
Assessing Phenotypic Variability in Some Eastern European Insular Populations of the Climatic Relict Ilex aquifolium L.

 , ,
, ,
Abstract
1. Introduction
2. Results
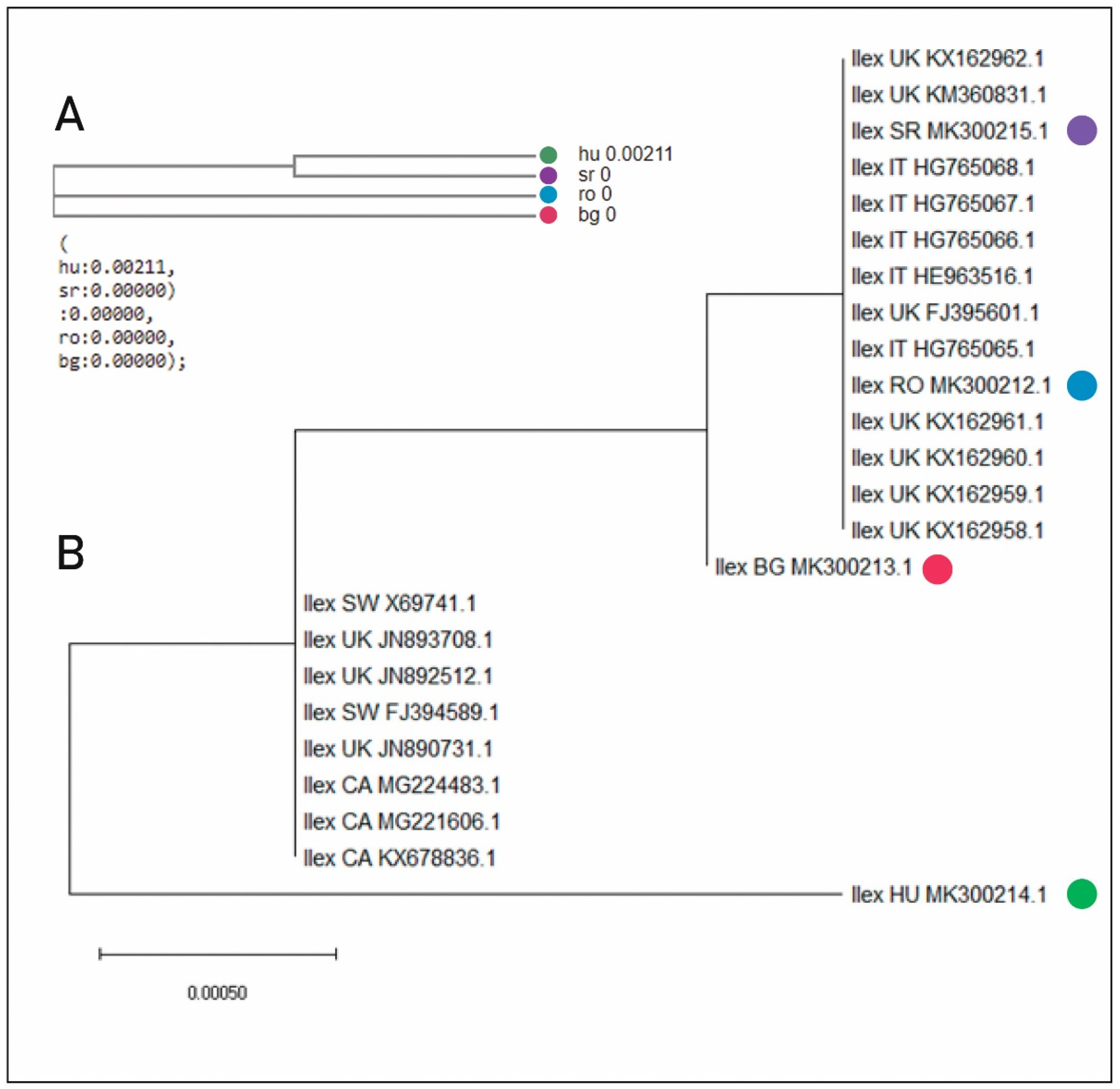
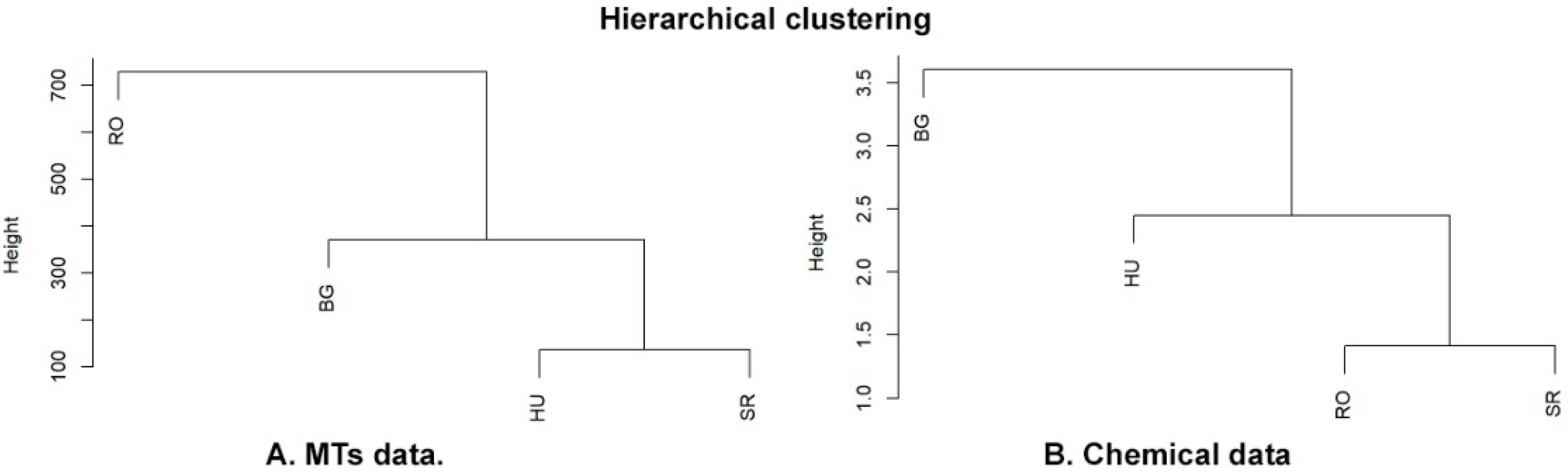
2.1. Molecular Analysis
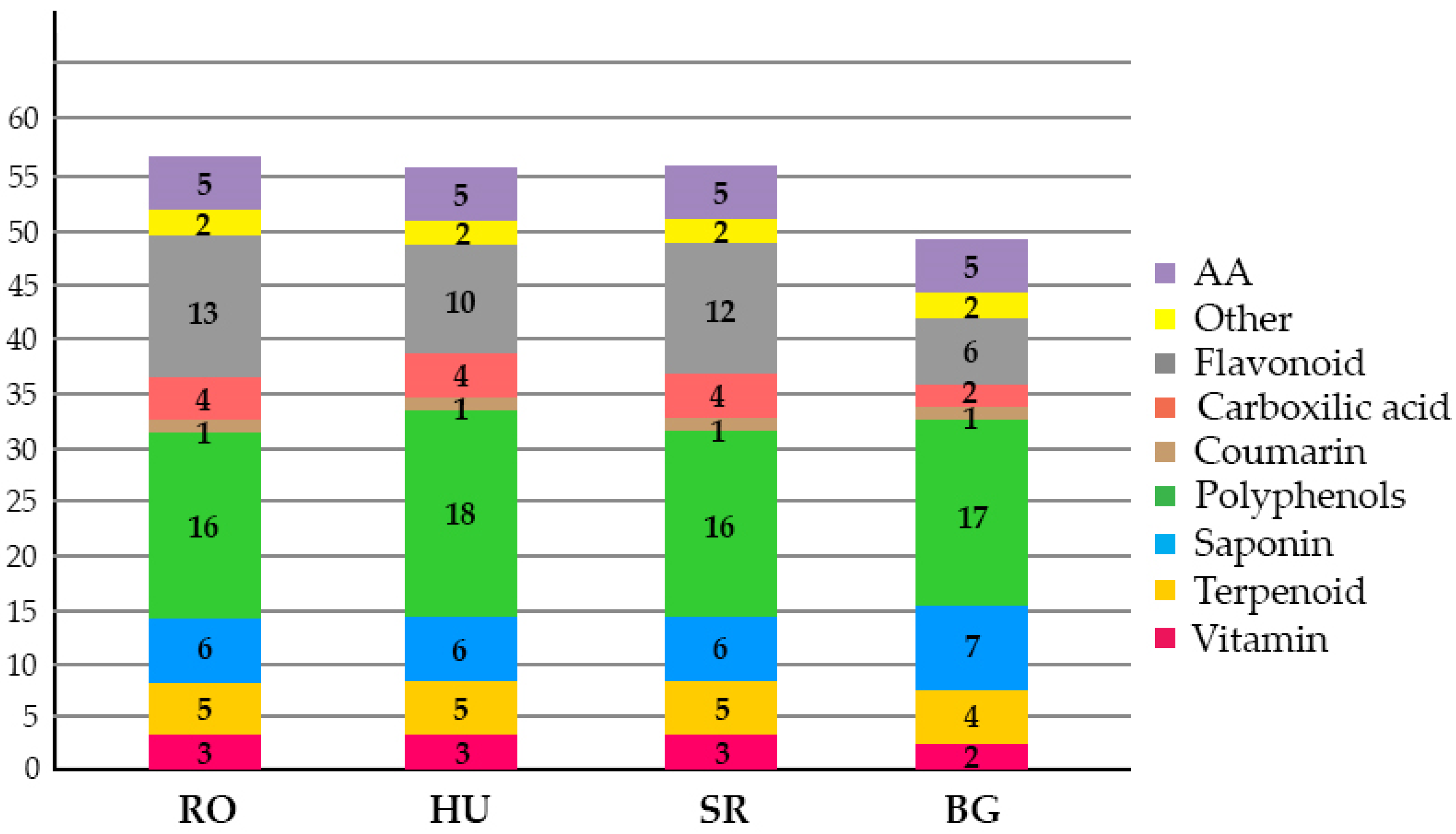
2.2. Leaf-Specific Phytochemical Profiles
2.3. Morphological Traits by SEM Descriptive Analysis
2.3.1. Leaf
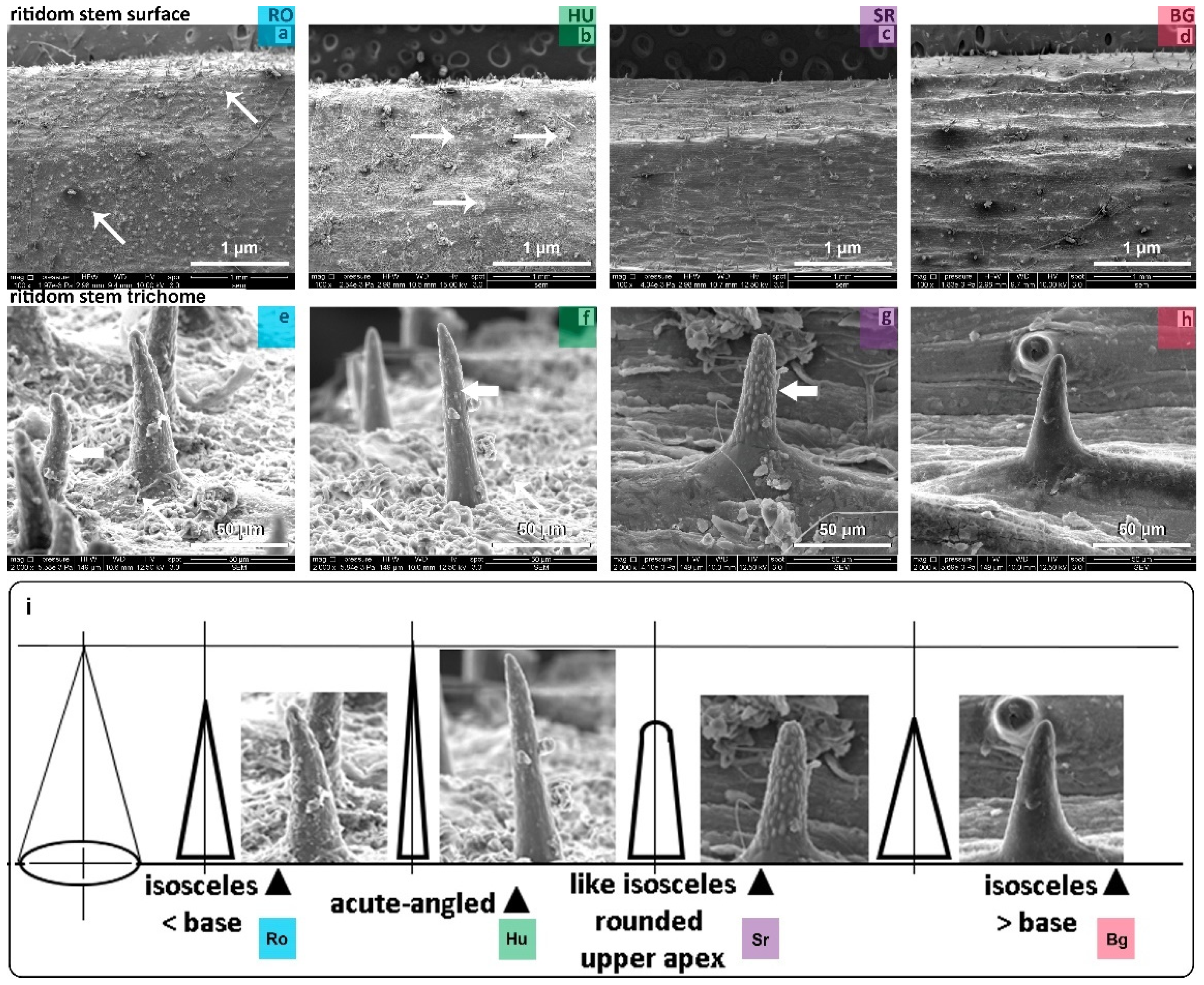
2.3.2. STEM Surface and Trichomes
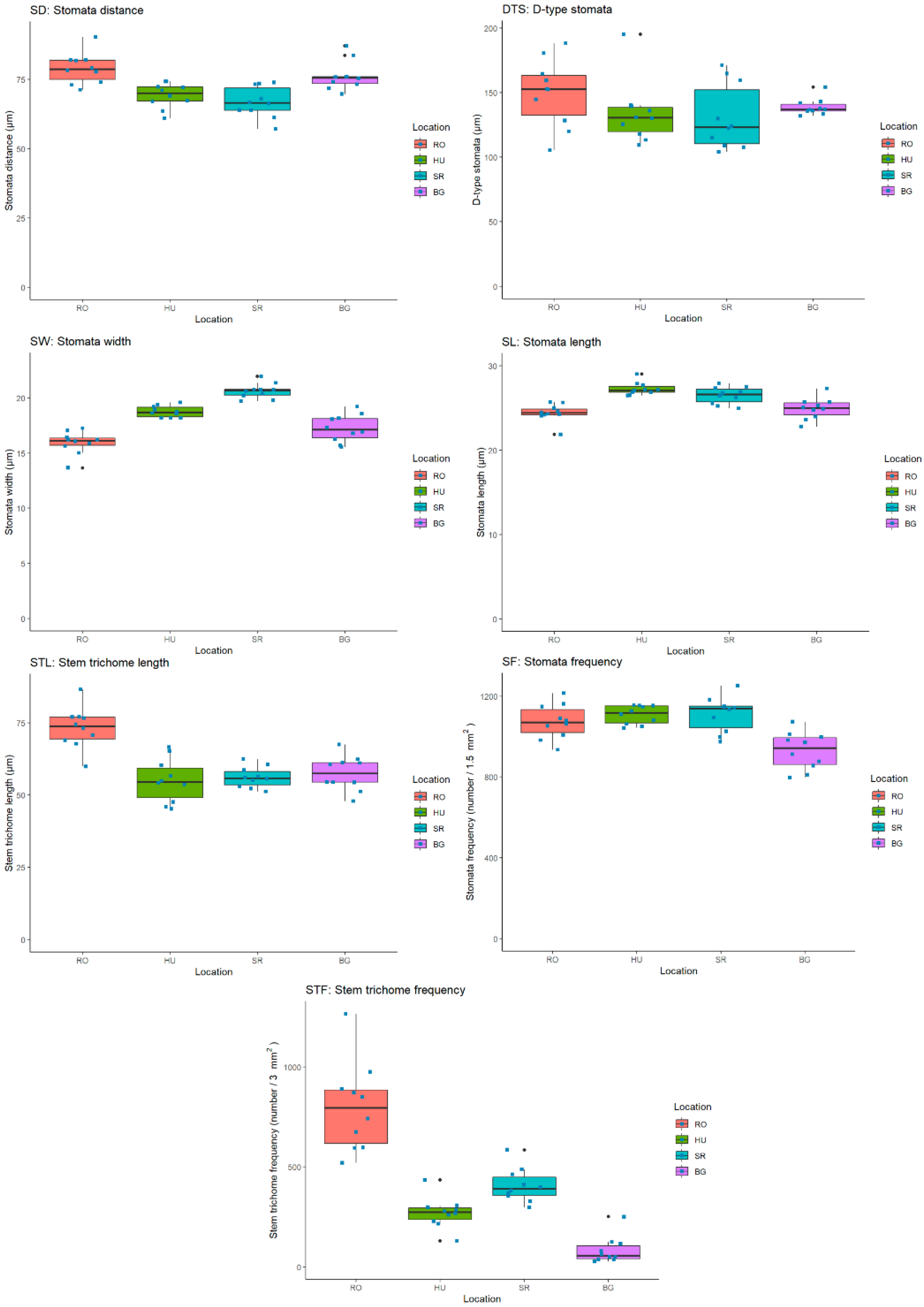
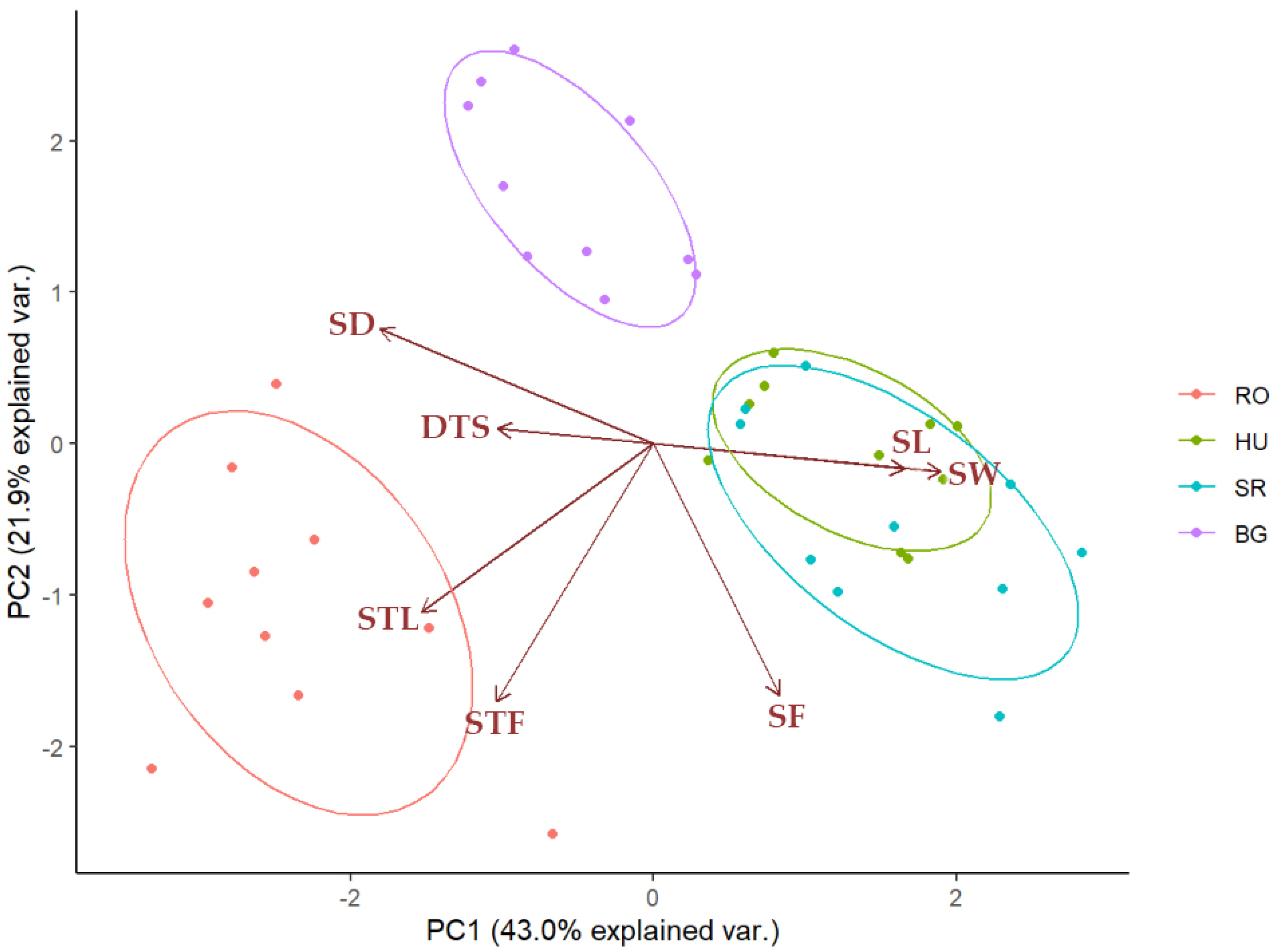
2.4. Quantification and Statistical Analysis of Leaf- and Stem-Specific Morphological Traits
3. Discussion
4. Materials and Methods
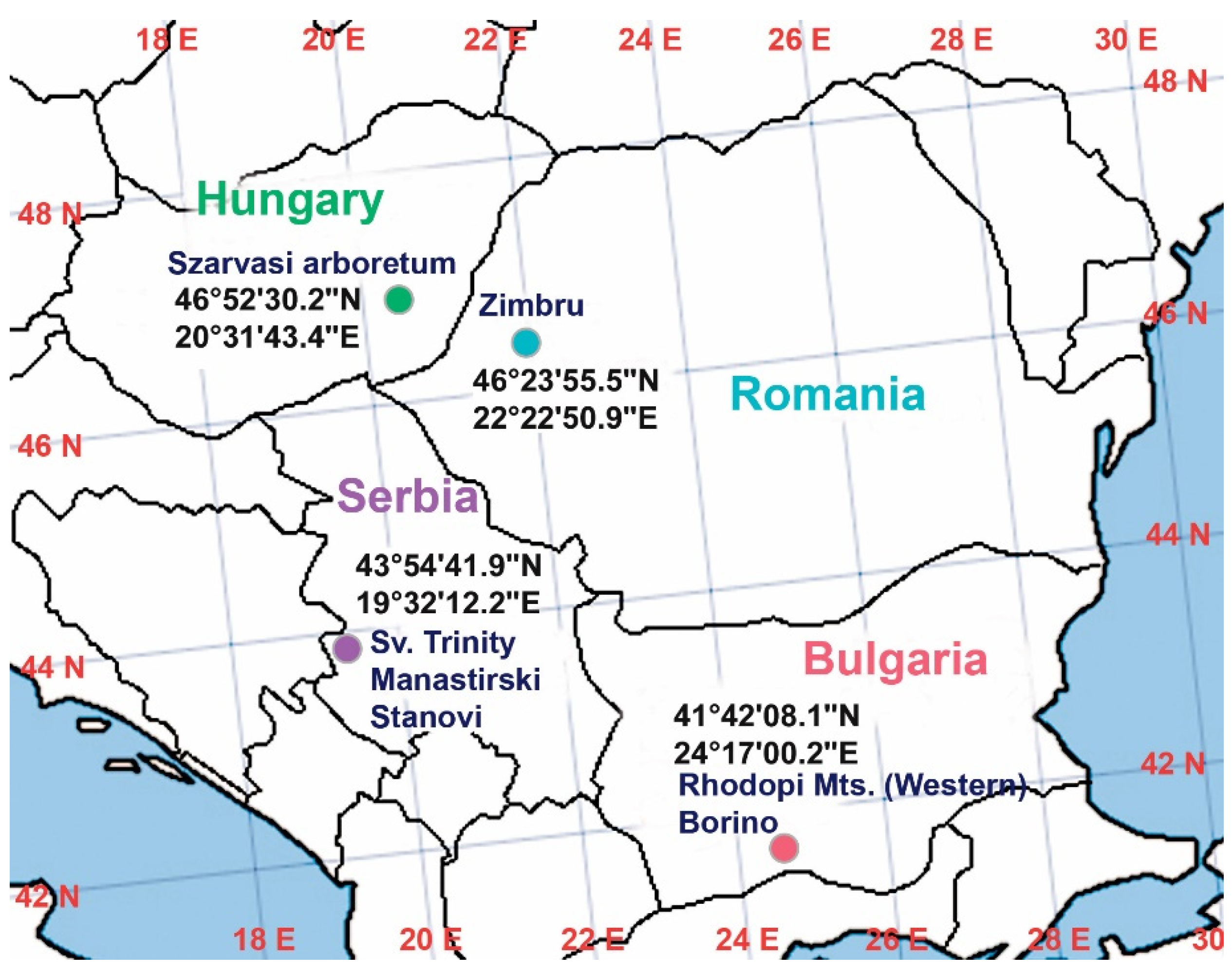
4.1. Taxon Sampling
4.2. DNA Extraction, PCR Amplification and Sequencing
4.3. Phytochemicals Analysis
4.4. Morphology Analysis and Morphometry
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peterken, G.F. Woodland Conservation and Management, 1st ed.; Reprint; Chapman and Hall: London, UK, 1981. [Google Scholar]
- Nimis, P.L.; Bolognini, G. Quantitative phytogeography of the Italian Beech Forests. Vegetatio 1993, 109, 125–143. [Google Scholar] [CrossRef]
- Pulido, F.; Valladares, F.; Calleja, J.A.; Moreno, G.; Gonzáles-Bornay, G. Tertiary relict trees in a Mediterranean climate: Abiotic constraints on the persistance of Prunus lusitanica at the eroding edge of its range. J. Biogeogr. 2008, 35, 1425–1435. [Google Scholar] [CrossRef]
- Manen, J.F.; Barriera, G.; Loizeau, P.A.; Naciri, Y. The history of extant Ilex species (Aquifoliacae): Evidence of hybridization within a Miocene radiation. Mol. Phylogenetics Evol. 2010, 57, 961–977. [Google Scholar] [CrossRef] [PubMed]
- Loizeau, P.A.; Barriera, G.; Manen, J.F.; Broennimann, O. Towards an understanding of the distribution of Ilex L. (Aquifoliaceae) on a World-wide scale. Biol. Skr. 2005, 55, 501–520. [Google Scholar]
- Soutar, R.G.; Spencer, J.W. The Conservation of Genetic Variation in Britain’s Native Trees. For. Int. J. For. Res. 1991, 64, 1–12. [Google Scholar]
