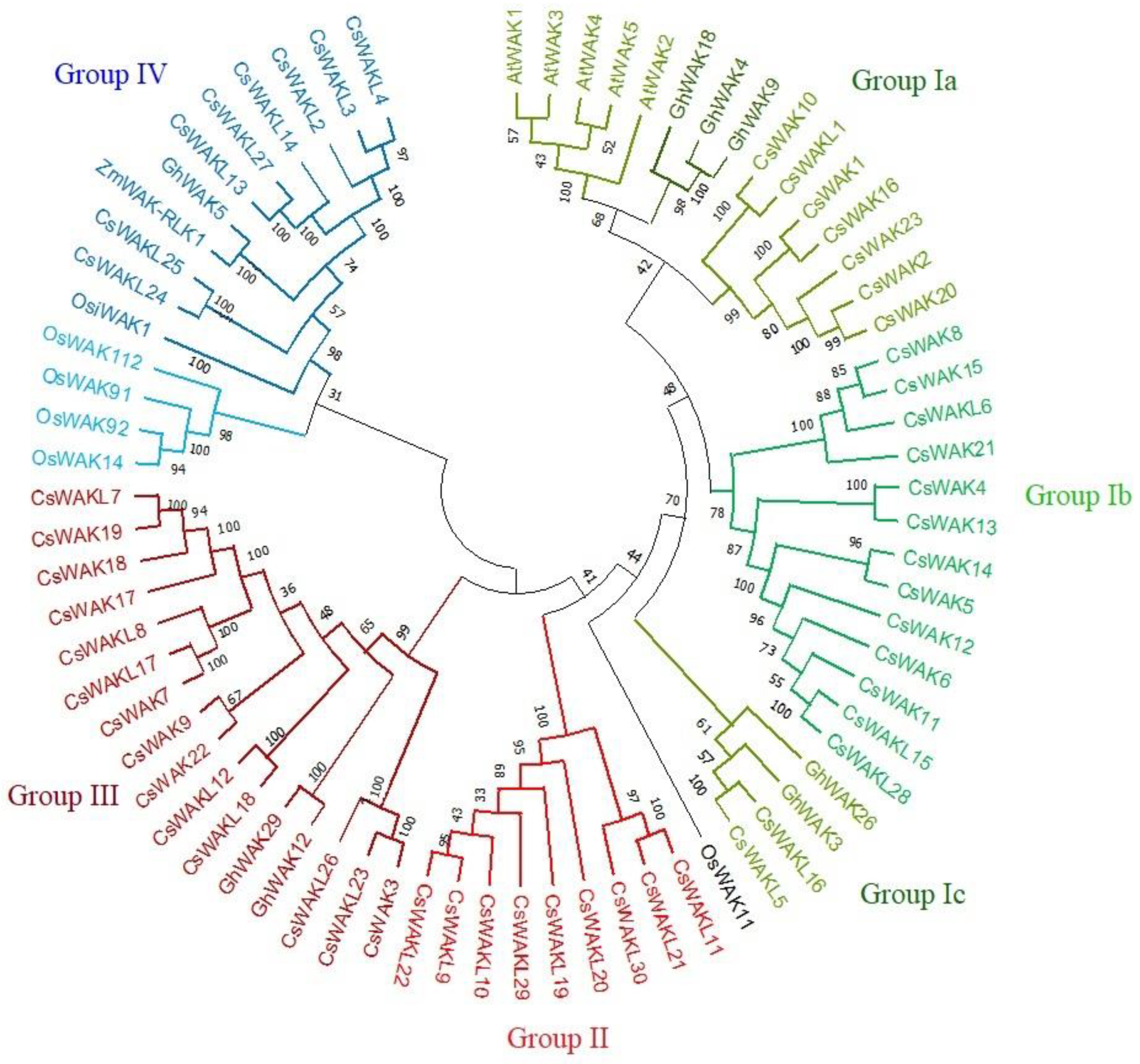
2.1. Identification and Phylogenetic Analysis of CsWAK/CsWAKL Gene Family in Cannabis
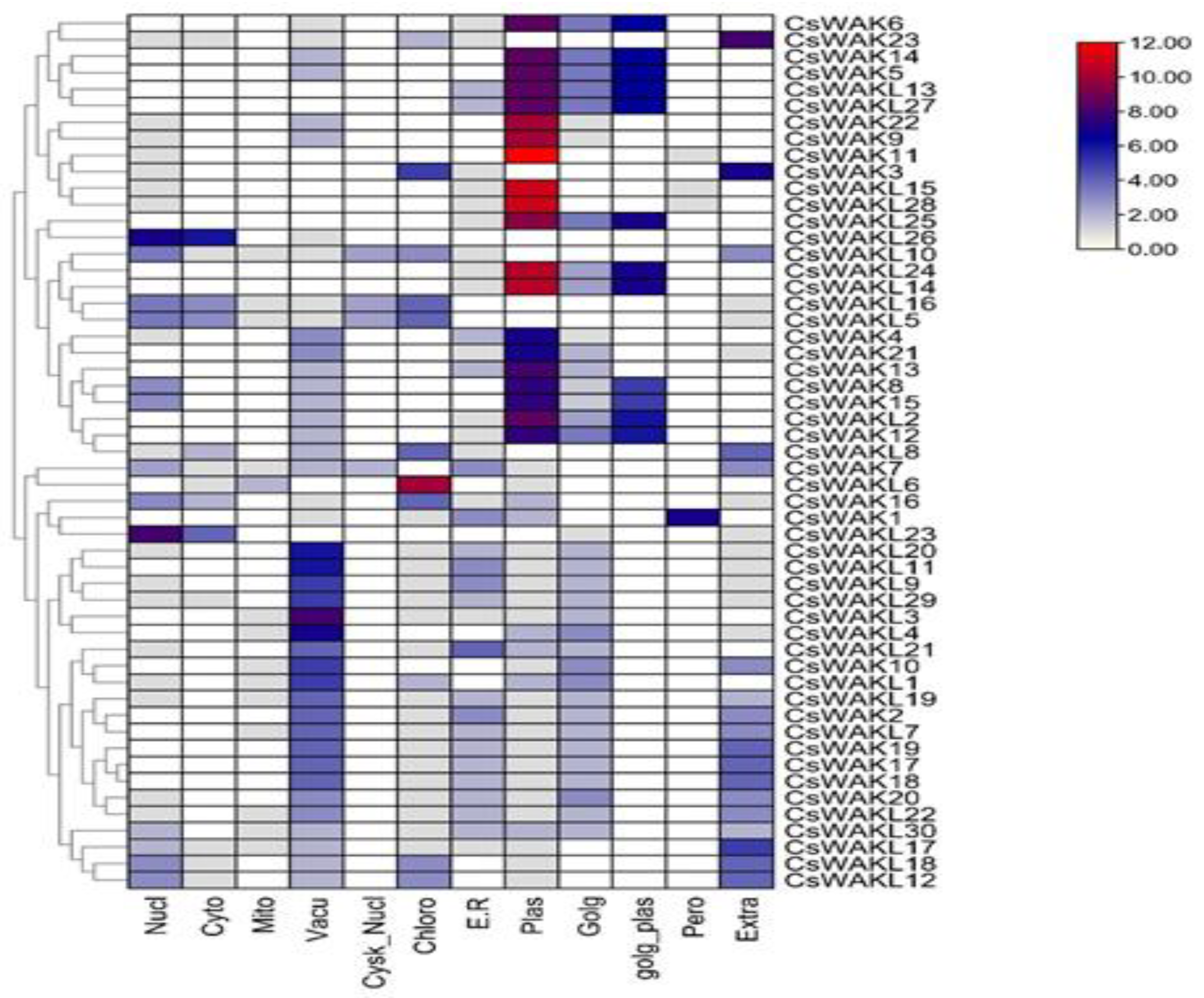
2.2. Physiochemical Property and Subcellular Localization Analysis of CsWAKs/CsWAKLs Proteins
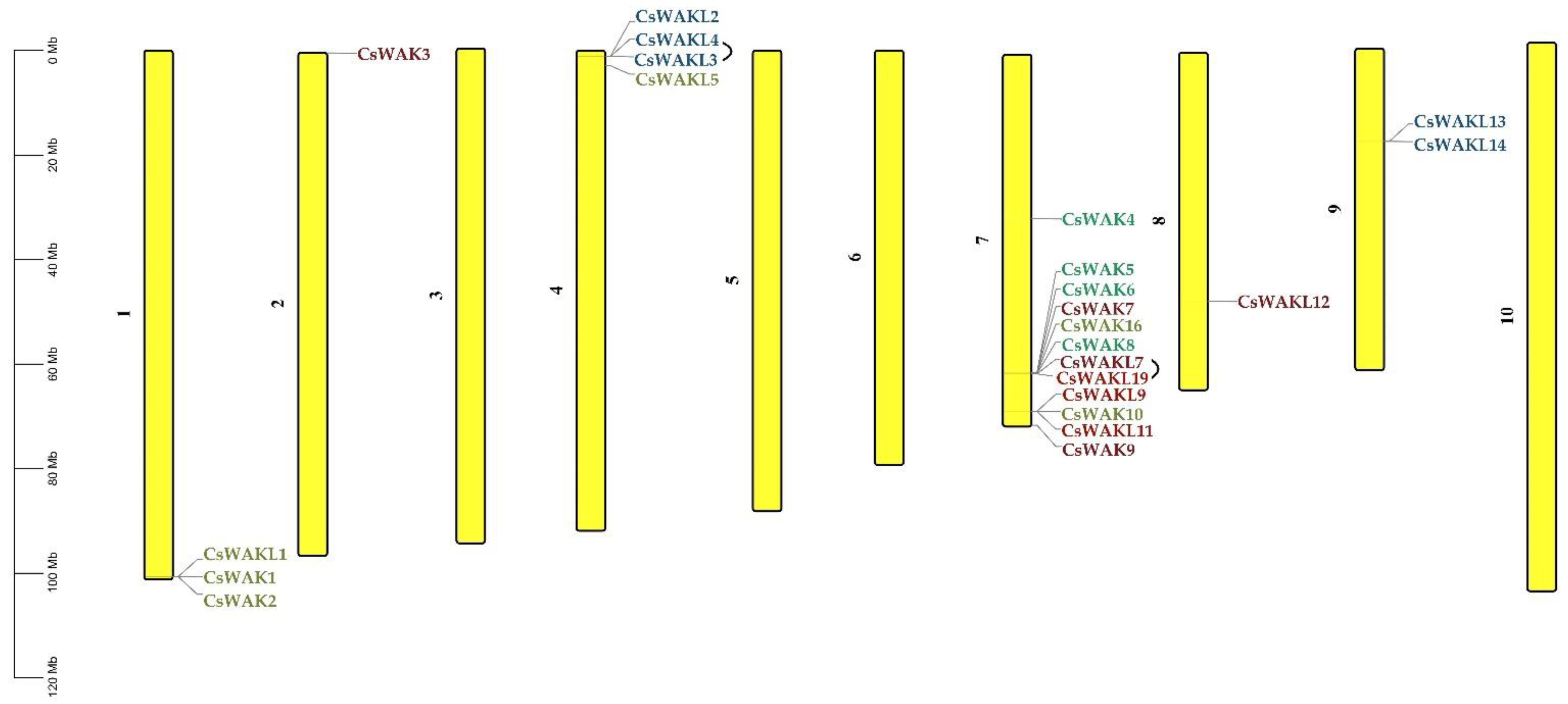
2.3. Chromosomal Location, Synteny, and Selective Pressure of CsWAK/CsWAKL Gene Family
2.4. Conserved Motif and Gene Structure of CsWAKs/CsWAKLs
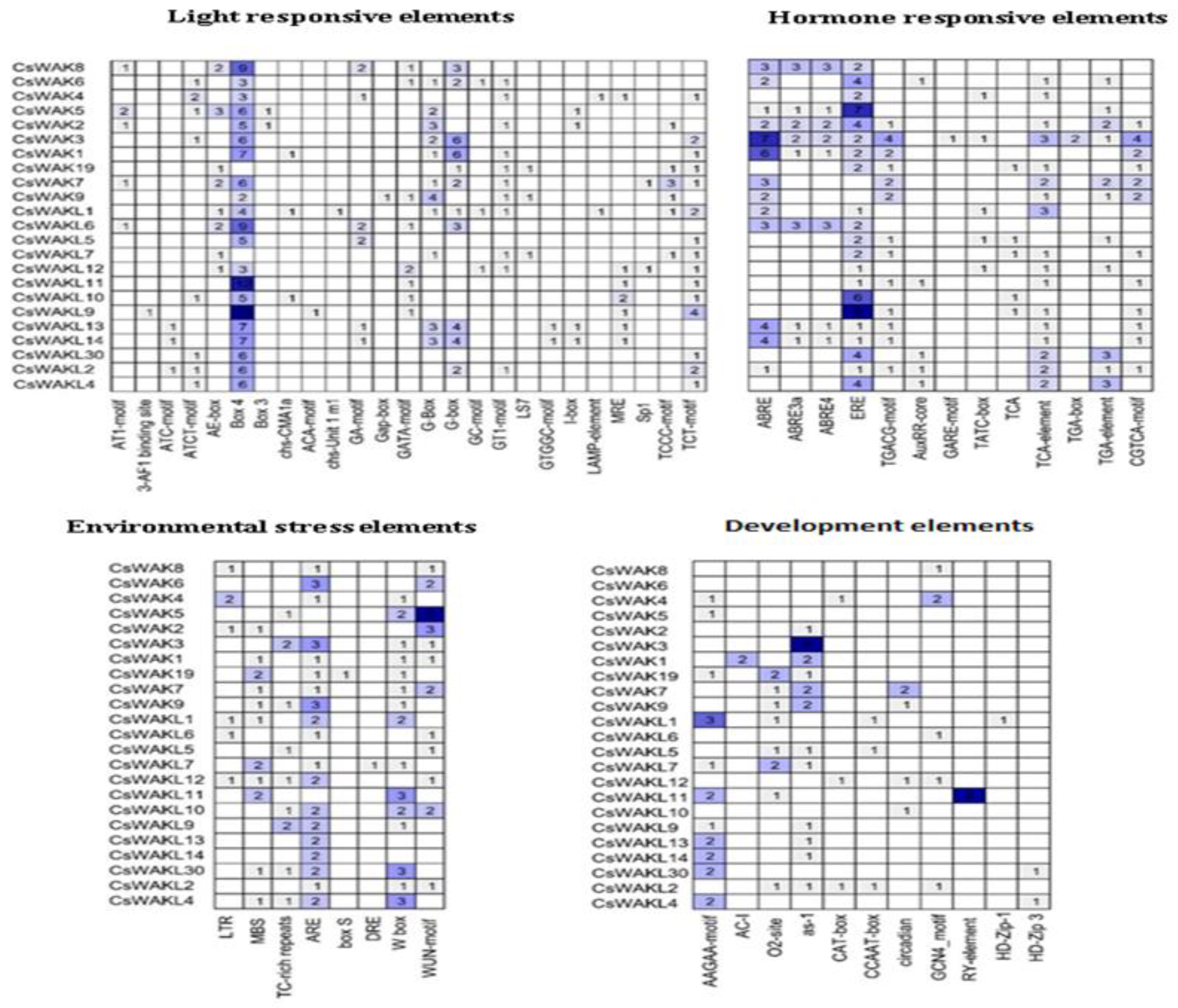
2.5. Cis-Acting Elements Analysis of Promoters of CsWAK/CsWAKL Genes
2.6. Expression Patterns of the Some CsWAKs/CsWAKLs Genes in Different Tissues
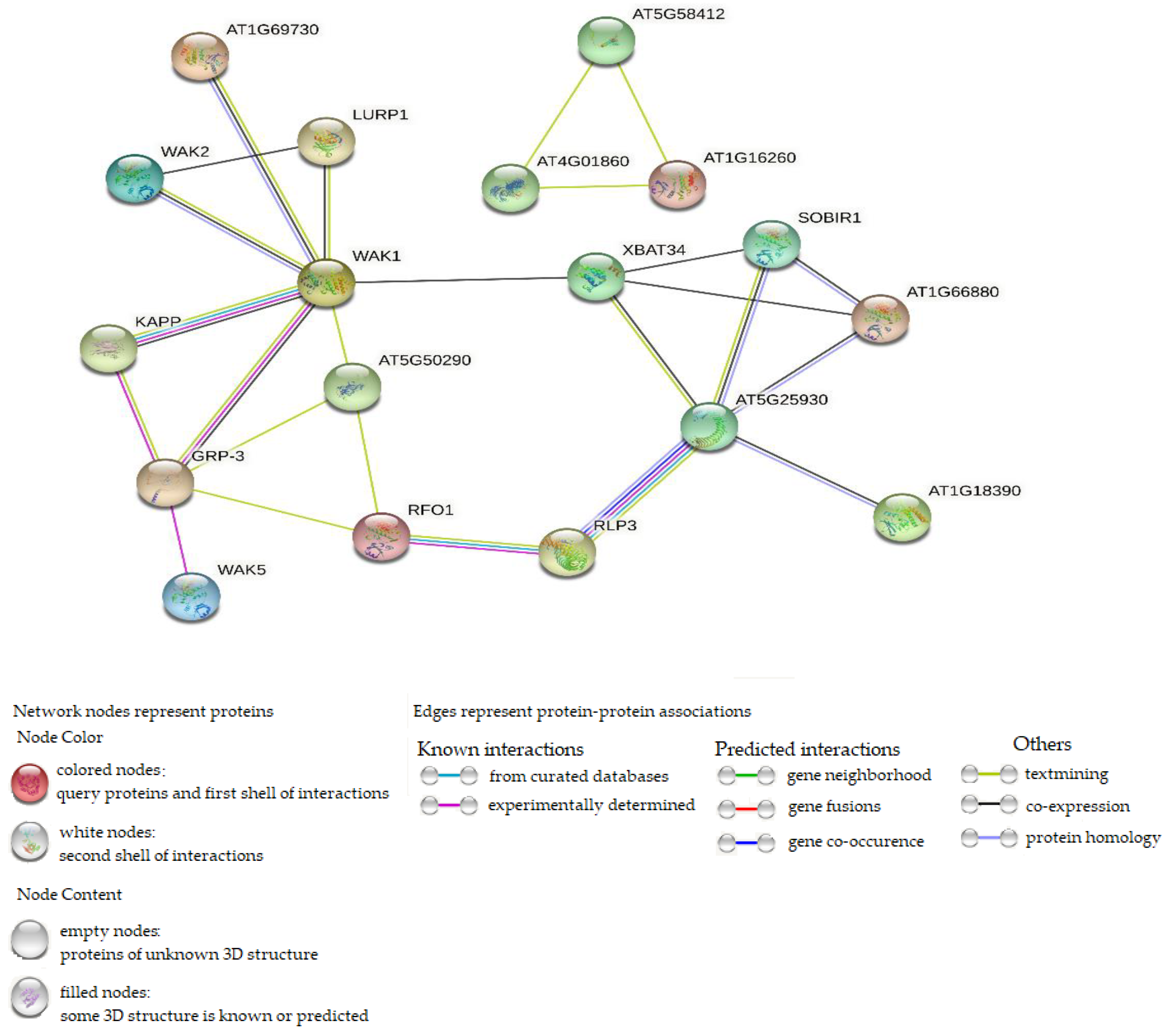
2.7. Identification of Protein–Protein Interaction and miRNA Targets
3. Discussion
4. Materials and Methods
4.1. Identification of WAK/WAKL Genes
4.2. Physicochemical Properties, Subcellular Prediction, and Conserved Motif Analysis of WAKs/WAKLs
4.3. Phylogenetic Tree Construction and Gene Structure Analysis Identification of WAK/WAKL Genes
4.4. Chromosome Localization, Synteny Analysis, and Ka/Ks Calculation
4.5. Prediction of Cis-Acting Elements
4.6. Plant Material
4.7. Total RNA Isolation, cDNA Synthesis, and Gene Expression Analysis
4.8. Isolation of Cannabis Trichomes
4.9. Identification of Protein-Protein interactions and miRNA Target Sites
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pilling, E.; Höfte, H. Feedback from the wall. Curr. Opin. Plant Biol. 2003, 6, 611–616. [Google Scholar] [CrossRef]
- Wolf, S.; Hofte, H. Growth control: A saga of cell walls, ros, and peptide receptors. Plant Cell 2014, 26, 1848–1856. [Google Scholar] [CrossRef]
- Ferrari, S.; Savatin, D.V.; Sicilia, F.; Gramegna, G.; Cervone, F.; Lorenzo, G.D. Oligogalacturonides: Plant damage-associated molecular patterns and regulators of growth and development. Front. Plant Sci. 2013, 4, 49. [Google Scholar] [CrossRef]
- Kohorn, B.D. Cell wall-associated kinases and pectin perception. J. Exp. Bot. 2016, 67, 489–494. [Google Scholar] [CrossRef]
- Verica, J.A.; He, Z.H. The cell wall-associated kinase (WAK) and WAK-like kinase gene family. Plant Physiol. 2002, 129, 455–459. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, C.; Li, L.; Meng, L.; Singh, J.; Jiang, N.; Deng, X.W.; He, Z.H.; Lemaux, P.G. Evolutionary expansion, gene structure, and expression of the rice wall-associated kinase gene family. Plant Physiol. 2005, 139, 1107–1124. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Bacic, A.; Johnson, K.L.; Humphries, J. The role of Brachypodium distachyon Wall-Associated Kinases (WAKs) in cell expansion and stress responses. Cells 2020, 9, 2478. [Google Scholar] [CrossRef]
- Sun, Z.; Song, Y.; Chen, D.; Zhang, Y.; Zhang, Q.; Yi, Y.; Qu, G. Genome-wide identification, classification, characterization, and expression analysis of the wall-associated kinase family during fruit development and under wound stress in tomato (Solanum lycopersicum L.). Genes 2020, 11, 1186. [Google Scholar] [CrossRef]
