
A Comparison of Three Types of “Vineyard Management” and Their Effects on the Structure of Plasmopara viticola Populations and Epidemic Dynamics of Grape Downy Mildew

Abstract
1. Introduction
2. Results
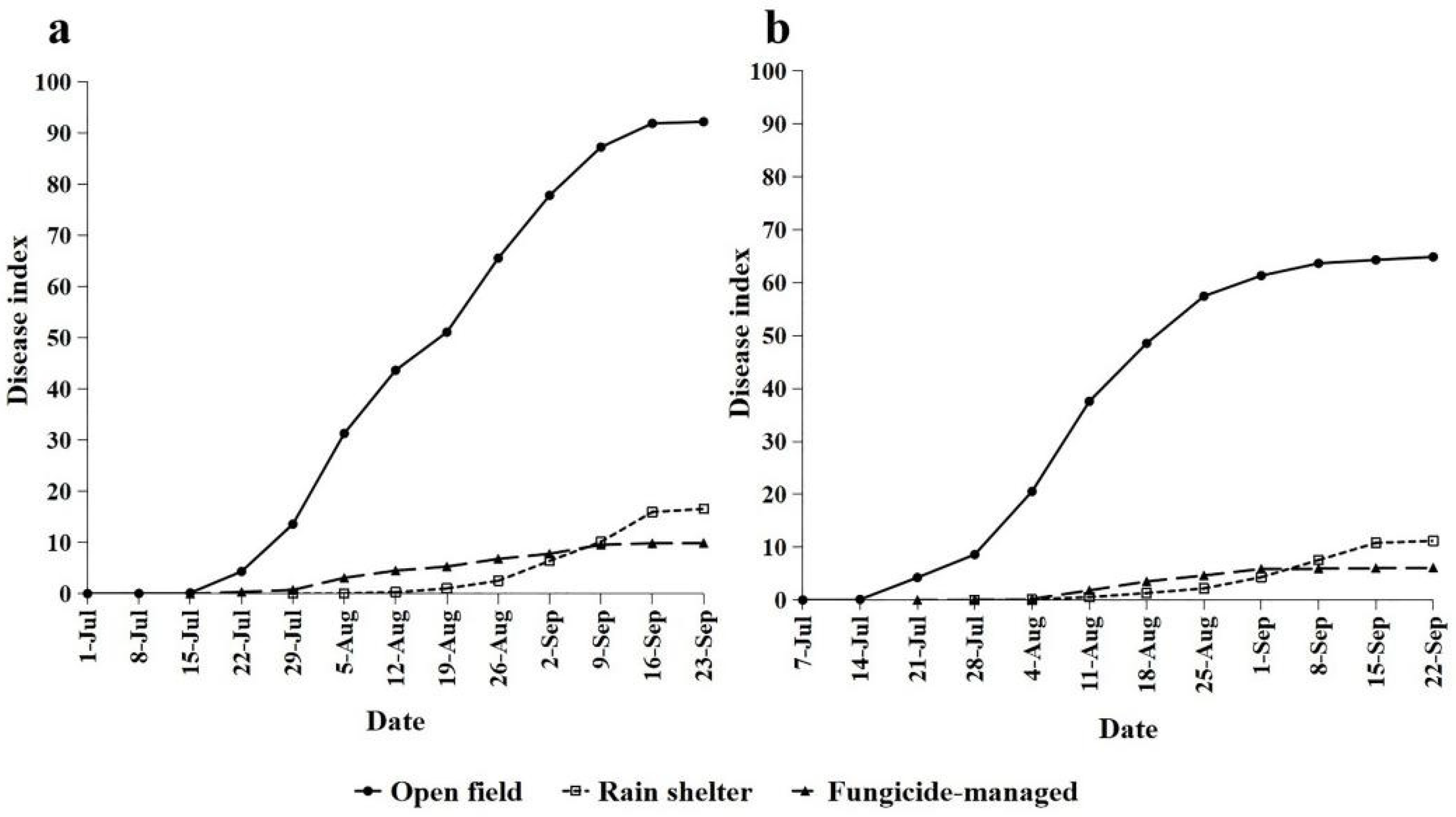
2.1. Disease Epidemic and Meteorological Conditions
2.2. Genotype Identification and Epidemic Mode of P. viticola
2.3. Genetic Structure and Population Differentiation
3. Discussion
4. Materials and Methods
4.1. Experimental Vineyard
4.2. Meteorological Data Collection
4.3. Disease Assessment
4.4. Sampling
4.5. Sample Processing and Simple Sequence Repeats (SSR) Genotyping
4.6. Genetic Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lafon, R.; Clerjeau, M. Downy mildew. In Compendium of Grape Diseases; Pearson, R.C., Goheen, A.C., Eds.; APS Press: St. Paul, MN, USA, 1988; pp. 11–13. [Google Scholar]
- Farlow, W.G. The American grape mildew. Bussy Inst. Bull 1876, 1, 423. [Google Scholar]
- Gessler, C.; Pertot, I.; Perazzolli, M. Plasmopara viticola: A review of knowledge on downy mildew of grapevine and effective disease management. Phytopathol. Mediterr. 2011, 50, 3–44. [Google Scholar]
- Kassemeyer, H.H.; Gadoury, H.; Wilcox, G. Downy Mildew. In Compendium of Grape Diseases, Disorders and Pest; Wilcox, W.F., Uyemoto, J.K., Eds.; APS Press: St. Paul, MN, USA, 2015; pp. 46–51. [Google Scholar]
- Zhao, K.H. An Atlas of Grape Diseases and Pests; China Agriculture Press: Beijing, China, 2005; pp. 6–11. (In Chinese) [Google Scholar]
- Blaeser, M.; Weltzien, H.C. Epidemiologische Studien an Plasmopara viticola zur Verbesserung der Spritzterminbestimmung/Epidemiological studies to improve the control of grapevine downy mildew (Plasmopara viticola). Z. Pflanzenkrankh. Pflanzenschutz/J. Plant Dis. Prot. 1979, 86, 489–498. [Google Scholar]
- Chen, M.; Brun, F.; Raynal, M.; Makowski, D. Forecasting severe grape downy mildew attacks using machine learning. PLoS ONE 2020, 15, e0230254. [Google Scholar] [CrossRef] [PubMed]
- Pertot, I.; Caffi, T.; Rossi, V.; Mugnai, L.; Hoffmann, C.; Grando, M.S.; Gary, C.; Lafond, D.; Duso, C.; Thiery, D.; et al. A critical review of plant protection tools for reducing pesticide use on grapevine and new perspectives for the implementation of IPM in viticulture. Crop Prot. 2017, 97, 70–84. [Google Scholar]
- Hong, C.F.; Brewer, M.T.; Brannen, P.M.; Scherm, H. Temporal disease dynamics and relative importance of sexual and asexual reproduction of grape downy mildew (Plasmopara viticola) in an isolated vineyard in the North Georgia Mountains, USA. Plant Pathol. 2020, 69, 1721–1730. [Google Scholar] [CrossRef]
