

Mitigation of Salinity Stress in Maize Seedlings by the Application of Vermicompost and Sorghum Water Extracts

 , , , , and
, , , , and 
Abstract
1. Introduction
2. Results
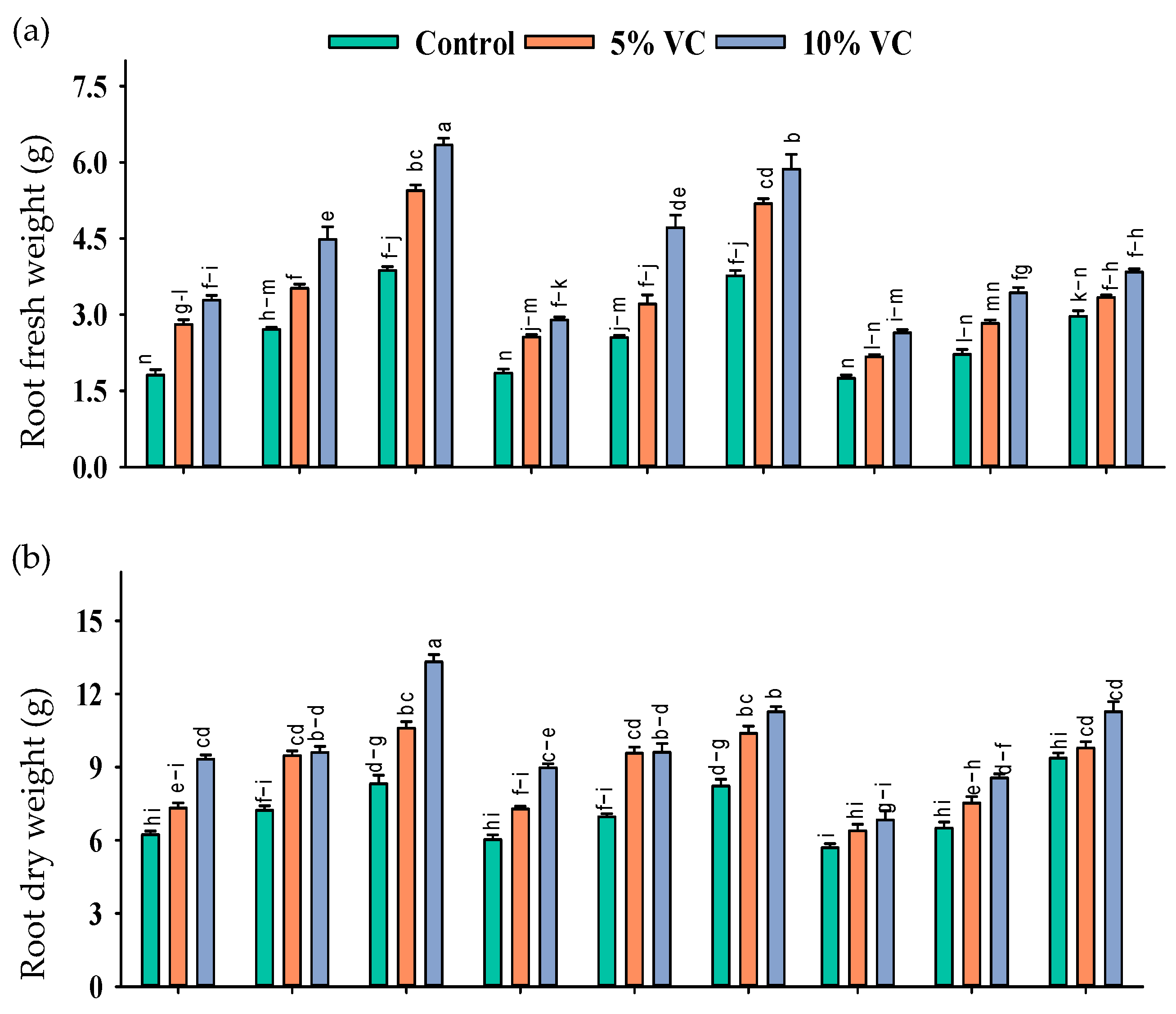
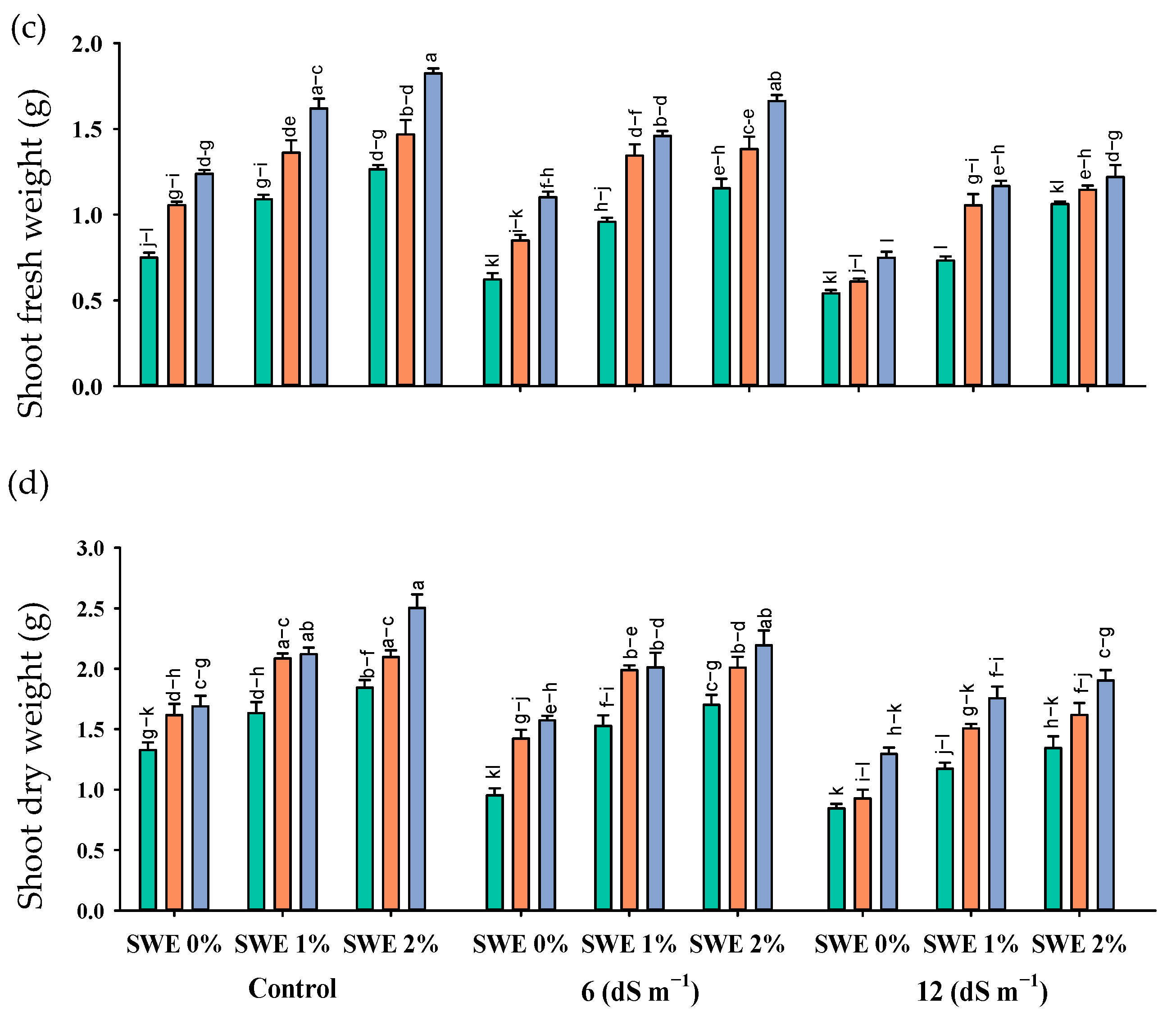
2.1. Morphological Parameters
Plant Biomass
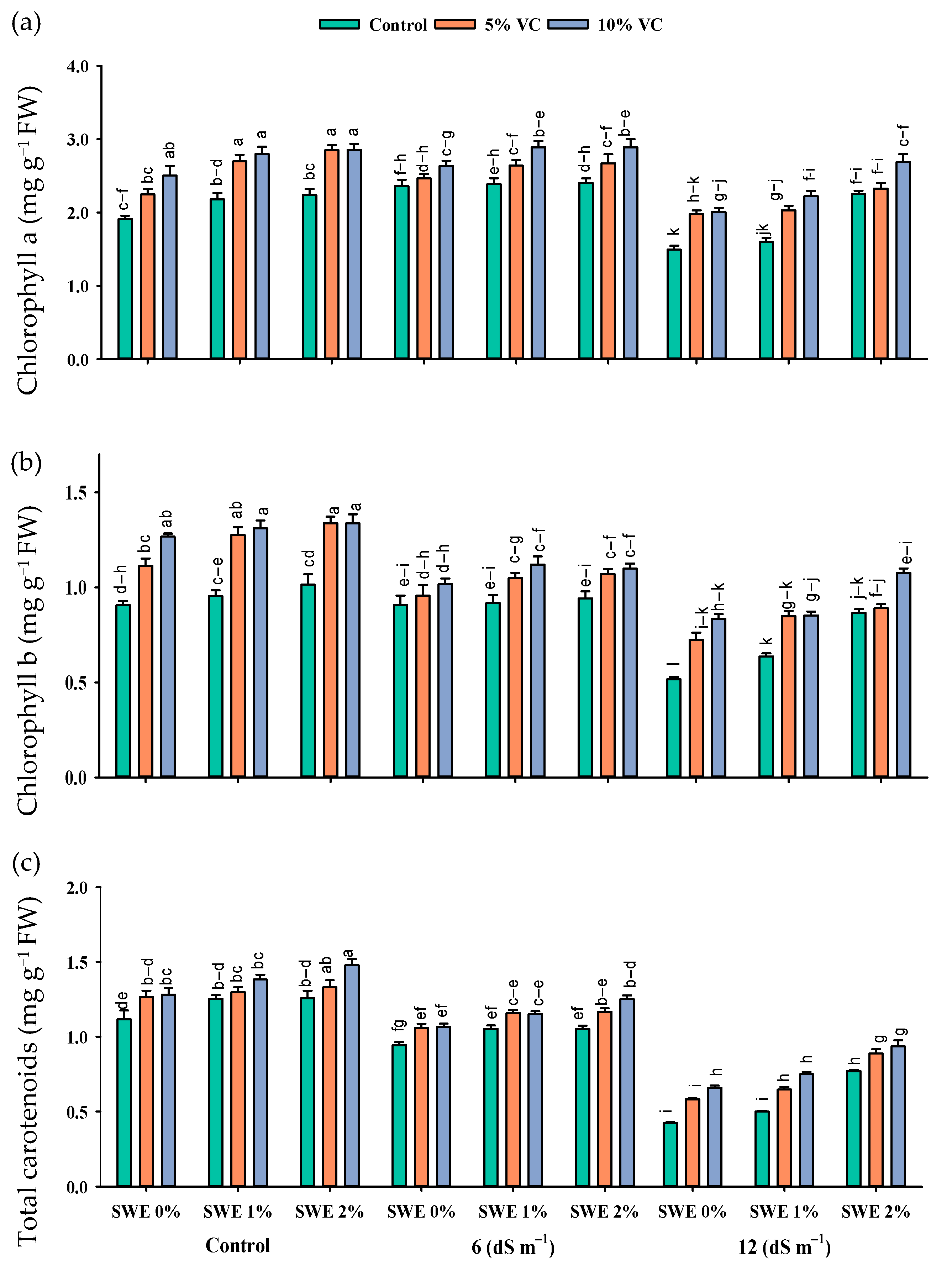
2.2. Photosynthetic Pigment
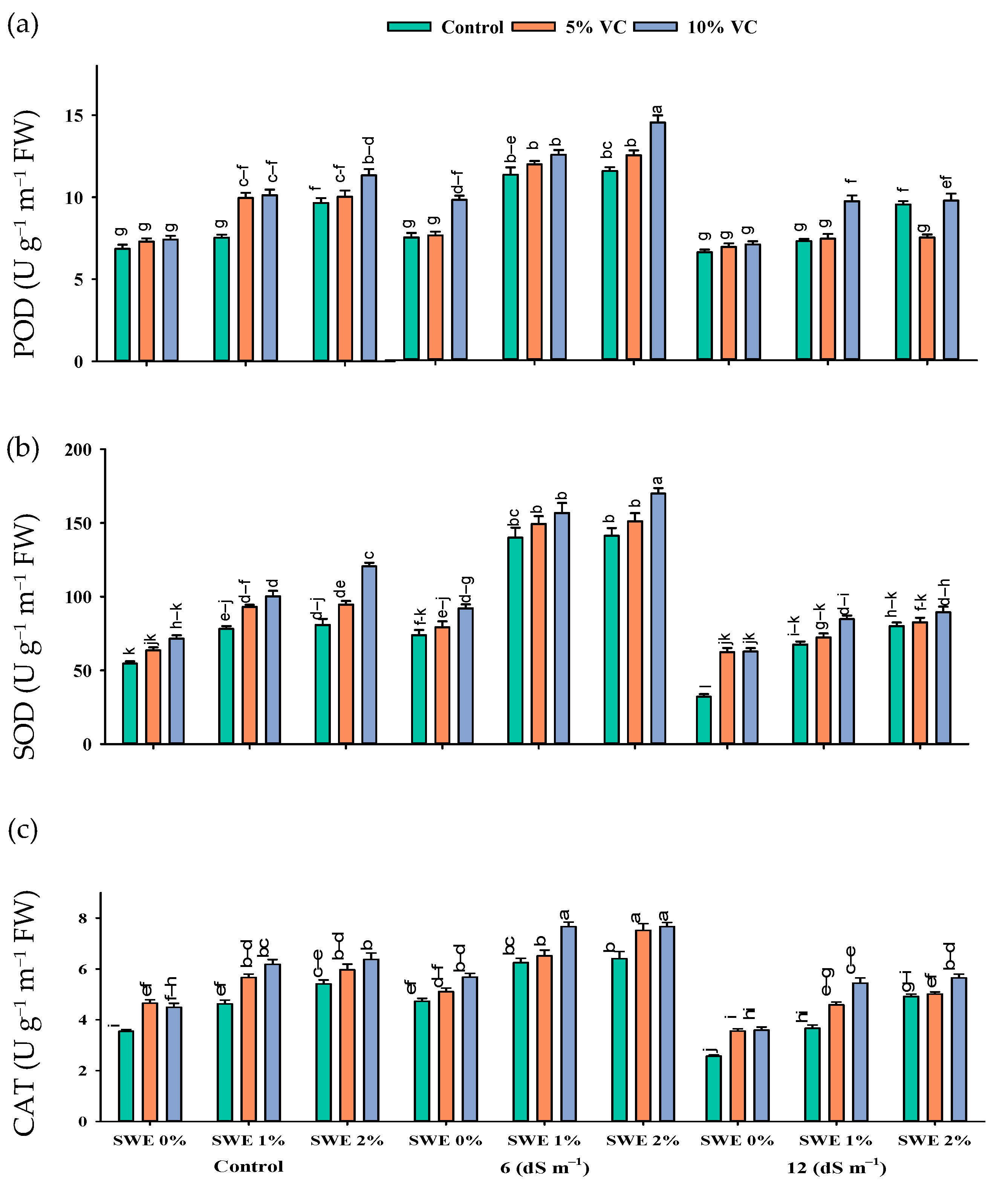
2.3. Antioxidant Enzyme Activities
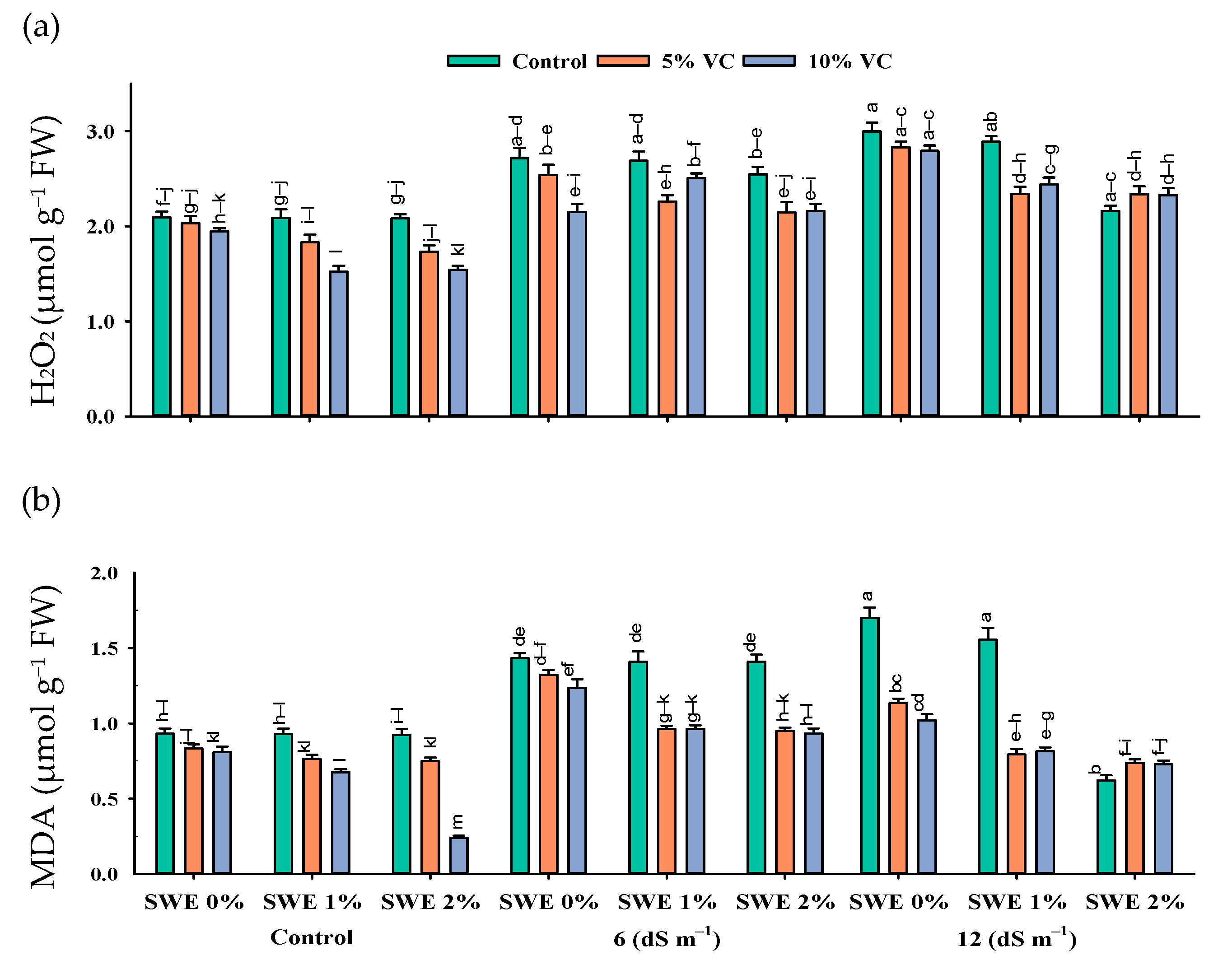
2.4. Leaf Malondialdehyde (MDA) and H2O2 Content
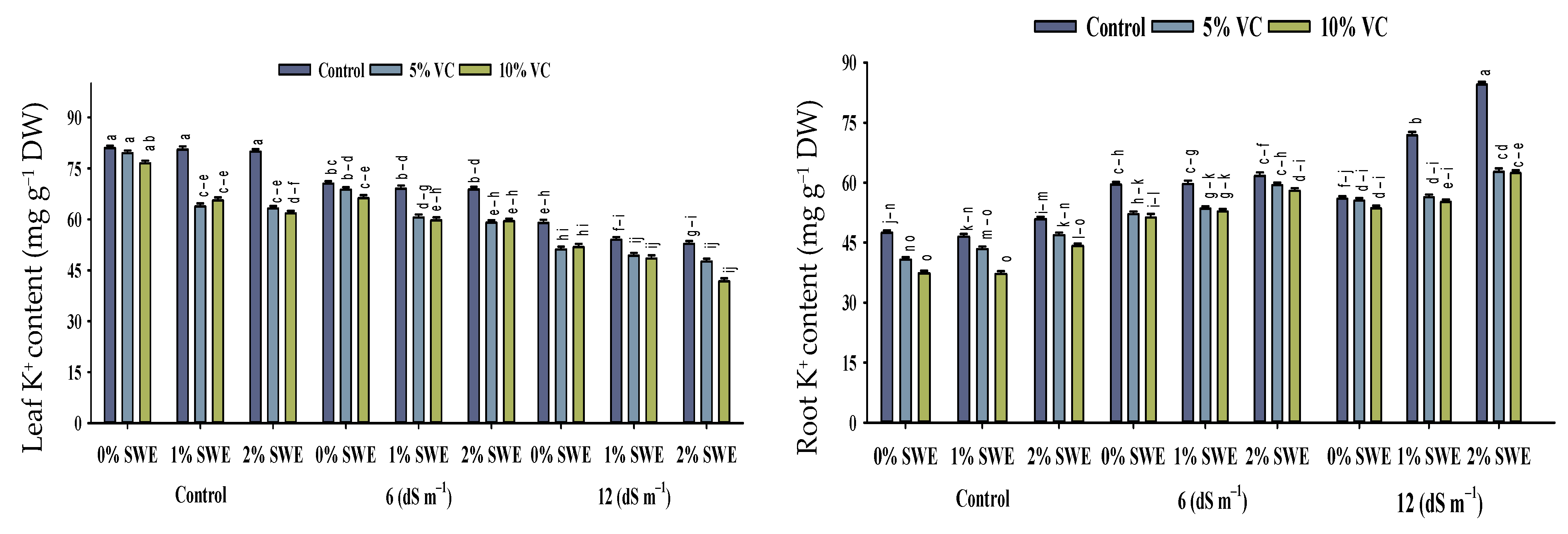
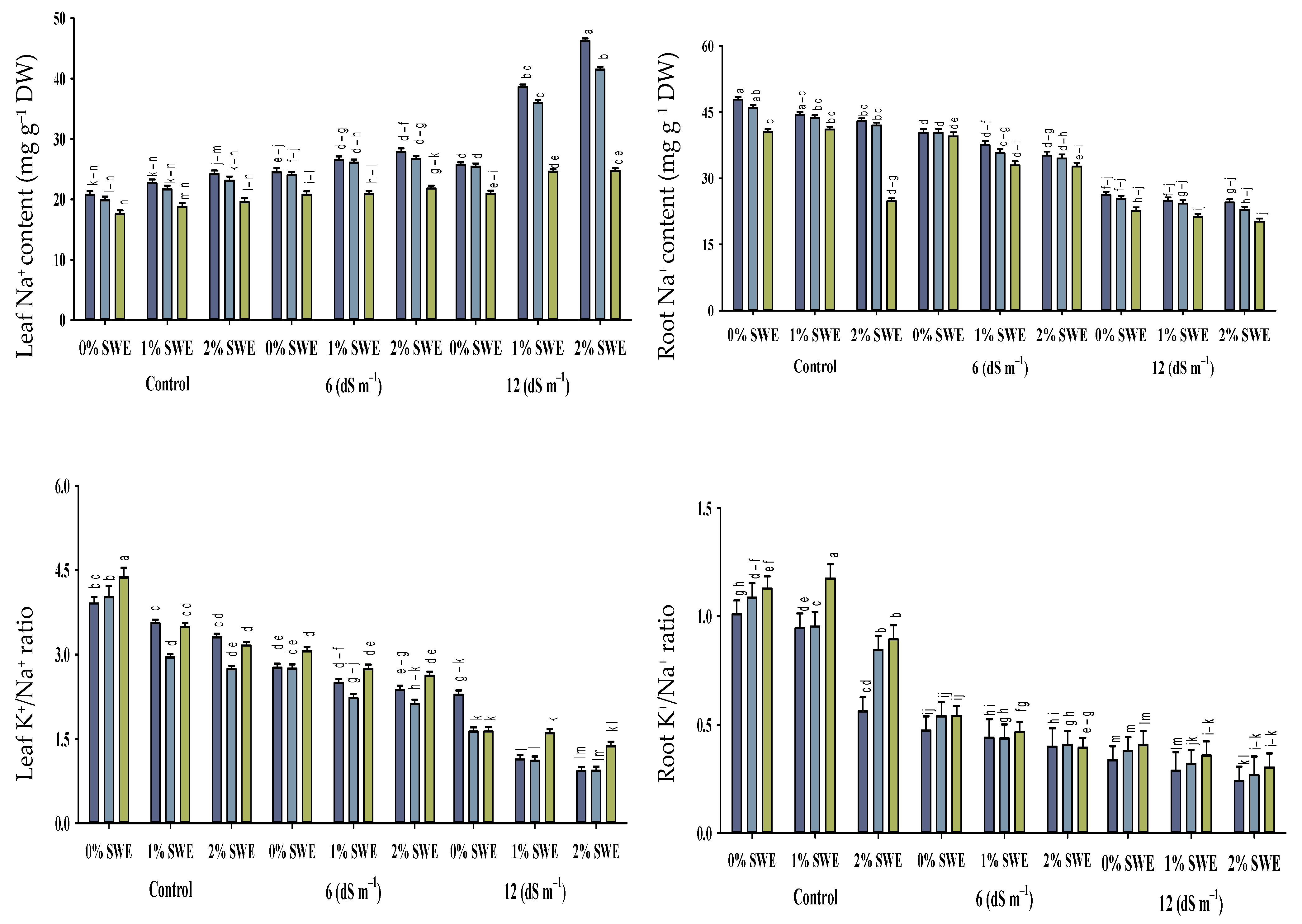
2.5. Mineral Ion Contents
2.5.1. Na+ and K+ Contents
2.5.2. Maize K+/Na+ Ratio
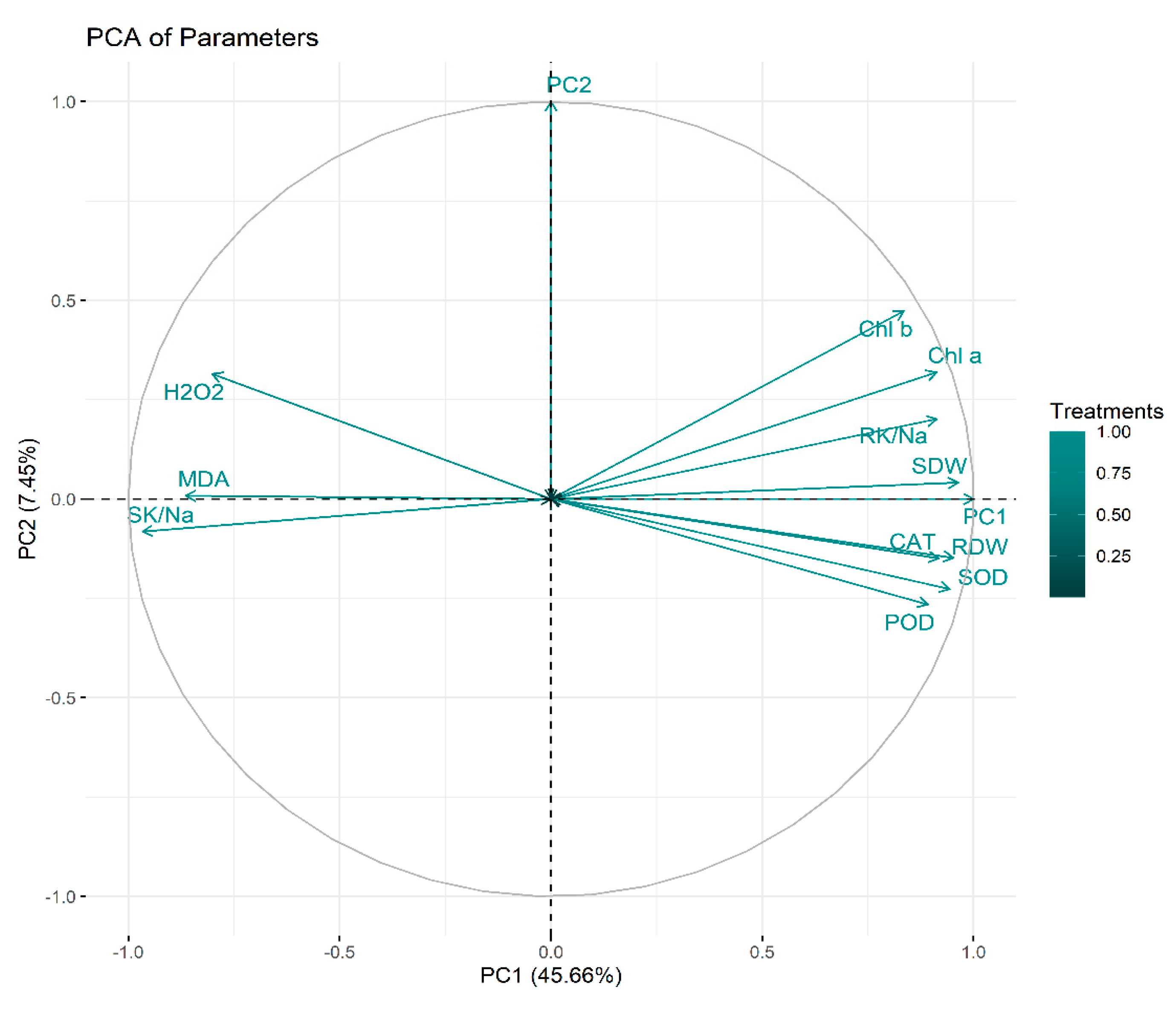
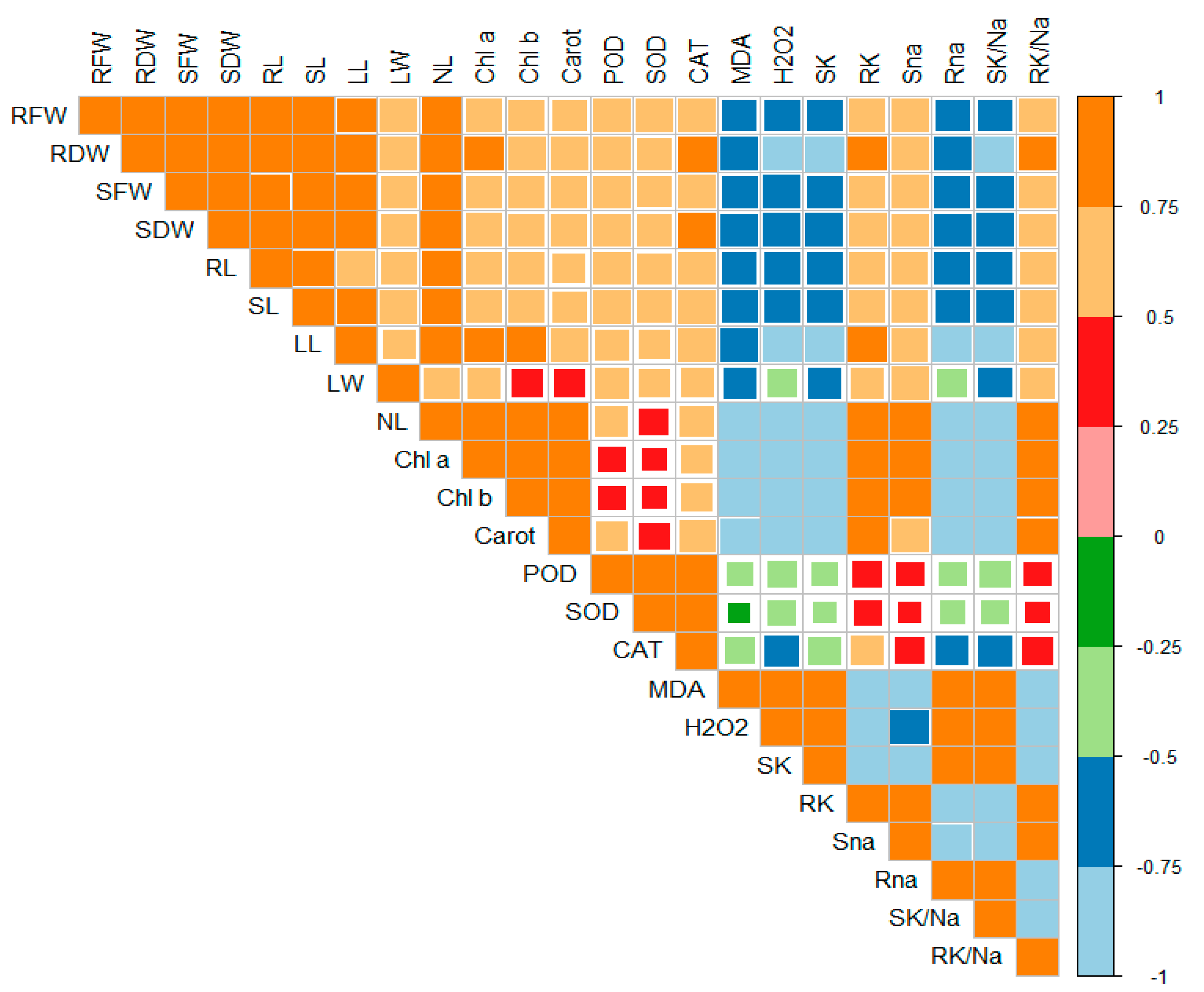
2.6. Principal Component Analysis
3. Discussion
4. Materials and Methods
4.1. Location of the Experiment
4.2. Sorghum Water Extract Preparation
4.3. Experimental Setup
4.4. Imposition of Salinity
4.5. Crop Husbandry
4.6. Measurement of Morphological Parameters
4.7. Determination of Photosynthetic Pigments
4.8. Determination of Enzymatic Antioxidant Activities
4.9. Determination of Reactive Oxygen Species (ROS)
H2O2 and MDA Contents
4.10. Determination of Mineral Ion (Na+ and K+) Concentration
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arora, N.K.; Fatima, T.; Mishra, J.; Mishra, I.; Verma, S.; Verma, R.; Verma, M.; Bhattacharya, A.; Verma, P.; Mishra, P.; et al. Halo-tolerant plant growth promoting rhizobacteria for improving productivity and remediation of saline soils. J. Adv. Res. 2020, 26, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Sahab, S.; Suhani, I.; Srivastava, V.; Chauhan, P.S.; Singh, R.P.; Prasad, V. Potential risk assessment of soil salinity to agroecosystem sustainability: Current status and management strategies. Sci. Total Environ. 2021, 764, 144164. [Google Scholar] [CrossRef]