- Zhang, Z.; Ma, W.; Ren, Z.; Wang, X.; Zhao, J.; Pei, X.; Liu, Y.; He, K.; Zhang, F.; Hou, W.; et al. Characterization and expression analysis of wall-associated kinase (WAK) and WAK-like family in cotton. Int. J. Biol. Macromol. 2021, 187, 867–879. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, Z.; Tian, Y.; Zhang, S.; Li, D.; Dong, W.; Zhang, C.; Zhang, Z. Characterization of wall-associated kinase/wallassociated kinase-like (WAK/WAKL) family in rose (Rosa chinensis) reveals the role of RcWAK4 in Botrytis resistance. BMC Plant Biol. 2021, 21, 526. [Google Scholar] [CrossRef]
- Tripathi, R.K.; Aguirre, J.A.; Singh, J. Genome-wide analysis of wall associated kinase (WAK) gene family in barley. Genomics 2021, 113, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Ma, J.; Liu, H.; Ou, M.; Ye, H.; Zhao, P. Identification and Characterization of Wall-Associated Kinase (WAK) and WAK-like (WAKL) Gene Family in Juglans regia and Its Wild Related Species Juglans mandshurica. Genes 2022, 13, 134. [Google Scholar] [CrossRef] [PubMed]
- Tocquard, K.; Lafon-Placette, C.; Auguin, D.; Muries, B.; Bronner, G.; Lopez, D.; Venisse, J.S. In silico study of wall-associated kinase family reveals large-scale genomic expansion potentially connected with functional diversification in Populus. Tree Genet. Genomes 2014, 10, 1135–1147. [Google Scholar] [CrossRef]
- Zuo, W.; Chao, Q.; Zhang, N.; Ye, J.; Tan, G.; Li, B.; Xing, Y.; Zhang, B.; Liu, H.; Fengler, K. A Maize wall-associated kinase confers quantitative resistance to head smut. Nat. Genet. 2015, 47, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Panchy, N.; Lehti-Shiu, M.; Shiu, S.H. Evolution of Gene Duplication in Plants. Plant Physiol. 2016, 171, 2294–2316. [Google Scholar] [CrossRef]
- Rosli, H.G.; Zheng, Y.; Pombo, M.A.; Zhong, S.; Bombarely, A.; Fei, Z.; Collmer, A.; Martin, G.B. Transcriptomics-based screen for genes induced by flagellin and repressed by pathogen effectors identifies a cell wall-associated kinase involved in plant immunity. Genome Biol. 2013, 14, R139. [Google Scholar] [CrossRef]
- Saintenac, C.; Lee, W.S.; Cambon, F.; Rudd, J.J.; King, R.C.; Marande, W.; Powers, S.J.; Berges, H.; Phillips, A.L.; Uauy, C.; et al. Wheat receptor-kinase-like protein Stb6 controls gene-for-gene resistance to fungal pathogen Zymoseptoria tritici. Nat. Genet. 2018, 50, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Praz, C.; Li, B.; Singla, J.; Robert, C.A.; Kessel, B.; Keller, B. Fungal resistance mediated by maize wall-associated kinase Zm WAK-RLK 1 correlates with reduced benzoxazinoid content. New Phytol. 2019, 221, 976–987. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Lv, Y.; Lei, W.; Li, X.; Chen, Y.; Zheng, L.; Xia, Y.; Shen, Z. Cloning and characterization of the Oryza sativa wall-associated kinase gene OsWAK11 and its transcriptional response to abiotic stresses. Plant Soil 2014, 384, 335–346. [Google Scholar] [CrossRef]
- Decreux, A.; Johan, M. Wall-associated kinase WAK1 interacts with cell wall pectins in a calcium-induced conformation. Plant Cell Physiol. 2005, 46, 268–278. [Google Scholar] [CrossRef]
- Hou, X.; Tong, H.; Selby, J.; DeWitt, J.; Peng, X.; He, Z.H. Involvement of a cell wall-associated kinase, WAKL4, in Arabidopsis mineral responses. Plant Physiol. 2005, 139, 1704–1716. [Google Scholar] [CrossRef] [PubMed]