- Gottlieb, A.M.; Giberti, G.C.; Poggio, L. Molecular analyses of the genus Ilex (Aquifoliaceae) in Southern South America, Evidence from AFLP and ITS sequence data. Am. J. Bot. 2005, 92, 352–369. [Google Scholar] [CrossRef]
- Setoguchi, H.; Watanabe, I. Intersectional gene flow between insular endemics of Ilex (Aquifoliaceae) on the Bonin Islands and the Ryukyu Islands. Am. J. Bot. 2000, 87, 793–810. [Google Scholar] [CrossRef]
- Herrera, C.M.; Bazaga, P. Epigenetic correlates of plant phenotypic plasticity: DNA methylation differs between prickly and nonprickly leaves in heterophyllous Ilex aquifolium (Aquifoliaceae) trees. Bot. J. Linn. Soc. 2013, 171, 441–452. [Google Scholar] [CrossRef]
- Yi, F.; Sun, L.; Hao, D.C.; Peng, Y.; Han, F.M.; Xiao, P.G. Complex phylogenetic placement of Ilex species (Aquifoliaceae): A case study of molecular phylogeny. Pak. J. Bot. 2017, 49, 215–225. [Google Scholar]
- Rendell, S.; Ennos, R.A. Chloroplast DNA diversity of the dioecious European tree Ilex aquifolium L. (English holly). Mol. Ecol. 2003, 12, 2681–2688. [Google Scholar] [CrossRef]
- Hollingsworth, P.M.; Graham, S.W.; Little, D.P. Choosing and using a plants DNA barcode. PLoS ONE 2011, 6, e19254. [Google Scholar] [CrossRef] [PubMed]
- Aranda, I.; Robson, T.M.; Rodríguez-Calcerrada, J.; Valladares, F. Limited capacity to cope with excessive light in the open and seasonal drought in the shade in Mediterranean Ilex aquifolium populations. Trees 2008, 22, 375–384. [Google Scholar] [CrossRef]
- Arrieta, S.; Suárez, F. Marginal holly (Ilex aquifolium L.) populations in Mediterranean central Spain are constrained by a low-seedling recruitment. Flora 2006, 201, 152–160. [Google Scholar] [CrossRef]
- San-Miguel, A.J.; de Rigo, D.; Caudullo, G.; Houston, D.T.; Mauri, A. European Atlas of Forest Tree Species; Publication Office of European Union: Luxembourg, 2016. [Google Scholar]
- Peterken, G.F.; Lloyd, P.S. Biological Flora of the British Isles, Ilex aquifolium L., List of the British Vascular Plant. J. Ecol. 1967, 55, 841–858. [Google Scholar] [CrossRef]