- Yu, S.Y.; Liu, C.Y.; Liang, C.H.; Zang, C.Q.; Liu, L.; Wang, H.; Guan, T.S. Effects of rain-shelter cultivation on the temporal dynamics of grape downy mildew epidemics. J. Phytopathol. 2017, 165, 331–341. [Google Scholar] [CrossRef]
- Meng, J.F.; Ning, P.F.; Xu, T.F.; Zhang, Z.W. Effect of rain shelter cultivation of Vitis vinifera cv. Cabernet Gernischet on the phenolic profifile of berry skins and the incidence of grape diseases. Molecules 2013, 18, 381–397. [Google Scholar]
- Detoni, A.M.; Clemente, E.; Fornari, C. Productivity and quality of grape ‘Cabernet Sauvignon’ produced in organic system under plastic covering. Rev. Bras. Frutic. 2007, 29, 530–534. [Google Scholar] [CrossRef]
- Du, F.; Deng, W.P.; Yang, M.; Wang, H.N.; Mao, R.J.; Shao, J.H.; Fan, J.X.; Chen, Y.D.; Fu, Y.; Li, C.Y.; et al. Protecting grapevines from rainfall in rainy conditions reduces disease severity and enhances profitability. Crop Prot. 2015, 67, 261–268. [Google Scholar] [CrossRef]
- Liang, C.H.; Zhao, K.H.; Liu, C.Y.; Guan, T.S.; Wang, H.; Li, B.H. Experiment on reducing the occurrence of grape downy mildew with simple rain-shelter. China Fruit 2010, 145, 43–45. (In Chinese) [Google Scholar]
- Gobbin, D.; Pertot, I.; Gessler, C. Identification of microsatellite markers for Plasmopara viticola and establishment of high throughput method for SSR analysis. Eur. J. Plant Pathol. 2003, 109, 153–164. [Google Scholar] [CrossRef]
- Gobbin, D.; Jermini, M.; Loskill, B.; Pertot, I.; Raynal, M.; Gessler, C. Importance of secondary inoculum of Plasmopara viticola to epidemics of grapevine downy mildew. Plant Pathol. 2005, 54, 522–534. [Google Scholar] [CrossRef]
- Gobbin, D.; Rumbou, A.; Linde, C.C.; Gessler, C. Population genetic structure of Plasmopara viticola after 125 years of colonization in European vineyards. Mol. Plant Pathol. 2006, 7, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Valsesia, G.; Gobbin, D.; Patocchi, A.; Vecchione, A.; Gessler, C. Development of a high-throughput method for quantification of Plasmopara viticola DNA in grapevine leaves by means of quantitative real-time polymerase chain reaction. Phytopathology 2005, 95, 672–678. [Google Scholar] [CrossRef]
- Delmotte, F.; Chen, W.; Richard–Cervera, S.; Greif, C.; Papura, D.; Giresse, X.; Mondor-genson, G.; Corio-costet, M.F. Microsatellite DNA markers for Plasmopara viticola, the causal agent of downy mildew of grapes. Mol. Ecol. Notes 2006, 6, 379–381. [Google Scholar] [CrossRef]