- Liu, Y.; Cao, X.; Yue, L.; Wang, C.; Tao, M.; Wang, Z.; Xing, B. Foliar-applied cerium oxide nanomaterials improve maize yield under salinity stress: Reactive oxygen species homeostasis and rhizobacteria regulation. Environ. Pollut. 2022, 299, 118900. [Google Scholar] [CrossRef]
- Ali, M.; Afzal, S.; Parveen, A.; Kamran, M.; Javed, M.R.; Abbasi, G.H.; Malik, Z.; Riaz, M.; Ahmad, S.; Chattha, M.S.; et al. Silicon mediated improvement in the growth and ion homeostasis by decreasing Na+ uptake in maize (Zea mays L.) cultivars exposed to salinity stress. Plant Physiol. Biochem. 2021, 158, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Hussain, M.; Wakeel, A.; Siddique, K.H. Salt stress in maize: Effects, resistance mechanisms, and management. A review. Agron. Sustain. Dev. 2015, 35, 461–481. [Google Scholar] [CrossRef]
- Haque, M.A.; Rafii, M.Y.; Yusoff, M.M.; Ali, N.S.; Yusuff, O.; Datta, D.R.; Anisuzzaman, M.; Ikbal, M.F. Advanced breeding strategies and future perspectives of salinity tolerance in rice. Agronomy 2021, 11, 1631. [Google Scholar] [CrossRef]
- Viator, R.P.; Johnson, R.M.; Grimm, C.C.; Richard, E.P., Jr. Allelopathic, autotoxic, and hormetic effects of postharvest sugarcane residue. Agron. J. 2006, 98, 1526–1531. [Google Scholar] [CrossRef]
- Abbas, T.; Nadeem, M.A.; Tanveer, A.; Chauhan, B.S. Can hormesis of plant-released phytotoxins be used to boost and sustain crop production? Crop Protect. 2017, 93, 69–76. [Google Scholar] [CrossRef]
- Ibrahim, M.U.; Khaliq, A.; Hussain, S.; Murtaza, G. Sorghum water extract application mediates antioxidant defense and confers drought stress tolerance in wheat. J. Plant Growth Regul. 2022, 41, 863–874. [Google Scholar] [CrossRef]