- Valladares, F.; Arrieta, S.; Aranda, I.; Lorenzo, D.; Sánchez-Gómez, D.; Tena, D.; Suárez, F.; Pardos, J.A. Shade tolerance, photoinhibition sensitivity and phenotypic plasticity of Ilex aquifolium in continental Mediterranean sites. Tree Physiol. 2005, 25, 1041–1052. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.; Batic, F.; Pfanz, H. Photosynthetic performance of leaves and twigs of evergreen holly (Ilex aquifolium L.). Phyton 2000, 40, 179–190. [Google Scholar]
- Obeso, J.R. Costs of reproduction in Ilex aquifolium: Effects at tree, branch and leaf levels. J. Ecol. 1997, 75, 159–166. [Google Scholar] [CrossRef]
- Collado, J.; Platas, G.; Peláez, F. Fungal endophytes in leaves, twigs and bark of Quercus ilex from Central Spain. Nova Hedwig. 1996, 63, 347–360. [Google Scholar]
- Nahar, L.; Russell, W.R.; Middleton, M.; Shoeb, M.; Sarker, S.D. Antioxidant phenylacetic acid derivates from the seeds of Ilex aquifolium. Acta Pharm. 2005, 55, 187–193. [Google Scholar]
- Zwyrzykowska, A.; Kupczyński, R.; Jarosz, B.; Szumny, A.; Kucharska, A.Z. Qualitative and quantitative analysis of polyphenolic compounds in Ilex sp. Open Chem. 2015, 13, 1303–1312. [Google Scholar] [CrossRef]
- Hao, D.; Gu, X.; Xiao, P.; Liang, Z.; Xu, L.; Peng, Y. Research progress in the phytochemistry and biology of Ilex pharmaceutical resources. Acta Pharm. Sin. 2013, 3, 8–19. [Google Scholar] [CrossRef]
- Skrøppa, T. State of Forest Genetic Resources in Norway, Norwegian Country Report to the preparation of the FAO Report on The State of the World’s Forest Genetic Resources. Norwegian Genetic Resource Center, Norwegian Forest and Landscape Institute. 2012. Available online: https://nibio.brage.unit.no/nibio-xmlui/bitstream/handle/11250/2453902/SoL-Rapport-2012-03.pdf?sequence=2&isAllowed=y (accessed on 19 April 2019).
- Gaskell, T.T. Flora Europaea, Vol. 2, Rosaceae to Umbelliferae; Cambridge University Press: Cambridge, UK, 1968. [Google Scholar]
- Kelcey, J.G.; Müller, N. Plants and Habitats of European Cities; Springer: New York, NY, USA, 2011. [Google Scholar]
- Ciocârlan, V. Flora Ilustrată a României: Pteridophyta et Spermathophyta; CeresEd: București, Romania, 2009. [Google Scholar]