- Fontaine, M.C.; Austerlitz, F.; Giraud, T.; Labbe, F.; Papura, D.; Richard–Cervera, S.; Delmotte, F. Genetic signature of a range expansion and leap-frog event after the recent invasion of Europe by the grapevine downy mildew pathogen Plasmopara viticola. Mol. Ecol. 2013, 22, 2771–2786. [Google Scholar] [CrossRef]
- Yin, L.; Zhang, Y.L.; Hao, Y.; Lu, J. Genetic diversity and population structure of Plasmopara viticola in China. Eur. J. Plant Pathol. 2014, 140, 365–376. [Google Scholar] [CrossRef]
- Li, X.L.; Yin, L.; Ma, L.J.; Zhang, Y.L.; Lu, J. Pathogenicity variation and population genetic structure of Plasmopara viticola in China. J. Phytopathol. 2016, 164, 863–873. [Google Scholar] [CrossRef]
- Hug, F. Genetic Structure and Epidemiology of Plasmopara viticola Populations from Australian Grape Growing Regions. Master’s Dissertation, ETH Zurich, Zurich, Switzerland, 2005. [Google Scholar]
- Koopman, T.; Linde, C.C.; Fourie, P.H.; McLeod, A. Population genetic structure of Plasmopara viticola in the Western Cape Province of South Africa. Mol. Plant Pathol. 2007, 8, 723–736. [Google Scholar] [CrossRef]
- Taylor, A.S.; Knaus, B.J.; Grünwald, N.J.; Burgess, T. Population genetic structure and cryptic species of Plasmopara viticola in Australia. Phytopathology 2019, 109, 1975–1983. [Google Scholar] [CrossRef]
- Zhang, W.; Manawasinghe, I.S.; Zhao, W.S.; Xu, J.P.; Brooks, S.; Zhao, X.Y.; Hyde, K.D.; Chethana, K.W.T.; Liu, J.H.; Li, X.H.; et al. Multiple gene genealogy reveals high genetic diversity and evidence for multiple origins of Chinese Plasmopara viticola population. Sci. Rep. 2017, 7, 17304. [Google Scholar] [CrossRef] [PubMed]
- Rossi, V.; Giosuè, S.; Caffi, T. Modelling the dynamic of infections caused by sexual and asexual spores during Plasmopara viticola epidemic. J. Plant Pathol. 2009, 91, 615–627. [Google Scholar]
- Kennelly, M.M.; Gadoury, D.M.; Wilcox, W.F.; Magarey, P.A.; Seem, R.C. Primary infection, lesion productivity, and survival of sporangia in the grapevine downy mildew pathogen Plasmopara viticola. Phytopathology 2007, 97, 512–522. [Google Scholar] [CrossRef]
- Milgroom, M.G. Recombination and the multilocus structure of fungal populations. Annu. Rev. Phytopathol. 1996, 34, 457–477. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.Y.; Liu, C.Y.; Wang, H.; Liu, L.; Guan, T.S. Effect of rain-shelter cultivation on temporal and spatial dynamics of airborne sporangia of Plasmopara viticola. Sci. Agric. Sin. 2016, 49, 1892–1902. (In Chinese) [Google Scholar]
- Rumbolz, J.; Wirtz, S.; Kassemeyer, H.H.; Guggenheim, R.; Schafer, E.; Buche, C. Sporulation of Plasmopara viticola: Differentiation and light regulation. Plant Biol. 2002, 4, 413–422. [Google Scholar] [CrossRef]
