

Relationship between the Antioxidant Activity and Allelopathic Activities of 55 Chinese Pharmaceutical Plants

 ,
,  and
and
Abstract
1. Introduction
2. Results
2.1. Antioxidative Capacity and Total Phenolics Content of the 55 Chinese Pharmaceutical Plants
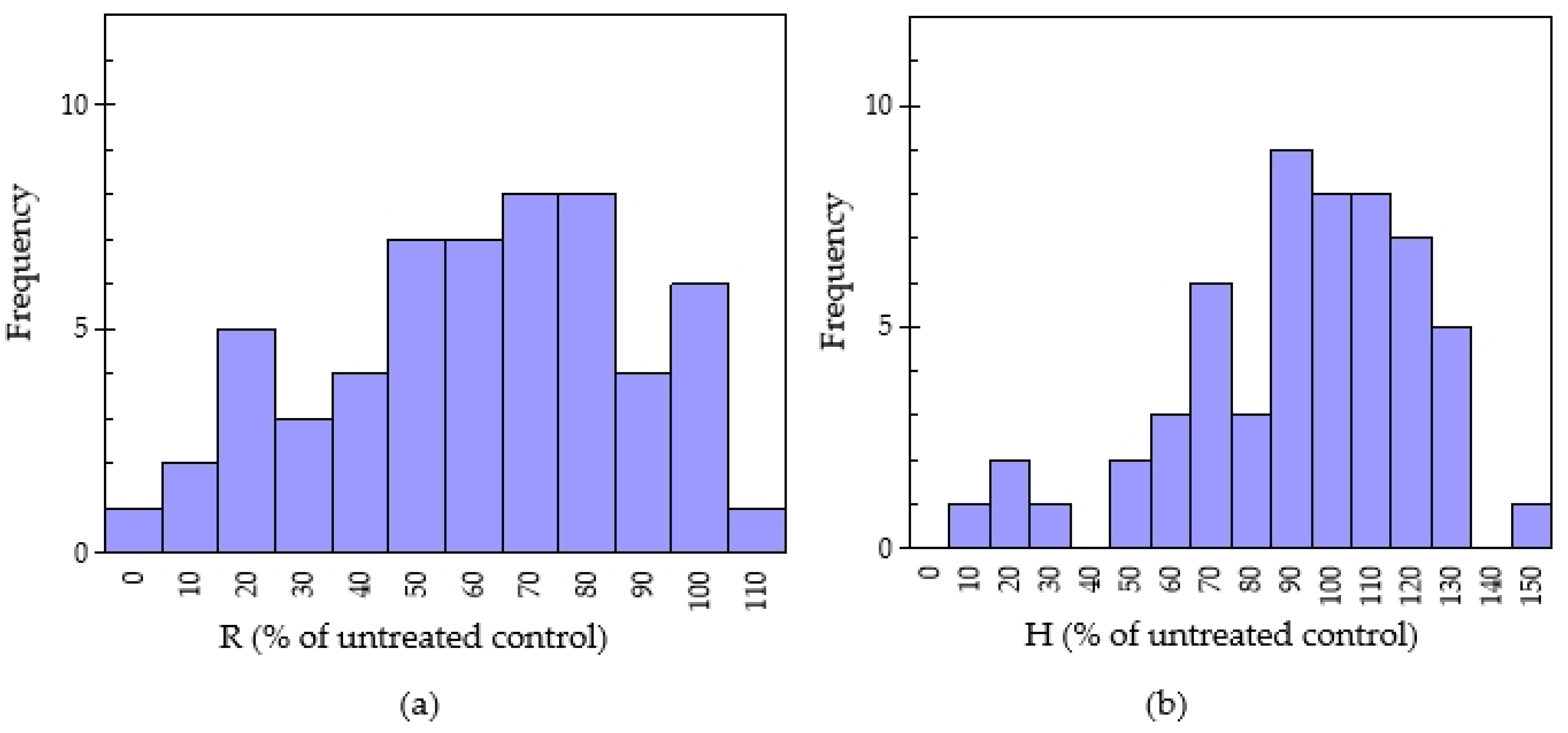
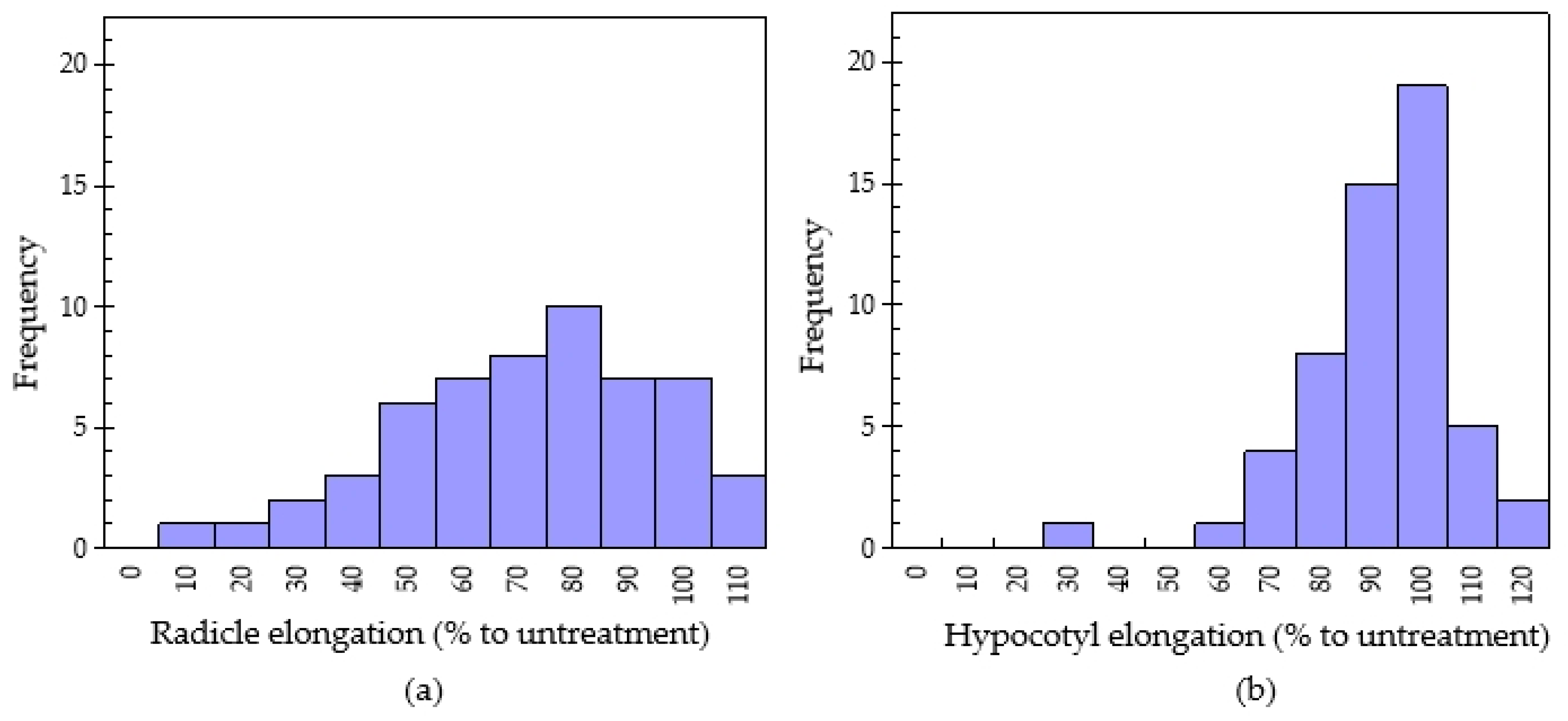
2.2. Evaluation of Allelopathic Potential from 55 Chinese Medicinal Plants
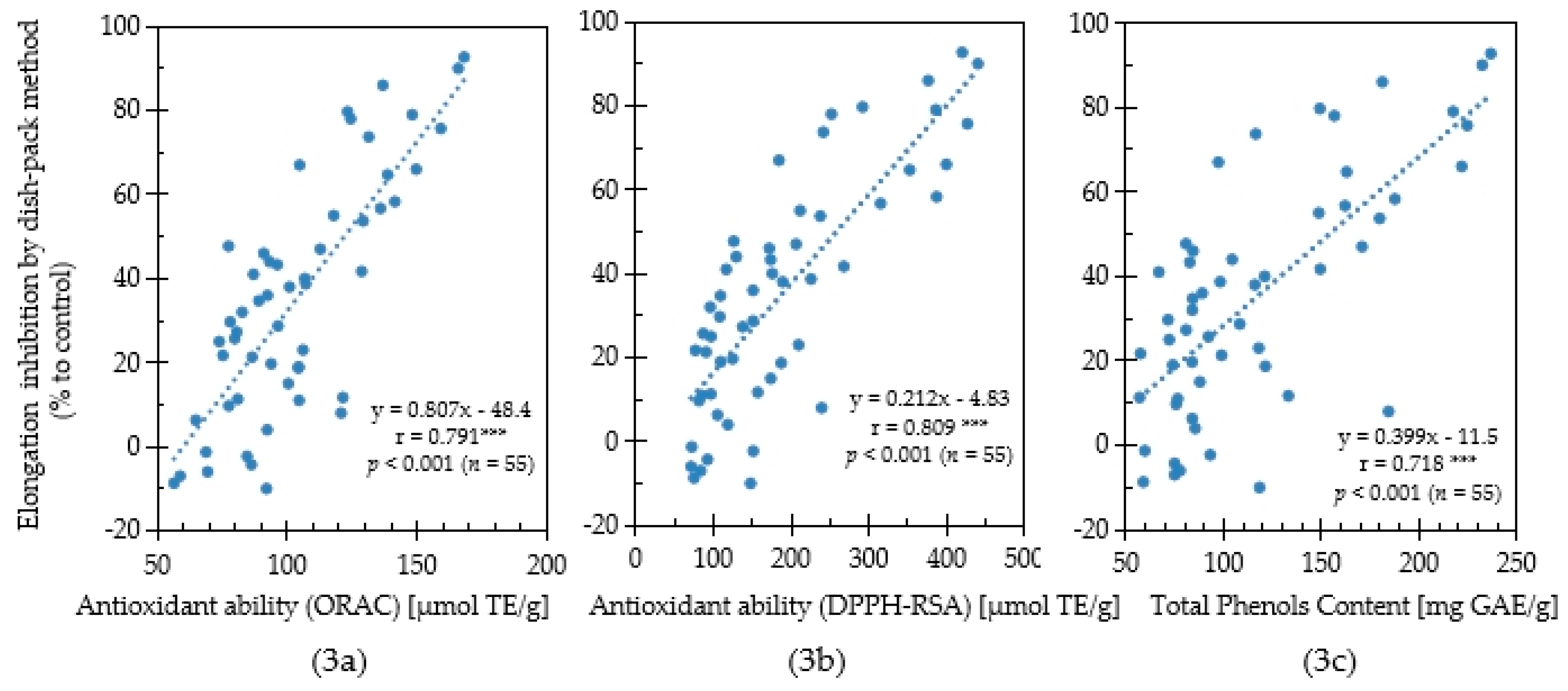
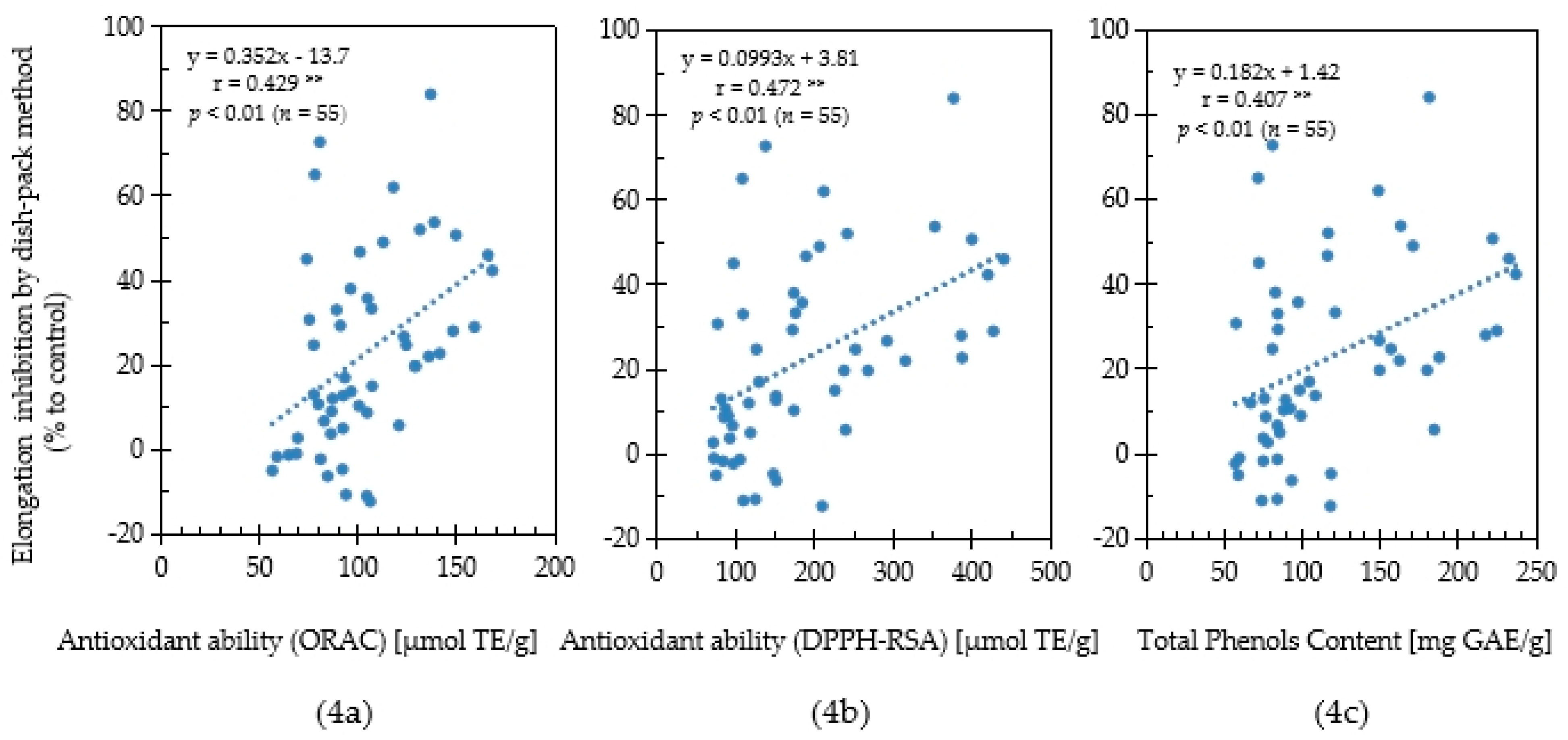
2.3. Relationship between Antioxidant Capacity and Allelopathic Activity
3. Discussion
4. Materials and Methods
4.1. Samples and Experimental Resources
4.2. ORAC, DPPH-RSA and Folin-Ciocalteu Assay
4.3. Evaluation of Allelopathy of Plant Samples through Sandwich and Dishpack Method
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ames, B.N.; Shigenaga, M.K.; Hagen, T.M. Oxidants, Antioxidant and the Degenerative Diseases of Ageing. Proc. Natl. Acad. Sci. USA 1993, 90, 7915–7922. [Google Scholar] [CrossRef]
- Washington, E.L. Allelochemicals: Role in Agriculture and Forestry. Am. Chem. Soc. 1987, 330, 9–22. [Google Scholar] [CrossRef]
- Shenoy, K.; Shrinker, A. Anti-inflammatory and Free Radical Scavenging Studies of Hypatia salvoes. Indian Drugs 2002, 39, 574–577. [Google Scholar]
- Drogue, W. Free Radicals in the Physiological Control of Cell Function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef]
- Valka, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Teiser, J. Free Radicals and Antioxidants in Normal Physiological Functions and Human Disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Fujii, Y.; Hiradate, S. Allelopathy: New Concepts and Methodology. Sci. Publ. 2007, 36, 173–183. [Google Scholar]
- Anderson, K.J.; Teuber, S.S.; Gobeille, A.; Cremin, P.; Waterhouse, A.L.; Steinburg, F.M. Walnut Polyphenolics Inhibit In vitro Human Plasma and LDL Oxidation. Biochem. Mol. Action Nutr. J. Nutr. 2001, 131, 2837–2842. [Google Scholar] [CrossRef]
- Saleem, A.; Ahotupa, M.; Pihlaja, K. Total Phenolics Concentration and Antioxidant Potential of Extracts of Medicinal Plants of Pakistan. Z. Nat. 2001, 56, 973–978. [Google Scholar] [CrossRef]
- Owens, D.K.; Nanayakkara, N.P.D.; Dayan, F.E. In planta mechanism of action of leptospermone: Impact of its physico-chemical properties on uptake, translocation, and metabolism. J. Chem. Ecol. 2013, 39, 262–270. [Google Scholar] [CrossRef]
