
Genetic Characterization of Spring Wheat Germplasm for Macro-, Microelements and Trace Metals

 and
and
Abstract
:1. Introduction
2. Results
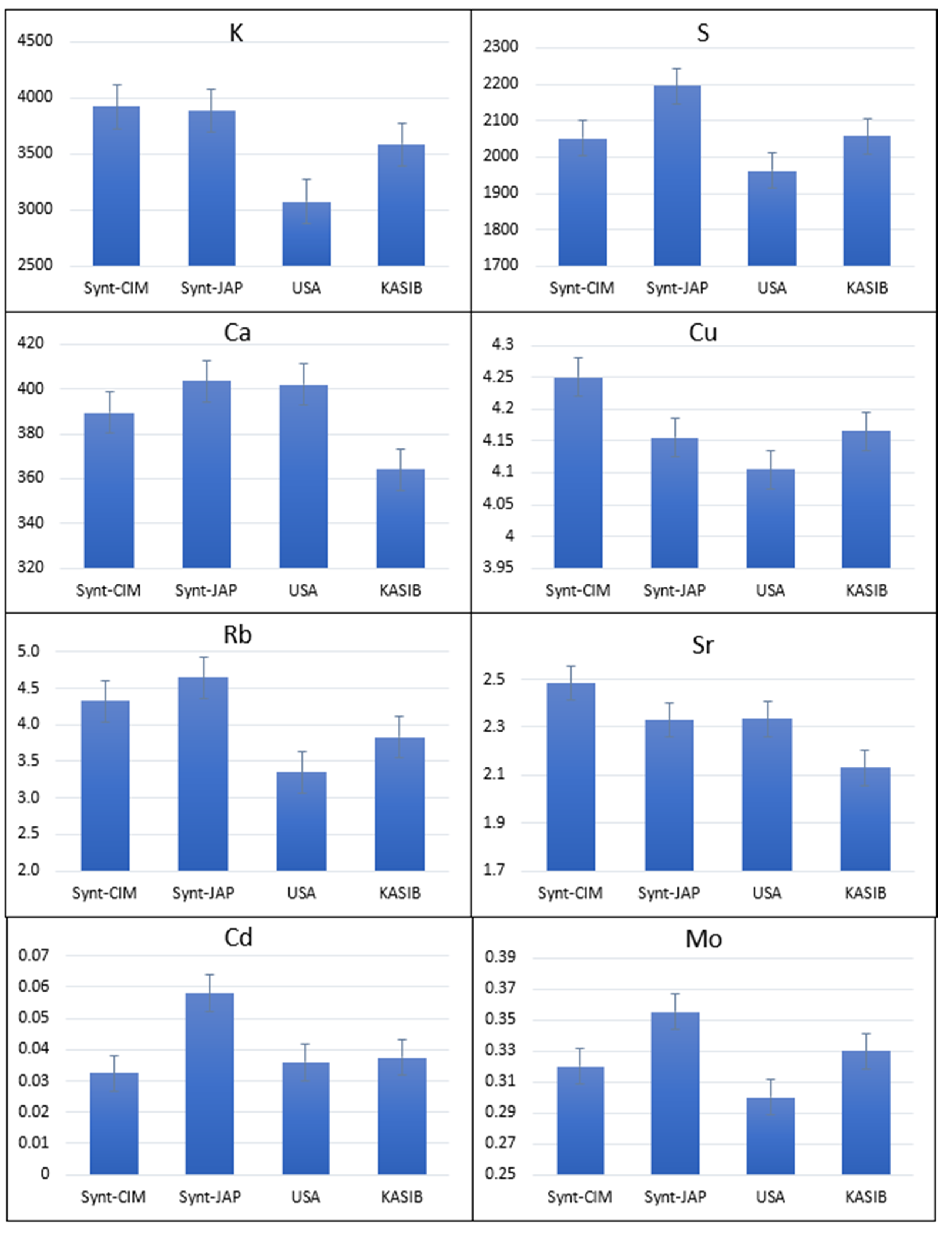
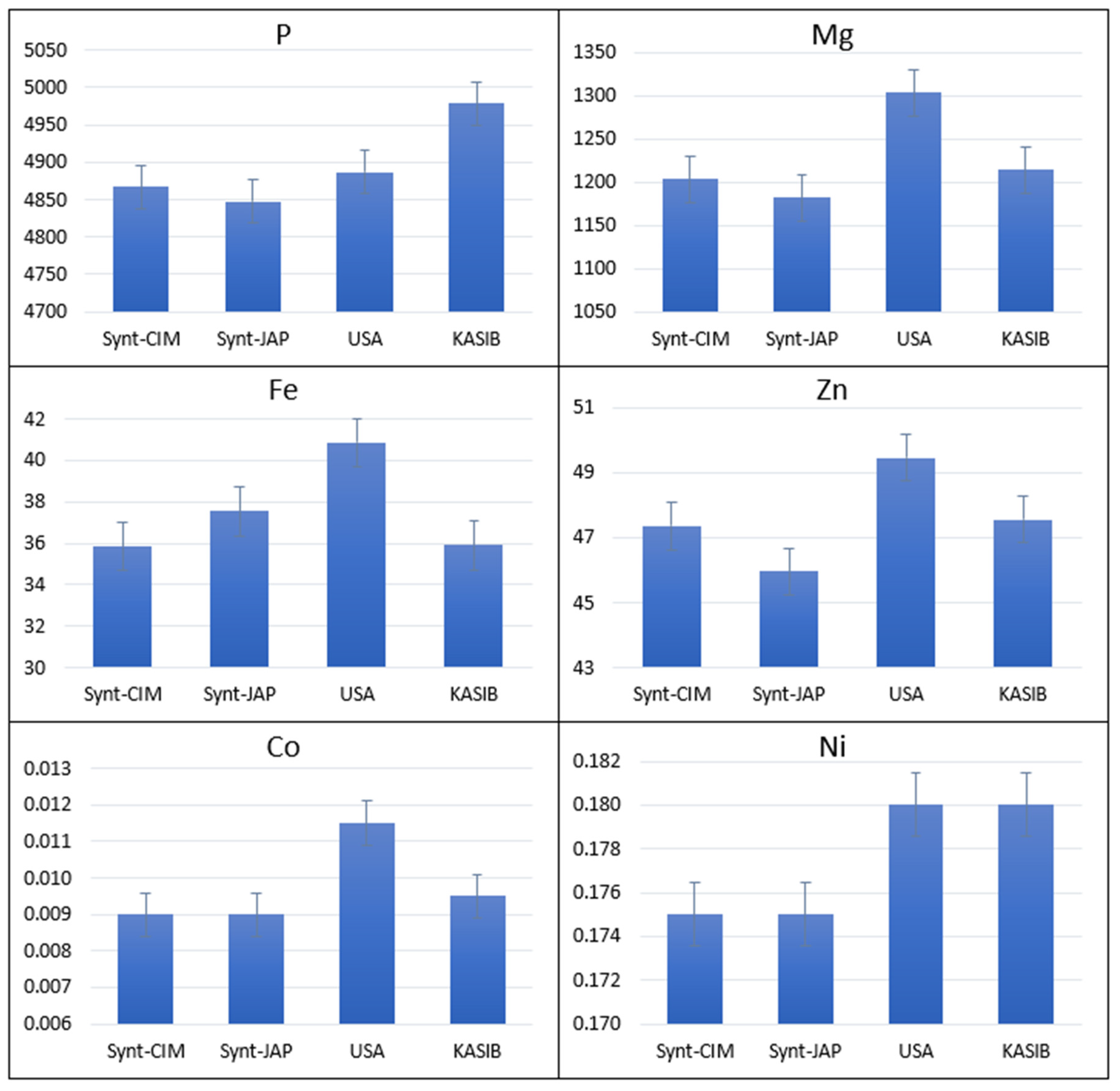
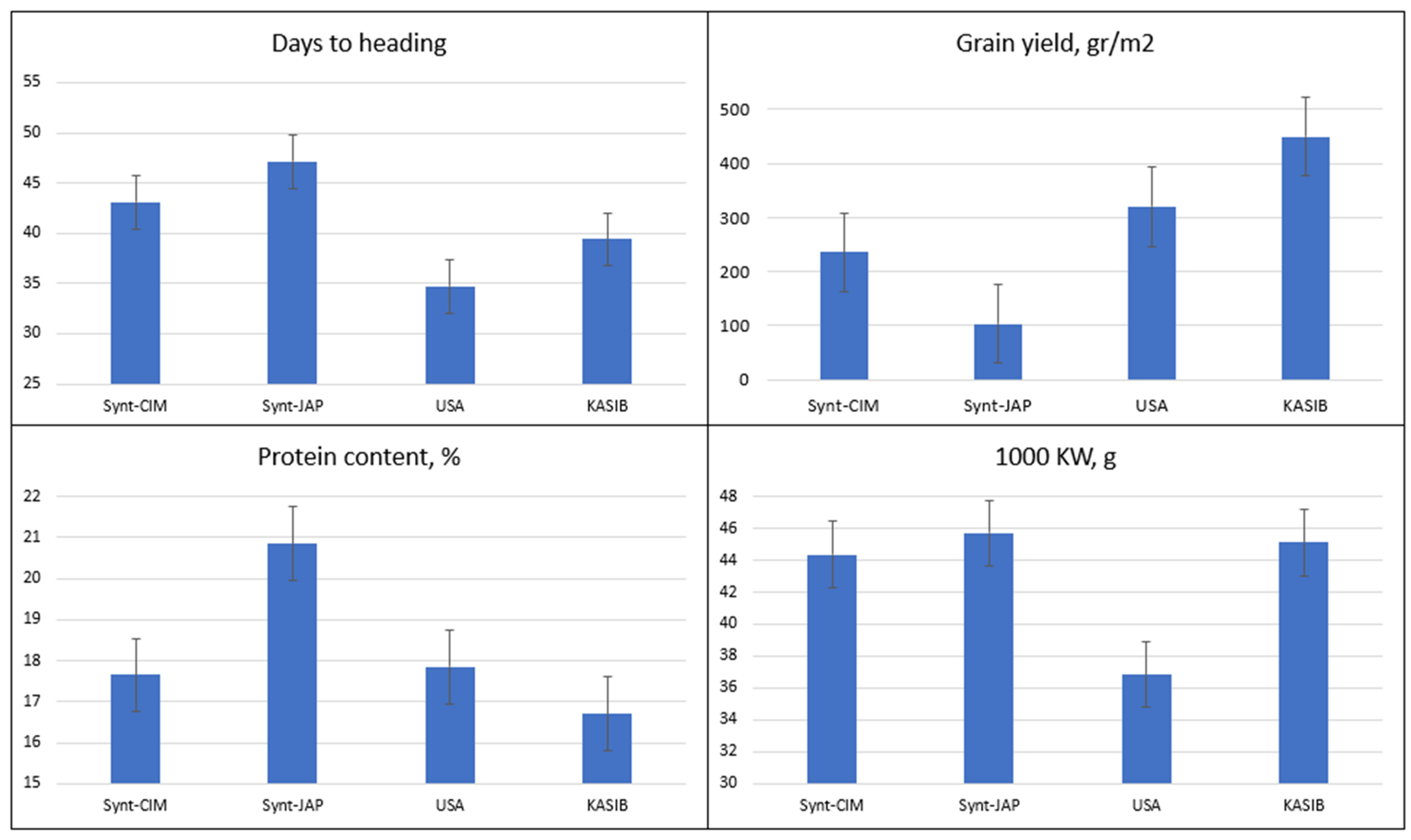
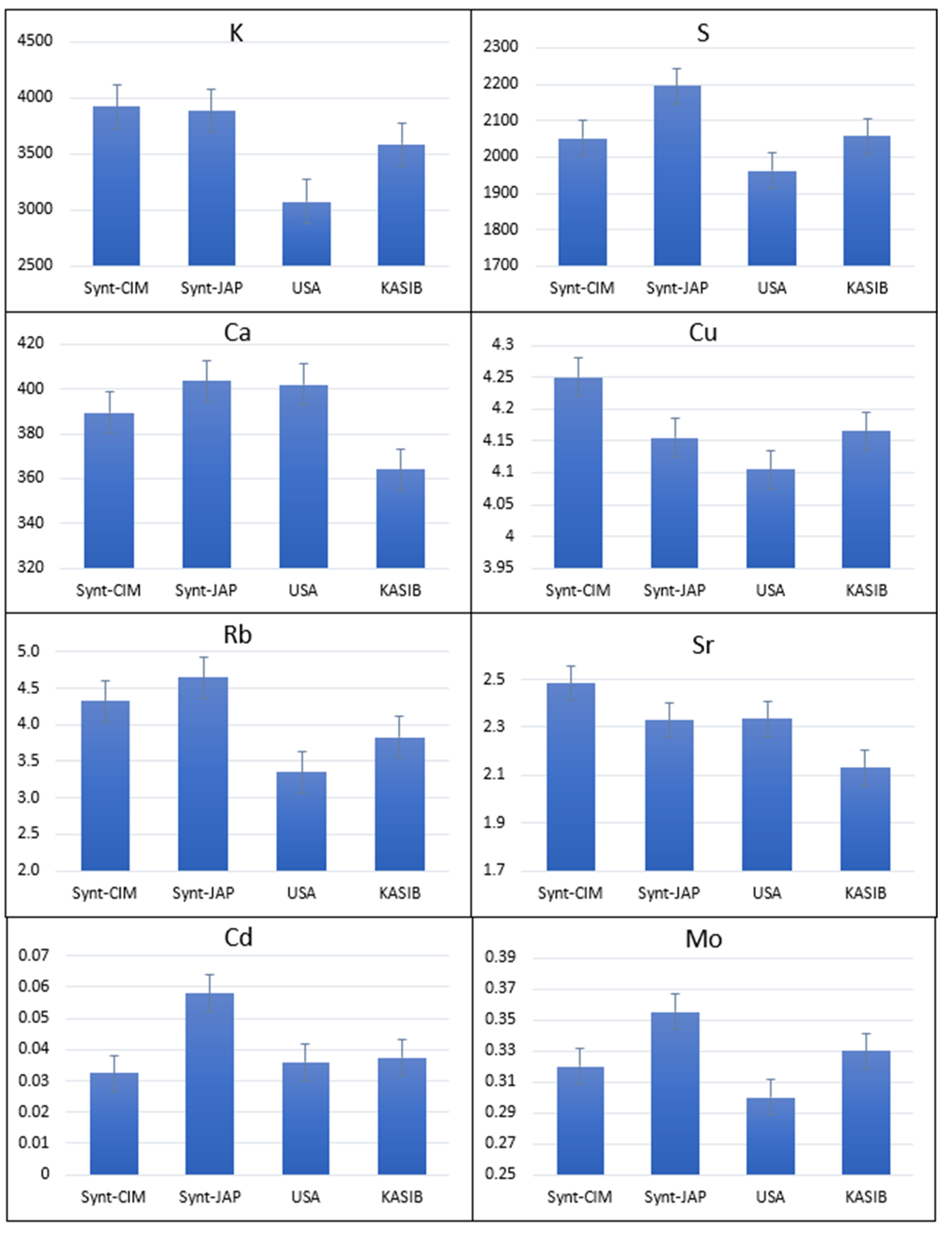
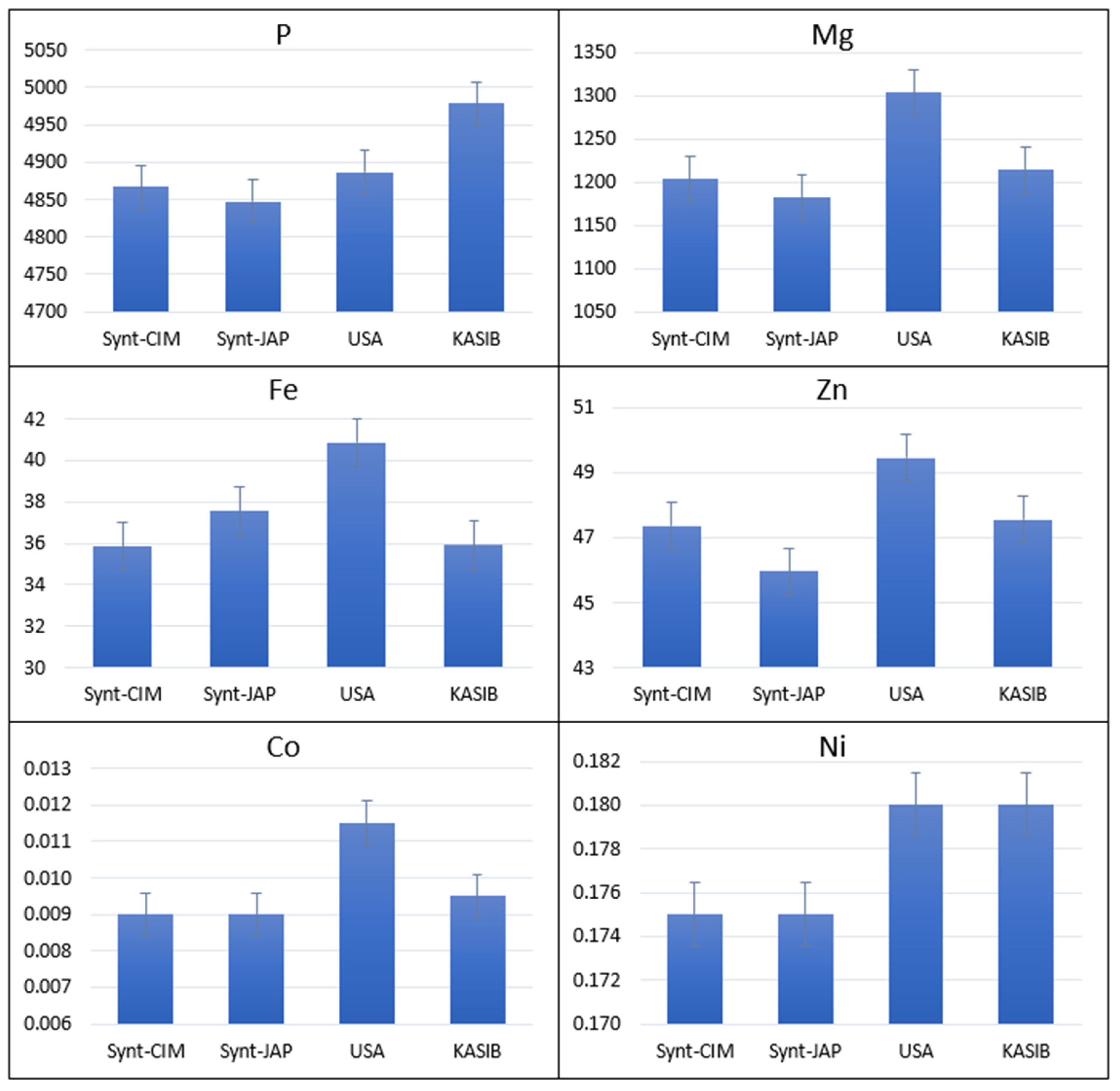
2.1. Agronomic Performance and Elemental Composition of Different Groups of Germplasm in GDP
2.2. Results from GWAS Analysis
2.3. Agronomic Performance and Elemental Composition of KASIB Validation Trial
2.4. Validated SNPs Affecting Elemental Concentration
2.5. Annotation of SNPs Contributing to Wheat Grain Elemental Composition
2.6. Distribution of SNP Markers among the Germplasm Groups
2.7. Superior Germplasm with Optimal Combination of Elements
3. Discussion
3.1. Elements and Underlying Physiology and Biochemistry
3.2. Differences between the Elements on Number of SNPs
