
Tobacco Mosaic Virus Infection of Chrysanthemums in Thailand: Development of Colorimetric Reverse-Transcription Loop-Mediated Isothermal Amplification (RT–LAMP) Technique for Sensitive and Rapid Detection

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection, RNA Extraction, and cDNA Synthesis
2.2. RT–PCR Detection
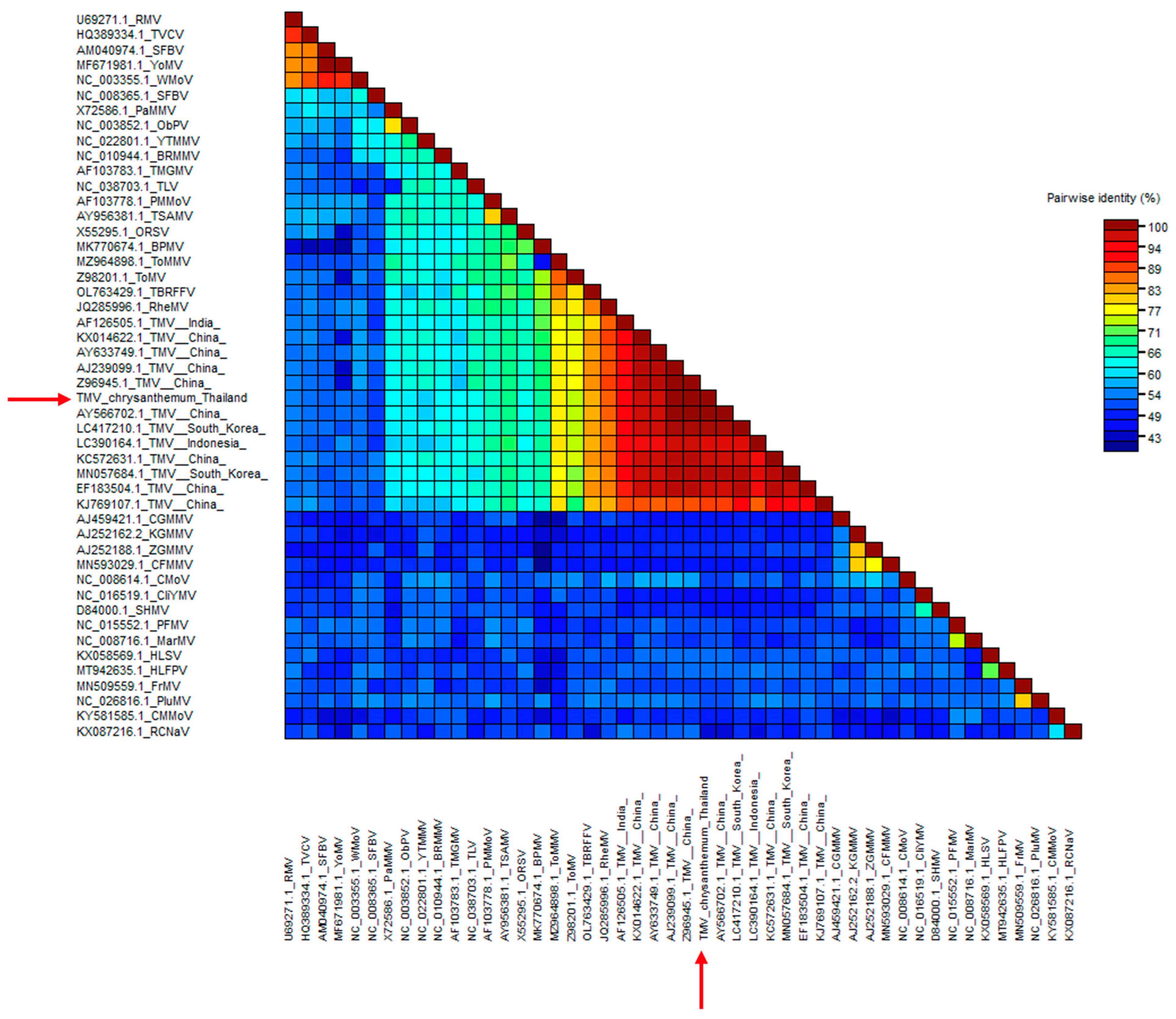
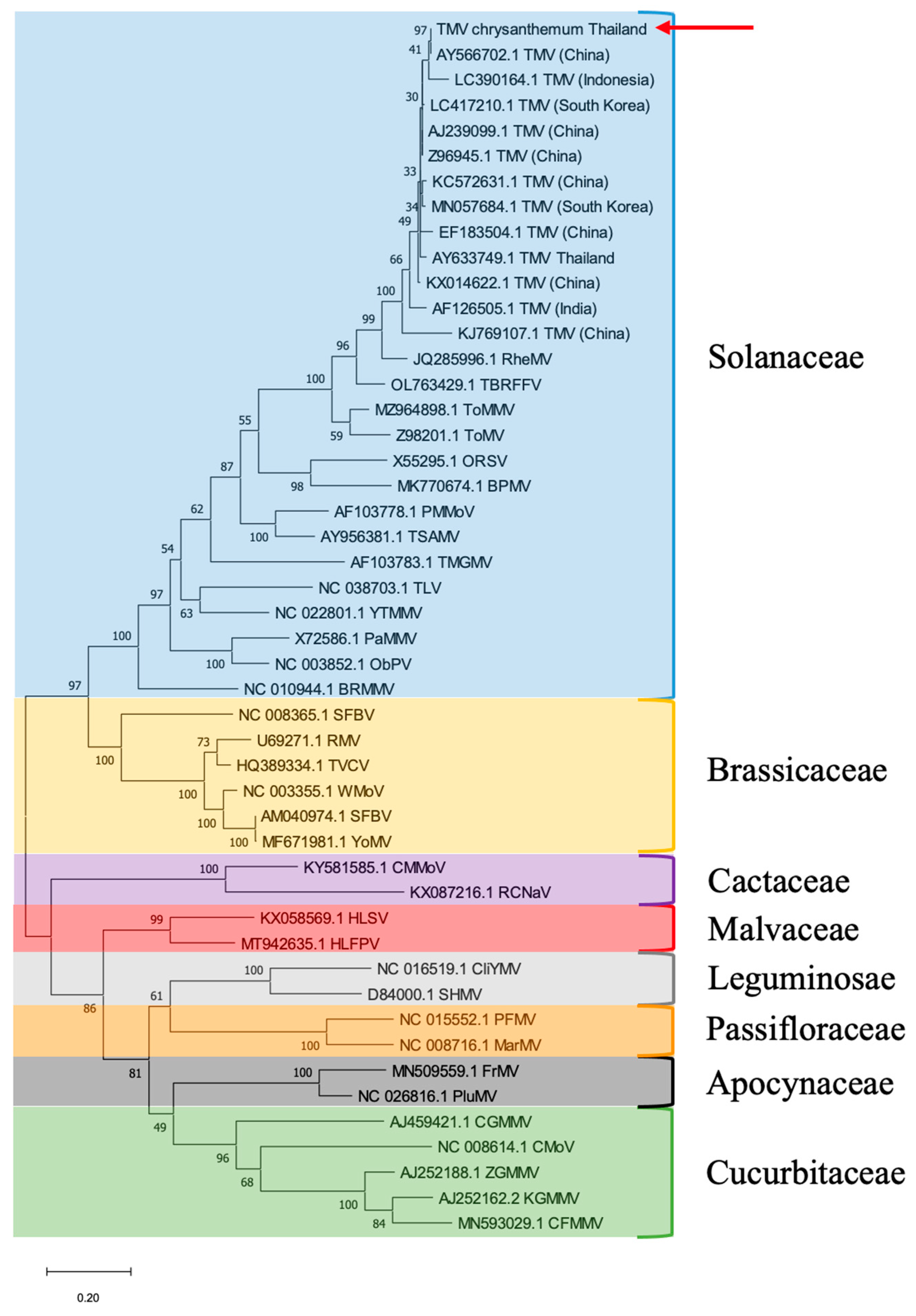
2.3. Sequence and Phylogenetic Analysis
2.4. Bioassay of Chrysanthemums and Indicator Plants
2.5. Transmission Electron Microscopy (TEM)
2.6. LAMP Primer Design
2.7. Optimization of RT–LAMP Conditions
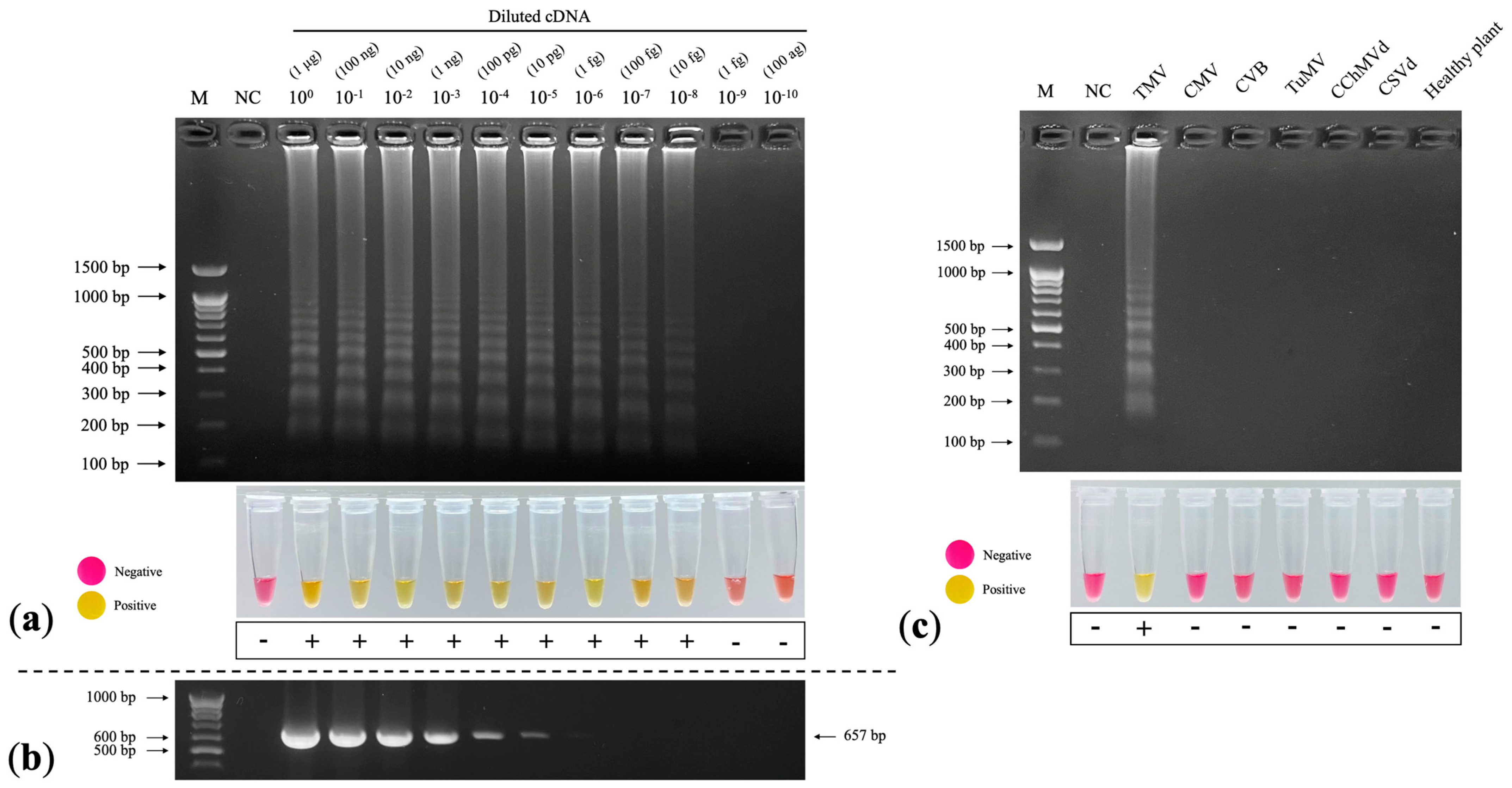
2.8. Sensitivity Assay
2.9. Specificity Assay
3. Results
3.1. RT–PCR Detection
3.2. Sequence and Phylogenetic Tree Analysis
3.3. Bioassay of Indicator Plants
3.4. Trasmission Electron Microscopy (TEM)
3.5. LAMP Primer Design and Optimization
3.6. Sensitivity Assay
3.7. Specificity Assay
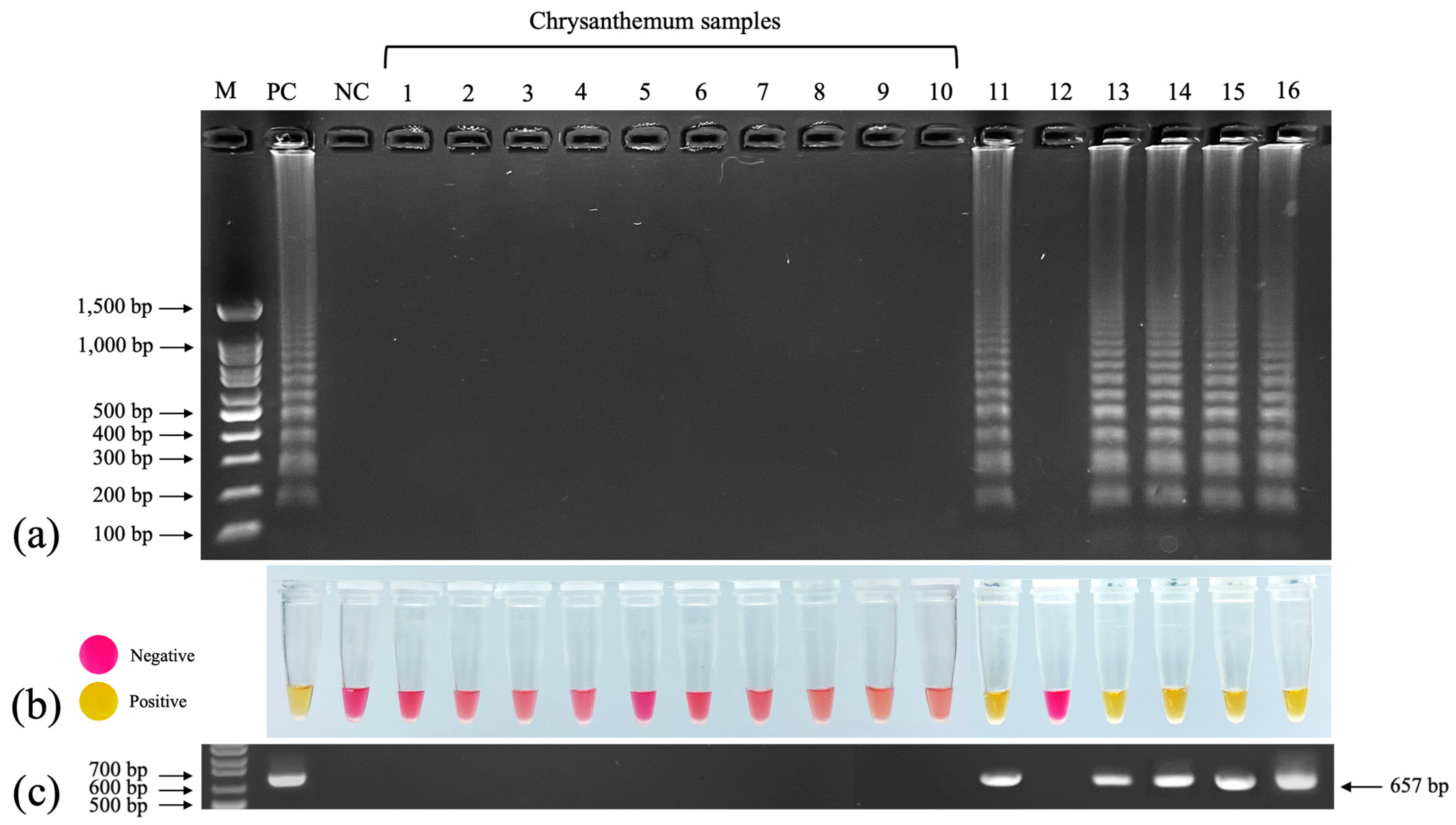
3.8. Evaluation of RT–LAMP for Detecting TMV
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zalewska, M.; Lema-Rumińska, J.; Miler, N. In vitro propagation using adventitious buds technique as a source of new variability in chrysanthemum. Sci. Hortic. 2007, 113, 70–73. [Google Scholar] [CrossRef]
- Trolinger, J.C.; McGovern, R.J.; Elmer, W.H.; Rechcigl, N.A.; Shoemaker, C.M. Diseases of chrysanthemum. In Handbook of Florists’ Crops Diseases; McGovern, R.J., Elmwe, W.H., Eds.; Springer: New York, NY, USA, 2017; pp. 1–66. [Google Scholar]