- Sutton, P.B.; Foxon, G.A.; Beraud, J.M.; Anderdon, J.; Wichert, R. Integrated weed management systems for maize using mesotrione, nicosulfuron and acetochlor. The 1999 Brighton Conference-Weeds. Br. Crop. Prot. Counc. 1999, 24, 225–230. [Google Scholar]
- Armel, G.R.; Wilson, H.P.; Richardson, R.J. Mesotrione alone and in mixtures with glyphosate in glyphosate-resistant corn (Zea mays). Weed Technol. 2003, 17, 680–685. [Google Scholar] [CrossRef]
- Nazir, T.; Uniyal, A.K.; Todari, N.P. Allelopathic Behaviour of Three Medicinal Plant Species on Traditional Agriculture Crops of Garhwal Himalaya, India. Agroforest Syst. 2007, 69, 183–187. [Google Scholar] [CrossRef]
- Huang, J.H.; Chen, B.; Liu, C.R.; Lai, J.S.; Zhang, J.L.; Ma, K.P. Identifying Hotspots of Endemic Woody Seed Plant Diversity in China. Divers. Distrib. 2012, 18, 673–688. [Google Scholar] [CrossRef]
- Dixon, R.A.; Achnine, L.; Kota, P. The phenylpropanoid pathway and plant defence-a genomics perspective. Mol. Plant Pathol. 2002, 3, 371–390. [Google Scholar] [CrossRef]
- Hartmann, T. From waste products to eco chemicals: Fifty years research of plant secondary metabolism. Phytochemistry 2007, 68, 2831–2846. [Google Scholar] [CrossRef]
- Jacob, J.; Sarada, S. Role of phenolics in allelopathic interactions. Allelopath. J. 2012, 29, 215–230. [Google Scholar] [CrossRef]
- Appiah, K.S.; Mardani, H.K.; Osivand, A.; Kpabitey, S.; Amoatey, C.A.; Oikawa, Y.; Fujii, Y. Exploring Alternative Use of Medicinal Plants for Sustainable Weed Management. Sustainability 2017, 9, 1468. [Google Scholar] [CrossRef]
- Kumar, M.S.Y.; Dutta, R.; Prasad, D.; Misra, K. Subcritical water extraction of antioxidant compounds from Seabuckthorn (Hippophae rhamnoides) leaves for the comparative evaluation of antioxidant activity. Food Chem. 2011, 127, 1309–1316. [Google Scholar] [CrossRef]
- Su, L.; Yin, J.J.; Charles, D.; Zhou, K.; Moore, J.; Yu, L. Total phenolic contents, chelating capacities, and radical-scavenging properties of black peppercorn, nutmeg, rosehip, cinnamon and oregano leaf. Food Chem. 2007, 100, 990–997. [Google Scholar] [CrossRef]
- Sepulveda, E.; Sáenz, C.; Peña, Á.; Robert, P.; Bartolomé, B.; Gómez, C. Influence of the genotype on the anthocyanin composition, antioxidant capacity and color of Chilean pomegranate (Punica granatum L.) juices. Chil. J. Agric. Res. 2010, 70, 50. [Google Scholar] [CrossRef]
