
Microbial Community Structure in Ancient European Arctic Peatlands


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results and Discussion
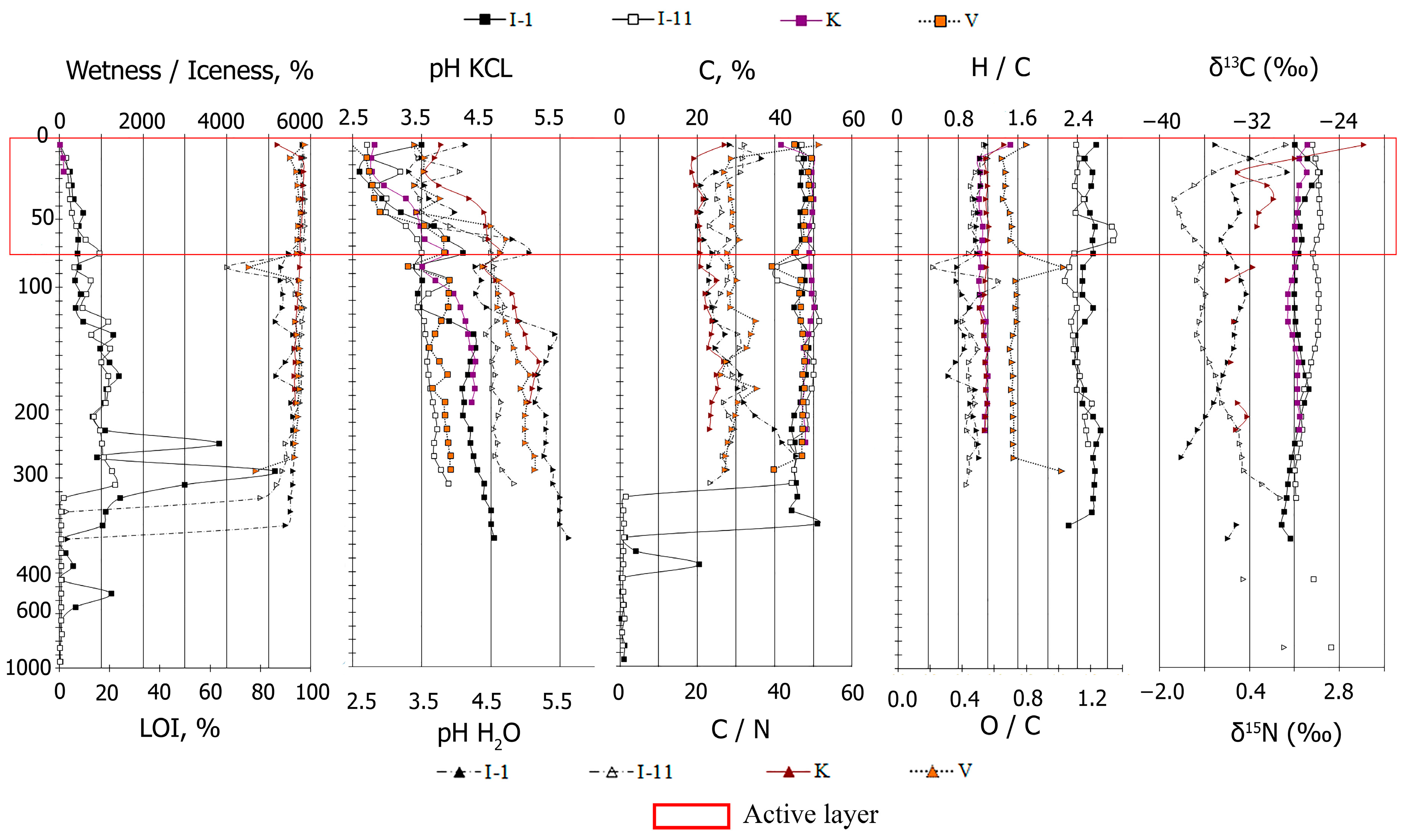
2.1. Soil Properties
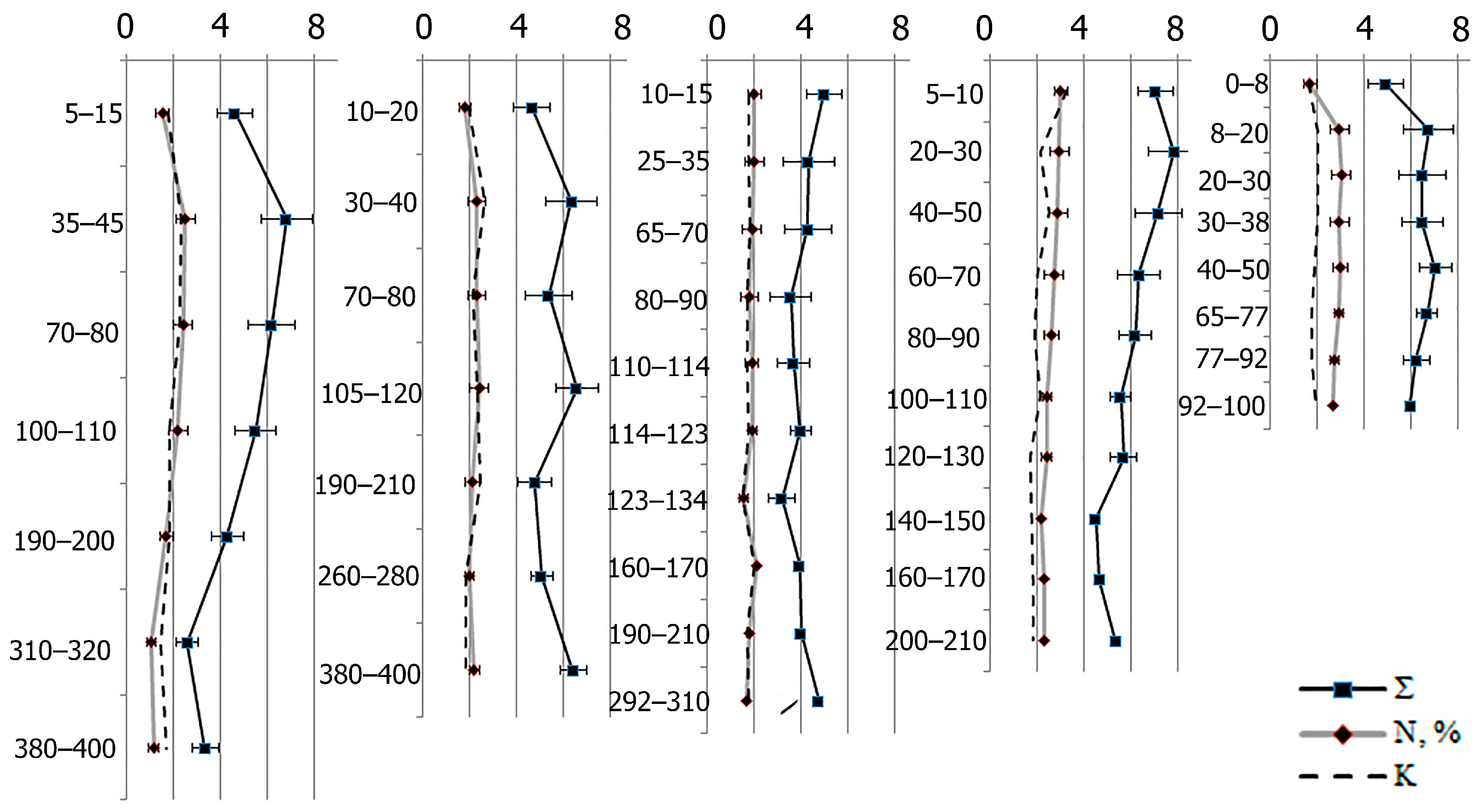
2.2. Amino Acid Composition
2.3. The Microbial Community Diversity and the Distribution of Microbial Groups in Peat Strata
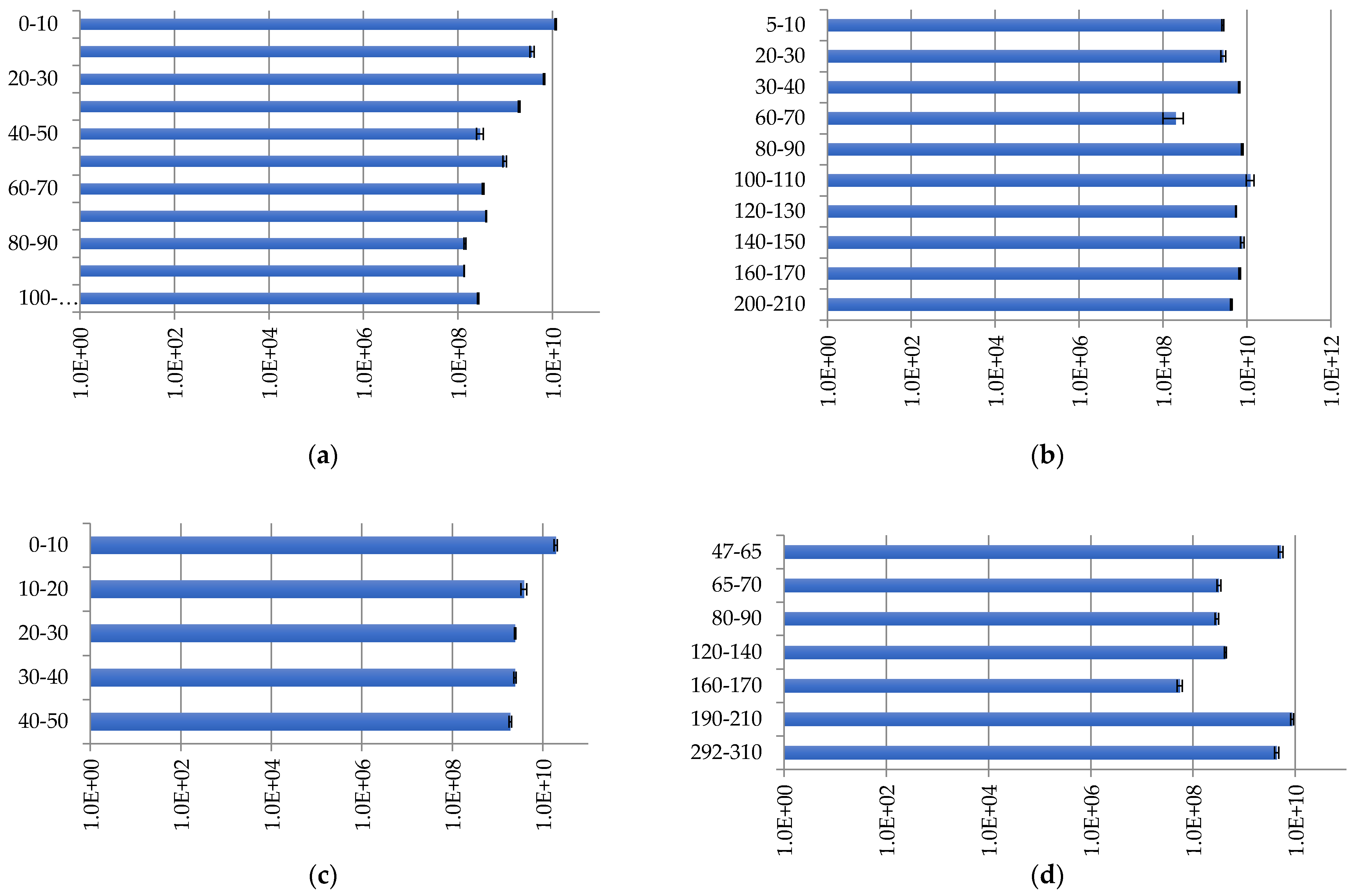
2.3.1. The Bacterial Community Distribution
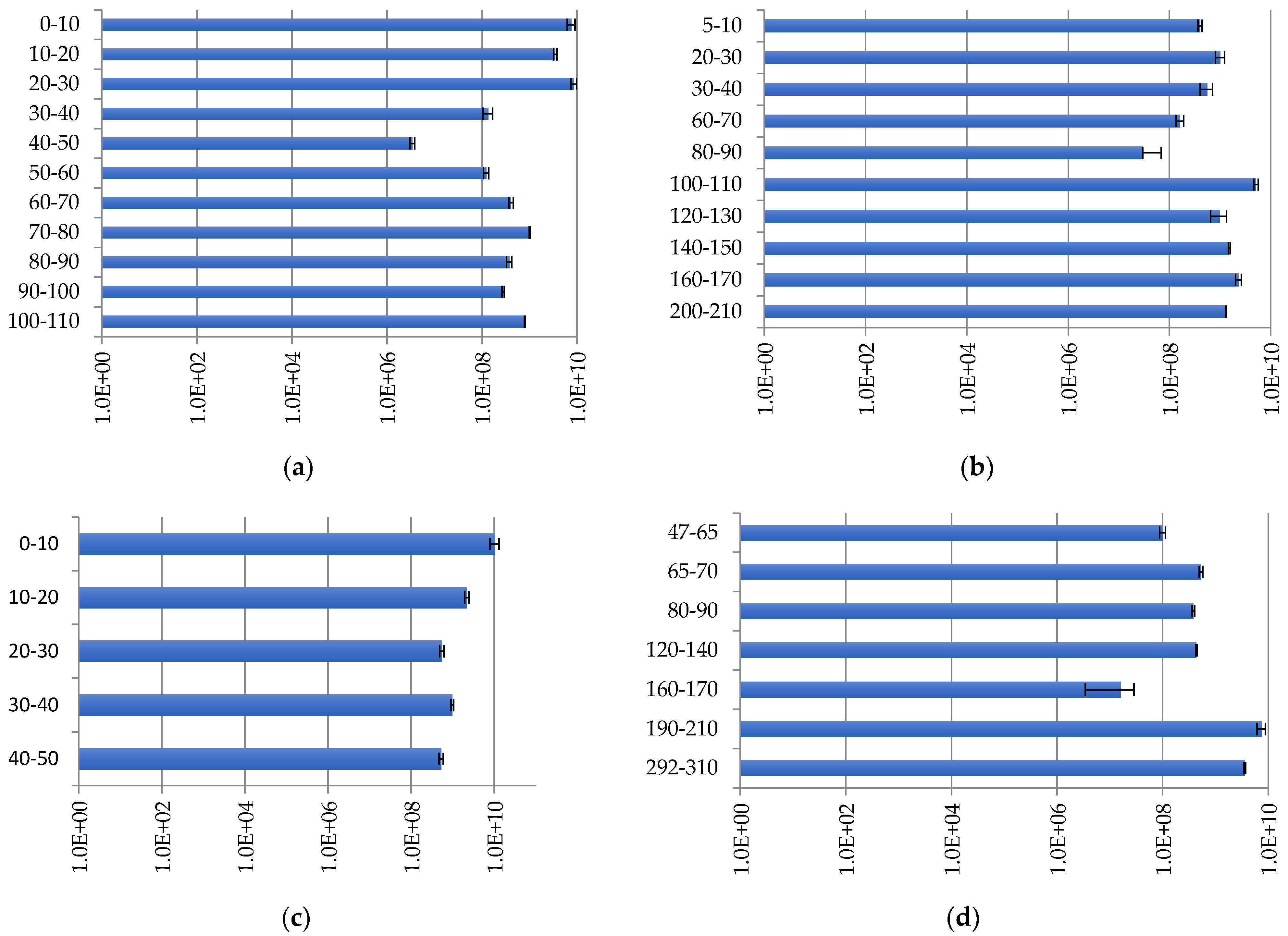
2.3.2. The Archaeal Community Distribution
2.3.3. Fungal Distribution
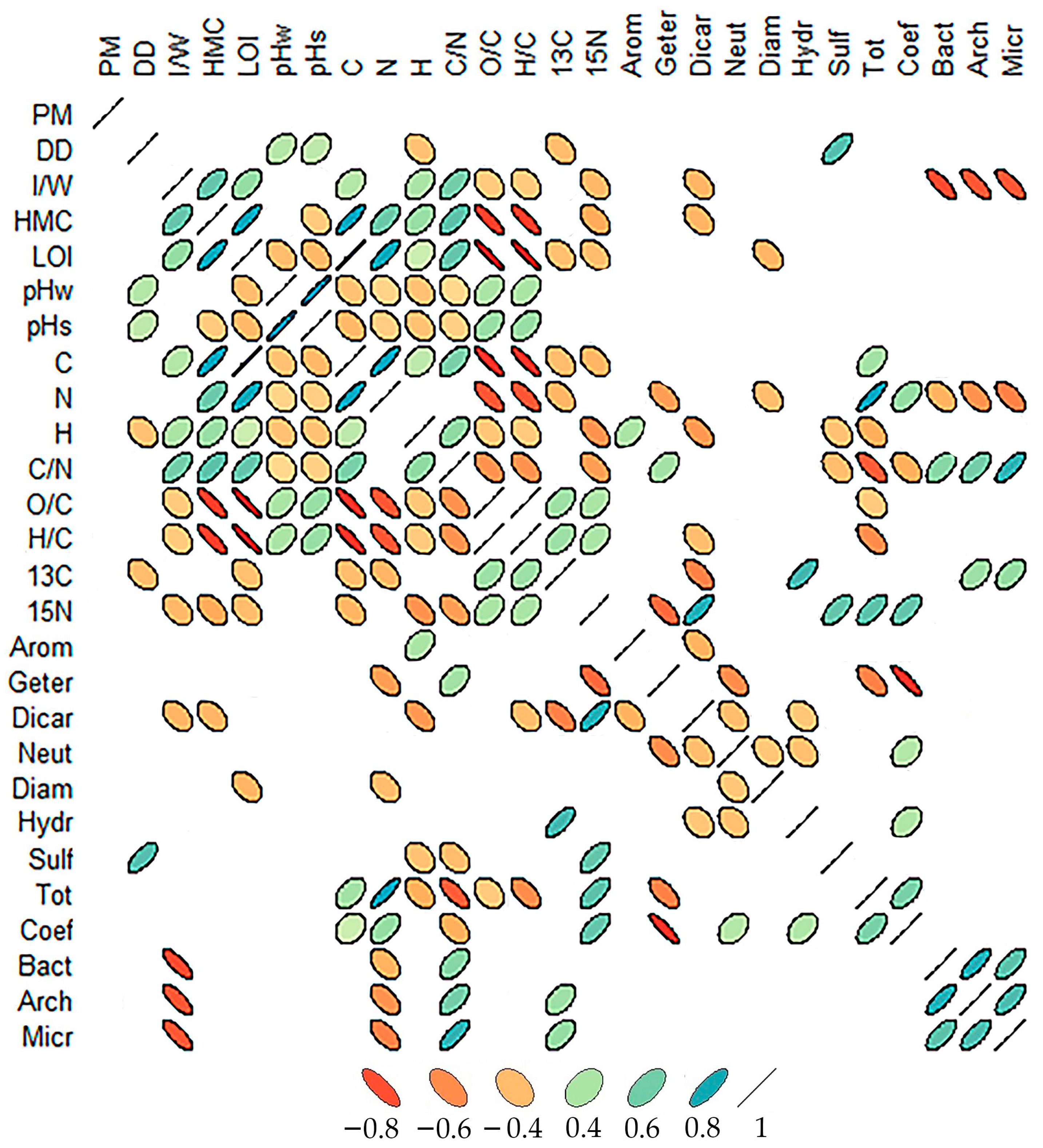
2.4. General Patterns and Correlations
3. Materials and Methods
3.1. Study Area
3.2. Soil Sampling and Laboratory Analyses
3.2.1. Soil Sampling
3.2.2. Plant Macrofossil Analyses
3.2.3. Element Analyses
3.2.4. Definition of Amino Acid Composition
3.2.5. Definition of Microbial Community Structure: Total Microbial DNA Extraction
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hugelius, G.; Loisel, J.; Chadburn, S.; Jackson, R.B.; Jones, M.; MacDonald, G.; Marushchak, M.; Olefeldt, D.; Packalen, M.; Siewert, M.B.; et al. Large stocks of peatland carbon and nitrogen are vulnerable to permafrost thaw. Proc. Natl. Acad. Sci. USA 2020, 117, 20438–20446. [Google Scholar] [CrossRef]
- Schuur, E.; McGuire, A.; Schädel, C.; Grosse, G.; Harden, J.W.; Hayes, D.J.; Hugelius, G.; Koven, C.D.; Kuhry, P.; Lawrence, D.M.; et al. Climate change and the permafrost carbon feedback. Nature 2015, 520, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Andersen, R.; Chapman, S.J.; Artz, R.R.E. Microbial communities in natural and disturbed peatlands: A review. Soil Biol. Biochem. 2013, 57, 979–994. [Google Scholar] [CrossRef]