- Cho, W.K.; Jo, Y.; Jo, K.M.; Kim, K.H. A current overview of two viroids that infect chrysanthemums: Chrysanthemum stunt viroid and chrysanthemum chlorotic mottle viroid. Viruses 2013, 5, 1099–1113. [Google Scholar] [CrossRef] [PubMed]
- Verma, N.; Ram, R.; Hallan, V.A.; Kumar, K.; Zaidi, A.A. Production of cucumber mosaic virus-free chrysanthemums by meristem tip culture. J. Crop Prot. 2004, 23, 469–473. [Google Scholar] [CrossRef]
- Kondo, T.; Yamashita, K.; Sugiyama, S. First report of impatiens necrotic spot virus infecting chrysanthemum (Chrysanthemum morifolium) in Japan. J. Gen. Plant Pathol. 2011, 77, 263–265. [Google Scholar] [CrossRef]
- Song, A.; You, Y.; Chen, F.; Li, P.; Jiang, J.; Chen, S. A multiplex RT-PCR for rapid and simultaneous detection of viruses and viroids in chrysanthemum. Lett. Appl. Microbiol. 2012, 56, 8–13. [Google Scholar] [CrossRef]
- Niu, E.B.; Chen, L.J.; Niu, Y.B. First report of zucchini yellow mosaic virus in chrysanthemum. Plant Dis. 2015, 99, 1289. [Google Scholar] [CrossRef]
- Renukadevi, P.; Nagendran, K.; Nakkeeran, S.; Karthikeyan, G.; Jawaharlal, M.; Alice, D.; Malathi, V.G.; Pappu, H.R. First report of tomato spotted wilt virus infection of chrysanthemum in India. Plant Dis. 2015, 99, 1190. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, X.; Ge, B.; Li, M.; Hong, B. A multiplex RT-PCR for simultaneous detection and identification of five viruses and two viroids infecting chrysanthemum. Arch Virol. 2015, 160, 1145–1152. [Google Scholar] [CrossRef]