- Moyer, R.A.; Hummer, K.E.; Finn, C.E.; Frei, B.; Wrolstad, R.E. Anthocyanins, Phenolics, and Antioxidant Capacity in Diverse Small Fruits: Vaccinium, Rubus, and Ribes. Agric. Food Chem. 2002, 50, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Suryakumar, G.; Gupta, A. Medicinal and Therapeutic Potential of Sea Buckthorn (Hippophae rhamnoides L.). J. Ethnopharmacol. 2011, 138, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, N.K.; Kumar, Y.; Gupta, A. Antioxidant, Cytoprotective and Antibacterial Effects of Sea Buckthorn (Hippophae rhamnoides L.) leaves. Food Chem. Toxicol. 2010, 48, 3443–3448. [Google Scholar] [CrossRef] [PubMed]
- Heo, H.; Kim, Y.; Chung, D.; Kim, D. Antioxidant Capacities of Individual and Combined Phenolics in a Model System. Food Chem. 2007, 104, 87–92. [Google Scholar] [CrossRef]
- Bao, L.; Bai, M.; Douqinfu, A.D.; Fujii, Y. Screening of Allelopathic Activity of Plants Growing in the Desert Area of Inner Mongolia Autonomous Region and discovery of Sea Buckthorn and Sweet Annie as Highly Active plants and Estimation of Their Active Substances. J. Arid Land Stud. 2019, 29, 1–10. [Google Scholar] [CrossRef]
- Shu, X.; Liu, X.M.; Fu, C.L.; Liang, Q.X. Extraction, Characterization and Antitumor Effect of the Polysaccharides from Star Anise (Illicium verum Hook. f). J. Med. Plant Res. 2010, 4, 2666–2673. [Google Scholar] [CrossRef]
- Chugh, C.A.; Mehta, S.; Dua, H. Phytochemical Screening and Evaluation of Biological Activities of Some Medicinal Plants of Phagwara, Punjab. Asian J. Chem. 2012, 12, 5903–5905. [Google Scholar]
- Sekine, T.; Appiah, K.S.; Azizi, M.; Fujii, Y. Plant Growth Inhibitory Activities and Volatile Active Compounds of 53 Spices and Herbs. Plants 2020, 9, 264. [Google Scholar] [CrossRef]
- Kang, G.W.; Mishyna, M.; Appiah, K.S.; Yamada, M.; Takano, A.; Prokhorov, V.; Fujii, Y. Screening for Plant Volatile Emissions with Allelopathic Activity and the Identification of L-Fenchone and 1,8-Cineole from Star Anise (Illicium verum) Leaves. Plants 2019, 8, 457. [Google Scholar] [CrossRef]
- Singh, G.; Maurya, S.; Lampason, M.P.; Catalan, C. Chemical constituent, antimicrobial investigations and antioxidative potential of volatile oil and acetone extract of star anise fruits. J. Sci. Food Agric. 2006, 86, 111–121. [Google Scholar] [CrossRef]
- Mungole, A.; Day, S.; Kamble, R.; Kanfade, H.; Chaturvedi, A.; Zanwar, P. Active Phytochemical and Antibacterial Potentiality of In-vitro Regenerated Plantlets of Canscora Decurrens (Dalzell). Indian J. Sci. Technol. 2010, 6, 679–683. [Google Scholar] [CrossRef]