3.3. Relation to SNPs Identified in Other Publications
3.4. Breeding Approaches Based on the Study Results
4. Materials and Methods
4.1. GWAS Genetic Diversity Panel Material, Field Experiments
4.2. Validation Panel KASIB Trial Material, Field Experiments
4.3. Grain Ionomics Analysis, Values Adjustment and Analysis of Variance
4.4. DNA Analysis, GWAS and Validation Methodology, Genes Annotation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- White, P.J.; Broadley, M.R. Biofortification of crops with seven mineral elements often lacking in human diets—Iron, zinc, copper, calcium, magnesium, selenium and iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef] [PubMed]
- Velu, G.; Ortiz-Monasterio, I.; Cakmak, I.; Hao, Y.; Singh, R.P. Biofortification strategies to increase grain zinc and iron concentrations in wheat. J. Cereal Sci. 2014, 59, 365–372. [Google Scholar] [CrossRef]
- Gupta, P.K.; Balyan, H.S.; Sharma, S.R. Biofortification and bioavailability of Zn, Fe and Se in wheat: Present status and future prospects. Theor. Appl. Genet. 2021, 134, 1–35. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, Q.; Liao, X.; Li, X.; Zheng, S.; Zhao, F. Phytoexclusion of heavy metals using low heavy metal accumulating cultivars: A green technology. J. Hazard. Mater. 2021, 413, 125427. [Google Scholar] [CrossRef]
- Bhatt, R.; Hossain, A.; Sharma, P. Zinc biofortification as an innovative technology to alleviate the zinc deficiency in human health: A review. Open Agric. 2020, 5, 176–187. [Google Scholar] [CrossRef]
- Velu, G.; Singh, R.P.; Joshi, A.K. A decade of progress on genetic enhancement of grain zinc and iron in CIMMYT wheat germplasm. In Wheat and Barley Grain Biofortification; Gupta, P., Pandey, V., Narwal, S., Sharma, P., Ram, S., Singh, G.P., Eds.; Woodhead Publishing: Cambridge, UK; pp. 129–138.