- Ashwathappa, K.V.; Venkataravanappa, V.; Reddy, C.N.; Reddy, M.K. Association of tomato leaf curl New Delhi virus with mosaic and leaf curl disease of chrysanthemum and its whitefly cryptic species. Indian Phytopathol. 2020, 73, 533–542. [Google Scholar] [CrossRef]
- Scholthof, K.B.G. Tobacco mosaic virus: A model system for plant biology. Annu. Rev. Phytopathol. 2004, 42, 13–34. [Google Scholar] [CrossRef]
- Abdelkhalek, A.; Sanan-Mishra, N. Comparative analysis of the suppressor activity of tobacco mosaic virus proteins in the tomato plant. Jordan J. Biol. Sci. 2018, 11, 469–473. [Google Scholar]
- Ng, J.C.; Perry, K.L. Transmission of plant viruses by aphid vectors. Mol. Plant Pathol. 2004, 5, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Zaitlin, M. Elucidation of the genome organization of tobacco mosaic virus. Philos. Trans. R. Soc. B 1999, 354, 587–591. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nassar, E.A.; El-Dougdoug, K.A.; Osman, M.E.; Dawoud, R.A.; Kinawy, A.H. Characterization and elimination of a TMV isolate infecting chrysanthemum plants in Egypt. Int. J. Virol. 2012, 8, 14–26. [Google Scholar] [CrossRef][Green Version]
- Liu, X.L.; Zhao, X.T.; Muhammad, I.; Ge, B.B.; Hong, B. Multiplex reverse transcription loop-mediated isothermal amplification for the simultaneous detection of CVB and CSVd in chrysanthemum. J. Virol. Methods 2014, 210, 26–31. [Google Scholar] [CrossRef]
- Park, M.; Baek, E.; Yoon, J.Y.; Palukaitis, P. The use of a tobacco mosaic virus-based expression vector system in chrysanthemum. Plant Pathol. J. 2017, 33, 429–433. [Google Scholar] [CrossRef]