- Valdés, B. Aquifoliaceae. Euro + Med Plantbase—The Information Resource for Euro-Mediterranean Plant Diversity. 2012. Available online: http://euromed.luomus.fi/euromed_map.php?taxon=435085&size=medium (accessed on 13 June 2019).
- Available online: https://www.iucnredlist.org/species/202963/68067360#habitat-ecology (accessed on 18 July 2018).
- Available online: https://library.hungaricana.hu/hu/view/SZAK_BAKO_Muzealis_Magyarorszag_viragos_novenyei/?pg=0&layout=s (accessed on 22 June 2020).
- Sütöri-Diószegi, M.; Schmidt, G. Urban Effects and Escaping of Exotic Woody Plants in the Buda Arboretum; Slovaca Universitas Agrogulturae: Budapest, Hungary, 2010; pp. 27–32. [Google Scholar]
- Simonkai, L. Aradvármegye és Arad Szabad királyi város, Természetrajzi Leirása, Monographiája; Kiadja a Monographia-Bizottság: Arad, Romania, 1893. [Google Scholar]
- Bedö, A.; Horváth, S. Erdészeti Lapok, az Országos Erdészeti Egyesület, Közlönye, Erdó és földbirtokosok, erdészeti ügyekkel foglalkozor és erdötisztek számára. Harminczkettedik Évfolya; Az Országgyélési Ertesitö kö es könyvnyomdája részv: Budapest, Hungary, 1893; pp. I–XII. [Google Scholar]
- Simonkai, L. Enumeratio Florae Transsilvanicae Versculoae Critica: Ex Mandato Societatis Scientiarum Naturalium Regiae Hungaricae; Nabu Press: Charleston, SC, USA, 1886. [Google Scholar]
- Muica, E.C.; Popova-Cucu, A. The composition and conservation of Romania’s plant cover. GeoJournal 1993, 29, 9–18. [Google Scholar] [CrossRef]
- UNEP-WCMC and IUCN. Protected Planet: Dosul Laurului in Romania. The World Database on. Protected Areas WDPA: 14574/The Global Database on Protected Areas Management Effectiveness. GD-PAME [On-line], UNEP-WCMC/IUCN, Cambridge, UK. 1999. Available online: www.protectedplanet.net (accessed on 3 June 2018).
- Available online: https://www.iucnredlist.org/species/202963/2758279 (accessed on 18 April 2019).
- Hanyecz, K. Szarvasi arboretum. Budapest: A projekt címe: PRO-PLANTS A projekt száma: HURO/1101/119.1.3.1. 2013. Available online: https://macea.uvvg.ro/muzeul-botanic/ (accessed on 23 June 2019).