- Artz, R.R.E. Microbial community structure and carbon substrate use in Northern peatlands. In Carbon Cycling in Northern Peatlands; Baird, A., Belyea, L., Comas, X., Reeve, A., Slater, L., Eds.; American Geophysical Union: Washington, DC, USA, 2009; pp. 111–129. [Google Scholar] [CrossRef]
- Arenz, B.E.; Held, B.W.; Jurgens, J.A.; Blanchette, R.A. Fungal colonization of exotic substrates in Antarctica. Fungal Divers. 2011, 49, 13–22. [Google Scholar] [CrossRef]
- Blaud, A.; Phoenix, G.K.; Osborn, A.M. Variation in bacterial, archaeal and fungal community structure and abundance in High Arctic tundra soil. Polar Biol. 2015, 38, 1009–1024. [Google Scholar] [CrossRef]
- Vinogradova, Y.A.; Lapteva, E.M.; Kovaleva, V.A.; Perminova, E.M. Profile distribution pattern of microfungi in the permafrost-affected peatland of forest-tundra. Micol. Phytopatol. 2019, 53, 342–353. [Google Scholar] [CrossRef]
- Vinogradova, Y.A.; Lapteva, E.M.; Kovaleva, V.A.; Perminova, E.M. Soil fungi biomass and diversity of soil microfungi in the active layer of south tundra peatlands. Micol. Phytopatol. 2021, 55, 105–118. [Google Scholar] [CrossRef]
- Dedysh, S.N.; Pankratov, T.A.; Belova, S.E.; Kulichevskaya, I.S.; Liesack, W. Phylogenetic analysis and in situ identification of Bacteria community composition in an acidic Sphagnum peat bog. Appl. Environ. Microbiol. 2006, 72, 2110–2117. [Google Scholar] [CrossRef] [PubMed]
- Dedysh, S.N. Cultivating uncultured bacteria from northern wetlands: Knowledge gained and remaining gaps. Front. Microbiol. 2011, 2, 184. [Google Scholar] [CrossRef]