- Singh, H.P.; Batish, D.R.; Kohli, R.K. Allelopathic Interactions and Allelochemicals: New Possibilities for Sustainable Weed Management. Crit. Rev. Plant Sci. 2003, 22, 239–311. [Google Scholar] [CrossRef]
- Krasowska, A.; Rosiak, D.; Szkapiak, K.; Oswiecimska, M.; Witek, S.; Lukas, Z.M. The Antioxidant Activity of BHT and New Phenolic Compounds PYA and PPA Measured by Chemiluminescence. Cell. Mol. Biol. Lett. 2001, 6, 71–81. [Google Scholar] [CrossRef]
- Aniya; Nomura, Y.; Fuerdeng; Appiah, K.S.; Fujii, Y. Evaluation of Allelopathic Activity of Chinese Medicinal Plants and Identification of Shikimic Acid as an Allelochemical from Illicium verum Hook. f. Plants 2020, 9, 684. [Google Scholar] [CrossRef] [PubMed]
- Golisz, A.; Lata, B.; Gawronski, S.W.; Fujii, Y. Specific and total activities of the allelochemicals identified in buckwheat. Weed Biol. Manag. 2007, 7, 164–171. [Google Scholar] [CrossRef]
- Wei, J.; Sha, L.; Tingting, S.; Jinmei, Z.; Yumei, J.; Yury, A.Z.; Yang, B. Phenolic compositions and antioxidant activities of Hippophae tibetana and H. rhamnoides ssp. sinensis berries produced in Qinghai-Tibet Plateau. Food Chem. X 2022, 15, 1–10. [Google Scholar] [CrossRef]
- Nakajima, N.; Hiradate, S.; Fujii, Y. Characteristics of Growth Inhibitory Effect of L-3,4-Dihydroxyphenylalanine (L-DOPA) on Cucumber Seedlings. Weed Res. 1999, 44, 132–138. [Google Scholar] [CrossRef]
- Ou, B.; Hampsch, W.M.; Prior, R.L. Development and Validation of an Improved Oxygen Radical Absorbance Capacity Assay Using Fluorescein as the Fluorescent Probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef]
- Watanabe, J.; Oki, T.; Takebayashi, J.; Yamasaki, K.; Takano-Ishikawa, Y.; Hino, A.; Yasui, A. Method Validation by Inter-laboratory Studies of Improved Hydrophilic Oxygen Radical Absorbance Capacity Methods for the Determination of Antioxidant Capacities of Antioxidant Solutions and Food Extracts. Anal. Sci. 2012, 28, 159–165. [Google Scholar] [CrossRef]
- Burda, S.; Oleszek, W. Antioxidant and Antiradical Activities of Flavonoids. J. Agric. Food Chem. 2001, 49, 2774–2779. [Google Scholar] [CrossRef]
- Choi, H.S.; Song, H.S.; Ukeda, H. Radical scavenging activities of citrus essential oils and their components: Detection using 1,1-diphenyl-2-picrylhydrazyl. J. Agric. Food Chem. 2000, 48, 4156–4161. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Fujii, Y.; Shibuya, T.; Nakatani, K.; Itani, T.; Hiradate, S.; Parvez, M.M. Assessment Method for Allelopathic from Leaf Litter Leachates. Weed Biol. Manag. 2004, 4, 19–23. [Google Scholar] [CrossRef]