- Bakhtavar, M.A.; Afzal, I.; Basra, S.M.A.; Ahmad, A.U.H.; Noor, M.A. Physiological strategies to improve the performance of spring maize (Zea mays L.) planted under early and optimum sowing conditions. PLoS ONE 2015, 10, e0124441. [Google Scholar] [CrossRef]
- Khaliq, A.; Perveen, S.; Alamer, K.H.; Zia Ul Haq, M.; Rafique, Z.; Alsudays, I.M.; Althobaiti, A.T.; Saleh, M.A.; Hussain, S.; Attia, H. Arbuscular Mycorrhizal Fungi Symbiosis to Enhance Plant–Soil Interaction. Sustainability 2022, 14, 7840. [Google Scholar] [CrossRef]
- Beyk-Khormizi, A.; Hosseini Sarghein, S.; Sarafraz-Ardakani, M.R.; Moshtaghioun, S.M.; Mousavi-Kouhi, S.M.; Ganjeali, A. Ameliorating effect of vermicompost on Foeniculum vulgare under saline condition. J. Plant Nutr. 2022, 1–15. [Google Scholar] [CrossRef]
- Ayyobi, H.; Hassanpour, E.; Alaqemand, S.; Fathi, S.; Olfati, J.A.; Peyvast, G. Vermicompost leachate and vermiwash enhance French dwarf bean yield. Int. J. Veg. Sci. 2014, 20, 21–27. [Google Scholar] [CrossRef]
- Beykkhormizi, A.; Abrishamchi, P.; Ganjeali, A.; Parsa, M. Effect of vermicompost on some morphological, physiological and biochemical traits of bean (Phaseolus vulgaris L.) under salinity stress. J. Plant Nutr. 2016, 39, 883–893. [Google Scholar] [CrossRef]
- Dikobe, T.B.; Mashile, B.; Sinthumule, R.R.; Ruzvidzo, O. Distinct morpho-physiological responses of maize to salinity stress. Am. J. Plant Sci. 2021, 12, 946–959. [Google Scholar] [CrossRef]
- Saddiq, M.S.; Iqbal, S.; Afzal, I.; Ibrahim, A.M.; Bakhtavar, M.A.; Hafeez, M.B.; Jahanzaib; Maqbool, M.M. Mitigation of salinity stress in wheat (Triticum aestivum L.) seedlings through physiological seed enhancements. J. Plant Nutr. 2019, 42, 1192–1204. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A.; Läuchli, A. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot. 2006, 57, 1025–1043. [Google Scholar] [CrossRef]