- Lin, X.; Green, S.; Tfaily, M.M.; Prakash, O.; Konstantinidis, K.T.; Corbett, J.E.; Chanton, J.P.; Cooper, W.T.; Kostka, J.E. Microbial community structure and activity linked to contrasting biogeochemical gradients in bog and fen environments of the Glacial Lake Agassiz Peatland. Appl. Environ. Microbiol. 2012, 78, 7023–7031. [Google Scholar] [CrossRef] [PubMed]
- Preston, M.D.; Smemo, K.A.; McLaughlin, J.W.; Basiliko, N. Peatland microbial communities and decomposition processes in the James Bay Lowlands, Canada. Front. Microbiol. 2012, 3, 70. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, S.; Barot, S.; Barre, P.; Bdioui, N.; Mary, B.; Rumpel, C. Stability of organic carbon in deep soil layers controlled by fresh carbon supply. Nature 2007, 450, 277–280. [Google Scholar] [CrossRef]
- Rumpel, C.; Kogel-Knabner, I. Deep soil organic matter—A key but poorly understood component of terrestrial C cycle. Plant Soil 2011, 338, 143–158. [Google Scholar] [CrossRef]
- Ye, R.Z.; Jin, Q.S.; Bohannan, B.; Keller, J.K.; McAllister, S.A.; Bridgham, S.D. pH controls over anaerobic carbon mineralization, the efficiency of methane production, and methanogenic pathways in peatlands across an ombrotrophic-minerotrophic gradient. Soil Biol. Biochem. 2012, 54, 36–47. [Google Scholar] [CrossRef]
- Cowie, G.; Hedges, J. Biochemical indicators of diagenetic alteration in natural organic matter mixtures. Nature 1994, 369, 304–307. [Google Scholar] [CrossRef]
- Lipson, D.A.; Monson, R.K. Plant-microbe competition for soil amino acids in the alpine tundra: Effects of freeze-thaw and dry-rewet events. Oecologia 1998, 113, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Lipson, D.; Näsholm, T. The unexpected versatility of plants: Organic nitrogen use and availability in terrestrial ecosystems. Oecologia 2001, 128, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Krab, E.; Cornelissen, J.; Lang, S.; van Logtestijn, R. Amino acid uptake among wide-ranging moss species may contribute to their strong position in high-latitude ecosystems. Plant Soil 2008, 304, 199–208. [Google Scholar] [CrossRef][Green Version]
- Umarov, M.M.; Kurakov, A.V.; Stepanov, A.L. Mikrobiologicheskaya Transformatsiya Azota v Pochve (Microbiological Transformation of Nitrogen in Soil); GEOS: Moscow, Russia, 2007; p. 138. Available online: https://rusneb.ru/catalog/000199_000009_003400783/ (accessed on 31 August 2022). (In Russian)
- Zaguralskaya, L.M. Mikrobnaya Transformatsiya Organicheskogo Veshchestva v Lesnykh Pochvakh Karelii (Microbial Transformation of Organic Matter in Forest Soils of Karelia); Nauka: Saint-Petersburg, Russia, 1993; p. 136. Available online: https://www.studmed.ru/zagypalskaya-lm-mikrobnaya-transformaciya-organicheskogo-veschestva-v-lesnyh-pochvah-karelii_2955590c953.html (accessed on 31 August 2022). (In Russian)
- Yu, Z.C.; Campbell, I.D.; Campbell, C.; Vitt, D.H.; Bond, G.C.; Apps, M.J. Carbon sequestration in western Canadian peat highly sensitive to Holocene wet-dry climate cycles at millennial timescales. Holocene 2003, 13, 801–808. [Google Scholar] [CrossRef]
- Marushchak, M.E.; Pitkämäki, A.; Koponen, H.; Biasi, C.; Seppälä, M.; Martikainen, P.J. Hot spots for nitrous oxide emissions found in different types of permafrost peatlands. Glob. Chang. Biol. 2011, 17, 2601–2614. [Google Scholar] [CrossRef]
- Pastukhov, A.V.; Kaverin, D.A.; Goncharova, N.N. Structure and properties of permafrost peatlands in the European north-east. Teor. Prikl. Ekol. 2015, 1, 77–84. Available online: http://envjournal.ru/ari/v2015/v1/files/15112.pdf (accessed on 9 October 2022).