- Rumbou, A.; Gessler, C. Particular structure of Plasmopara viticola populations evolved under Greek island conditions. Phytopathology 2006, 96, 501–509. [Google Scholar] [CrossRef]
- Kennelly, M.M.; Eugster, C.; Gadoury, D.M.; Smart, C.D.; Seem, R.C.; Gobbin, D.; Gessler, C. Contributions of oospore inoculum to epidemics of grapevine downy mildew (Plasmopara viticola). Phytopathology 2005, 94, S50–S51. [Google Scholar]
- Killigrew, B.X.; Sivasithamparam, K. Absence of oospores of downy mildew of grape caused by Plasmopara viticola as the source of primary inoculum in most Western Australian vineyards. Plant Dis. 2005, 89, 777. [Google Scholar] [CrossRef]
- Kast, W.K.; Stark-Urnau, M. Survival of sporangia from Plasmopara viticola, the downy mildew of grapevine. Vitis Geilweilerhof 1999, 38, 185–186. [Google Scholar]
- Altizer, S.; Harvell, D.; Friedle, E. Rapid evolutionary dynamics and disease threats to biodiversity. Trends Ecol. Evol. 2003, 18, 589–596. [Google Scholar] [CrossRef]
- GB/T 17980.122-2004; Pesticide-Guidelines for the Field Effificacy Trials(I)—Fungicides against Grape Downy Mildew. Ministry of Agriculture of the People’s Republic of China: Beijing, China, 2004.
- Madden, L.V.; Hughes, G.; Bosch, F.V.D. Temporal analysis II: The components of disease. In The Study of Plant Disease Epidemics; APS Press: St. Paul, MN, USA, 2007; pp. 117–143. [Google Scholar]
- Nei, M. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef] [PubMed]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1948, 24, 379–423. [Google Scholar] [CrossRef]
- Belkhir, K.; Borsa, P.; Chikhi, L.; Raufaste, N.; Bonhomme, F. GENETIX 4.05, Logiciel sous WindowsTM pour la Génétique des Populations; Laboratoire Génome, Populations, Interactions, CNRS UMR 5000, Université de Montpellier II: Montpellier, France, 2004. [Google Scholar]
- Weir, B.S.; Cockerham, C.C. Estimating F-statistics for the analysis of population structure. Evolution 1984, 38, 1358–1370. [Google Scholar]
- Peakall, R.; Smouse, P.E. GenAlEx v.6.5: Genetic analysis in Excel. Population genetic software for teaching and research. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Weir, B.S. Genetic Data Analysis II; Sinauer Associates Inc.: Sunderland, MA, USA, 1997. [Google Scholar]


{kind=link}
| Treatment | First Visibility of the Disease | Epidemic Phase (d) | Disease Index | Average Apparent Infection Rate | ||||
|---|---|---|---|---|---|---|---|---|
| 2016 | 2017 | 2016 | 2017 | 2016 | 2017 | 2016 | 2017 | |
| Open field | 7/1 a | 7/7 a | 92 a | 86 a | 92.22 a | 64.89 a | 0.121 a | 0.112 a |
| Rain shelter | 7/29 c | 7/28 b | 64 c | 65 c | 16.58 b | 11.21 b | 0.114 a | 0.106 a |