- Farooq, M.; Rehman, A.; Al-Alawi, A.K.; Al-Busaidi, W.M.; Lee, D.J. Integrated use of seed priming and biochar improves salt tolerance in cowpea. Sci. Hortic. 2020, 272, 109507. [Google Scholar] [CrossRef]
- Khaliq, A.; Ibrahim, M.U.; Hussain, S.; Zia Ul Haq, M.; Al-Huqail, A.A.; Nawaz, M.; Ali, B.; Khan, F.; Ali, H.M.; Siddiqui, M.H. The hormetic effects of a brassica water extract triggered wheat growth and antioxidative defense under drought stress. App. Sci. 2022, 12, 4582. [Google Scholar] [CrossRef]
- Cai, J.F.; Liu, X.S.; Sun, K.; Jiang, F.; Zhang, M.X.; Li, H.L.; Yu, F.H. Vermicompost application enhances halophyte Suaeda salsa performance and improves coastal saline soil quality. J. Soil Sci. Plant Nutr. 2022, 22, 294–305. [Google Scholar] [CrossRef]
- Kamran, M.; Malik, Z.; Parveen, A.; Zong, Y.; Abbasi, G.H.; Rafiq, M.T.; Shaaban, M.; Mustafa, A.; Bashir, S.; Rafay, M.; et al. Biochar alleviates Cd phytotoxicity by minimizing bioavailability and oxidative stress in pak choi (Brassica chinensis L.) cultivated in Cd-polluted soil. J. Environ. Manag. 2019, 250, 109500. [Google Scholar] [CrossRef] [PubMed]
- Tammam, A.A.; Shehata, M.R.A.M.; Pessarakli, M.; El-Aggan, W.H. Vermicompost and its role in alleviation of salt stress in plants–I. Impact of vermicompost on growth and nutrient uptake of salt-stressed plants. J. Plant Nutr. 2022, 1–12. [Google Scholar] [CrossRef]
- Khaliq, A.; Zia Ul Haq, M.; Ali, F.; Aslam, F.; Matloob, A.; Navab, A.; Hussain, S. Salinity tolerance in wheat cultivars is related to enhanced activities of enzymatic antioxidants and reduced lipid peroxidation. CLEAN–Soil Air Water 2015, 43, 1248–1258. [Google Scholar] [CrossRef]