- Liu, J.; Möller, M.; Gao, L.M.; Zhang, D.Q.; Li, D.Z. DNA barcoding for the discrimination of Eurasian yews (Taxus L., Taxaceae) and the discovery of cryptic species. Mol. Ecol. Resour. 2011, 11, 89–100. [Google Scholar] [CrossRef]
- Bafeel, S.O.; Arif, I.A.; Bakir, M.A.; Al Homaidan, A.A.; Al Farhan, A.H.; Khan, H.A. DNA barcoding of arid wild plants using rbcL gene sequences. Genet Mol. Res. 2012, 11, 1934–1941. [Google Scholar] [CrossRef] [PubMed]
- Majada, J.P.; Sánchez-Tamés, R.; Revilla, M.A.; Casares, A. Micropropagation of Ilex aquifolium L. In Vitro Cellular and Developmental Biology. Plant 2000, 36, 521–526. [Google Scholar]
- Marchiosi, R.; dos Santos, W.D.; Constantin, R.P.; de Lima, R.B.; Soares, A.R.; Finger-Teixeira, A.; Mota, T.R.; de Oliveira, D.M.; Foletto-Felipe, M.d.P.; Abrahão, J. Biosynthesis and metabolic actions of simple phenolic acids in plants. Phytochem. Rev. 2020, 19, 865–906. [Google Scholar] [CrossRef]
- Paluch, E.; Okińczyc, P.; Zwyrzykowska-Wodzińska, A.; Szperlik, J.; Zarowska, B.; Duda-Madej, A.; Babelewski, P.; Wlodarczyk, M.; Wojtasik, W.; Kupczyński, R. Composition and Antimicrobial Activity of Ilex Leaves Water Extracts. Molecules 2021, 26, 7442. [Google Scholar] [CrossRef]
- Sudhir, K.K.; Satish, C.S.; Mohan, S.M.R.; Manisha, D.S.; Deepak, K.S.; Ruchi, B.S.; Amita, S.; Bipin, R.; Ashok, K. Chemical constituents and biological significance of the genus Ilex (Aquifoliaceae). Nat. Prod. J. 2012, 2, 212–224. [Google Scholar]
- Anand, P.; Samit, R.; Amit, R. Molecular Markers in Phylogenetic Studies-A Review. J. Phylogenetics Evol. Biol. 2014, 2, 1–9. [Google Scholar]
- Zahariev, D. Biodiversity of Relict Vascular Plants in Bulgaria. Int. J. Res. Stud. Biosci. 2016, 4, 38–51. [Google Scholar]
- Molnár, A.; Végvári, Z.; Tóthmérész, B. Identification of floral relicts based on spatial distance of isolation. Forests 2017, 8, 459. [Google Scholar] [CrossRef]
- Varga, Z. Extra-Mediterranean Refugia, Post-Glacial Vegetation History and Area Dynamics in Eastern Central Europe. In Relict Species: Phylogeography and Conservation Biology; Habel, J.C., Assmann, T., Eds.; Springer Science & Business Media: New York, NY, USA, 2009. [Google Scholar]
- Vukojičić, S.; Jakovljević, K.; Matevski, V.; Randjelović, V.; Niketić, M.; Lakušić, D. Distribution, Diversity and Conservation of Boreo-Montane Plant Species in the Central Part of the Balkan Peninsula and the Southern Part of the Pannonian Plain. Folia Geobot. 2014, 49, 487–505. [Google Scholar] [CrossRef]
- Yao, X.; Tan, Y.H.; Yang, J.B.; Wang, Y.; Corlett, R.T.; Manen, J.F. Exceptionally high rates of positive selection on the rbcL gene in the genus Ilex (Aquifoliaceae). BMC Evol. Biol. 2019, 19, 192. [Google Scholar] [CrossRef] [PubMed]
- Bojić, M.; Haas, V.S.; Šarić, D.; Maleš, Ž. Determination of Flavonoids, Phenolic Acids, and Xanthines in Mate Tea (Ilex paraguariensis St.-Hil.). J. Anal. Methods Chem. 2016, 2013, 658596. [Google Scholar] [CrossRef]
- Heck, C.I.; De Mejia, E.G. Yerba Mate Tea (Ilex paraguariensis): A Comprehensive Review on Chemistry, Health Implications, and Technological Considerations. J. Food Sci. 2007, 72, R138–R151. [Google Scholar] [CrossRef] [PubMed]
- Peres, R.G.; Tonin, F.G.; Tavares, M.F.; Rodriguez-Amaya, D.B. HPLC-DAD-ESI/MS Identification and Quantification of Phenolic Compounds in Ilex paraguariensis Beverages and On-Line Evaluation of Individual Antioxidant Activity. Molecules 2013, 18, 3859–3871. [Google Scholar] [CrossRef]
- Jaiswal, R.; Sovdat, T.; Vivan, F.; Kuhnert, N. Profiling and characterization by LC-MSn of the chlorogenic acids and hydroxycinnamoylshikimate esters in maté (Ilex paraguariensis). J. Agric. Food Chem. 2010, 58, 5471–5484. [Google Scholar] [CrossRef] [PubMed]
- Butiuk, A.P.; Martos, M.A.; Adachi, O.; Hours, R.A. Study of the chlorogenic acid content in yerba mate (Ilex paraguariensis St. Hil.): Effect of plant fraction, processing step and harvesting season. J. Appl. Res. Med. Aromat. Plants 2016, 3, 27–33. [Google Scholar] [CrossRef]
- Available online: https://drugs.ncats.io/drug/6RO47OBY4X (accessed on 16 June 2022).
- Sugimoto, S.; Nakamura, S.; Yamamoto, S.; Yamashita, C.; Oda, Y.; Matsuda, H.; Yoshikawa, M. Brazilian natural medicines. III. structures of triterpene oligoglycosides and lipase inhibitors from mate, leaves of Ilex paraguariensis. Chem. Pharm. Bull. 2009, 57, 257–261. [Google Scholar] [CrossRef]
- Coelho, G.C.; Gnoatto, S.B.; Bassani, V.L.; Schenkel, E.P. Quantification of saponins in extractive solution of mate leaves (Ilex paraguariensis A. St. Hil.). J. Med. Food 2010, 13, 439–443. [Google Scholar] [CrossRef]
- Acevedo, R.M.; Avico, E.H.; González, S.; Salvador, A.R.; Rivarola, M.; Paniego, N.; Nunes-Nesi, A.; Ruiz, O.A.; Sansberro, P.A. Transcript and metabolic adjustments triggered by drought in Ilex paraguariensis leaves. Planta 2019, 250, 445–462. [Google Scholar] [CrossRef] [PubMed]
- Valenca, S.S.; Lanzetti, M.; Porto, L.C. Mate Tea: From the Camp to the Bench. In Tea in Health and Disease Prevention; Academic Press: Cambridge, MA, USA, 2013; pp. 161–170. [Google Scholar] [CrossRef]
- Available online: http://www.thegoodscentscompany.com/data/rw1296251.html (accessed on 18 May 2022).
- Kim, D.K.; Nam, I.Y.; Kim, J.W.; Shin, T.Y.; Lim, J.P. Pentacyclic triterpenoids fromllex macropoda. Arch. Pharmacal Res. 2002, 25, 617–620. [Google Scholar] [CrossRef] [PubMed]
- Radice, M.; Scalvenzi, L.; Cossio, N.S. Ilex guayusa: A systematic review of its Traditional Uses, Chemical Constituents, Biological Activities and Biotrade Opportunities. In Proceedings of the MOL2NET’16, Conference on Molecular, Biomedical & Computational Sciences and Engineering, 2nd ed.; MDPI: Basel, Switzerland, 2017; Section M; p. 3868. [Google Scholar] [CrossRef]
- Kang, Y.F.; Wu, H.M.; Chen, S.J.; Chen, W.Y.; Li, H.T. Secondary Metabolites from the Leaves of Ilex Cornuta. Chem. Nat. Compd. 2014, 50, 355–356. [Google Scholar] [CrossRef]
- Tori, M.; Mukai, Y.; Nakashima, K.; Sono, M. A Carboxylic Acid from Ilex integra. Molecules 2003, 8, 882–885. [Google Scholar] [CrossRef]
- Chen, Y.-L.; Hwang, T.-L.; Yu, H.-P.; Fang, J.-Y.; Chong, K.Y.; Chang, Y.-W.; Chen, C.-Y.; Yang, H.-W.; Chang, W.-Y.; Hsieh, P.-W. Ilex kaushue and Its Bioactive Component 3,5-Dicaffeoylquinic Acid Protected Mice from Lipopolysaccharide-Induced Acute Lung Injury. Sci. Rep. 2016, 6, 34243. [Google Scholar] [CrossRef]
- Kuropka, P.; Zwyrzykowska-Wodzińska, A.; Kupczyński, R.; Włodarczyk, M.; Szumny, A.; Nowaczyk, R.M. The Effect of Ilex × meserveae S. Y. Hu Extract and Its Fractions on Renal Morphology in Rats Fed with Normal and High-Cholesterol Diet. Foods 2021, 10, 818. [Google Scholar] [CrossRef]
- Palu, D.; Bighelli, A.; Casanova, J.; Paoli, M. Identification and Quantitation of Ursolic and Oleanolic Acids in Ilex aquifolium L. Leaf Extracts Using 13C and 1H-NMR Spectroscopy. Molecules 2019, 24, 4413. [Google Scholar] [CrossRef]
- Pachura, N.; Kupczyński, R.; Sycz, J.; Kuklińska, A.; Zwyrzykowska-Wodzińska, A.; Wińska, K.; Owczarek, A.; Kuropka, P.; Nowaczyk, R.; Bąbelewski, P.; et al. Biological Potential and Chemical Profile of European Varieties of Ilex. Foods 2021, 11, 47. [Google Scholar] [CrossRef]
- Yang, L.; Wen, K.S.; Ruan, X.; Zhao, Y.X.; Wei, F.; Wang, Q. Response of Plant Secondary Metabolites to Environmental Factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef]
- Available online: https://www.worldweatheronline.com/ (accessed on 16 June 2022).