- Fujii, Y.; Parvez, S.S.; Parvez, M.M.; Ohmae, Y.; Iida, O. Screening of 239 Medicinal Plant Species for Allelopathic Activity Using the Sandwich Method. Weed Biol. Manag. 2003, 3, 233–241. [Google Scholar] [CrossRef]
- Amini, S.; Azizi, M.; Joharchi, M.R.; Shafei, M.N.; Moradinezhad, F.; Fujii, Y. Determination of Allelopathic Potential in Some Medicinal and Wild Plant Species of Iran by Dish Pack Method. Theor. Exp. Plant Physiol. 2014, 26, 189–199. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Name | Plant Part Used | ORAC (μmol TE/g) | DPPH-RSA (μmol TE/g) | Total Phenolic Contents (mg GAE/g) |
|---|---|---|---|---|
| Hippophae rhamnoides L. | fruit | 168 ± 7.04 | 420 ± 7.15 | 236 ± 7.62 |
| Hippophae rhamnoides L. | leaf | 166 ± 6.05 | 440 ± 7.32 | 232 ± 7.40 |
| Rosa multiflora Thunb. | fruit | 159 ± 5.38 | 426 ± 7.02 | 224 ± 7.51 |
| Punica granatum L. | peel | 149 ± 5.62 | 399 ± 6.40 | 221 ± 6.48 |
| Ribes nigrum L. | fruit | 148 ± 5.23 | 386 ± 5.17 | 217 ± 7.33 |
| Lycium ruthenicum Murray | fruit | 141 ± 5.01 | 387 ± 5.21 | 187 ± 6.04 |
| Capsicum annuum L. | fruit | 138 ± 5.21 | 352 ± 4.58 | 162 ± 4.01 |
| Illicium verum Hook. f. | fruit | 136 ± 4.20 | 376 ± 5.04 | 181 ± 6.20 |
| Physalis alkekengi L. | fruit | 135 ± 3.80 | 315 ± 4.20 | 162 ± 3.98 |
| Nitraria tangutorum Bobr. | leaf | 131 ± 4.05 | 241 ± 3.80 | 116 ± 2.75 |
| Rubus idaeus L. | fruit | 129 ± 3.52 | 237 ± 3.63 | 179 ± 5.31 |
| Cuminum cyminum Linn. | fruit | 125 ± 3.74 | 243 ± 2.15 | 162 ± 1.52 |
| Stachys geobombycis C. Y. Wu | root | 125 ± 2.10 | 249 ± 1.81 | 164 ± 2.30 |
| Gossypium herbaceum L. | root | 122 ± 2.15 | 290 ± 2.40 | 145 ± 2.13 |
| Adenia chevalieri Gagnep | fruit | 121 ± 3.22 | 157 ± 2.78 | 133 ± 3.44 |
| Dendrobium nobile Lindl. | fruit | 120 ± 3.10 | 239 ± 3.35 | 184 ± 4.86 |
| Indigofera tinctoria L. | leaf | 117 ± 2.18 | 211 ± 3.21 | 148 ± 3.67 |
| Schisandra chinensis Baill. | fruit | 112 ± 2.12 | 206 ± 3.10 | 170 ± 4.53 |
| Calophyllum inophyllum L. | fruit | 110 ± 1.67 | 208 ± 4.11 | 104 ± 2.25 |
| Perilla frutescens Britt. | leaf | 107 ± 0.94 | 217 ± 1.51 | 110 ± 1.40 |
| Ziziphus jujuba Mill. | fruit | 105 ± 0.51 | 200 ± 1.75 | 113 ± 1.30 |
| Chenopodium glaucum L. | leaf | 104 ± 1.73 | 184 ± 2.33 | 97.3 ± 2.07 |
| Heracleum scabridum Franch. | root | 104 ± 2.03 | 85.3 ± 1.60 | 76.4 ± 0.75 |
| Cicer arietinum L. | fruit | 104 ± 1.62 | 109 ± 2.46 | 74.0 ± 0.42 |