- Gupta, N.K.; Meena, S.K.; Gupta, S.; Khandelwal, S.K. Gas exchange, membrane permeability, and ion uptake in two species of Indian jujube differing in salt tolerance. Photosynthetica 2002, 40, 535–539. [Google Scholar] [CrossRef]
- Ashraf, M.; Shahbaz, M. Assessment of genotypic variation in salt tolerance of early CIMMYT hexaploid wheat germplasm using photosynthetic capacity and water relations as selection criteria. Photosynthetica 2003, 41, 273–280. [Google Scholar] [CrossRef]
- Kwon, O.K.; Mekapogu, M.; Kim, K.S. Effect of salinity stress on photosynthesis and related physiological responses in carnation (Dianthus caryophyllus). Hortic. Environ. Biotechnol. 2019, 60, 831–839. [Google Scholar] [CrossRef]
- de Azevedo Neto, A.D.; Prisco, J.T.; Enéas-Filho, J.; de Abreu, C.E.B.; Gomes-Filho, E. Effect of salt stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salt-tolerant and salt-sensitive maize genotypes. Environ. Exp. Bot. 2006, 56, 87–94. [Google Scholar] [CrossRef]
- Ajmal, Z.; Kashif Irshad, M.; Qadeer, A.; Zia Ul Haq, M.; Ullah, R.; Aqeel Sarwar, M.; Saeed, T.; Abid, M.; Hayat, A.; Ali, A.; et al. Novel magnetite nano-rods-modified biochar: A promising strategy to control lead mobility and transfer in soil-rice system. Int. J. Environ. Sci. Technol. 2022. [Google Scholar] [CrossRef]
- Mahmood, U.; Hussain, S.; Hussain, S.; Ali, B.; Ashraf, U.; Zamir, S.; Al-Robai, S.A.; Alzahrani, F.O.; Hano, C.; El-Esawi, M.A. Morpho-physio-biochemical and molecular responses of maize hybrids to salinity and waterlogging during stress and recovery phase. Plants 2021, 10, 1345. [Google Scholar] [CrossRef]
- Murat, A.T.; Abdelkarim, H.A.E.; Nilguumln, T.; Suleyman, T. Effect of salt stress on growth and ion distribution and accumulation in shoot and root of maize plant. Afr. J. Agric. Res. 2010, 5, 584–588. [Google Scholar]