| Fungicide | 7/15 b | 7/21 c | 78 b | 72 b | 9.92 c | 6.13 c | 0.089 b | 0.079 b |
| Treatment | Precipitation (mm) | Average Temperature (°C) | Average Relative Humidity (%) | Average Leaf Wetness Duration per Day (h) | ||||
|---|---|---|---|---|---|---|---|---|
| 2016 | 2017 | 2016 | 2017 | 2016 | 2017 | 2016 | 2017 | |
| Open field | 428.6 a | 290.3 a | 23.4 a | 23.3 a | 67.7 a | 62.1 a | 2.7 a | 1.7 a |
| Rain shelter | 0 b | 0 b | 23.8 a | 23.7 a | 60.1 b | 57.5 b | 0.4 b | 0.4 b |
| Fungicide | 425.9 a | 277.7 a | 23.7 a | 23.5 a | 65.0 a | 61.3 a | 2.5 a | 1.6 a |
| Cultivation Mode | Date | Collecting Method | Lesion Collected | Number of Genotypes | G1 (%) | G2 (%) | G3 (%) | Single Genotypes (%) | Other Genotypes (%) | I |
|---|---|---|---|---|---|---|---|---|---|---|
| 2016 Open field | 1 July | TSS | 1 | 1 | 1 | - | - | - | - | 0.594 |
| 15 July | TSS | 316 | 281 | 31 | 9 | 5 | 144 | 92 | 0.967 | |
| 29 July | PSS | 554 | 504 | 116 | 47 | 21 | 213 | 107 | 1.097 | |
| 12 August | PSS | 1102 | 1019 | 273 | 126 | 84 | 387 | 149 | 1.270 | |
| 26 August | PSS | 225 | 204 | 58 | 22 | 15 | 61 | 48 | 1.263 | |
| 9 September | PSS | 116 | 101 | 22 | 18 | 7 | 33 | 21 | 1.243 | |
| Total | 2314 | 2110 | 501 (23.7) | 222 (10.5) | 132 (6.3) | 838 (39.7) | 417 (19.8) | 1.226 | ||
| 2016 Fungicide | 15 July | TSS | 22 | 19 | 2 | 1 | - | 11 | 3 | 0.906 |
| 29 July | TSS | 78 | 70 | 5 | 3 | 2 | 54 | 6 | 1.079 | |
| 12 August | TSS | 189 | 162 | 11 | 8 | 3 | 128 | 12 | 1.109 | |
| 26 August | TSS | 125 | 116 | 8 | 4 | 1 | 95 | 8 | 1.265 | |
| 9 September | TSS | 62 | 56 | 4 | 5 | 2 | 41 | 4 | 1.261 | |
| Total | 476 | 423 | 30 (7.1) | 21 (5.0) | 8 (1.9) | 329 (77.8) | 33 (7.8) | 1.222 | ||
| 2016 Rain shelter | 29 July | TSS | 2 | 2 | 2 | - | - | - | - | 0.594 |
| 12 August | TSS | 72 | 66 | 25 | 9 | 6 | 12 | 14 | 1.101 | |
| 26 August | PSS | 265 | 246 | 115 | 54 | 28 | 11 | 38 | 1.104 | |
| 9 September | PSS | 264 | 233 | 102 | 36 | 24 | 17 | 54 | 1.124 | |
| 23 September | PSS | 166 | 148 | 48 | 23 | 16 | 15 | 46 | 1.141 | |
| Total | 769 | 695 | 292 (42.0) | 122 (17.6) | 74 (10.6) | 55 (7.9) | 152 (21.9) | 1.124 | ||
| 2017 Open field | 7 July | TSS | 1 | 1 | 1 | - | - | - | - | 0.297 |
| 21 July | TSS | 183 | 167 | 18 | 11 | 5 | 95 | 38 | 0.896 | |
| 4 August | PSS | 636 | 524 | 113 | 49 | 27 | 219 | 116 | 1.060 | |
| 18 August | PSS | 598 | 557 | 137 | 65 | 28 | 195 | 132 | 1.118 | |
| 1 September | PSS | 129 | 120 | 27 | 12 | 4 | 53 | 24 | 1.121 | |
| Total | 1547 | 1369 | 296 (21.6) | 137 (10.0) | 64 (4.7) | 562 (41.1) | 310 (22.6) | 1.093 | ||