| Adenophora stricta Miq. | fruit | 104 ± 1.28 | 187 ± 2.35 | 121 ± 3.27 |
| Lonicera dasystyla Rehd. | root | 103 ± 2.30 | 185 ± 1.40 | 112 ± 1.80 |
| Paeonia lactiflora Pall. | fruit | 100 ± 1.37 | 173 ± 2.26 | 87.5 ± 2.05 |
| Aster tataricus Linn. | leaf | 98.4 ± 0.85 | 140 ± 1.60 | 93.0 ± 1.75 |
| Tribulus terrester Linn. | fruit | 95.4 ± 0.70 | 166 ± 3.01 | 102 ± 2.04 |
| Ziziphus jujuba Mill. | fruit | 93.8 ± 0.92 | 124 ± 1.52 | 83.5 ± 1.23 |
| Plantago asiatica Linn. | leaf | 93.7 ± 1.03 | 139 ± 1.30 | 94.7 ± 1.44 |
| Eriobotrya japonica Lindl. | fruit | 93.5 ± 2.41 | 151 ± 0.97 | 95.3 ± 1.37 |
| Munronia sinica Diels | root | 92.3 ± 0.68 | 118 ± 1.83 | 85.4 ± 1.90 |
| Morus macroura Miq. | fruit | 92.0 ± 0.77 | 147 ± 1.64 | 118 ± 2.90 |
| Aralia chinensis L. | root | 90.7 ± 0.55 | 171 ± 1.75 | 84.3 ± 1.82 |
| Elaeagnus angustifolia Linn. | fruit | 90.6 ± 1.2 | 100 ± 2.40 | 106 ± 1.51 |
| Dendranthema indicum Moul | leaf | 90.5 ± 2.10 | 110 ± 0.85 | 92.4 ± 1.32 |
| Morus alba Linn. | root | 88.2 ± 1.30 | 100 ± 0.82 | 73.4 ± 1.60 |
| Phyllanthus emblica L. | fruit | 86.5 ± 0.48 | 90.6 ± 1.21 | 98.7 ± 2.30 |
| Arnebia euchroma Johnst. | leaf | 86.0 ± 0.45 | 92.4 ± 1.03 | 74.8 ± 0.91 |
| Amaranthus viridis L. | leaf | 84.5 ± 0.37 | 151 ± 1.32 | 93.1 ± 2.13 |
| Armeniaca vulgaris Lam. | fruit | 82.7 ± 0.90 | 94.2 ± 1.41 | 87.5 ± 1.73 |
| Fritillaria walujewii Regel | fruit | 81.0 ± 0.27 | 96.7 ± 0.97 | 56.3 ± 0.26 |
| Angelica sinensis Diels | root | 80.7 ± 1.50 | 102 ± 2.10 | 83.4 ± 2.03 |
| Pueraria lobata Ohwi | root | 80.3 ± 1.73 | 90.6 ± 0.91 | 97.5 ± 4.23 |
| Rheum officinale Baill. | root | 79.3 ± 0.70 | 102 ± 0.91 | 79.3 ± 1.43 |
| Tulipa iliensis Regel | leaf | 77.4 ± 0.26 | 81.5 ± 0.80 | 75.6 ± 0.87 |
| Salix sinopurpurea C. Wang | fruit | 77.4 ± 0.22 | 126 ± 0.76 | 80.3 ± 1.04 |
| Trigonella foenum-graecum L. | fruit | 75.2 ± 0.30 | 76.9 ± 0.72 | 57.0 ± 0.41 |
| Dimocarpus longan L. | fruit | 75.0 ± 0.27 | 101 ± 0.51 | 80.0 ± 1.03 |
| Sinapis alba L. | fruit | 69.3 ± 0.13 | 71.3 ± 0.55 | 77.6 ± 0.95 |
| Senecio scandens Buch. | fruit | 68.7 ± 0.25 | 72.2 ± 0.64 | 59.6 ± 0.47 |
| Equisetum hyemale L. | leaf | 64.8 ± 0.14 | 105 ± 0.72 | 83.7 ± 1.64 |
| Armeniaca vulgaris Lam | leaf | 58.8 ± 0.17 | 83.7 ± 0.42 | 74.7 ± 0.80 |
| Carya cathayensis Sarg | leaf | 56.3 ± 0.12 | 75.1 ± 0.38 | 58.3 ± 0.30 |
| Scientific Name | Plant Part Used | Radicle Growth by Sandwich Method | Radicle Growth by Dishpack Method | ||