- Jeong, J.J.; Ju, H.J.; Noh, J. A review of detection methods for the plant viruses. Res. Plant Dis. 2014, 20, 173–181. [Google Scholar] [CrossRef]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, e63. [Google Scholar] [CrossRef]
- Panno, S.; Matić, S.; Tiberini, A.; Caruso, A.G.; Bella, P.; Torta, L.; Stassi, R.; Davino, S. Loop mediated isothermal amplification: Principles and applications in plant virology. Plants 2020, 9, 461. [Google Scholar] [CrossRef]
- Poole, C.B.; Li, Z.; Alhassan, A.; Guelig, D.; Diesburg, S.; Tanner, N.A.; Zhang, Y.; Evans, T.C., Jr.; LaBarre, P.; Wanji, S.; et al. Colorimetric tests for diagnosis of filarial infection and vector surveillance using non-instrumented nucleic acid loop-mediated isothermal amplification (NINA-LAMP). PLoS ONE 2017, 12, e0169011. [Google Scholar] [CrossRef]
- Letschert, B.; Adam, G.; Lesemann, D.E.; Willingmann, P.; Heinze, C. Detection and differentiation of serologically cross-reacting tobamoviruses of economical importance by RT-PCR and RT-PCR-RFLP. J. Virol. Method 2002, 106, 1–10. [Google Scholar] [CrossRef]
- Ali, A.; Hussain, A.; Ahmad, M. Occurrence and molecular characterization of cucumber green mottle mosaic virus in cucurbit crops of KPK, Pakistan. Braz. J. Microbiol. 2013, 45, 1247–1253. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef] [PubMed]
- Supakitthanakorn, S.; Akarapisan, A.; Ruangwong, O.U. First record of melon yellow spot virus in pumpkin and its occurrence in cucurbitaceous crops in Thailand. Australas. Plant Dis. Notes 2018, 13, 32. [Google Scholar] [CrossRef]