- Pastukhov, A.V.; Knoblauch, C.; Yakovleva, E.V.; Kaverin, D.A. Markers of soil organic matter transformation in permafrost peat mounds of northeastern Europe. Eurasian Soil Sci. 2018, 51, 42–53. [Google Scholar] [CrossRef]
- Pastukhov, A.; Knoblauch, C.; Beer, C.; Ryzhova, I.; Kaverin, D. Development of permafrost-affected peatlands in the southern limit of the European Russian cryolithozone and their vulnerability to future warming. Sci. Total Environ. 2022, 828, 154350. [Google Scholar] [CrossRef] [PubMed]
- Pastukhov, A.V.; Kaverin, D.A. Ecological state of peat plateaus in northeastern European Russia. Russ. J. Ecol. 2016, 47, 125–132. [Google Scholar] [CrossRef]
- Pastukhov, A.V.; Marchenko-Vagapova, T.I.; Kaverin, D.A.; Kulizhskii, S.P.; Kuznetsov, O.L.; Panov, V.S. Dynamics of peat plateau near the southern boundary of the east European permafrost zone. Eurasian Soil Sci. 2017, 50, 526–538. [Google Scholar] [CrossRef]
- Pastukhov, A.; Marchenko-Vagapova, T.; Loiko, S.; Kaverin, D. Vulnerability of the ancient permafrost peatlands in Western Siberia. Plants 2021, 10, 2813. [Google Scholar] [CrossRef] [PubMed]
- Vasilevich, R.S.; Beznosikov, V.A. Amino acid composition of humic substances in tundra soils. Eurasian Soil Sci. 2015, 48, 593–599. [Google Scholar] [CrossRef]
- Vasilevich, R.; Lodygin, E.; Abakumov, E. Molecular composition of humic substances isolated from permafrost peat soils of the eastern European Arctic. Pol. Polar Res. 2018, 39, 481–503. [Google Scholar] [CrossRef]
- Kumar, V.; Sharma, A.; Kaur, R.; Thukral, A.K.; Bhardwaj, R.; Ahmad, P. Differential distribution of amino acids in plants. Amino Acids 2017, 49, 821–869. [Google Scholar] [CrossRef] [PubMed]