|---|---|---|---|---|---|
| (% to Control) | Criteria * | (% to Control) | Criteria * | ||
| Hippophae rhamnoides L. | fruit | 7.30 ± 0.35 | *** | 57.7 ± 2.51 | * |
| Hippophae rhamnoides L. | leaf | 10.0 ± 0.52 | *** | 54.0 ± 1.75 | * |
| Illicium verum Hook. f. | fruit | 14.0 ± 0.48 | *** | 16.0 ± 0.14 | *** |
| Gossypium herbaceum L. | root | 19.3 ± 0.21 | *** | 78.3 ± 1.03 | |
| Ribes nigrum L. | fruit | 21.0 ± 0.70 | *** | 72.0 ± 4.38 | |
| Stachys geobombycis C. Y. Wu | root | 22.3 ± 0.65 | *** | 75.3 ± 4.71 | |
| Rosa multiflora Thunb. | fruit | 24.3 ± 0.73 | ** | 71.0 ± 4.13 | |
| Nitraria tangutorum Bobr. | leaf | 26.3 ± 0.78 | ** | 48.0 ± 1.28 | ** |
| Chenopodium glaucum L. | leaf | 33.0 ± 0.83 | ** | 64.3 ± 2.30 | * |
| Punica granatum L. | peel | 34.0 ± 0.81 | ** | 49.3 ± 1.86 | ** |
| Capsicum annuum L. | fruit | 35.3 ± 0.90 | ** | 46.3 ± 1.90 | ** |
| Lycium ruthenicum Murray | fruit | 41.7 ± 0.95 | * | 77.3 ± 4.26 | |
| Physalis alkekengi L. | fruit | 43.3 ± 1.03 | * | 78.0 ± 3.97 | |
| Indigofera tinctoria L. | leaf | 45.0 ± 1.20 | * | 38.0 ± 0.82 | *** |
| Rubus idaeus L. | fruit | 46.3 ± 1.08 | * | 80.3 ± 4.66 | |
| Salix sinopurpurea C. Wang | leaf | 52.3 ± 1.27 | 75.3 ± 5.10 | ||
| Schisandra chinensis Baill. | fruit | 53.0 ± 1.30 | 51.0 ± 1.90 | ** | |
| Aralia chinensis L. | root | 54.0 ± 1.42 | 70.7 ± 4.40 | ||
| Plantago asiatica Linn. | leaf | 56.3 ± 1.10 | 84.0 ± 2.10 | ||
| Tribulus terrester Linn. | fruit | 57.7 ± 0.81 | 63.0 ± 2.03 | * | |
| Cuminum cyminum Linn. | fruit | 58.7 ± 1.47 | 80.1 ± 4.11 | ||
| Morus alba Linn. | root | 59.3 ± 2.03 | 82.3 ± 3.50 | ||
| Perilla frutescens Britt. | leaf | 61.0 ± 3.70 | 65.7 ± 1.53 | ||
| Calophyllum inophyllum L. | fruit | 61.7 ± 2.10 | 88.0 ± 5.42 | ||
| Lonicera dasystyla Rehd. | root | 62.3 ± 2.70 | 56.1 ± 3.01 | ** | |
| Eriobotrya japonica Lindl. | fruit | 65.3 ± 1.80 | 89.0 ± 0.73 | ||
| Elaeagnus angustifolia Linn. | fruit | 66.7 ± 2.70 | 65.0 ± 1.53 | ||
| Dendranthema indicum Moul. | fruit | 67.0 ± 1.10 | 69.1 ± 1.81 | ||
| Armeniaca vulgaris Lam. | fruit | 70.0 ± 2.30 | 98.5 ± 4.30 | ||
| Rheum officinale Baill. | root | 70.7 ± 1.85 | 36.2 ± 1.13 | *** | |
| Aster tataricus Linn. | leaf | 71.0 ± 1.60 | 85.2 ± 3.23 | ||
| Angelica sinensis Diels | root | 71.7 ± 3.24 | 29.1 ± 0.81 | *** | |
| Pueraria lobata Ohwi | root | 74.7 ± 2.51 | 87.3 ± 4.15 | ||
| Dimocarpus longan L. | fruit | 76.7 ± 3.20 | 56.0 ± 1.70 | * | |