- Nogués, S.; Allen, D.J.; Morison, J.I.; Baker, N.R. Ultraviolet-B radiation effects on water relations, leaf development, and photosynthesis in droughted pea plants. Plant Physiol. 1998, 117, 173–181. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Drought stress enhances nutritional and bioactive compounds, phenolic acids and antioxidant capacity of Amaranthus leafy vegetable. BMC Plant Biol. 2018, 18, 258. [Google Scholar] [CrossRef] [PubMed]
- Jaafar, H.Z.; Ibrahim, M.H.; Karimi, E. Phenolics and flavonoids compounds, phenylanine ammonia lyase and antioxidant activity responses to elevated CO₂ in Labisia pumila (Myrisinaceae). Molecules 2012, 17, 6331–6347. [Google Scholar] [CrossRef] [PubMed]
- Holopainen, J.K.; Virjamo, V.; Ghimire, R.P.; Blande, J.D.; Julkunen-Tiitto, R.; Kivimäenpää, M. Climate Change Effects on Secondary Compounds of Forest Trees in the Northern Hemisphere. Front. Plant Sci. 2018, 9, 1445. Available online: https://www.frontiersin.org/articles/10.3389/fpls.2018.01445 (accessed on 23 April 2021). [CrossRef] [PubMed]
- Poljak, I.; Vahčić, N.; Liber, Z.; Šatović, Z.; Idžojtić, M. Morphological and Chemical Variation of Wild Sweet Chestnut (Castanea sativa Mill.) Populations. Forests 2022, 13, 55. [Google Scholar] [CrossRef]
- Jia, K.; Zhao, W.; Maier, P.A.; Hu, X.; Jin, Y.; Zhou, S.; Jiao, S.; A El-Kassaby, Y.; Wang, T.; Wang, X.; et al. Landscape genomics predicts climate change-related genetic offset for the widespread Platycladus orientalis(Cupressaceae). Evol. Appl. 2019, 13, 665–676. [Google Scholar] [CrossRef] [PubMed]
- Richard, I.M.; Richard, J.A. The Origin and Evolution of Tertiary Relict Floras; Advances in Botanical Research; Academic Press: Cambridge, MA, USA, 2002; Volume 38, pp. 281–314. ISBN 9780120059386. [Google Scholar] [CrossRef]
- Lendvay, B.; Kadereit, J.W.; Westberg, E.; Cornejo, C.; Pedryc, A.; Höhn, M. Phylogeography of Syringa josikaea (Oleaceae): Early Pleistocene divergence from East Asian relatives and survival in small populations in the Carpathians. Biol. J. Linn. Soc. 2015, 119, 689–703. [Google Scholar] [CrossRef]
- Bordács, S.; Popescu, F.; Slade, D.; Csaikl, U.M.; Lesur, I.; Borovics, A.; Kézdy, P.; König, A.O.; Gömöry, D.; Brewer, S.; et al. Chloroplast DNA variation of white oaks in northern Balkans and in the Carpathian Basin. For. Ecol. Manag. 2002, 156, 197–209. [Google Scholar] [CrossRef]
- Moldovan, I.C.; Sofletea, N.; Curtu, A.L.; Abrudan, I.V.; Postolache, D.; Popescu, F. Chloroplast DNA diversity of oak species in Eastern Romania. Not. Bot. Horti Agrobot. ClujNapoca 2010, 38, 302–307. [Google Scholar]
- Robert, B.; Dalibor, B.; Faruk, B.; Martin, B.; Marilena, I. Leaflet morphometric variation of service tree (Sorbus domestica L.) in the Balkan Peninsula. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2011, 145, 278–285. [Google Scholar] [CrossRef]
- Nikolić, D.; Šinžar-Sekulić, J.; Ranđelović, V.; Lakušić, D. Morphological variation of Jovibarba heuffelii (Crassulaceae) in the central Balkan Peninsula—The impact of geological, orographical and bioclimatic factors on the differentiation of populations. Phytotaxa 2015, 203, 213–230. [Google Scholar] [CrossRef][Green Version]
- Reed, D.W.; Tukey, H.B. Light-intensity and temperature effects on epicuticular wax morphology and internal cuticle ultrastructure of carnation and brusselssprouts leaf cuticles. J. Am. Soc. Hortic. Sci. 1982, 107, 417–420. [Google Scholar]
- Valkama, E.; Salminen, J.-P.; Koricheva, J.; Pihlaja, K. Changes in leaf trichomes and epicuticular flavonoids during leaf development in three birch taxa. Ann. Bot. 2004, 94, 233–242. [Google Scholar] [CrossRef]
- Wagner, G.J. New Approaches for Studying and Exploiting an Old Protuberance, the Plant Trichome. Ann. Bot. 2004, 93, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Mauricio, R. Ontogenetics of QTL: The genetic architecture of trichome density over time in Arabidopsis thaliana. Genetica 2005, 123, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Stuart, C.; Julie, E.G. Influence of environmental factors on stomatal development. New Phytol. 2008, 178, 9–23. [Google Scholar]
- Jane, E.C.; Christopher, A.A.; Kent, E.H. Intraspecific variation in stomatal traits, leaf traits and physiology reflects adaptation along aridity gradients in a South African shrub. Ann. Bot. 2016, 117, 195–207. [Google Scholar]
- Javier, G.A.; Mateu, F.P.; Pere, A.M.; Miquel, A.C.; Cathie, M.; Jeroni, G. The ratio of trichomes to stomata is associated with water use efficiency in Solanum lycopersicum (tomato). Plant J. 2018, 96, 607–619. [Google Scholar]
- Maloukh, L.; Kumarappan, A.; Jarrar, M.; Salehi, J.; El-Wakil, H.; Lakshmi, T.V.R. Discriminatory power of rbcL barcode locus for authentication of some of United Arab Emirates (UAE) native plants. 3 Biotech 2017, 7, 144. [Google Scholar] [CrossRef]
- Samarakoon, T.; Wang, S.Y.; Alford, M.H. Enhancing PCR Amplification of DNA from Recalcitrant Plant Specimens Using a Trehalose-Based Additive. Appl. Plant Sci. 2013, 1, 1200236. [Google Scholar] [CrossRef]
- Available online: https://www.macrogen-europe.com/services/sanger-sequencing/standard (accessed on 30 June 2019).
- McGinnis, S.; Madden, T.L. BLAST: At the core of a powerful and diverse set of sequence analysis tools. Nucleic Acids Res. 2004, 32, W20–W25. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [PubMed]
- Nihal, T.E.; Ferda, S.; Gökçe, P.; Sedat, Y.V. Antioxidant and Antibacterial Activities of Various Extracts and Fractions of Fresh Tea Leaves and Green Tea. Tarim Bilimleri Derg. 2009, 15, 371–378. [Google Scholar] [CrossRef]
- Vígh, S.; Cziaky, Z.; Sinka, L.T.; Pribac, C.; Moş, L.; Turcuş, V.; Remenyik, J.; Mathe, E. Analysis of phytoconstituent profile of fenugreek–Trigonella foenuem-graecum L.-seed extracts. Studia Ubb Chem. 2017, 62, 145–166. [Google Scholar] [CrossRef]
- Available online: https://scandium.software.informer.com/ (accessed on 30 June 2018).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 19 April 2022).
- Revelle, W. psych: Procedures for Personality and Psychological Research; Northwestern University: Evanston, IL, USA, 2022; Available online: https://cran.microsoft.com/snapshot/2022-04-12/web/packages/psych/index.html (accessed on 20 April 2022).
- Alboukadel, K. rstatix: Pipe-Friendly Framework for Basic Statistical Tests. R package Version 0.7.0. 2021. Available online: https://CRAN.R-project.org/package=rstatix (accessed on 20 April 2022).
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testin. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Jari, O.; Gavin, L.S.; Guillaume, B.; Roeland, K.; Pierre, L.; Peter, R.M.; O’Hara, R.B.; Peter, S.; Stevens, H.M.H.; Eduard, S.; et al. vegan: Community Ecology Package. R Package Version 2.6-2. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 20 April 2022).
- Available online: https://www.worldclim.org/data/bioclim.html (accessed on 16 June 2022).