| 2017 Fungicide | 21 July | TSS | 13 | 13 | - | - | - | 13 | - | 0.779 |
| 4 August | TSS | 32 | 27 | 2 | - | - | 25 | - | 0.907 | |
| 18 August | TSS | 112 | 99 | 6 | 2 | - | 86 | 5 | 0.915 | |
| 1 September | TSS | 98 | 88 | 11 | 5 | 2 | 67 | 3 | 0.996 | |
| 15 September | TSS | 52 | 48 | 4 | 2 | 1 | 38 | 3 | 0.970 | |
| Total | 307 | 275 | 23 (8.4) | 9 (3.3) | 3 (1.1) | 229 (83.3) | 11 (4.0) | 0.958 | ||
| 2017 Rain shelter | 4 August | TSS | 5 | 4 | 2 | - | 2 | - | - | 0.743 |
| 18 August | TSS | 42 | 37 | 12 | 8 | 3 | 5 | 9 | 0.940 | |
| 1 September | PSS | 95 | 86 | 35 | 17 | 9 | 8 | 17 | 0.956 | |
| 15 September | PSS | 168 | 153 | 59 | 32 | 21 | 12 | 29 | 0.954 | |
| 29 September | PSS | 66 | 59 | 21 | 14 | 9 | 5 | 10 | 0.966 | |
| Total | 376 | 339 | 129 (38.1) | 71 (20.9) | 44 (13.0) | 30 (8.8) | 65 (19.2) | 0.964 |
| Population | Number of Alleles (k) | A | Ae | Pa | H | I | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| kPv7 | kPv13 | kPv14 | kPv16 | kPv17 | kPv31 | kPv39 | ||||||
| 2016 Open field | 5 | 4 | 3 | 3 | 5 | 4 | 3 | 4.143 | 3.257 | 2 | 0.674 | 1.226 |
| 2016 Fungicide | 5 | 3 | 3 | 3 | 7 | 4 | 3 | 4.000 | 3.390 | 2 | 0.680 | 1.122 |
| 2016 Rain shelter | 4 | 4 | 3 | 2 | 5 | 4 | 3 | 3.571 | 2.995 | 0 | 0.640 | 1.224 |
| 2017 Open field | 8 | 4 | 5 | 2 | 6 | 3 | 2 | 4.143 | 2.940 | 4 | 0.518 | 1.093 |
| 2017 Fungicide | 3 | 3 | 3 | 2 | 5 | 3 | 2 | 3.286 | 2.578 | 2 | 0.518 | 0.958 |
| 2017 Rain shelter | 3 | 3 | 5 | 2 | 5 | 3 | 2 | 3.286 | 2.542 | 0 | 0.444 | 0.964 |
| 2016 Open Field | 2016 Fungicide | 2016 Rain Shelter | 2017 Open Field | 2017 Fungicide | 2017 Rain Shelter | |
|---|---|---|---|---|---|---|
| 2016 Open field | - | 12.540 | 20.699 | 0.757 | 0.582 | 0.670 |
| 2016 Fungicide | 0.005 * | - | 5.159 | 0.740 | 0.570 | 0.654 |
| 2016 Rain shelter | 0.003 * | 0.012 * | - | 0.776 | 0.530 | 0.660 |
| 2017 Open field | 0.076 ns | 0.083 ns | 0.073 | - | 4.548 | 6.221 |
| 2017 Fungicide | 0.097 ns | 0.101 ns | 0.096 | 0.016 * | - | 1.548 |
| 2017 Rain shelter | 0.089 ns | 0.096 ns | 0.083 | 0.013 * | 0.047 * | - |
| Population | Pv7 | Pv13 | Pv14 | Pv16 | Pv17 | Pv31 | Pv39 | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ho/He | Fis | Das | Ho/He | Fis | Das | Ho/He | Fis | Das | Ho/He | Fis | Das | Ho/He | Fis | Das | Ho/He | Fis | Das | Ho/He | Fis | Das | |
| 2016 Open field | 0.76/0.79 | 0.05 | 291 | 0.72/0.76 | 0.05 | 218 | 0.33/0.49 | 0.32 | 122 | 0.68/0.66 | −0.04 | 251 | 0.89/0.94 | 0.05 | 162 | 0.81/0.83 | 0.02 | 241 | 0.64/0.65 | 0.02 | 174 |