- Zörb, C.; Geilfus, C.M.; Dietz, K.J. Salinity and crop yield. Plant Biol. 2019, 21, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Shabala, S.; Cuin, T.A. Potassium transport and plant salt tolerance. Physiol. Plant. 2008, 133, 651–669. [Google Scholar] [CrossRef] [PubMed]
- Tuna, A.L.; Kaya, C.; Higgs, D.; Murillo-Amador, B.; Aydemir, S.; Girgin, A.R. Silicon improves salinity tolerance in wheat plants. Environ. Exp. Bot. 2008, 62, 10–16. [Google Scholar] [CrossRef]
- Wakeel, A. Potassium–sodium interactions in soil and plant under saline-sodic conditions. J. Plant Nutr. Soil Sci. 2013, 176, 344–354. [Google Scholar] [CrossRef]
- Eker, S.; Cömertpay, G.; Konuşkan, Ö.; Ülger, A.C.; Öztürk, L.; Çakmak, İ. Effect of salinity stress on dry matter production and ion accumulation in hybrid maize varieties. Turk. J. Agric. Forest. 2006, 30, 365–373. [Google Scholar]
- Akhzari, D.; Pessarakli, M.; Khedmati, M. Effects of vermicompost and salinity stress on growth and physiological traits of Medicago rigidula L. J. Plant Nutr. 2016, 39, 2106–2114. [Google Scholar] [CrossRef]
- Khaliq, A.; Matloob, A.; Mahmood, S.; Wahid, A. Seed pre-treatments help improve maize performance under sorghum allelopathic stress. J. Crop Improv. 2013, 27, 586–605. [Google Scholar] [CrossRef]
- Hussain, M.I.; Danish, S.; Sánchez-Moreiras, A.M.; Vicente, Ó.; Jabran, K.; Chaudhry, U.K.; Branca, F.; Reigosa, M.J. Unraveling sorghum allelopathy in agriculture: Concepts and implications. Plants 2021, 10, 1795. [Google Scholar] [CrossRef]
- Aslam, Z.; Bashir, S.; Hassan, W.; Bellitürk, K.; Ahmad, N.; Niazi, N.K.; Khan, A.; Khan, M.I.; Chen, Z.; Maitah, M. Unveiling the efficiency of vermicompost derived from different biowastes on wheat (Triticum aestivum L.) plant growth and soil health. Agronomy 2019, 9, 791. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Bailly, C.; Benamar, A.; Corbineau, F.; Côme, D. Changes in malondialdehyde content and in superoxide dismutase, catalase and glutathione reductase activities in sunflower seeds as related to deterioration during accelerated aging. Physiol. Plant. 1996, 97, 104–110. [Google Scholar] [CrossRef]