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Name | RO | HU | SR | BG |
|---|---|---|---|---|---|
| Amino acid | Glutamic acid | + | + | + | + |
| Isoleucine/Leucine | + | + | + | + | |
| Phenylalanine | + | + | + | + | |
| Tryptophan | + | + | + | + | |
| Other | 1-Benzofuranecarbaldehyde | + | + | + | + |
| Indole-4-carbaldehyde | + | + | + | + | |
| Flavonoid | Eriodictyol-O-hexoside | + | − | + | − |
| Homoeriodictyol (3′-Methoxy-4′,5,7-trihydroxyflavanone) | + | + | + | + | |
| Isoquercitrin (Hirsutrin, Quercetin-3-O-glucoside) | + | + | + | − | |
| Isorhamnetin-3-O-glucoside | + | + | + | + | |
| Isorhamnetin-3-O-rutinoside (Narcissin) | + | + | + | + | |
| Kaempferol-O-(rhamnosyl)hexoside | + | + | + | + | |
| Kaempferol-O-hexoside | + | − | + | − | |
| Naringenin | + | + | + | + | |
| Quercetin-3-O-rutinoside-7-O-glucoside | + | + | + | − | |
| Quercetin-O-(pentosyl)hexoside | + | − | − | − | |
| Quercetin-O-(rhamnosyl)hexoside-O-hexoside isomer | + | + | + | − | |
| Reinutrin (Reynoutrin, Quercetin-3-O-xyloside) | + | + | + | − | |
| Rutin | + | + | + | + | |
| Carboxylic acid | 12-Oxo phytodienoic acid | + | + | + | − |
| Citric acid | + | + | + | + | |
| Hexadecanedioic acid | + | + | + | − | |
| Quinic acid | + | + | + | + | |
| Coumarin | Dihydroxycoumarin-O-glucoside | + | + | + | + |
| Polyphenol | 12-Hydroxyjasmonic acid-12-O-glucoside like Tuberonic acid glucoside | + | + | + | + |
| 3-O-Feruloylquinic acid | + | + | + | + | |
| 4-O-Feruloylquinic acid | + | + | + | + | |
| 5-O-Feruloylquinic acid | + | + | + | + | |
| Chlorogenic acid (3-O-Caffeoylquinic acid) | + | + | + | + | |
| Chryptochlorogenic acid (4-O-Caffeoylquinic acid) | + | + | + | + | |
| cis-3-O-(4-Coumaroyl)quinic acid | + | + | + | + | |
| cis-4-O-(4-Coumaroyl)quinic acid | + | + | + | + | |
| cis-5-O-(4-Coumaroyl)quinic acid | + | + | + | + | |
| Di-O-caffeoylquinic acid isomer 1 | + | + | + | + | |
| Di-O-caffeoylquinic acid isomer 2 | + | + | + | + | |
| Feruloylquinic acid isomer | − | − | − | + | |
| Feruloylquinic acid isomer 1 | − | + | − | − | |
| Feruloylquinic acid isomer 2 | − | + | − | − | |
| Neochlorogenic acid (5-O-Caffeoylquinic acid) | + | + | + | + | |
| trans-3-O-(4-Coumaroyl)quinic acid | + | + | + | + | |
| trans-4-O-(4-Coumaroyl)quinic acid | + | + | + | + | |
| trans-5-O-(4-Coumaroyl)quinic acid | + | + | + | + | |
| trans-Melilotoside (trans-Glucosyl-2-hydroxycinnamate) | + | + | + | + | |
| Saponin | Mateglycoside B (Ilexpernoside H, Matenoside A) like Mateglycoside B′ | + | + | + | + |
| Mateglycoside C like Mateglycoside C’ | + | + | + | + | |
| Mateglycoside D (J3a) like J3b | + | + | + | + | |
| Matesaponin 1 like Matesaponin 1′ | + | + | + | + | |
| Matesaponin 2 like Matesaponin 2′ | + | + | + | + | |
| Matesaponin 3 (Araliasaponin X) like Matesaponin 3′ | − | + | + | + | |
| Matesaponin 4 like Mateglycoside A (Matenoside C) | + | + | + | + | |
| Terpenoid | Abscisic acid | + | + | + | + |
| Betulinic acid | + | + | + | + | |
| Oleanolic acid | + | + | + | + | |
| Ursolic acid | + | + | + | + | |
| Uvaol | + | + | + | − | |
| Vitamin | Adenine (B4) | + | + | + | + |
| Nicotinic acid (Niacin, B3) | + | + | + | + | |
| Pyridoxine (B6) | + | + | + | − | |
| Riboflavin (B2) | + | + | + | + |
| RO | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Vars | n | Mean | sd | Median | Trimmed | Mad | Min | Max | Range | Skew | |
| SD | 1 | 10 | 78.97 | 5.56 | 78.69 | 78.51 | 4.93 | 71.25 | 90.38 | 19.13 | 0.45 |
| DTS | 2 | 10 | 149.67 | 26.05 | 152.60 | 150.39 | 26.87 | 105.34 | 188.29 | 82.95 | −0.18 |
| SW | 3 | 10 | 15.95 | 1.03 | 16.14 | 16.07 | 0.57 | 13.67 | 17.28 | 3.60 | −0.81 |
| SL | 4 | 10 | 24.40 | 1.07 | 24.40 | 24.56 | 0.42 | 21.85 | 25.68 | 3.83 | −1.03 |
| STL | 5 | 10 | 73.22 | 7.11 | 73.76 | 73.22 | 4.92 | 59.87 | 86.60 | 26.73 | −0.03 |
| SF | 6 | 10 | 1073.40 | 85.57 | 1071.00 | 1073.00 | 103.78 | 935.00 | 1215.00 | 280.00 | 0.04 |
| STF | 7 | 10 | 797.80 | 221.96 | 795.50 | 774.12 | 223.13 | 520.00 | 1265.00 | 745.00 | 0.63 |
| HU | |||||||||||
| vars | n | Mean | sd | Median | Trimmed | Mad | Min | Max | Range | Skew | |
| SD | 1 | 10 | 69.21 | 4.47 | 70.05 | 69.60 | 4.20 | 60.98 | 74.32 | 13.35 | −0.48 |
| DTS | 2 | 10 | 133.80 | 24.02 | 130.56 | 129.18 | 13.79 | 109.47 | 195.08 | 85.61 | 1.46 |
| SW | 3 | 10 | 18.78 | 0.51 | 18.70 | 18.75 | 0.74 | 18.20 | 19.62 | 1.42 | 0.23 |
| SL | 4 | 10 | 27.26 | 0.76 | 27.11 | 27.14 | 0.62 | 26.46 | 29.00 | 2.54 | 1.02 |
| STL | 5 | 10 | 55.02 | 7.48 | 54.58 | 54.80 | 9.35 | 45.25 | 66.58 | 21.33 | 0.16 |
| SF | 6 | 10 | 1108.50 | 45.55 | 1119.00 | 1110.75 | 53.37 | 1043.00 | 1156.00 | 113.00 | −0.26 |
| STF | 7 | 10 | 270.40 | 77.81 | 273.00 | 267.50 | 43.74 | 129.00 | 435.00 | 306.00 | 0.31 |
| SR | |||||||||||
| vars | n | Mean | sd | Median | Trimmed | Mad | Min | Max | Range | Skew | |
| SD | 1 | 10 | 66.79 | 5.57 | 66.53 | 67.10 | 5.90 | 57.17 | 73.91 | 16.74 | −0.10 |
| DTS | 2 | 10 | 130.76 | 25.19 | 123.19 | 129.03 | 22.03 | 104.03 | 171.33 | 67.30 | 0.52 |
| SW | 3 | 10 | 20.65 | 0.68 | 20.70 | 20.60 | 0.55 | 19.73 | 21.98 | 2.26 | 0.42 |
| SL | 4 | 10 | 26.47 | 1.00 | 26.59 | 26.48 | 1.24 | 24.94 | 27.90 | 2.96 | −0.16 |
| STL | 5 | 10 | 56.13 | 3.60 | 55.85 | 55.95 | 4.30 | 51.24 | 62.42 | 11.18 | 0.30 |
| SF | 6 | 10 | 1110.20 | 87.13 | 1139.00 | 1109.38 | 64.49 | 975.00 | 1252.00 | 277.00 | −0.17 |
| STF | 7 | 10 | 407.40 | 84.66 | 390.00 | 399.00 | 71.91 | 297.00 | 585.00 | 288.00 | 0.70 |
| BG | |||||||||||
| vars | n | Mean | sd | Median | Trimmed | Mad | Min | Max | Range | Skew | |
| SD | 1 | 10 | 76.28 | 5.27 | 75.65 | 75.76 | 2.93 | 69.71 | 87.07 | 17.36 | 0.85 |
| DTS | 2 | 10 | 138.78 | 6.43 | 136.92 | 137.70 | 3.53 | 131.89 | 154.29 | 22.40 | 1.24 |
| SW | 3 | 10 | 17.27 | 1.24 | 17.14 | 17.24 | 1.48 | 15.56 | 19.23 | 3.67 | 0.06 |
| SL | 4 | 10 | 24.90 | 1.26 | 24.96 | 24.86 | 1.08 | 22.78 | 27.30 | 4.52 | 0.12 |
| STL | 5 | 10 | 57.51 | 5.97 | 57.55 | 57.47 | 5.31 | 47.88 | 67.50 | 19.62 | 0.00 |
| SF | 6 | 10 | 929.10 | 92.30 | 942.50 | 927.62 | 100.08 | 797.00 | 1073.00 | 276.99 | −0.05 |