- Sazawal, S.; Dhingra, U.; Dhingra, P.; Dutta, A.; Deb, S.; Kumar, J.; Devi, P.; Prakash, A. Efficacy of high zinc biofortified wheat in improvement of micronutrient status, and prevention of morbidity among preschool children and women—A double masked, randomized, controlled trial. Nutr. J. 2018, 17, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgounov, A.; Gómez-Becerra, H.F.; Abugalieva, A.; Dzhunusova, M.; Yessimbekova, M.; Muminjanov, H.; Zelenskiy, Y.; Ozturk, L.; Cakmak, I. Iron and zinc grain density in common wheat grown in Central Asia. Euphytica 2007, 155, 193–203. [Google Scholar] [CrossRef] [Green Version]
- Tattibayeva, D.; Nebot, C.; Miranda, J.M.; Abuova, A.B.; Baibatyrov, T.A.; Kizatova, M.Z.; Cepeda, A.; Franco, C.M. A study on toxic and essential elements in wheat grain from the Republic of Kazakhstan. Environ. Sci. Pollut. Res. 2016, 23, 5527–5537. [Google Scholar] [CrossRef]
- Morgounov, A.I.; Belan, I.; Zelenskiy, Y.; Roseeva, L.; Tömösközi, S.; Békés, F.; Abugalieva, A.; Cakmak, I.; Vargas, M.; Crossa, J. Historical changes in grain yield and quality of spring wheat varieties cultivated in Siberia from 1900 to 2010. Can. J. Plant Sci. 2013, 93, 425–433. [Google Scholar] [CrossRef]
- Berseneva, M.L. The content of some heavy metals in wheat grain. Newsl. Krasn. State Univ. 2018, 2, 9–14. [Google Scholar]
- Gryaznov, A.A.; Senkova, L.A.; Grinets, L.V. Heavy metals in wheat and barley in Chelyabinsk region. In Proceedings of the VI International Scientific and Practical Conference, Online, 15–16 November 2018; Ural SAU: Yekaterinburg, Russia, 2018; pp. 233–238. [Google Scholar]
- Abugalieva, A.I.; Flis, P.; Shamanin, V.P.; Savin, T.; Morgounov, A. Ionomic analysis of spring wheat grain produced in Ka-zakhstan and Russia. Comm. Soil Sci. Plant Anal. 2021, 52, 704–7011. [Google Scholar] [CrossRef]
- Morgounov, A.; Savin, T.; Flis, P.; Babkenov, A.; Chudinov, V.; Kazak, A.; Koksel, H.; Likhenko, I.; Sharma, R.; Shelaeva, T.; et al. Effects of environments and cultivars on grain ionome of spring wheat grown in Kazakhstan and Russia. Crop Pasture Sci. 2022, 73, 515–527. [Google Scholar] [CrossRef]
- Shepelev, S.; Morgounov, A.; Flis, P.; Koksel, H.; Li, H.; Savin, T.; Sharma, R.; Wang, J.; Shamanin, V. Variation of macro- and microelements, and trace metals in spring wheat genetic resources in Siberia. Plants 2022, 11, 149. [Google Scholar] [CrossRef] [PubMed]
- Bhatta, M.; Shamanin, V.; Shepelev, S.; Baenziger, P.S.; Pozherukova, V.; Pototskaya, I.; Morgounov, A. Genetic diversity and population structure analysis of synthetic and bread wheat accessions in Western Siberia. J. Appl. Genet. 2019, 60, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Bhatta, M.; Shamanin, V.; Shepelev, S.; Baenziger, P.S.; Pozherukova, V.; Pototskaya, V.; Morgounov, A. Marker-trait associations for enhancing agronomic performance, disease resistance, and grain quality in synthetic and bread wheat accessions in Western Siberia. G3 2019, 9, 4209–4222. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Wen, W.; He, Z.; Liu, J.; Jin, H.; Cao, S.; Geng, H.; Yan, J.; Zhang, P.; Wan, Y.; et al. Genome-wide linkage mapping of yield-related traits in three Chinese bread wheat populations using high-density SNP markers. Theor. Appl. Genet. 2018, 131, 1903–1924. [Google Scholar] [CrossRef]
- Bhatta, M.; Morgounov, A.; Belamkar, V.; Baenziger, P.S. Genome-wide association study reveals novel genomic regions for grain yield and yield-related traits in drought-stressed synthetic hexaploid wheat. Int. J. Mol. Sci. 2018, 19, 3011. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Wen, W.; Liu, J.; Zhang, Y.; Cao, S.; He, Z.; Rasheed, A.; Jin, H.; Zhang, C.; Yan, J.; et al. Genetic architecture of grain yield in bread wheat based on genome-wide association studies. BMC Plant Biol. 2019, 19, 168. [Google Scholar] [CrossRef]
- Alqudah, A.M.; Haile, J.K.; Alomari, D.Z.; Pozniak, C.J.; Kobiljski, B.; Börner, A. Genome-wide and SNP network analyses reveal genetic control of spikelet sterility and yield-related traits in wheat. Sci. Rep. 2020, 10, 2098. [Google Scholar] [CrossRef]
- Rahimi, Y.; Bihamta, M.R.; Taleei, A.; Alipour, H.; Ingvarsson, P.K. Genome-wide association study of agronomic traits in bread wheat reveals novel putative alleles for future breeding programs. BMC Plant Biol. 2019, 19, 541. [Google Scholar] [CrossRef] [Green Version]
- Cuthbert, J.L.; Somers, D.J.; Brûlé-Babel, A.L.; Brown, P.D.; Crow, G.H. Molecular mapping of quantitative trait loci for yield and yield components in spring wheat (Triticum aestivum L.). Theor. Appl. Genet. 2008, 117, 595–608. [Google Scholar] [CrossRef]
- Kumar, A.; Mantovani, E.E.; Seetan, R.; Soltani, A.; Echeverry-Solarte, M.; Jain, S.; Simsek, S.; Doehlert, D.; Alamri, M.S.; Elias, E.M.; et al. Dissection of genetic factors underlying wheat kernel shape and size in an Elite × Nonadapted cross using a high density SNP linkage map. Plant Genome 2016, 9, plantgenome2015.09.0081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatta, M.; Morgounov, A.; Belamkar, V.; Poland, J.; Baenziger, P.S. Unlocking the novel genetic diversity and population structure of synthetic hexaploid wheat. BMC Genom. 2018, 19, 591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCartney, C.A.; Somers, D.J.; McCallum, B.D.; Thomas, J.; Humphreys, D.G.; Menzies, J.G.; Brown, P.D. Microsatellite tagging of the leaf rust resistance gene Lr16 on wheat chromosome 2BSc. Mol Breed. 2005, 15, 329–337. [Google Scholar] [CrossRef]
- Anzueto-Ponciano, H.A. Identification of Quantitative Trait Loci (QTL) for Tolerance to Metribuzin in Soft Red Winter Wheat (Triticum aestivum L.). Master’s Thesis, Louisiana State University, Baton Rouge, LA, USA, 2018. [Google Scholar]