| Solanum nigrum Linn. | fruit | 78.0 ± 4.05 | 112 ± 10.5 | ||
| Trigonella foenum-graecum L. | fruit | 78.3 ± 5.20 | 69.3 ± 3.34 | ||
| Phyllanthus emblica L. | fruit | 78.7 ± 5.35 | 91.0 ± 6.55 | ||
| Ziziphus jujuba Mill. | fruit | 80.3 ± 6.24 | 110 ± 10.4 | ||
| Adenophora stricta Miq. | fruit | 81.3 ± 7.40 | 58.3 ± 2.62 | * | |
| Cicer arietinum L. | fruit | 81.0 ± 6.58 | 108 ± 9.22 | ||
| Paeonia lactiflora Pall. | fruit | 85.0 ± 6.93 | 89.7 ± 6.90 | ||
| Adenia chevalieri Gagnep. | root | 87.0 ± 7.56 | 69.0 ± 3.72 | ||
| Fritillaria walujewii Regel. | fruit | 88.3 ± 8.03 | 102 ± 3.95 | ||
| Heracleum scabridum Franch. | root | 88.7 ± 7.78 | 91.3 ± 7.31 | ||
| Tulipa iliensis Regel | root | 89.0 ± 8.31 | 87.0 ± 6.70 | ||
| Dendrobium nobile Lindl. | fruit | 90.3 ± 9.04 | 94.3 ± 8.11 | ||
| Equisetum hyemale L. | fruit | 92.0 ± 8.61 | 101 ± 6.45 | ||
| Munronia sinica Diels | fruit | 93.7 ± 9.30 | 95.0 ± 7.44 | ||
| Senecio scandens Buch.-Ham. | root | 96.0 ± 9.22 | 101 ± 3.02 | ||
| Amaranthus viridis L. | fruit | 101 ± 9.23 | 106 ± 9.63 | ||
| Arnebia euchroma Johnst. | leaf | 102 ± 9.64 | 96.3 ± 7.80 | ||
| Sinapis alba L. | leaf | 104 ± 9.70 | 97.3 ± 8.90 | ||
| Armeniaca vulgaris Lam. | fruit | 106 ± 10.0 | 101 ± 8.86 | ||
| Carya cathayensis Sarg. | leaf | 107 ± 9.85 | 105 ± 10.2 | ||
| Morus macroura Miq. | leaf | 108 ± 10.10 | 104 ± 9.31 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aniya; Nomura, Y.; Appiah, K.S.; Fuerdeng; Suzuki, Y.; Fujii, Y.; Xia, Q. Relationship between the Antioxidant Activity and Allelopathic Activities of 55 Chinese Pharmaceutical Plants. Plants 2022, 11, 2481. https://doi.org/10.3390/plants11192481
Aniya, Nomura Y, Appiah KS, Fuerdeng, Suzuki Y, Fujii Y, Xia Q. Relationship between the Antioxidant Activity and Allelopathic Activities of 55 Chinese Pharmaceutical Plants. Plants. 2022; 11(19):2481. https://doi.org/10.3390/plants11192481
Chicago/Turabian StyleAniya, Yoshihiro Nomura, Kwame Sarpong Appiah, Fuerdeng, Yoko Suzuki, Yoshiharu Fujii, and Qile Xia. 2022. "Relationship between the Antioxidant Activity and Allelopathic Activities of 55 Chinese Pharmaceutical Plants" Plants 11, no. 19: 2481. https://doi.org/10.3390/plants11192481
APA StyleAniya, Nomura, Y., Appiah, K. S., Fuerdeng, Suzuki, Y., Fujii, Y., & Xia, Q. (2022). Relationship between the Antioxidant Activity and Allelopathic Activities of 55 Chinese Pharmaceutical Plants. Plants, 11(19), 2481. https://doi.org/10.3390/plants11192481