- Shavrukov, Y.; Langridge, P.; Tester, M. Salinity tolerance and sodium exclusion in genus Triticum. Breed. Sci. 2009, 59, 671–678. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean Sum of Square | ||||||
|---|---|---|---|---|---|---|
| Characteristics | Salinity | VC | SWE | Salinity × VC | Salinity × SWE | Salinity × VC × SWE |
| Root fresh weight (g) | 11.02 ** | 21.77 ** | 24.10 ** | 2.11 ** | 1.00 ** | 0.33 ** |
| Root dry weight (g) | 1.25 ** | 1.66 ** | 1.35 ** | 0.02 ** | 0.05 ** | 0.03 ** |
| Shoot fresh weight (g) | 22.97 ** | 48.01 ** | 59.82 ** | 1.71 ** | 2.00 ** | 1.47 ** |
| Shoot dry weight (g) | 1.53 ** | 2.93 ** | 4.04 ** | 0.05 ** | 0.07 ** | 0.05 ** |
| Chlorophyll a (mg g−1 FW) | 6.70 ** | 0.55 ** | 1.35 ** | 0.05 ** | 0.08 ** | 0.01 ns |
| Chlorophyll b (mg g−1 FW) | 1.84 ** | 0.11 ** | 0.46 ** | 0.006 ns | 0.04 ** | 0.01 ns |
| Total carotenoid (mg g−1 FW) | 4.22 ** | 0.29 ** | 0.68 ** | 0.04 ** | 0.20 ** | 0.044 ** |
| Peroxidase (U g−1m−1FW) | 123.37 ** | 103.97 ** | 34.59 ** | 3.31 ** | 0.66 * | 0.95 ** |
| Superoxide dismutase (U g−1m−1FW) | 29,908.5 ** | 17,737 ** | 5092.1 ** | 2478.6 ** | 150.7 ** | 240.7 ** |
| Catalase (U g−1m−1FW) | 33.93 ** | 22.47 ** | 11.91 ** | 0.25 * | 0.21 * | 0.18 * |
| Leaf Na+ (mg g−1 DW) | 4592.72 ** | 511.36 ** | 770.07 ** | 225.86 ** | 189.59 ** | 34.47 ** |
| Leaf K+ (mg g−1 DW) | 3289.3 ** | 447.81 ** | 694.25 ** | 24.37 ** | 26.68 ** | 23.68 ** |
| Leaf K+/Na+ ratio | 36.33 ** | 4.94 ** | 7.38 ** | 0.57 ** | 0.58 ** | 0.04 * |
| Root Na+ (mg g−1 DW) | 2905.5 ** | 442.88 ** | 858.07 ** | 35.65 ** | 113.85 ** | 23.84 ** |