- Winn, Z.J. An Evaluation of Hybrid Traits, yield, and Major QTL Effect on Heterosis in Hybrid Soft Red Winter Wheat. Master’s Thesis, University of Arkansas, Fayetteville, AR, USA, December 2019. [Google Scholar]
- Wang, W.; Guo, H.; Wu, C. Identification of novel genomic regions associated with nine mineral elements in Chinese winter wheat grain. BMC Plant Biol. 2021, 21, 311. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.; Zhao, C.; Ding, A.; Jun, L.; Wang, L.; Li, X.; Bao, Y.; Li, J.; Wang, H. Construction of an integrative linkage map and QTL mapping of grain yield-related traits using three related wheat RIL populations. Theor. Appl. Genet. 2014, 127, 659–675. [Google Scholar] [CrossRef]
- Bhatta, M.; Baenziger, P.S.; Waters, B.M.; Poudel, R.; Belamkar, V.; Poland, J.; Morgounov, A. Genome-wide association study reveals novel genomic regions associated with 10 grain minerals in synthetic hexaploid wheat. Int. J. Mol. Sci. 2018, 19, 3237. [Google Scholar] [CrossRef] [Green Version]
- Salt, D.E.; Baxter, I.; Lahner, B. Ionomics and the study of the plant ionome. Annu. Rev. Plant Biol. 2008, 59, 709–733. [Google Scholar] [CrossRef] [Green Version]
- Wani, S.H.; Gaikwad, K.; Razzaq, A.; Kajal Samantara, M.K.; Velu, G. Improving zinc and iron biofortification in wheat through genomics approaches. Mol. Biol. Rep. 2022, 49, 8007–80023. [Google Scholar] [CrossRef]
- Kamaral, C.; Neate, S.M.; Gunasinghe, N.; Milham, P.J.; Paterson, D.J.; Kopittke, P.M. Genetic biofortification of wheat with zinc: Opportunities to fine-tune zinc uptake, transport and grain loading. Physiol. Plant. 2022, 174, e13612. [Google Scholar] [CrossRef]
- Rathan, N.D.; Krishna, H.; Ellur, R.K.; Sehgal, D.; Govindan, V.; Ahlawat, A.K.; Krishnappa, G.; Jaiswal, J.P.; Singh, J.B.; Sv, S.; et al. Genome-wide association study identifies loci and candidate genes for grain micronutrients and quality traits in wheat (Triticum aestivum L.). Sci. Rep. 2022, 12, 7037. [Google Scholar] [CrossRef] [PubMed]
- Hao, B.; Ma, J.; Si, L.; Jiang, L.; Wang, X.; Yao, C.; Ma, S.; Li, C.; Gao, Z. Did wheat breeding simultaneously alter grain concentrations of macro- and micro-nutrient over the past 80 years of cultivar releasing in China? Front. Plant Sci. 2022, 13, 872781. [Google Scholar] [CrossRef] [PubMed]
- Alomari, D.Z.; Alqudah, A.M.; Pillen, K.; von Wirén, N.; Röder, M.S. Toward identification of a putative candidate gene for nutrient mineral accumulation in wheat grains for human nutrition purposes. J. Exp. Bot. 2021, 72, 6305–6318. [Google Scholar] [CrossRef] [PubMed]
- El-Soda, M.; Aljabri, M. Genome-wide association mapping of grain metal accumulation in wheat. Genes 2022, 13, 1052. [Google Scholar] [CrossRef] [PubMed]
- Morgounov, A.; Sonder, K.; Abugalieva, A.; Bhadauria, V.; Cuthbert, R.D.; Shamanin, V. Effect of climate change on spring wheat yields in North America and Eurasia in 1981-2015 and implications for breeding. PLoS ONE 2018, 13, e0204932. [Google Scholar] [CrossRef] [PubMed]
- Alomari, D.Z.; Eggert, K.; Von Wirén, N.; Polley, A.; Plieske, J.; Ganal, M.W.; Liu, F.; Pillen, K.; Röder, M.S. Whole-genome association mapping and genomic prediction for iron concentration in wheat grains. Int. J. Mol. Sci. 2019, 20, 76. [Google Scholar] [CrossRef] [Green Version]
- Shamanin, V.; Salina, E.; Wanyera, R.; Zelenskiy, Y.; Olivera, P.; Morgounov, A. Genetic diversity of spring wheat from Kazakhstan and Russia for resistance to stem rust Ug99. Euphytica 2016, 212, 287–296. [Google Scholar] [CrossRef]
- Aberkane, H.; Payne, T.; Kishi, M.; Smale, M.; Amri, A.; Jamora, N. Transferring diversity of goat grass to farmers’ fields through the development of synthetic hexaploid wheat. Food Secur. 2020, 12, 1017–1033. [Google Scholar] [CrossRef]
- Shamanin, V.; Shepelev, S.; Pozherukova, V.; Gultyaeva, E.; Kolomiets, T.; Pakholkova, E.; Morgounov, A. Primary hexaploid synthetics: Novel sources of wheat disease resistance. Crop Prot. 2019, 121, 7–10. [Google Scholar] [CrossRef]
- Morgounov, A.; Abugalieva, A.; Akan, K.; Akın, B.; Baenziger, S.P.; Bhatta, M.; Zelenskiy, Y. High-yielding winter synthetic hexaploid wheats resistant to multiple diseases and pests. Plant Genet. Resour. 2018, 16, 273–278. [Google Scholar] [CrossRef]
- Matsuoka, Y.; Takumi, S.; Kawahara, T. Natural variation for fertile triploid F1 hybrid formation in allohexaploid wheat speciation. Theor. Appl. Genet. 2007, 115, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Pask, A.J.D.; Pietragalla, J.; Mullan, D.M.; Reynolds, M.P. Physiological Breeding II: A Field Guide to Wheat Phenotyping; CIMMYT: Mexico City, Mexico, 2012. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Glaubitz, J.C.; Casstevens, T.M.; Lu, F.; Harriman, J.; Elshire, R.J.; Sun, Q.; Buckler, E.S. TASSEL-GBS: A high capacity genotyping by sequencing analysis pipeline. PLoS ONE 2014, 9, e90346. [Google Scholar] [CrossRef] [PubMed]
- International Wheat Genome Sequencing Consortium. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, 7191. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
| Elements | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ca | K | Mg | P | S | Cu | Fe | Mn | Zn | Cd | Co | Ni | Mo | Rb | Sr | All | |
| Number of significant marker-element associations (Groups 1–4) | ||||||||||||||||
| All | 229 | 32 | 499 | 394 | 399 | 111 | 63 | 224 | 212 | 243 | 19 | 381 | 43 | 78 | 212 | 2997 |
| Chr. 1 | 35 | 2 | 180 | 165 | 13 | 2 | 18 | 18 | 40 | 33 | 0 | 30 | 6 | 3 | 40 | 548 |
| Chr. 2 | 5 | 5 | 49 | 42 | 47 | 17 | 14 | 20 | 46 | 63 | 1 | 45 | 11 | 8 | 46 | 384 |
| Chr. 3 | 9 | 6 | 41 | 38 | 28 | 26 | 6 | 23 | 7 | 31 | 5 | 111 | 9 | 31 | 7 | 372 |
| Chr. 4 | 8 | 2 | 31 | 21 | 58 | 19 | 5 | 67 | 33 | 35 | 0 | 23 | 3 | 2 | 33 | 307 |
| Chr. 5 | 51 | 3 | 110 | 41 | 50 | 8 | 7 | 20 | 30 | 17 | 3 | 59 | 4 | 16 | 30 | 436 |