- He, Z.H.; Cheeseman, I.; He, D.; Kohorn, B.D. A cluster of five cell wall-associated receptor kinase genes, Wak1–5, are expressed in specific organs of Arabidopsis. Plant Mol. Biol. 1999, 39, 1189–1196. [Google Scholar] [CrossRef]
- Wagner, T.A.; Kohorn, B.D. Wall-associated kinases are expressed throughout plant development and are required for cell expansion. Plant Cell 2001, 13, 303–318. [Google Scholar] [CrossRef] [PubMed]
- Sivaguru, M.; Ezaki, B.; He, Z.H.; Tong, H.; Matsumoto, H. Aluminium-induced gene expression and protein localization of a cell wall-associated receptor kinase in Arabidopsis. Plant Physiol. 2003, 132, 2256–2266. [Google Scholar] [CrossRef] [PubMed]
- Lally, D.; Ingmire, P.; Tong, H.Y.; He, Z.H. Antisense expression of a cell wall-associated protein kinase, WAK4, inhibits cell elongation and alters morphology. Plant Cell 2001, 13, 1317–1331. [Google Scholar] [CrossRef]
- Kohorn, B.D.; Kobayashi, M.; Johansen, S.; Riese, J.; Huang, L.F.; Koch, K.; Fu, S.; Dotson, A.; Byers, N. An Arabidopsis cell wall-associated kinase required for invertase activity and cell growth. Plant J. 2006, 46, 307–316. [Google Scholar] [CrossRef]
- Kanneganti, V.; Gupta, A.K. RNAi mediated silencing of a wall associated kinase, OsiWAK1 in Oryza sativa results in impaired root development and sterility due to anther indehiscence. Physiol. Mol. Biol. Plants 2011, 17, 65–77. [Google Scholar] [CrossRef]
- Wang, N.; Huang, H.J.; Ren, S.T.; Li, J.J.; Sun, Y.; Sun, D.Y. The rice wall-associated receptor-like kinase gene OsDEES1 plays a role in female gametophyte development. Plant Physiol. 2012, 160, 696–707. [Google Scholar] [CrossRef] [PubMed]
- He, Z.H.; He, D.; Kohorn, B.D. Requirement for the induced expression of a cell wall associated receptor kinase for survival during the pathogen response. Plant J. 1998, 14, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Diener, A.C.; Ausubel, F.M. Resistance to Fusarium oxysporum 1, a dominant Arabidopsis disease-resistance gene, is not race specific. Genetics 2005, 171, 305–321. [Google Scholar] [CrossRef]
- Brutus, A.; Sicilia, F.; Macone, A.; Cervone, F.; De Lorenzo, G. A domain swap approach reveals a role of the plant wall-associated kinase 1 (WAK1) as a receptor of oligogalacturonides. Proc. Natl. Acad. Sci. USA 2010, 107, 9452–9457. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhou, S.Y.; Zhao, W.S.; Su, S.C.; Peng, Y.L. A novel wall-associated receptor-like protein kinase gene, OsWAK1, plays important roles in rice blast disease resistance. Plant Mol. Biol. 2009, 69, 337–346. [Google Scholar] [CrossRef]
- Hurni, S.; Scheuermann, D.; Krattinger, S.G.; Kessel, B.; Wicker, T.; Herren, G.; Mirjam, N.; Breen, J.; Presterl, T.; Ouzunova, M.; et al. The maize disease resistance gene Htn1 against northern corn leaf blight encodes a wall-associated receptor-like kinase. Proc. Natl. Acad. Sci. USA 2015, 112, 8780–8785. [Google Scholar] [CrossRef]