| Root K+ (mg g−1 DW) | 3400.81 ** | 121.68 ** | 218.18 ** | 33.74 ** | 39.3 ** | 22.78 ** |
| Root K+/Na+ ratio | 2.66 ** | 0.22 ** | 0.42 ** | 0.025 ** | 0.02 ** | 0.008 ** |
| H2O2 (µmol g−1 FW) | 5.03 ** | 0.79 ** | 2.48 ** | 0.10 ** | 0.45 ** | 0.04 ** |
| MDA (µmol g−1 FW) | 4.22 ** | 0.48 ** | 1.20 ** | 0.06 ** | 0.03 ns | 0.04 ** |
| Chemical Analysis | |
| ECe | 1.64 dS m−1 |
| pHs | 7.83 |
| SAR | 12.02 |
| TSS | 45.05 m·mol L−1 |
| Na+ | 30.95 m·mol L−1 |
| K+ | 0.45 m·mol L−1 |
| Cl− | 20.88 m·mol L−1 |
| Soil organic carbon | 2.54 g kg−1 soil |
| Physical analysis | |
| Sand | 46% |
| Silt | 30% |
| Clay | 25% |
| Textual class | Sandy loam |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alamer, K.H.; Perveen, S.; Khaliq, A.; Zia Ul Haq, M.; Ibrahim, M.U.; Ijaz, B. Mitigation of Salinity Stress in Maize Seedlings by the Application of Vermicompost and Sorghum Water Extracts. Plants 2022, 11, 2548. https://doi.org/10.3390/plants11192548
Alamer KH, Perveen S, Khaliq A, Zia Ul Haq M, Ibrahim MU, Ijaz B. Mitigation of Salinity Stress in Maize Seedlings by the Application of Vermicompost and Sorghum Water Extracts. Plants. 2022; 11(19):2548. https://doi.org/10.3390/plants11192548
Chicago/Turabian StyleAlamer, Khalid H., Shaista Perveen, Abdul Khaliq, Muhammad Zia Ul Haq, Muhammad Usman Ibrahim, and Bader Ijaz. 2022. "Mitigation of Salinity Stress in Maize Seedlings by the Application of Vermicompost and Sorghum Water Extracts" Plants 11, no. 19: 2548. https://doi.org/10.3390/plants11192548
APA StyleAlamer, K. H., Perveen, S., Khaliq, A., Zia Ul Haq, M., Ibrahim, M. U., & Ijaz, B. (2022). Mitigation of Salinity Stress in Maize Seedlings by the Application of Vermicompost and Sorghum Water Extracts. Plants, 11(19), 2548. https://doi.org/10.3390/plants11192548