| Chr. 6 | 108 | 7 | 24 | 54 | 189 | 23 | 6 | 12 | 47 | 23 | 9 | 62 | 3 | 8 | 47 | 598 |
| Chr. 7 | 13 | 7 | 64 | 43 | 14 | 16 | 7 | 64 | 9 | 41 | 1 | 51 | 7 | 10 | 9 | 352 |
| Gen. A | 58 | 17 | 203 | 99 | 130 | 35 | 25 | 66 | 47 | 88 | 3 | 94 | 20 | 31 | 47 | 937 |
| Gen. B | 33 | 10 | 222 | 178 | 119 | 40 | 16 | 140 | 97 | 93 | 15 | 154 | 10 | 26 | 97 | 1191 |
| Gen. D | 138 | 5 | 74 | 117 | 150 | 36 | 22 | 18 | 68 | 62 | 1 | 133 | 13 | 21 | 68 | 869 |
| Number of SNPs with significant effects | ||||||||||||||||
| Group 1 | 3 | 0 | 14 | 5 | 0 | 1 | 0 | 8 | 0 | 13 | 1 | 3 | 0 | 0 | 2 | 50 |
| Group 2 | 30 | 2 | 2 | 4 | 11 | 2 | 2 | 5 | 15 | 23 | 0 | 95 | 4 | 0 | 2 | 197 |
| # | SNP | QTL Region | Element | REF | ALT | 2017 | 2018 | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Effect, µg/g | Effect, % | −log10(p-Value) > 3.5 | Effect, µg/g−1 | Effect, % | −log10(p-Value) > 3.5 | ||||||
| 1 | S3B_807804964 | 442 | Ca | A | G | 39.9 | 11.0 | 3.57 | 37.7 | 9.6 | 3.67 |
| 2 | S6D_27846508 | 878 | Ca | T | A | 30.4 | 8.4 | 4.05 | 28.0 | 7.1 | 3.75 |
| 3 | S1B_468389275 | 112 | Mg | C | A | 38.2 | 3.2 | 3.78 | 60.7 | 4.9 | 3.71 |
| 4 | S1B_470419196 | 113 | Mg | C | T | 38.2 | 3.2 | 3.99 | 60.0 | 4.9 | 3.85 |
| 5 | S1B_483598145 | 114 | Mg | T | G | 51.4 | 4.2 | 4.24 | 75.6 | 6.2 | 3.63 |
| 6 | S2A_738732586 | 227 | Mg | T | G | 48.2 | 4.0 | 3.81 | 80.8 | 6.6 | 4.15 |
| 7 | S4B_64816370 | 562 | Mg | C | T | 46.7 | 3.9 | 3.77 | 75.4 | 6.1 | 3.83 |
| 8 | S5B_679675578 | 738 | Mg | T | C | 33.3 | 2.7 | 3.58 | 21.2 | 1.7 | 4.38 |
| 9 | S6D_469161928 | 958 | Mg | C | G | 32.9 | 2.7 | 4.03 | 17.5 | 1.4 | 3.78 |
| 10 | S7B_723334278 | 1040 | Mg | C | T | −52.7 | −4.4 | 3.78 | −80.8 | −6.6 | 3.50 |
| 11 | S6B_610963068 | 852 | P | G | A | −246 | −4.7 | 3.80 | −131 | −2.8 | 3.67 |
| 12 | S6B_610963076 | 852 | P | G | T | −135 | −2.6 | 3.88 | −131 | −2.8 | 3.67 |
| 13 | S6D_376894590 | 932 | P | G | A | −319 | −6.2 | 3.87 | −188 | −4.0 | 4.49 |
| 14 | S6D_29369738 | 879 | Cu | C | G | 0.320 | 6.9 | 3.59 | 0.281 | 7.5 | 3.78 |
| 15 | S2A_24200649 | 206 | Mn | G | A | −3.22 | −7.5 | 4.25 | −3.13 | −7.0 | 3.96 |
| 16 | S3A_697506434 | 374 | Mn | G | T | −2.41 | −5.6 | 3.88 | −2.35 | −5.3 | 3.66 |
| 17 | S4B_603519569 | 579 | Mn | C | A | −3.52 | −8.2 | 3.56 | −3.53 | −7.9 | 3.98 |
| 18 | S7B_574853540 | 1015 | Mn | G | T | −1.82 | −4.2 | 3.60 | −1.93 | −4.3 | 3.94 |
| 19 | S7B_720831474 | 1039 | Mn | C | T | −3.60 | −8.3 | 4.15 | −3.93 | −8.8 | 4.83 |
| 20 | S2A_751844369 | 231 | Cd | A | G | 0.008 | 18.9 | 3.87 | 0.007 | 21.0 | 4.88 |
| 21 | S2B_772063522 | 297 | Cd | C | G | 0.016 | 35.3 | 4.61 | 0.010 | 31.5 | 3.87 |
| 22 | S2B_88259062 | 246 | Cd | G | T | 0.007 | 16.8 | 3.95 | 0.005 | 15.5 | 3.53 |
| 23 | S3D_550209436 | 482 | Cd | G | A | −0.008 | −18.1 | 4.03 | 0.012 | −39.6 | 3.72 |
| 24 | S4D_11471805 | 592 | Cd | C | T | 0.010 | 23.0 | 3.87 | 0.007 | 24.0 | 4.25 |
| 25 | S5D_486749507 | 768 | Cd | C | T | 0.014 | 32.8 | 4.50 | 0.009 | 28.3 | 3.55 |
| 26 | S7B_677743542 | 1033 | Cd | A | G | 0.008 | 19.0 | 3.69 | 0.006 | 19.1 | 3.85 |
| 27 | S6D_454012454 | 953 | Sr | G | A | 0.338 | 16.2 | 3.62 | 0.329 | 13.6 | 3.79 |
| Element | Mean Concentration Across 2017–2018, µg/g | LSD 0.05 | |||||
|---|---|---|---|---|---|---|---|
| Karabalyk, KZ | Shortandy, KZ | Chelyabinsk, RU | Omsk, RU | Novosibirsk, RU | Tyumen, RU | ||
| Ca | 306 | 319 | 309 | 373 | 391 | 341 | 29 |
| K | 4176 | 3908 | 4276 | 4022 | 4093 | 4313 | 185 |
| Mg | 1314 | 1089 | 1225 | 1293 | 1213 | 1177 | 59 |
| P | 4944 | 3960 | 5343 | 5387 | 5269 | 5320 | 360 |
| S | 1818 | 1948 | 1719 | 1949 | 1829 | 1515 | 111 |
| Cu | 3.82 | 5.28 | 3.46 | 4.67 | 4.19 | 4.20 | 2.6 |
| Fe | 34.6 | 32.5 | 33.7 | 39.7 | 33.8 | 31.0 | 0.45 |
| Mn | 40.8 | 42.0 | 34.1 | 41.8 | 38.6 | 30.3 | 3.1 |
| Zn | 37.2 | 26.9 | 44.3 | 50.3 | 44.9 | 44.9 | 6.2 |
| Cd | 0.021 | 0.025 | 0.026 | 0.033 | 0.030 | 0.018 | 0.004 |
| Co | 0.020 | 0.027 | 0.008 | 0.011 | 0.007 | 0.004 | 0.006 |
| Ni | 0.402 | 0.249 | 0.541 | 0.257 | 0.123 | 0.336 | 0.091 |
| Mo | 0.461 | 0.571 | 0.190 | 0.290 | 0.268 | 0.488 | 0.10 |
| Rb | 1.88 | 1.76 | 4.07 | 3.28 | 7.06 | 4.88 | 1.22 |
| Sr | 2.83 | 2.34 | 1.67 | 2.05 | 1.69 | 1.65 | 0.34 |
| # | SNP | Element | QTL Region | Ref. SNP | Alt. SNP | GWAS Panel * | KASIB Validation Trial ** | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | Effect, µg/g | % | −log10(p-Value) > 3.5 | No. of Sites | Effect, µg/g | % | ||||||
| 1 | S1B_9711623 | P | 58 | C | T | 2017 | 157 | 3.0 | 4.95 | 2 | 598 | 12.0 |
| 2 | S1B_10111796 | P | 58 | G | T | 17 | 149 | 2.9 | 4.69 | 7 | 514 | 10.0 |
| 3 | S1B_13242483 | P | 59 | A | G | 17 | 150 | 2.9 | 4.89 | 4 | 539 | 10.6 |
| 4 | S1B_114437220 | P | 82 | T | C | 17 | 156 | 3.0 | 4.87 | 3 | 385 | 7.3 |
| 5 | S1B_176291121 | Mg | 96 | A | G | 17 | −31.8 | −2.6 | 4.01 | 1 | −75 | −5.6 |
| P | 96 | A | G | 17 | 152 | 2.9 | 4.79 | 3 | 608 | 12.1 | ||
| 6 | S1B_184771090 | P | 99 | T | C | 17 | 153 | 2.9 | 4.76 | 4 | 509 | 9.9 |
| 7 | S2A_726322626 | Mo | 223 | A | G | 17–18 | 0.041 | 12.1 | 9.29 | 10 | 0.25 | 63.1 |
| 8 | S2B_780115106 | Cd | 300 | A | G | 18 | −0.003 | −9.7 | 4.24 | 4 | −0.09 | −26.6 |
| Mn | 300 | A | G | 17 | −1.96 | −4.5 | 5.19 | 6 | −4.51 | −10.7 | ||
| 9 | S2B_780665986 | Cd | 300 | T | C | 17–18 | 0.001 | 3.2 | 4.50 | 4 | 0.09 | 28.1 |
| Mn | 300 | T | C | 17 | −2.06 | −4.8 | 5.57 | 4 | −4.68 | −11.5 | ||
| Zn | 300 | T | C | 17 | −3.01 | −5.6 | 3.90 | 1 | −7.0 | −13.5 | ||
| 10 | S3B_758201335 | Ni | 432 | A | G | 17–18 | 0.004 | 2.7 | 4.11 | 3 | 0.054 | 33.4 |
| 11 | S4B_23355392 | S | 561 | C | T | 17 | 87.9 | 4.3 | 5.25 | 6 | 203 | 11.6 |
| 12 | S5A_568799967 | Ca | 658 | C | G | 18 | −23.2 | −5.9 | 6.28 | 6 | −44 | −13.7 |
| 13 | S5A_569526776 | Ca | 658 | C | T | 18 | 19.8 | 5.0 | 4.78 | 4 | 38 | 12.2 |
