
Current State of Knowledge Regarding WHO Critical Priority Pathogens: Mechanisms of Resistance and Proposed Solutions through Candidates Such as Essential Oils


 , , ,
, , ,  ,
,  , and
, and 
Abstract
1. Introduction
2. WHO Critical Priority Pathogens
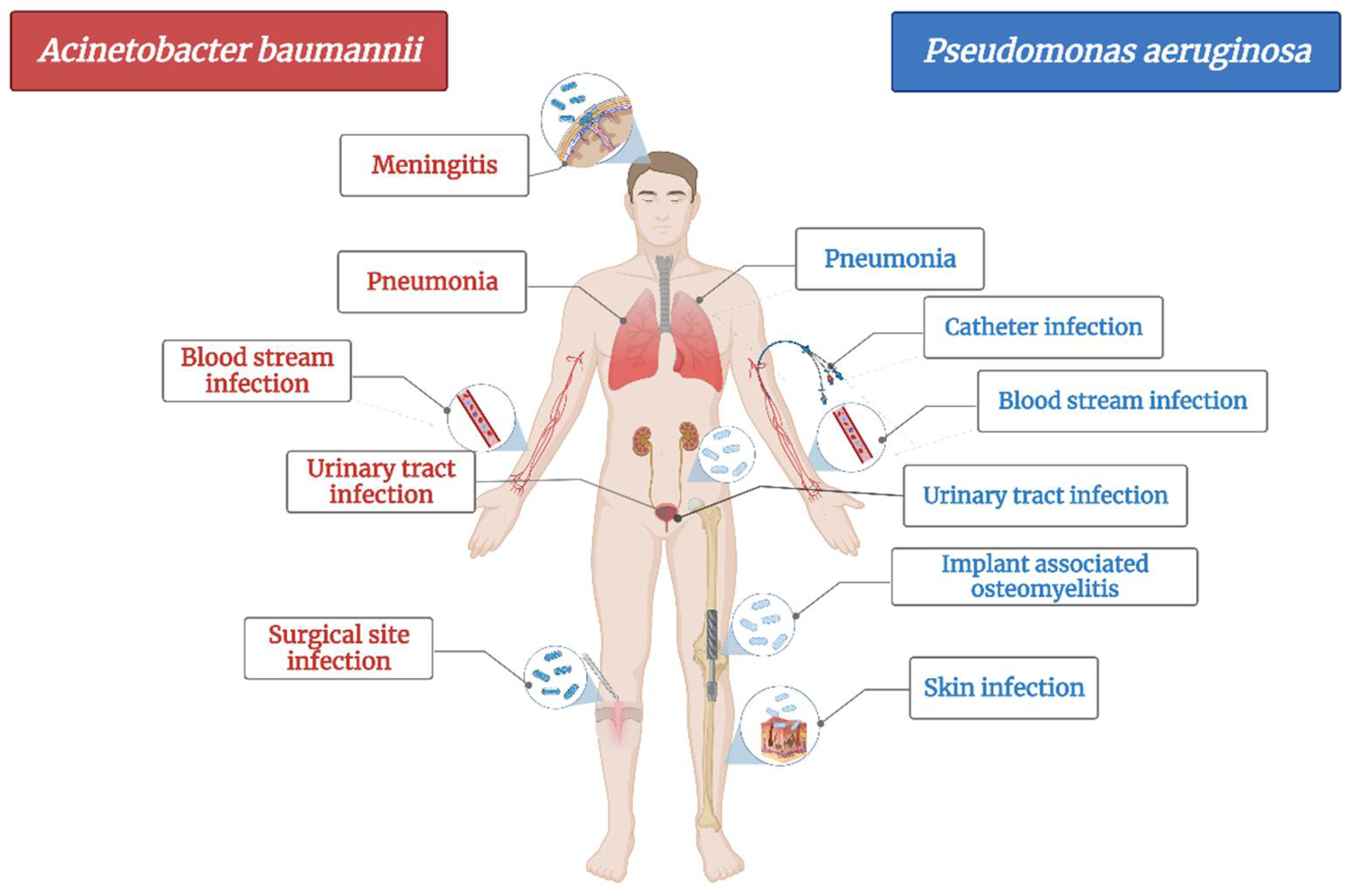
2.1. Carbapenem-Resistant Acinetobacter baumannii
2.2. Carbapenem-Resistant Pseudomonas aeruginosa
2.3. Carbapenem-Resistant and Extended Spectrum Beta-Lactamase (ESBL)-Producing Enterobacterales
2.3.1. Carbapenem-Resistant and ESBL-Producing Klebsiella pneumoniae
2.3.2. Carbapenem-Resistant and ESBL-Producing Escherichia coli
3. Materials and Methods
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| ABs | Antibiotics |
| AMR | Antimicrobial resistance |
| CNS | Central nervous system |
| EOs | Essential oils |
| ESBL | Extended spectrum beta-lactamase. |
| FICI | Fractional inhibitory concentration index |
| KPC | K. Pneumoniae carbapenemase |
| LPS | Lipopolysaccharides |
| MBEC50% | Minimum biofilm inhibitory concentration |
| MDR | Multidrug resistant |
| MIC | Minimum inhibitory concentration |
| MRSA | Methicillin-resistant Staphylococcus aureus |
| OmpA | Outer membrane protein |
| PDR | Pandrug-resistant |
| TTO | Tea tree oil |
| WHO | World Health Organization |
| XDR | Extensively drug-resistant |
References
- Aslam, B.; Wang, W.; Arshad, M.I.; Khurshid, M.; Muzammil, S.; Rasool, M.H.; Nisar, M.A.; Alvi, R.F.; Aslam, M.A.; Qamar, M.U.; et al. Antibiotic Resistance: A Rundown of a Global Crisis. Infect. Drug Resist. 2018, 11, 1645–1658. [Google Scholar] [CrossRef] [PubMed]
- Dadgostar, P. Antimicrobial Resistance: Implications and Costs. Infect. Drug Resist. 2019, 12, 3903–3910. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, G.; Midiri, A.; Gerace, E.; Biondo, C. Bacterial Antibiotic Resistance: The Most Critical Pathogens. Pathogens 2021, 10, 1310. [Google Scholar] [CrossRef] [PubMed]
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 39, 629–655. [Google Scholar] [CrossRef]
- World Health Organization. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. Cad. Pesqui. 2017, 43, 348–365. [Google Scholar]
- Surveillance Atlas of Infectious Diseases. Available online: https://www.ecdc.europa.eu/en/surveillance-atlas-infectious-diseases (accessed on 25 June 2022).