- Lumata, J.L.; Ball, D.; Shahrivarkevishahi, A.; Luzuriaga, M.A.; Herbert, F.C.; Brohlin, O.; Lee, H.; Hagge, L.M.; D’Arcy, S.; Gassensmith, J.J. Identification and physical characterization of a spontaneous mutation of the tobacco mosaic virus in the laboratory environment. Sci. Rep. 2021, 11, e0169011. [Google Scholar] [CrossRef]
- Jung, H.W.; Yun, W.S.; Hahm, Y.I.; Kim, K.H. Characterization of tobacco mosaic virus isolated from potato showing yellow leaf mosaic and stunting symptoms in Korea. Plant Dis. 2002, 65, 112–117. [Google Scholar] [CrossRef]
- Asthana, S.; Roytberg, M.; Stamatoyannopoulos, J.; Sunyaev, S. Analysis of sequence conservation at nucleotide resolution. PLoS Comput. Biol. 2007, 3, e254. [Google Scholar] [CrossRef]
- Min, B.E.; Chung, B.N.; Kim, M.J.; Ha, J.H.; Lee, B.Y.; Ryu, K.H. Cactus mild mottle virus is a new cactus-infecting tobamovirus. Arch Virol. 2006, 151, 13–21. [Google Scholar] [CrossRef]
- Song, Y.S.; Min, B.E.; Hong, J.S.; Rhie, M.J.; Kim, M.J.; Ryu, K.H. Molecular evidence supporting the confirmation of Maracuja mosaic virus as a species of the genus Tobamovirus and production of an infectious cDNA transcript. Arch Virol. 2006, 151, 2337–2348. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.; Hu, J.; Xiao, L.; Tan, G.; Lan, P.; Liu, Y.; Li, F. The complete genome sequence, occurrence and host range of tomato mottle mosaic virus Chinese isolate. Virol. J. 2017, 14, 15. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Liu, Y.; Wu, Y.; Hao, X. Rapid detection of watermelon viruses by reverse transcription loop-mediated isothermal amplification. J. Phytopathol. 2016, 164, 330–336. [Google Scholar] [CrossRef]