- Ustunol, I.B.; Gonzalez-Pech, N.; Grassian, V.H. pH-dependent adsorption of α-amino acids, lysine, glutamic acid, serine and glycine, on TiO2 nanoparticle surfaces. J. Colloid Interface Sci. 2019, 554, 362–375. [Google Scholar] [CrossRef]
- Aerts, R.; Verhoeven, J.T.A.; Whigham, D.F. Plant-mediated controls on nutrient cycling in temperate fens and bogs. Ecology 1999, 80, 2170–2181. Available online: https://www.jstor.org/stable/176901 (accessed on 9 October 2022). [CrossRef]
- Thormann, M.N.; Currah, R.S.; Bayley, S.E. Patterns of distribution of microfungi in decomposing bog and fen plants. Can. J. Bot. 2004, 82, 710–720. [Google Scholar] [CrossRef]
- Dobrovol’skaya, T.G.; Golovchenko, A.V.; Zvyagintsev, D.G. Analysis of ecological factors limiting the destruction of high-moor peat. Eurasion Soil Sci. 2014, 47, 182–193. [Google Scholar] [CrossRef]
- Grodnitskaya, I.D.; Sorokin, N.D. Soil-microbiological monitoring of forest-mire ecosystems in Western Siberia. Eurasion Soil Sci. 2004, 37, 824–829. Available online: https://www.researchgate.net/publication/290750809_Soil-microbiological_monitoring_of_forest-mire_ecosystems_in_Western_Siberia (accessed on 9 October 2022).
- Leininger, S.; Urich, T.; Schloter, M.; Schwark, L.; Qi, J.; Nicol, G.W.; Prosser, J.I.; Schuster, S.C.; Schleper, C. Archaea predominate among ammonia-oxidizing prokaryotes in soils. Nature 2006, 442, 806–809. [Google Scholar] [CrossRef] [PubMed]
- Thauer, R.; Kaster, A.K.; Seedorf, H.; Buckel, W.; Hedderich, R. Methanogenic archaea: Ecologically relevant differences in energy conservation. Nat. Rev. Microbiol. 2008, 6, 579–591. [Google Scholar] [CrossRef]