- Fetch, T.G., Jr.; Steffenson, B.J. Identification of Cochliobolus sativus isolates expressing differential virulence on two-row barley genotypes from North Dakota. Can. J. Plant Pathol. 1994, 16, 202–206. [Google Scholar] [CrossRef]
- Ameen, G.; Solanki, S.; Drader, T.; Sager-Bittara, L.; Steffenson, B.; Kleinhofs, A.; Vogiatzis, C.; Brueggeman, R.S. rcs5-mediated spot blotch resistance in barley is conferred by wall-associated kinases that resist pathogen manipulation. BioRxiv 2020. [CrossRef]
- Fidan, M.; Karaismailoğlu, M.C. Kenevirin Tarihçesi ve Sistematiği. Kenevir (Cannabis sativa L.); Onay, A., Yıldırım, H., Ekinci, R., Eds.; Palme Yayınevi: Istanbul, Turkey, 2020; pp. 15–31. [Google Scholar]
- Johnson, M.S.; Wallace, J.G. Genomic and chemical diversity of commercially available high-CBD industrial hemp accessions. Front. Genet. 2021, 12, 1160. [Google Scholar] [CrossRef]
- Ahmad, R.; Tehsin, Z.; Malik, S.T.; Asad, S.A.; Shahzad, M.; Bilal, M.; Khan, S.A. Phytoremediation potential of hemp (Cannabis sativa L.): Identification and characterization of heavy metals responsive genes. Clean–Soil Air Water 2016, 44, 195–201. [Google Scholar] [CrossRef]
- Horton, P.; Park, K.J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef]
- Yadav, C.B.; Bonthala, V.S.; Muthamilarasan, M.; Pandey, G.; Khan, Y.; Prasad, M. Genome-wide development of transposable elements-based markers in foxtail millet and construction of an integrated database. DNA Res. 2015, 22, 79–90. [Google Scholar] [CrossRef]
- Dou, L.; Li, Z.; Shen, Q.; Shi, H.; Li, H.; Wang, W.; Zou, C.; Shang, H.; Li, H.; Xiao, G. Genome-wide characterization of the WAK gene family and expression analysis under plant hormone treatment in cotton. BMC Genom. 2021, 22, 85. [Google Scholar] [CrossRef]
- Yang, J.; Xie, M.; Wang, X.; Wang, G.; Zhang, Y.; Li, Z.; Ma, Z. Identification of cell wall-associated kinases as important regulators involved in Gossypium hirsutum resistance to Verticillium dahliae. BMC Plant Biol. 2021, 21, 220. [Google Scholar] [CrossRef]
- Das, A.; Chaudhury, S.; Kalita, M.C.; Mondal, T.K. In silico identification, characterization and expression analysis of miRNAs in Cannabis sativa L. Plant Gene 2015, 2, 17–24. [Google Scholar] [CrossRef]
- Hasan, M.A.; Hussain, M.H.; Chowdhury, A.S.; Dhar, S.B.; Abedin, M.; Fima, I.N. Computational identification of potential microRNAs and their targets from expressed sequence tags of marijuana (Cannabis sativa). Meta Gene 2016, 10, 45–55. [Google Scholar] [CrossRef]
- Van Bakel, H.; Stout, J.M.; Cote, A.G.; Tallon, C.M.; Sharpe, A.G.; Hughes, T.R.; Page, J.E. The draft genome and transcriptome of Cannabis sativa. Genome Biol. 2011, 12, R102. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Wang, B.; Xie, S.; Xu, X.; Zhang, J.; Pei, L.; Yu, Y.; Yang, W.; Zhang, Y. A high-quality reference genome of wild Cannabis sativa. Hortic. Res. 2020, 7, 73. [Google Scholar] [CrossRef]
- Ponzoni, E.; Brambilla, I.M.; Galasso, I. Genome-wide identification and organization of seed storage protein genes of Cannabis sativa. Biol. Plant 2018, 62, 693–702. [Google Scholar] [CrossRef]