- Ren, J.; Wang, J.; Zhu, Y.; Han, C.; Shang, Q. A reverse transcription loop-mediated isothermal amplification assay for the detection of strawberry mottle virus. J. Phytopathol. 2021, 169, 295–302. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, X.; Li, M.; Qiu, Y.; Li, G.; Li, Y.; Zhu, S. Development of reverse transcription loop-mediated isothermal amplification assay for sensitive and rapid detection of hosta virus X. J. Phytopathol. 2018, 166, 291–297. [Google Scholar] [CrossRef]
- Karwitha, M.; Feng, Z.K.; Shen, Y.; Essendi, W.; Zhang, W.N.; Li, J.Y.; Tao, X.R. Rapid detection of tomato chlorosis virus from infected plant and whitefly by one-step reverse transcription loop-mediated isothermal amplification. J. Phytopathol. 2015, 164, 255–263. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Z.; Qian, Y.; Mu, J.; Shen, L.; Wang, F.; Yang, J. Rapid detection of tobacco mosaic virus using the reverse transcription loop-mediated isothermal amplification method. Arch Virol. 2010, 155, 1681–1685. [Google Scholar] [CrossRef]
- Nagamine, K.; Hase, T.; Notomi, T. Accelerated reaction by loop-mediated isothermal amplification using loop primers. Mol. Cell Probes 2022, 16, 223–229. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chrysanthemum Cultivar | No. of Collected and Detected | No. of Positive to TMV | PDI * |