| 14 | S5A_570718644 | Ca | 659 | A | G | 18 | −21.1 | −5.3 | 4.81 | 5 | −28 | −9.2 |
| 15 | S5A_570788577 | Ca | 659 | T | G | 18 | −23.9 | −6.0 | 4.97 | 5 | −40 | −13.1 |
| 16 | S5A_594133493 | Sr | 661 | A | T | 18 | −0.158 | −6.5 | 4.86 | 5 | −0.225 | −11.2 |
| 17 | S5A_698528417 | Sr | 677 | C | G | 17–18 | 0.256 | 10.6 | 5.69 | 4 | 0.475 | 21.1 |
| 18 | S5D_43408942 | Ca | 749 | A | G | 17–18 | 30.1 | 7.6 | 3.61 | 3 | 88 | 24.5 |
| 19 | S6B_562488824 | P | 846 | C | T | 17 | −308 | −5.9 | 3.98 | 2 | −552 | −10.7 |
| Zn | 846 | C | T | 17 | −4.66 | −8.7 | 3.56 | 2 | −10.1 | −20.6 | ||
| 20 | S6B_601138481 | P | 848 | C | T | 17 | −308 | −6.0 | 3.98 | 1 | −709 | −14.4 |
| Zn | 848 | C | T | 17 | −4.66 | −8.7 | 3.56 | 2 | −10.1 | −22.4 | ||
| SNP | Position (Mb) | Annotation | Gene ID * | Description | References |
|---|---|---|---|---|---|
| S1B_9711623 | 9.711623 | Intergenic region | TraesCS1B03G0039600-TraesCS1B03G0040000 | G-type lectin S-receptor-like serine/threonine-protein kinase At2g19130, Putative 12-oxophytodienoate reductase 4 | [18,19] |
| S1B_10111796 | 10.111796 | Upstream gene variant | TraesCS1B03G0041500 | Rust resistance kinase Lr10 | |
| S1B_13242483 | 13.242483 | Intergenic region | TraesCS1B03G0051800-TraesCS1B03G0051900 | Bowman-Birk type trypsin inhibitor | |
| S1B_114437220 | 114.43722 | Intergenic region | TraesCS1B03G0269300-TraesCS1B03G0269500 | Chitinase 10, Hydrophobic protein LTI6B | |
| S1B_176291121 | 176.291121 | Intergenic region | TraesCS1B03G0363900-TraesCS1B03G0364700 | LRR receptor-like serine/threonine-protein kinase | |
| S1B_184771090 | 184.77109 | Intergenic region | TraesCS1B03G0374100-TraesCS1B03G0374900 | Pre-mRNA splicing factor SR-like 1, E3 ubiquitin-protein ligase | |
| S1B_468389275 | 468.389275 | Intergenic region | TraesCS1B03G0736700-TraesCS1B03G0737500 | NADH dehydrogenase [ubiquinone] iron-sulfur protein 2 | |
| S1B_470419196 | 470.419196 | Intergenic region | TraesCS1B03G0739000-TraesCS1B03G0739200 | Uncharacterized protein, Autophagy-related protein 18 h | |
| S1B_483598145 | 483.598145 | Intron variant | TraesCS1B03G0758900 | Guanosine nucleotide diphosphate dissociation inhibitor 1 | [20] |
| S2A_24200649 | 24.200649 | Intergenic region | TraesCS2A03G0098800-TraesCS2A03G0099400 | Bidirectional sugar transporter SWEET6b | [19] |
| S2B_88259062 | 88.259062 | Intergenic region | TraesCS2B03G0283000-TraesCS2B03G0283400 | Probable pectinesterase 53 | [19] |
| S2B_772063522 | 772.063522 | Intergenic region | TraesCS2B03G1432900-TraesCS2B03G1433300 | BTB/POZ and MATH domain-containing protein 2 | [21] |
| S2B_780115106 | 780.115106 | Downstream gene variant | TraesCS2B03G1459800 | N/A | [21] |
| S2B_780665986 | 780.665986 | Intergenic region | TraesCS2B03G1462400-TraesCS2B03G1462900 | N/A | [21] |
| S7B_574853540 | 574.85354 | Intergenic region | TraesCS7B03G0857100-TraesCS7B03G0858100 | Eukaryotic initiation factor 4A, Cadmium/zinc-transporting ATPase HMA2 | |
| S7B_677743542 | 677.743542 | Intergenic region | TraesCS7B03G1084500-TraesCS7B03G1085100 | MMS19 nucleotide excision repair protein homolog, Ethylene-responsive transcription factor 12 | [19,22] |
| S7B_720831474 | 720.831474 | Downstream gene variant | TraesCS7B03G1209100 | Receptor-like cytoplasmic kinase 176 | [18,19] |
| S3A_697506434 | 697.506434 | Intergenic region | TraesCS3A03G1072400-TraesCS3A03G1072800 | N/A | |
| S3B_758201335 | 758.201335 | Intergenic region | TraesCS3B03G1233100-TraesCS3B03G1234800 | Amino acid transporter AVT1I, Glutathione transferase GST 23 | [19,20,22,23] |
| S3B_807804964 | 807.804964 | Intergenic region | TraesCS3B03G1388900-TraesCS3B03G1389300 | Probable disease resistance protein | [21] |
| S3D_550209436 | 550.209436 | Intergenic region | TraesCS3D03G0963300-TraesCS3D03G0963700 | LRR receptor-like serine/threonine-protein kinase EFR, Serine carboxypeptidase 1 | |
| S4B_23355392 | 23.355392 | Intergenic region | TraesCS4B03G0059500-TraesCS4B03G0060200 | E3 ubiquitin-protein ligase UPL7 | [22,24] |
| S4B_64816370 | 64.81637 | Intergenic region | TraesCS4B03G0150900-TraesCS4B03G0151600 | N/A | [25] |
| S4B_603519569 | 603.519569 | Intergenic region | TraesCS4B03G0818900-TraesCS4B03G0819000 | Mitogen-activated protein kinase kinase 1 | |
| S4D_11471805 | 11.471805 | Intergenic region | TraesCS4D03G0040900-TraesCS4D03G0041000 | Cysteine proteinase inhibitor 8, Pollen allergen Phl p 5.0101 | [21,26] |
| S5A_568799967 | 568.799967 | Intergenic region | TraesCS5A03G0881300-TraesCS5A03G0882600 | RuBisCO large subunit-binding protein subunit alpha, chloroplastic (Fragment), B2 protein | |
| S5A_569526776 | 569.526776 | Intergenic region | TraesCS5A03G0882600-TraesCS5A03G0882900 | B2 protein | [27] |
| S5A_570718644 | 570.718644 | Intergenic region | TraesCS5A03G0885700-TraesCS5A03G0887200 | Acyl-coenzyme A thioesterase 13 | [20] |
| S5A_570788577 | 570.788577 | Intergenic region | TraesCS5A03G0887200-TraesCS5A03G0887300 | Acyl-coenzyme A thioesterase 13 | [20] |
| S5A_594133493 | 594.133493 | Intergenic region | TraesCS5A03G0949400-TraesCS5A03G0949800 | Gibberellin-regulated protein 8, Snakin-1 | [20] |
| S5A_698528417 | 698.528417 | Intergenic region | TraesCS5A03G1257600-TraesCS5A03G1258800 | CCR4-NOT transcription complex subunit 1 | [22,28] |
| S5B_679675578 | 679.675578 | Intergenic region | TraesCS5B03G1245900-TraesCS5B03G1246800 | N/A | [21] |
| S5D_43408942 | 43.408942 | Upstream gene variant | TraesCS5D03G0110300 | BTB/POZ and MATH domain-containing protein 1 | |
| S5D_486749507 | 486.749507 | Intergenic region | TraesCS5D03G0937000-TraesCS5D03G0938500 | N/A | [21,22] |
| S6B_562488824 | 562.488824 | Intergenic region | TraesCS6B03G0886600-TraesCS6B03G0886800 | Fatty acid desaturase DES2 | [20] |
| S6B_601138481 | 601.138481 | Intergenic region | TraesCS6B03G0954400-TraesCS6B03G0954500 | Methyltransferase-like protein 7A | |