- Allen, K.D.; McKernan, K.; Pauli, C.; Roe, J.; Torres, A.; Gaudino, R. Genomic characterization of the complete terpene synthase gene family from Cannabis sativa. PLoS ONE 2019, 14, e0222363. [Google Scholar] [CrossRef]
- Guerriero, G.; Deshmukh, R.; Sonah, H.; Sergeant, K.; Hausman, J.F.; Lentzen, E.; Valle, N.; Siddiqui, K.S.; Exley, C. Identification of the aquaporin gene family in Cannabis sativa and evidence for the accumulation of silicon in its tissues. Plant Sci. 2019, 287, 110167. [Google Scholar] [CrossRef]
- Pan, G.; Li, Z.; Yin, M.; Huang, S.; Tao, J.; Chen, A.; Li, J.; Tang, H.; Chang, L.; Deng, Y.; et al. Genome-wide identification, expression, and sequence analysis of CONSTANS-like gene family in cannabis reveals a potential role in plant flowering time regulation. BMC Plant Biol. 2021, 21, 142. [Google Scholar] [CrossRef]
- Wei, H.; Chen, S.; Niyitanga, S.; Liu, T.; Qi, J.; Zhang, L. Genome-wide identification and expression analysis response to GA3 stresses of WRKY gene family in seed hemp (Cannabis sativa L.). Gene 2022, 822, 146290. [Google Scholar] [CrossRef]
- Ma, G.; Zelman, A.K.; Apicella, P.V.; Berkowitz, G. Genome-Wide Identification and Expression Analysis of Homeodomain Leucine Zipper Subfamily IV (HD-ZIP IV) Gene Family in Cannabis sativa L. Plants 2022, 11, 1307. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Dai, Z.G.; Zhang, X.Y.; Tang, Q.; Cheng, C.H.; Liu, C.; Yu, Y.; Xu, G.C.; Xie, D.W.; Su, J.G. Identification of TPS and TPP gene families in Cannabis sativa and their expression under abiotic stresses. Biol. Plant 2022, 66, 14–23. [Google Scholar] [CrossRef]
- Lu, M.; Meng, X.X.; Zhang, Y.M.; Zhu, X.W.; Li, J.; Chen, W.Q.; Wang, H.H.; Cao, X.; Sun, W.; Mi, Y.L.; et al. Genome-Wide Identification and Expression Profiles of bZIP Genes in Cannabis sativa L. Cannabis Cannabinoid Res. 2022. [CrossRef] [PubMed]
- Wan, J.; He, M.; Hou, Q.; Zou, L.; Yang, Y.; Wei, Y.; Chen, X. Cell wall associated immunity in plants. Stress Biol. 2021, 1, 3. [Google Scholar] [CrossRef]
- de Oliveira, L.F.V.; Christoff, A.P.; de Lima, J.C.; de Ross, B.C.F.; Sachetto-Martins, G.; Margis-Pinheiro, M.; Margis, R. The Wall-associated Kinase gene family in rice genomes. Plant Sci. 2014, 229, 181–192. [Google Scholar] [CrossRef]
- Zuo, C.; Liu, Y.; Guo, Z.; Mao, J.; Chu, M.; Chen, B. Genome-wide annotation and expression responses to biotic stresses of the WALL-ASSOCIATED KINASE-RECEPTOR-LIKE KINASE (WAK-RLK) gene family in Apple (Malus domestica). Eur. J. Plant Pathol. 2019, 153, 771–785. [Google Scholar] [CrossRef]
- White, S.E.; Habera, L.F.; Wessler, S.R. Retrotransposons in the flanking regions of normal plant genes: A role for copia-like elements in the evolution of gene structure and expression. Proc. Natl. Acad. Sci. USA 1994, 91, 11792–11796. [Google Scholar] [CrossRef]
- Kumar, A.; Bennetzen, J.L. Plant retrotransposons. Annu. Rev. Genet. 1999, 33, 479–532. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, W.; Kang, Y.; Fan, Y.; Yang, X.; Shi, M.; Zhang, R.; Wang, Y.; Qin, S. Genome-wide identification and expression analysis of wall-associated kinase (WAK) gene family in potato (Solanum tuberosum L.). Plant Biotechnol. Rep. 2022, 16, 317–331. [Google Scholar] [CrossRef]