|---|---|---|---|
| Candor Pink | 10 | 0 | 0 |
| Celebrate | 12 | 0 | 0 |
| Feeling Green | 7 | 0 | 0 |
| Huay Leuk-1 | 10 | 0 | 0 |
| Huay Leuk-4 | 15 | 1 | 6.66% |
| Leopard | 9 | 0 | 0 |
| New Day | 7 | 0 | 0 |
| Center | 5 | 0 | 0 |
| Orange Day | 7 | 0 | 0 |
| One Way Improve | 5 | 0 | 0 |
| Vanora | 6 | 0 | 0 |
| Cornallia | 8 | 0 | 0 |
| Explorer | 9 | 0 | 0 |
| Total | 110 | 1 | 0.91% |
| Indicator Plant | Symptom | |
|---|---|---|
| Inoculated Leaf | Upper Leaf | |
| Capsicum annum | CS *, VC | M |
| Chenopodium amaranticolor | NS | - |
| Chenopodium quinoa | NS | - |
| Cassia siamea | - | - |
| Chrysanthemum x morifolium | - | Mo |
| Chrysanthemum coronarium | Y | Ma, Y |
| Datura stramonium | NS, N | - |
| Nicotiana benthamiana | - | M, Ma |
| Nicotiana glutinosa | NS, N | - |
| Nicotiana tabacum cv. Samsun NN | NS | - |
| Nicotiana tabacum cv. Xanthi | - | M, Ma |
| Petunia x hybrida | - | M, Ma |
| Solanum lycopersicum | NS | M, VN |
| Vigna unguiculata | NS | - |
| Primer | Sequence (5′–3′) | Attachment Position (5′–3′) |
|---|---|---|
| F3 * | CCGGAAAAAAGAGTGATGTC | 45–64 |
| B3 | ACAAGAACACGAACTGAGAT | 237–256 |
| FIP | ACTCATTCCTCCAAAATCCTTAACA- AAAGGGAAAAATAGTAGTAGTGATC | F2: 68–92 F1c: 121–145 |
| BIP | AATCGATGATGATTCGGAGGCT- GGAGTAGTGATACTGTAAGACAT | B2: 214–236 B1c: 163–184 |
| LF | TCTTGTTCGGCACTGACC | 93–110 |