| STF | 7 | 10 | 83.49 | 67.38 | 56.59 | 69.50 | 31.88 | 27.00 | 251.99 | 224.99 | 1.42 |
| whole data sets | |||||||||||
| vars | n | Mean | sd | Median | Trimmed | Mad | Min | Max | Range | Skew | |
| SD | 1 | 10 | 72.81 | 7.12 | 73.26 | 72.70 | 6.46 | 57.17 | 90.38 | 33.21 | 0.16 |
| DTS | 2 | 10 | 138.25 | 22.33 | 135.99 | 136.52 | 21.97 | 104.03 | 195.08 | 91.05 | 9.63 |
| SW | 3 | 10 | 18.16 | 1.98 | 18.20 | 18.18 | 2.32 | 13.67 | 21.98 | 8.31 | −0.10 |
| SL | 4 | 10 | 25.76 | 1.54 | 25.68 | 25.80 | 1.74 | 21.85 | 29.00 | 7.15 | −0.25 |
| STL | 5 | 10 | 60.47 | 9.60 | 59.26 | 59.84 | 8.65 | 45.25 | 86.60 | 41.35 | 0.67 |
| SF | 6 | 10 | 1055.30 | 107.49 | 1068.00 | 1064.41 | 116.38 | 797.00 | 1252.00 | 455.00 | −0.60 |
| STF | 7 | 10 | 389.75 | 293.19 | 317.50 | 357.44 | 283.18 | 27.00 | 1265.00 | 1238.00 | 0.94 |
| SD: K-W p-val = 0.0001319 | |||||||
| No. | MT | L1 | L.2 | Est.1 | Est.2 | p adj. | p adj. signf. |
| 1 | SD | RO | HU | 30.5 | 13.9 | 0.0038884472 | ** |
| 2 | SD | RO | SR | 30.5 | 10.7 | 0.0009141059 | *** |
| 3 | SD | RO | BG | 30.5 | 26.9 | 0.5404890339 | ns |
| 4 | SD | HU | SR | 13.9 | 10.7 | 0.5404890339 | ns |
| 5 | SD | HU | BG | 13.9 | 26.9 | 0.0193483109 | * |
| 6 | SD | SR | BG | 10.7 | 26.9 | 0.0038884472 | ** |
| DTS: K-W p-val = 0.1733 | |||||||
| No. | MT | Gr.1 | Gr.2 | Est.1 | Est.2 | p adj. | p adj. signf. |
| 1 | DTS | RO | HU | 25.9 | 17.4 | 0.3119635 | ns |
| 2 | DTS | RO | SR | 25.9 | 15.7 | 0.3063532 | ns |
| 3 | DTS | RO | BG | 25.9 | 23.0 | 0.6949266 | ns |
| 4 | DTS | HU | SR | 17.4 | 15.7 | 0.7450569 | ns |
| 5 | DTS | HU | BG | 17.4 | 23.0 | 0.4261672 | ns |
| 6 | DTS | SR | BG | 15.7 | 23.0 | 0.3252526 | ns |
| SW: K-W p-val = 4.212 × 10−7 | |||||||
| No. | MT | Gr.1 | Gr.2 | Est.1 | Est.2 | p adj. | p adj. signf. |
| 1 | SW | RO | HU | 7.7 | 24.5 | 2.618087 × 10−3 | ** |
| 2 | SW | RO | SR | 7.7 | 35.5 | 6.281448 × 10−7 | *** |
| 3 | SW | RO | BG | 7.7 | 14.3 | 2.067190 × 10−1 | ns |
| 4 | SW | HU | SR | 24.5 | 35.5 | 5.301561 × 10−2 | ns |
| 5 | SW | HU | BG | 24.5 | 14.3 | 6.121838 × 10−2 | ns |
| 6 | SW | SR | BG | 35.5 | 14.3 | 1.499218 × 10−4 | *** |
| SL: K-W p-val = 3.995 × 10−5 | |||||||
| No. | MT | Gr.1 | Gr.2 | Est.1 | Est.2 | p adj. | p adj. signf. |
| 1 | SL | RO | HU | 9.9 | 31.9 | 0.0001545802 | *** |
| 2 | SL | RO | SR | 9.9 | 26.1 | 0.0038884472 | ** |
| 3 | SL | RO | BG | 9.9 | 14.1 | 0.4217743968 | ns |
| 4 | SL | HU | SR | 31.9 | 26.1 | 0.3207177427 | ns |
| 5 | SL | HU | BG | 31.9 | 14.1 | 0.0019873969 | ** |
| 6 | SL | SR | BG | 26.1 | 14.1 | 0.0325759538 | * |
| STL: K-W p-val = 0.0001979 | |||||||
| No. | MT | Gr.1 | Gr.2 | Est.1 | Est.2 | p adj. | p adj. signf. |
| 1 | SL | RO | HU | 34.5 | 13.9 | 0.0004884332 | *** |
| 2 | SL | RO | SR | 34.5 | 15.8 | 0.0010434569 | ** |
| 3 | SL | RO | BG | 34.5 | 17.8 | 0.0028038024 | ** |
| 4 | SL | HU | SR | 13.9 | 15.8 | 0.7162921150 | ns |
| 5 | SL | HU | BG | 13.9 | 17.8 | 0.6835330598 | ns |
| 6 | SL | SR | BG | 15.8 | 17.8 | 0.7162921150 | ns |
| SF: K-W p-val = 0.0009187 | |||||||
| No. | MT | Gr.1 | Gr.2 | Est.1 | Est.2 | p adj. | p adj. signf. |
| 1 | SF | RO | HU | 22.00 | 26.45 | 0.574900837 | ns |
| 2 | SF | RO | SR | 22.00 | 25.70 | 0.574900837 | ns |
| 3 | SF | RO | BG | 22.00 | 7.85 | 0.013588382 | * |
| 4 | SF | HU | SR | 26.45 | 25.70 | 0.885920409 | ns |
| 5 | SF | HU | BG | 26.45 | 7.85 | 0.001916669 | ** |
| 6 | SF | SR | BG | 25.70 | 7.85 | 0.001916669 | ** |
| STF: K-W p-val = 1.367 × 10−7 | |||||||
| No. | MT | Gr.1 | Gr.2 | Est.1 | Est.2 | p adj. | p adj. signf. |
| 1 | SF | RO | HU | 35.4 | 16.1 | 6.005562 × 10−4 | *** |
| 2 | SF | RO | SR | 35.4 | 24.7 | 5.859064 × 10−2 | ns |
| 3 | SF | RO | BG | 35.4 | 5.8 | 8.993826 × 10−8 | *** |
| 4 | SF | HU | SR | 16.1 | 24.7 | 9.998055 × 10−2 | ns |
| 5 | SF | HU | BG | 16.1 | 5.8 | 5.859064 × 10−2 | ns |
| 6 | SF | SR | BG | 24.7 | 5.8 | 6.005562 × 10−4 | *** |
| Zimbru/RO | Szarvas/HU | Mala-Reka/SR | Borino/BG | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Min/Avg/Max temp. (°C)—October | 6 | 13 | 17 | 10 | 15 | 18 | 10 | 15 | 18 | 7 | 10 | 13 |
| Min/Avg/Max temp. (°C)—2017 | −11 | 14 | 32 | −9 | 14 | 32 | −7 | 14 | 32 | −11 | 9.6 | 24 |
| Avg. humidity(%)—October | 69 | 62 | 63 | 70 | ||||||||
| Min/Avg/Max humidity(%)—2017 | 52 | 74 | 90 | 43 | 67 | 83 | 50 | 68 | 86 | 56 | 75 | 91 |
| Avg. rainfall amount(mm)—October | 67.1 | 76.9 | 63 | 123.3 | ||||||||
| Min/Avg/Max rainfall amount(mm)—2017 | 23.2 | 66.6 | 160.1 | 42.3 | 67.5 | 114.6 | 19.9 | 71 | 132.7 | 70.3 | 155 | 333 |
| Min/Avg/Max rainy days/month—2017 | 1 | 6 | 12 | 2 | 4.9 | 7 | 1 | 5.8 | 13 | 4 | 10 | 24 |
| Altitude(m) | 258 | 79 | 1033 | 1143 | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mihali, C.V.; Petrescu, C.M.; Ciolacu-Ladasiu, C.F.; Mathe, E.; Popescu, C.; Bota, V.; Mizeranschi, A.E.; Ilie, D.E.; Neamț, R.I.; Turcus, V. Assessing Phenotypic Variability in Some Eastern European Insular Populations of the Climatic Relict Ilex aquifolium L. Plants 2022, 11, 2022. https://doi.org/10.3390/plants11152022
Mihali CV, Petrescu CM, Ciolacu-Ladasiu CF, Mathe E, Popescu C, Bota V, Mizeranschi AE, Ilie DE, Neamț RI, Turcus V. Assessing Phenotypic Variability in Some Eastern European Insular Populations of the Climatic Relict Ilex aquifolium L. Plants. 2022; 11(15):2022. https://doi.org/10.3390/plants11152022
Chicago/Turabian StyleMihali, Ciprian Valentin, Constantin Marian Petrescu, Calin Flavius Ciolacu-Ladasiu, Endre Mathe, Cristina Popescu, Viviane Bota, Alexandru Eugeniu Mizeranschi, Daniela Elena Ilie, Radu Ionel Neamț, and Violeta Turcus. 2022. "Assessing Phenotypic Variability in Some Eastern European Insular Populations of the Climatic Relict Ilex aquifolium L." Plants 11, no. 15: 2022. https://doi.org/10.3390/plants11152022
APA StyleMihali, C. V., Petrescu, C. M., Ciolacu-Ladasiu, C. F., Mathe, E., Popescu, C., Bota, V., Mizeranschi, A. E., Ilie, D. E., Neamț, R. I., & Turcus, V. (2022). Assessing Phenotypic Variability in Some Eastern European Insular Populations of the Climatic Relict Ilex aquifolium L. Plants, 11(15), 2022. https://doi.org/10.3390/plants11152022