- Wang, W.; Barnaby, J.Y.; Tada, Y.; Li, H.; Tör, M.; Caldelari, D.; Lee, D.; Fu, X.D.; Dong, X. Timing of plant immune responses by a central circadian regulator. Nature 2011, 470, 110–114. [Google Scholar] [CrossRef]
- VanGuilder, H.D.; Vrana, K.E.; Freeman, W.M. Twenty-five years of quantitative PCR for gene expression analysis. Biotechniques 2008, 44, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Zhang, X.; Li, Y.; Ridout, K.; Serrano-Serrano, M.L.; Yang, Y.; Ravikanth, G.; Nawaz, M.A.; Mumtaz, A.S.; Salamin, N.; et al. Large-scale whole-genome resequencing unravels the domestication history of Cannabis sativa. Sci. Adv. 2021, 7, eabg2286. [Google Scholar] [CrossRef] [PubMed]
- Tanney, C.A.; Backer, R.; Geitmann, A.; Smith, D.L. Cannabis glandular trichomes: A cellular metabolite factory. Front. Plant Sci. 2021, 12, 721986. [Google Scholar] [CrossRef]
- Park, A.R.; Cho, S.K.; Yun, U.J.; Jin, M.Y.; Lee, S.H.; Sachetto-Martins, G. Interaction of the Arabidopsis receptor protein kinase Wak1 with a glycine-rich protein, AtGRP-3. J. Biol. Chem. 2001, 276, 26688–26693. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.M.; Wagner, T.A.; Perret, M.; He, Z.H.; He, D.; Kohorn, B.D. WAKs: Cell wall-associated kinases linking the cytoplasm to the extracellular matrix. In Plant Cell Walls; Springer: Dordrecht, The Netherlands, 2001; pp. 197–206. [Google Scholar]
- Narjala, A.; Nair, A.; Tirumalai, V.; Hari Sundar, G.V.; Shivaprasad, P.V. A conserved sequence signature is essential for robust plant miRNA biogenesis. Nucleic Acids Res. 2020, 48, 3103–3118. [Google Scholar] [CrossRef]
- Sunkar, R.; Zhu, J.K. Novel and stress-regulated microRNAs and other small RNAs from Arabidopsis. Plant Cell 2004, 16, 2001–2019. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein analysis tools on the ExPASy server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME Suite: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Guo, H.; Zhang, Q.; Guo, M.; Xu, Y.; Zeng, M.; Lv, P.; Chen, X.; Yang, M. Evaluation of reference genes for RT-qPCR analysis in wild and cultivated Cannabis. Biosci. Biotechnol. Biochem. 2018, 82, 1902–1910. [Google Scholar] [CrossRef]
- Marshall, O.J. PerlPrimer: Cross-platform, graphical primer design for standard, bisulphite and real-time PCR. Bioinformatics 2004, 20, 2471–2472. [Google Scholar] [CrossRef]
- Livingston, S.J.; Quilichini, T.D.; Booth, J.K.; Wong, D.; Rensing, K.H.; Laflamme-Yonkman, J.; Castellarin, S.D.; Bohlmann, J.; Page, J.E.; Samuels, A.L. Cannabis glandular trichomes alter morphology and metabolite content during flower maturation. Plant J. 2020, 101, 37–56. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Zhuang, Z.; Zhao, P.X. PsRNATarget: A plant small RNA target analysis server. Nucleic Acids Res. 2018, 46, W49–W54. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein ID | Gene Name | Length (aa) | Molecular Weight (kDa) | pI | Grand Average of Hydropathicity (GRAVY) | Aliphatic Index | Instability Index |
|---|---|---|---|---|---|---|---|
| XP_030487710.1 | CsWAK1 | 983 | 108.78 | 6.45 | −0.109 | 94.67 | 34.45 |