| LB | GTCGCCGAATCGGATTCGTTTTA | 188–210 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Supakitthanakorn, S.; Vichittragoontavorn, K.; Sunpapao, A.; Kunasakdakul, K.; Thapanapongworakul, P.; Ruangwong, O.-U. Tobacco Mosaic Virus Infection of Chrysanthemums in Thailand: Development of Colorimetric Reverse-Transcription Loop-Mediated Isothermal Amplification (RT–LAMP) Technique for Sensitive and Rapid Detection. Plants 2022, 11, 1788. https://doi.org/10.3390/plants11141788
Supakitthanakorn S, Vichittragoontavorn K, Sunpapao A, Kunasakdakul K, Thapanapongworakul P, Ruangwong O-U. Tobacco Mosaic Virus Infection of Chrysanthemums in Thailand: Development of Colorimetric Reverse-Transcription Loop-Mediated Isothermal Amplification (RT–LAMP) Technique for Sensitive and Rapid Detection. Plants. 2022; 11(14):1788. https://doi.org/10.3390/plants11141788
Chicago/Turabian StyleSupakitthanakorn, Salit, Kanjana Vichittragoontavorn, Anurag Sunpapao, Kaewalin Kunasakdakul, Pilunthana Thapanapongworakul, and On-Uma Ruangwong. 2022. "Tobacco Mosaic Virus Infection of Chrysanthemums in Thailand: Development of Colorimetric Reverse-Transcription Loop-Mediated Isothermal Amplification (RT–LAMP) Technique for Sensitive and Rapid Detection" Plants 11, no. 14: 1788. https://doi.org/10.3390/plants11141788
APA StyleSupakitthanakorn, S., Vichittragoontavorn, K., Sunpapao, A., Kunasakdakul, K., Thapanapongworakul, P., & Ruangwong, O.-U. (2022). Tobacco Mosaic Virus Infection of Chrysanthemums in Thailand: Development of Colorimetric Reverse-Transcription Loop-Mediated Isothermal Amplification (RT–LAMP) Technique for Sensitive and Rapid Detection. Plants, 11(14), 1788. https://doi.org/10.3390/plants11141788