- Aerts, R.; Wallen, B.; Malmer, N. Growth-limiting nutrients in Sphagnum-dominated bogs subject to low a high atmospheric nitrogen supply. J. Ecol. 1992, 80, 131–140. [Google Scholar] [CrossRef]
- Freeman, C.; Ostie, N.; Kang, H. An enzymic ‘latch’ on a global carbon store—A shortage of oxygen locks up carbon in peatlands by restraining a single enzyme. Nature 2001, 409, 149. [Google Scholar] [CrossRef]
- Kim, J.; Rochefort, L.; Hogue-Hugron, S.; Alqulaiti, Z.; Dunn, C.; Pouliot, R.; Jones, T.G.; Freeman, C.; Kang, H. Water table fluctuation in peatlands facilitates fungal proliferation, impedes Sphagnum growth and accelerates decomposition. Front. Earth Sci. 2021, 8, 579329. [Google Scholar] [CrossRef]
- Golovchenko, A.V.; Semenova, T.A.; Morozov, A.A.; Glukhova, T.V.; Inisheva, L.I. The mycobiota of eutrophic peatlands. Eurasian Soil Sci. 2022, 55, 348–356. [Google Scholar] [CrossRef]
- Bogdanova, O.Y.; Golovchenko, A.V.; Lysak, L.V.; Zvyagintsev, D.G.; Glukhova, T.V. Viability of bacteria in peatlands. Eurasian Soil Sci. 2014, 47, 297–303. [Google Scholar] [CrossRef]
- Golovchenko, A.V.; Zvyagintsev, D.G.; Kurakov, A.V.; Semenova, T.A. Abundance, diversity, viability, and factorial ecology of fungi in peatbogs. Eurasian Soil Sci. 2013, 46, 74–90. [Google Scholar] [CrossRef]
- Myhre, G.; Shindell, D.; Breon, F.M.; Collins, W.; Fuglestvedt, J.; Huang, J.; Koch, D.; Lamarque, J.-F.; Lee, D.; Mendoza, B.; et al. Anthropogenic and natural radiative forcing. In Climate Change 2013: The Physical Science Basis. Contributions of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University: Cambridge, UK; New York, NY, USA, 2013; pp. 659–740. Available online: https://www.ipcc.ch/site/assets/uploads/2018/02/WG1AR5_Chapter08_FINAL.pdf (accessed on 31 August 2022).