| XP_030492016.1 | CsWAK2 | 761 | 84.63 | 6.55 | −0.125 | 91.94 | 34.58 |
| XP_030503825.1 | CsWAK3 | 784 | 87.58 | 5.29 | −0.302 | 80.54 | 41.03 |
| XP_030480968.1 | CsWAK4 | 747 | 83.83 | 6.25 | −0.210 | 89.63 | 36.68 |
| XP_030481386.1 | CsWAK5 | 767 | 85.73 | 6.74 | −0.234 | 86.53 | 31.08 |
| XP_030479582.1 | CsWAK6 | 764 | 85.62 | 8.09 | −0.250 | 86.24 | 33.42 |
| XP_030479448.1 | CsWAK7 | 749 | 84.33 | 5.65 | −0.332 | 79.63 | 38.18 |
| XP_030481338.1 | CsWAK8 | 750 | 84.58 | 5.64 | −0.236 | 78.61 | 26.68 |
| XP_030479562.1 | CsWAK9 | 726 | 80.08 | 5.67 | −0.059 | 90.61 | 35.50 |
| KAF4346698.1 | CsWAK10 | 766 | 84.17 | 5.08 | −0.203 | 88.15 | 36.29 |
| KAF4398384.1 | CsWAK11 | 788 | 88.67 | 8.33 | −0.264 | 85.34 | 35.12 |
| KAF4376937.1 | CsWAK12 | 784 | 87.67 | 7.79 | −0.247 | 86.52 | 31.11 |
| KAF4391022.1 | CsWAK13 | 747 | 83.82 | 8.96 | −0.212 | 88.84 | 37.39 |
| KAF4376938.1 | CsWAK14 | 747 | 83.69 | 6.46 | −0.264 | 84.67 | 31.97 |
| KAF4376931.1 | CsWAK15 | 751 | 84.65 | 5.65 | −0.226 | 79.55 | 26.74 |
| KAF4346705.1 | CsWAK16 | 769 | 85.46 | 6.26 | −0.221 | 89.45 | 35.87 |
| KAF4379309.1 | CsWAK17 | 771 | 86.44 | 5.52 | −0.154 | 84.03 | 33.17 |
| KAF4376930.1 | CsWAK18 | 771 | 86.39 | 5.58 | −0.159 | 84.03 | 33.75 |
| XP_030480237.1 | CsWAK19 | 771 | 86.36 | 5.52 | −0.162 | 83.90 | 32.68 |
| KAF4353681.1 | CsWAK20 | 711 | 78.60 | 6.06 | −0.091 | 91.81 | 31.83 |
| KAF4398399.1 | CsWAK21 | 748 | 83.54 | 5.08 | −0.153 | 82.47 | 26.33 |
| KAF4377370.1 | CsWAK22 | 725 | 79.97 | 5.81 | −0.057 | 90.59 | 35.02 |
| KAF4346699.1 | CsWAK23 | 695 | 76.92 | 5.73 | −0.112 | 90.27 | 33.03 |
| XP_030492015.1 | CsWAKL1 | 769 | 84.33 | 5.15 | −0.179 | 89.08 | 34.43 |
| XP_030496407.1 | CsWAKL2 | 646 | 72.77 | 6.67 | −0.274 | 87.51 | 45.51 |
| XP_030496276.1 | CsWAKL3 | 682 | 76.40 | 5.55 | −0.200 | 87.04 | 42.61 |
| XP_030496277.1 | CsWAKL4 | 656 | 74.00 | 6.92 | −0.295 | 83.95 | 42.72 |
| XP_030499458.1 | CsWAKL5 | 741 | 83.02 | 6.26 | −0.266 | 87.44 | 33.63 |
| XP_030481339.1 | CsWAKL6 | 594 | 67.26 | 5.92 | −0.295 | 80.89 | 26.93 |
| XP_030480238.1 | CsWAKL7 | 704 | 78.95 | 5.62 | −0.163 | 85.53 | 32.25 |
| KAF4376932.1 | CsWAKL8 | 708 | 80.00 | 6.28 | −0.284 | 82.87 | 34.43 |
| XP_030479527.1 | CsWAKL9 | 787 | 87.47 | 7.63 | −0.320 | 80.97 | 35.08 |
| XP_030480670.1 | CsWAKL10 | 763 | 84.55 | 8.27 | −0.292 | 81.73 | 34.78 |
| XP_030480956.1 | CsWAKL11 | 769 | 85.81 | 6.63 | −0.356 | 82.46 | 30.83 |
| XP_030484049.1 | CsWAKL12 | 732 | 81.76 | 6.90 | −0.164 | 87.84 | 37.74 |
| XP_030508821.1 | CsWAKL13 | 692 | 77.11 | 5.28 | −0.319 | 79.31 | 36.59 |
| XP_030508820.1 | CsWAKL14 | 694 | 76.93 | 5.58 | −0.269 | 81.59 | 36.30 |
| KAF4379299.1 | CsWAKL15 | 761 | 85.76 | 7.42 | −0.259 | 86.32 | 35.33 |
| KAF4355457.1 | CsWAKL16 | 741 | 83.08 | 6.18 | −0.263 | 86.77 | 33.15 |
| KAF4398398.1 | CsWAKL17 | 740 | 83.41 | 5.52 | −0.307 | 81.26 | 38.47 |
| KAF4381619.1 | CsWAKL18 | 735 | 82.07 | 6.65 | −0.165 | 88.54 | 39.53 |
| KAF4365316.1 | CsWAKL19 | 788 | 87.85 | 8.10 | −0.253 | 87.42 | 34.39 |
| KAF4365315.1 | CsWAKL20 | 773 | 85.62 | 7.64 | −0.334 | 82.42 | 32.16 |