| S6B_610963068 | 610.963068 | Intergenic region | TraesCS6B03G0966200-TraesCS6B03G0966500 | Probable galacturonosyltransferase-like 9, Formin-like protein 16 | [19] |
| S6B_610963076 | 610.963076 | Intergenic region | TraesCS6B03G0966200-TraesCS6B03G0966500 | Probable galacturonosyltransferase-like 9, Formin-like protein 16 (Bhatta et al., 2018b) [19] | [19] |
| S6D_27846508 | 27.846508 | Intergenic region | TraesCS6D03G0114300-TraesCS6D03G0114400 | 60S ribosomal protein L13-1, AT-hook motif nuclear-localized protein 20 | |
| S6D_29369738 | 29.369738 | Intergenic region | TraesCS6D03G0114300-TraesCS6D03G0114400 | 60S ribosomal protein L13-1, AT-hook motif nuclear-localized protein 20 | |
| S6D_376894590 | 376.89459 | Intergenic region | TraesCS6D03G0607800-TraesCS6D03G0608400 | Abscisic acid 8′-hydroxylase 1, WAT1-related protein | |
| S6D_454012454 | 454.012454 | Intergenic region | TraesCS6D03G0764500-TraesCS6D03G0764700 | Guanine nucleotide exchange factor subunit RIC1 | [19] |
| S6D_469161928 | 469.161928 | Intergenic region | TraesCS6D03G0810400-TraesCS6D03G0810500 | N/A | [18,19,22] |
| S7B_723334278 | 723.334278 | Intergenic region | TraesCS7B03G1219500-TraesCS7B03G1220400 | N/A | [19,20,29] |
| S2A_726322626 | 726.322626 | Intergenic region | TraesCS2A03G1140100-TraesCS2A03G1140200 | VQ motif-containing protein 25 | [30] |
| S2A_738732586 | 738.732586 | Downstream gene variant | TraesCS2A03G1180900 | Golgin candidate 2 | [20,21,31] |
| S2A_751844369 | 751.844369 | Intergenic region | TraesCS2A03G1227800-TraesCS2A03G1227900 | N/A | [20] |
| Element | No. of SNPs | % of Reference SNP Alleles in Germplasm Groups: | |||
|---|---|---|---|---|---|
| Synthetics-CIMMYT | Synthetics-Japan | USA Cultivars | KASIB Germplasm | ||
| Frequency of germplasm | 28.5 | 5.1 | 10.2 | 56.2 | |
| Ca | 7 | 26.7 | 4.3 | 10.6 | 58.4 |
| Mg | 8 | 29.4 | 4.0 | 8.3 | 58.2 |
| P | 7 | 31.8 | 5.6 | 12.0 | 50.7 |
| S | 1 | 20.7 | 6.3 | 7.2 | 65.8 |
| Cu | 1 | 27.9 | 2.9 | 13.5 | 55.8 |
| Mn | 5 | 25.4 | 1.1 | 9.4 | 64.1 |
| Cd | 7 | 28.8 | 2.2 | 8.2 | 60.8 |
| Ni | 1 | 24.1 | 0.0 | 3.4 | 72.4 |
| Mo | 1 | 32.5 | 5.3 | 12.3 | 50.0 |
| Sr | 3 | 23.2 | 3.3 | 8.1 | 65.4 |
| Mg, P | 1 | 37.8 | 7.1 | 12.2 | 42.9 |
| P, Zn | 2 | 26.9 | 4.8 | 11.2 | 57.0 |
| Cd, Mn | 1 | 18.6 | 8.1 | 14.0 | 59.3 |
| Cd, Mn, Zn | 1 | 13.0 | 9.1 | 16.9 | 61.0 |
| Entry # | Genotype | Yield | Protein Content | 1000 Kernel Weight | Optimal Concentration of the Following Elements * | |||
|---|---|---|---|---|---|---|---|---|
| g/m2 | +LC | % | +LC | g | +LC | |||
| - | Pamyati Azieva, local check (LC) | 399 | 16.5 | 43.5 | - | |||
| 12 | Aisberg/Ae. squarrosa (511) | 245 | −38.7 | 16.8 | 2.3 | 43.4 | −0.3 | Ca, S, Cu, Fe, Mo |
| 13 | Ukr-Od 1530.94/Ae. squarrosa (392) | 313 | −21.7 | 17.2 | 4.2 | 42.0 | −3.6 | Mg, Fe, Mn, Rb, Sr |
| 36 | Aisberg/Ae. squarrosa (369)//Demir | 273 | −31.7 | 16.3 | −0.7 | 47.5 | 9.1 | Mg, Cu, Zn, Cd, Co, Mo |
| 57 | Ukr-Od 1530.94/Ae. squarrosa (392) | 209 | −47.7 | 18.2 | 10.7 | 47.6 | 9.4 | K, P, S, Zn, Cd, Mo, Sr |
| 14 | Langdon/KU-2075 | 92 | −77.1 | 21.1 | 28.1 | 40.3 | −7.6 | P, Cu, Fe, Mn, Cd, Rb |
| 22 | Langdon/IG 48042 | 141 | −64.7 | 20.7 | 25.8 | 41.8 | −4.1 | Ca, Fe, Cd, Ni, Mo |
| 47 | Langdon/KU-2093 | 105 | −73.8 | 21.5 | 30.5 | 47.0 | 7.8 | K, Mg, S, Zn, Co, Sr |
| 72 | Tom | 285 | −28.7 | 19.2 | 16.7 | 40.0 | −8.1 | K, S, Cd, Mo, Sr |
| 73 | Freyr | 354 | −11.5 | 19.0 | 15.4 | 35.0 | −19.6 | P, Zn, Cd, Co, Ni, Mo |
| 94 | Element-22 | 535 | 33.8 | 17.0 | 3.4 | 44.7 | 2.7 | Ca, P, S, Cu, Ni |
| 96 | Lutescens-96-12 | 432 | 8.1 | 16.8 | 2.2 | 44.0 | 1.0 | Mg, Cu, Fe, Mo, Sr |
| 99 | Lutescens-6-04-4 | 486 | 21.8 | 18.1 | 9.7 | 47.3 | 8.6 | K, S, Cu, Mn, Cd |
| 103 | Lutescens-15-12 | 372 | −6.8 | 17.6 | 6.7 | 43.8 | 0.5 | P, Cu, Co, Ni, Sr |
| 114 | OmGAU-90 | 466 | 16.7 | 15.7 | −4.6 | 40.4 | −7.2 | Ca, Mg, S, Cu, Fe |
| 116 | Uralosibirskaya | 515 | 28.9 | 17.6 | 6.6 | 49.5 | 13.6 | K, Mg, S, Fe |
| 119 | Duet | 427 | 6.8 | 16.0 | −2.8 | 41.0 | −5.8 | Ca, S, Mn, Ni, Mo |
| 128 | GVK-2161 | 449 | 12.3 | 17.3 | 5.1 | 42.0 | −3.6 | Mg, Fe, Mn, Zn |
| 132 | Lutescens-248-01 | 394 | −1.3 | 15.9 | −3.3 | 49.8 | 14.2 | Mn, Zn, Mo, Sr |
| 136 | Lutescens-48-204-03 | 369 | −7.5 | 16.1 | −2.1 | 49.0 | 12.6 | P, Zn, Cd, Co, Ni, Mo |
| 143 | Lutescens-1103 | 469 | 17.5 | 16.1 | −2.2 | 43.6 | 0.0 | Ca, Mg, Mn, Ni, Mo |
| 156 | Novosibirskaya-41 | 482 | 20.6 | 19.0 | 15.3 | 39.7 | −8.7 | P, Cu, Cd, Co, Ni, Mo |
| 157 | OmGAU-100 | 518 | 29.7 | 16.4 | −0.2 | 43.2 | −0.9 | Ca, P, Cu, Zn, Cd, Ni, Mo, Rb |
| 164 | Silach | 541 | 35.4 | 16.7 | 1.2 | 49.8 | 14.3 | Ca, Mg, Fe, Ni, Rb |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morgounov, A.; Li, H.; Shepelev, S.; Ali, M.; Flis, P.; Koksel, H.; Savin, T.; Shamanin, V. Genetic Characterization of Spring Wheat Germplasm for Macro-, Microelements and Trace Metals. Plants 2022, 11, 2173. https://doi.org/10.3390/plants11162173
Morgounov A, Li H, Shepelev S, Ali M, Flis P, Koksel H, Savin T, Shamanin V. Genetic Characterization of Spring Wheat Germplasm for Macro-, Microelements and Trace Metals. Plants. 2022; 11(16):2173. https://doi.org/10.3390/plants11162173
Chicago/Turabian StyleMorgounov, Alexey, Huihui Li, Sergey Shepelev, Mohsin Ali, Paulina Flis, Hamit Koksel, Timur Savin, and Vladimir Shamanin. 2022. "Genetic Characterization of Spring Wheat Germplasm for Macro-, Microelements and Trace Metals" Plants 11, no. 16: 2173. https://doi.org/10.3390/plants11162173
APA StyleMorgounov, A., Li, H., Shepelev, S., Ali, M., Flis, P., Koksel, H., Savin, T., & Shamanin, V. (2022). Genetic Characterization of Spring Wheat Germplasm for Macro-, Microelements and Trace Metals. Plants, 11(16), 2173. https://doi.org/10.3390/plants11162173