| 2016 Fungicide | 0.77/0.72 | −0.07 | 292 | 0.59/0.64 | 0.07 | 218 | 0.55/0.64 | 0.14 | 122 | 0.53/0.56 | 0.04 | 251 | 0.85/0.80 | −0.07 | 166 | 0.71/0.75 | 0.05 | 241 | 0.50/0.56 | 0.11 | 174 |
| 2016 Rain shelter | 0.84/0.89 | 0.05 | 291 | 0.71/0.76 | 0.06 | 218 | 0.27/0.48 | 0.45 | 122 | 0.75/0.48 | −0.55 | 251 | 0.95/0.72 | −0.33 | 162 | 0.90/0.92 | 0.02 | 241 | 0.70/0.73 | 0.04 | 174 |
| 2017 Open field | 0.30/0.54 | 0.44 | 293 | 0.74/0.78 | 0.06 | 218 | 0.34/0.55 | 0.39 | 124 | 0.63/0.47 | −0.34 | 251 | 0.47/0.62 | 0.24 | 168 | 0.75/0.77 | 0.02 | 245 | 0.27/0.34 | 0.20 | 174 |
| 2017 Fungicide | 0.45/0.54 | 0.20 | 293 | 0.59/0.53 | 0.06 | 218 | 0.38/0.54 | 0.45 | 124 | 0.55/0.50 | −0.55 | 251 | 0.65/0.77 | −0.33 | 168 | 0.51/0.53 | 0.02 | 245 | 0.48/0.49 | 0.04 | 174 |
| 2017 Rain shelter | 0.12/0.58 | 0.79 | 293 | 0.68/0.63 | −0.08 | 218 | 0.33/0.66 | 0.50 | 124 | 0.70/0.48 | −0.45 | 251 | 0.30/0.68 | 0.56 | 168 | 0.84/0.91 | 0.08 | 245 | 0.12/0.21 | 0.41 | 174 |
| Source of Variation | df | Sum of Squares | Estimate Variance | Percentage of Variation |
|---|---|---|---|---|
| Between years | 1 | 4458.657 | 0.742 | 10.73 |
| Among populations within years | 2 | 808.672 | 0.230 | 4.07 |
| Within Populations | 5205 | 24,672.350 | 4.875 | 85.20 |
| Total | 5208 | 29,939.680 | 5.847 | 100.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, S.; Li, B.; Guan, T.; Liu, L.; Wang, H.; Liu, C.; Zang, C.; Huang, Y.; Liang, C. A Comparison of Three Types of “Vineyard Management” and Their Effects on the Structure of Plasmopara viticola Populations and Epidemic Dynamics of Grape Downy Mildew. Plants 2022, 11, 2175. https://doi.org/10.3390/plants11162175
Yu S, Li B, Guan T, Liu L, Wang H, Liu C, Zang C, Huang Y, Liang C. A Comparison of Three Types of “Vineyard Management” and Their Effects on the Structure of Plasmopara viticola Populations and Epidemic Dynamics of Grape Downy Mildew. Plants. 2022; 11(16):2175. https://doi.org/10.3390/plants11162175
Chicago/Turabian StyleYu, Shuyi, Baihong Li, Tianshu Guan, Li Liu, Hui Wang, Changyuan Liu, Chaoqun Zang, Yuqian Huang, and Chunhao Liang. 2022. "A Comparison of Three Types of “Vineyard Management” and Their Effects on the Structure of Plasmopara viticola Populations and Epidemic Dynamics of Grape Downy Mildew" Plants 11, no. 16: 2175. https://doi.org/10.3390/plants11162175
APA StyleYu, S., Li, B., Guan, T., Liu, L., Wang, H., Liu, C., Zang, C., Huang, Y., & Liang, C. (2022). A Comparison of Three Types of “Vineyard Management” and Their Effects on the Structure of Plasmopara viticola Populations and Epidemic Dynamics of Grape Downy Mildew. Plants, 11(16), 2175. https://doi.org/10.3390/plants11162175