| KAF4385963.1 | CsWAKL21 | 771 | 86.07 | 6.51 | −0.357 | 82.24 | 30.78 |
| KAF4385975.1 | CsWAKL22 | 798 | 88.67 | 7.63 | −0.278 | 84.25 | 35.22 |
| KAF4385447.1 | CsWAKL23 | 723 | 80.87 | 5.47 | −0.391 | 77.63 | 37.34 |
| KAF4363501.1 | CsWAKL24 | 794 | 87.89 | 8.16 | −0.111 | 91.56 | 35.43 |
| KAF4346333.1 | CsWAKL25 | 636 | 69.66 | 6.30 | −0.143 | 88.58 | 35.78 |
| KAF4349269.1 | CsWAKL26 | 582 | 65.63 | 5.36 | −0.388 | 84.55 | 38.31 |
| KAF4349872.1 | CsWAKL27 | 692 | 77.01 | 5.28 | −0.318 | 79.31 | 36.59 |
| KAF4379299.1 | CsWAKL28 | 761 | 85.76 | 7.42 | −0.259 | 86.32 | 35.33 |
| KAF4365319.1 | CsWAKL29 | 770 | 85.42 | 6.27 | −0.286 | 85.29 | 32.59 |
| KAF4365318.1 | CsWAKL30 | 791 | 88.21 | 6.25 | −0.271 | 88.31 | 33.10 |
| Gene Pair | Ka | Ks | Ka/Ks | |
|---|---|---|---|---|
| CsWAKL15 | CsWAKL28 | 0 | 0 | NaN |
| CsWAK14 | CsWAK5 | 0.004595 | 0.002025 | 2.268982 |
| CsWAK4 | CsWAK13 | 0.004019 | 0.004055 | 0.991173 |
| CsWAK8 | CsWAK15 | 0.003109 | 0.018028 | 0.172439 |
| CsWAK10 | CsWAKL1 | 0.009062 | 0.025339 | 0.357635 |
| CsWAK1 | CsWAK16 | 0.012437 | 0.042493 | 0.292677 |
| CsWAK2 | CsWAK20 | 0.00731 | 0.029908 | 0.244432 |
| CsWAKL16 | CsWAKL5 | 0.003498 | 0.036624 | 0.095504 |
| CsWAKL13 | CsWAKL27 | 6.31E-04 | 0.006127 | 0.103065 |
| CsWAKL3 | CsWAKL4 | 0.206354 | 0.304622 | 0.67741 |
| CsWAKL24 | CsWAKL25 | 0.025859 | 0.05379 | 0.480727 |
| CsWAK22 | CsWAK9 | 0.003592 | 0.006016 | 0.597084 |
| CsWAKL18 | CsWAKL12 | 0.005622 | 0.03376 | 0.166527 |
| CsWAK3 | CsWAKL23 | 0.01581 | 0.022952 | 0.688858 |
| CsWAK7 | CsWAKL17 | 0.002932 | 0.016705 | 0.175506 |
| CsWAK19 | CsWAKL7 | 0 | 0.002134 | 0 |
| CsWAKL11 | CsWAKL21 | 0 | 0 | NaN |
| Gene Name | Forward qPCR Primer | Reverse qPCR Primer |
|---|---|---|
| CsWAK1 | CAAGCCTCTCAAAGGAAGTATCTC | CATGAAAGAGCCCAATTAAGTCC |
| CsWAK4 | TGCACTTGGTACAACAATTGG | GTTGTTGCAGTATTAACCCACC |
| CsWAK7 | GGTTGCATAGATATTGATGAGTGC | GCACAAGGTTTCATTCTGTATTGG |
| CsWAKL1 | GGATCTAAAGGAGATGGCACC | CCTATAGAGACACCCAAGGC |
| CsWAKL7 | GAAGAACAAGGCAACTTACGGT | CCAAGAGCAGAACCTAAGCA |
| CsUBQ * | TACTGCGCCAGCTAACAAACC | GCACCCGTCTGACCTGAATC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sipahi, H.; Whyte, T.D.; Ma, G.; Berkowitz, G. Genome-Wide Identification and Expression Analysis of Wall-Associated Kinase (WAK) Gene Family in Cannabis sativa L. Plants 2022, 11, 2703. https://doi.org/10.3390/plants11202703
Sipahi H, Whyte TD, Ma G, Berkowitz G. Genome-Wide Identification and Expression Analysis of Wall-Associated Kinase (WAK) Gene Family in Cannabis sativa L. Plants. 2022; 11(20):2703. https://doi.org/10.3390/plants11202703
Chicago/Turabian StyleSipahi, Hülya, Terik Djabeng Whyte, Gang Ma, and Gerald Berkowitz. 2022. "Genome-Wide Identification and Expression Analysis of Wall-Associated Kinase (WAK) Gene Family in Cannabis sativa L." Plants 11, no. 20: 2703. https://doi.org/10.3390/plants11202703
APA StyleSipahi, H., Whyte, T. D., Ma, G., & Berkowitz, G. (2022). Genome-Wide Identification and Expression Analysis of Wall-Associated Kinase (WAK) Gene Family in Cannabis sativa L. Plants, 11(20), 2703. https://doi.org/10.3390/plants11202703