- Climate Database Ust-Usa. Available online: https://ru.climate-data.org/aзия/poccийcкaя-фeдepaция/pecпyбликa-кoми/ycть-yca-333966/ (accessed on 31 August 2022). (In Russian).
- Climate Database Pechora. Available online: https://ru.climate-data.org/aзия/poccийcкaя-фeдepaция/pecпyбликa-кoми/пeчopa-32650/ (accessed on 31 August 2022). (In Russian).
- Yurkovskaya, T.K. Geografiya i Kartografiya Rastitel’nosti Bolot Yevropeyskoy Rossii i Sopredel’nykh Territoriy (Geography and Cartography of the Vegetation of the Swamps of European Russia and Adjacent Territories); Botanical Institute named V.L. Komarov: Saint-Peterburg, Russia, 1992; p. 256. Available online: https://www.studmed.ru/yurkovskaya-t-k-geografiya-i-kartografiya-rastitelnosti-bolot-evropeyskoy-rossii-i-sopredelnyh-territoriy_b1e4d33cc14.html (accessed on 31 August 2022). (In Russian)
- IUSS Working Group WRB. World Reference Base for Soil Resources 2014: Update 2015; Rep. 106; Food and Agriculture Organization of the United Nations: Rome, Italy, 2015; p. 203. Available online: http://www.fao.org/3/a-i3794en.pdf (accessed on 31 August 2022).
- Mossberg, B.; Stenberg, L. Den. Nya Nordiska Floran; Wahlström & Widstrand: Stockholm, Sweden, 2003; p. 928. Available online: https://www.bonnierfakta.se/bocker/151725/den-nya-nordiska-floran/ (accessed on 31 August 2022). (In Norwegian)
- Laine, J.; Harju, P.; Timonen, T.; Laine, A.; Tuittila, E.S.; Minkkinen, K.; Vasander, H. The Intricate Beauty of Sphagnum Mosses: A Finnish Guide to Identification; University of Helsinki Department of Forest Sciences Publications: Helsinki, Finland, 2011; p. 191. Available online: https://www.nhbs.com/the-intricate-beauty-of-sphagnum-mosses-book-2 (accessed on 31 August 2022).
- Kutenkov, S.A. The computer program «Korpi» for the construction of stratigraphic charts of peat composition. Trans. Karel. Res. Cent. Russ. Acad. Sci. 2013, 6, 171–176. Available online: https://www.elibrary.ru/item.asp?id=21112812 (accessed on 31 August 2022). (In Russian).
- Heiri, O.; Lotter, A.F.; Lemcke, G. Loss on ignition as a method for estimating organic and carbonate content in sediments: Reproducibility comparability of results. J. Paleolimnol. 2001, 25, 101–110. [Google Scholar] [CrossRef]
- Jurgens, G.; Saano, A. Diversity of soil archaea in boreal forest before and after clear-cutting and prescribed burning. FEMS Microbiol. Ecol. 1999, 29, 205–213. [Google Scholar] [CrossRef]
- Moskalenko, S.E.; Chabelskaya, S.V.; Inge-Vechtomov, S.G.; Philippe, M.; Zhouravleva, G.A. Viable nonsense mutants for the essential gene SUP45 of Saccharomyces cerevisiae. BMC Mol. Biol. 2003, 4, 2. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stacke-brandt, E., Goodfellow, M., Eds.; Wiley: Hoboken, NJ, USA, 1991; pp. 115–175. [Google Scholar]
- Yu, Y.; Lee, C.; Kim, J.; Hwang, S. Group-specific primer and probe sets to detect methanogenic communities using quantitative real-time polymerase chain reaction. Biotechnol. Bioeng. 2005, 89, 670–679. [Google Scholar] [CrossRef]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pastukhov, A.; Kovaleva, V.; Kaverin, D. Microbial Community Structure in Ancient European Arctic Peatlands. Plants 2022, 11, 2704. https://doi.org/10.3390/plants11202704
Pastukhov A, Kovaleva V, Kaverin D. Microbial Community Structure in Ancient European Arctic Peatlands. Plants. 2022; 11(20):2704. https://doi.org/10.3390/plants11202704
Chicago/Turabian StylePastukhov, Alexander, Vera Kovaleva, and Dmitry Kaverin. 2022. "Microbial Community Structure in Ancient European Arctic Peatlands" Plants 11, no. 20: 2704. https://doi.org/10.3390/plants11202704
APA StylePastukhov, A., Kovaleva, V., & Kaverin, D. (2022). Microbial Community Structure in Ancient European Arctic Peatlands. Plants, 11(20), 2704. https://doi.org/10.3390/plants11202704