- Mayor, S. Antibiotic resistance is highest in south and east Europe. BMJ 2005, 330, 383. [Google Scholar] [CrossRef][Green Version]
- European Centre for Disease Prevention and Control (ECDC). Summary of the Latest Data on Antibiotic Resistance in the European Union; ECDC: Stockholm, Sweden, 2012; pp. 1–4. [Google Scholar]
- Elmahi, O.K.O.; Uakkas, S.; Olalekan, B.Y.; Damilola, I.A.; Adedeji, O.J.; Hasan, M.M.; dos Santos Costa, A.C.; Ahmad, S.; Essar, M.Y.; Thomson, D.J. Antimicrobial Resistance and One Health in the Post COVID-19 Era: What Should Health Students Learn? Antimicrob. Resist. Infect. Control 2022, 11, 58. [Google Scholar] [CrossRef]
- Chokshi, A.; Sifri, Z.; Cennimo, D.; Horng, H. Global Contributors to Antibiotic Resistance. J. Glob. Infect. Dis. 2019, 11, 36–42. [Google Scholar] [CrossRef]
- Martin, M.J.; Thottathil, S.E.; Newman, T.B. Antibiotics Overuse in Animal Agriculture: A Call to Action for Health Care Providers. Am. J. Public Health 2015, 105, 2409–2410. [Google Scholar] [CrossRef]
- McEwen, S.A.; Collignon, P.J. Antimicrobial Resistance: A One Health Colloquium. Microbiol. Spectr. 2018, 6, 1–26. [Google Scholar] [CrossRef]
- Zalewska, M.; Błażejewska, A.; Czapko, A.; Popowska, M. Antibiotics and Antibiotic Resistance Genes in Animal Manure–Consequences of Its Application in Agriculture. Front. Microbiol. 2021, 12, 610656. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Lee, S.; Jeong, K.C. Mitigating Antibiotic Resistance at the Livestock-Environment Interface:A Review. J. Microbiol. Biotechnol. 2019, 29, 1683–1692. [Google Scholar] [CrossRef] [PubMed]
- Aslam, B.; Khurshid, M.; Arshad, M.I.; Muzammil, S.; Rasool, M.; Yasmeen, N.; Shah, T.; Chaudhry, T.H.; Rasool, M.H.; Shahid, A.; et al. Antibiotic Resistance: One Health One World Outlook. Front. Cell. Infect. Microbiol. 2021, 11, 771510. [Google Scholar] [CrossRef]
- World Health Organization. Global Action Plan on Antimicrobial Resistance. 2015. Available online: https://apps.who.int/iris/handle/10665/193736 (accessed on 1 March 2022).
- Butler, M.S.; Gigante, V.; Sati, H.; Paulin, S.; Al-Sulaiman, L.; Rex, J.H.; Fernandes, P.; Arias, C.A.; Paul, M.; Thwaites, G.E.; et al. Analysis of the Clinical Pipeline of Treatments for Drug-Resistant Bacterial Infections: Despite Progress, More Action Is Needed. Antimicrob. Agents Chemother. 2022, 66, e01991-21. [Google Scholar] [CrossRef]
- Terreni, M.; Taccani, M.; Pregnolato, M. New Antibiotics for Multidrug-Resistant Bacterial Strains: Latest Research Developments and Future Perspectives. Molecules 2021, 26, 2671. [Google Scholar] [CrossRef] [PubMed]
- Murugan, M.S.; Sinha, D.K.; Vinodh Kumar, O.R.; Yadav, A.K.; Pruthvishree, B.S.; Vadhana, P.; Nirupama, K.R.; Bhardwaj, M.; Singh, B.R. Epidemiology of Carbapenem-Resistant Escherichia coli and First Report of BlaVIM Carbapenemases Gene in Calves from India. Epidemiol. Infect. 2019, 147, e159. [Google Scholar] [CrossRef]
- Ramsey, J.T.; Shropshire, B.C.; Nagy, T.R.; Chambers, K.D.; Li, Y.; Korach, K.S. Essential Oils and Health. Yale J. Biol. Med. 2020, 93, 291–305. [Google Scholar]
- Plant, R.M.; Dinh, L.; Argo, S.; Shah, M. The Essentials of Essential Oils. Adv. Pediatr. 2019, 66, 111–122. [Google Scholar] [CrossRef]
- Wińska, K.; Mączka, W.; Łyczko, J.; Grabarczyk, M.; Czubaszek, A.; Szumny, A. Essential Oils as Antimicrobial Agents—Myth or Real Alternative? Molecules 2019, 24, 2130. [Google Scholar] [CrossRef]
- Yu, Z.; Tang, J.; Khare, T.; Kumar, V. The Alarming Antimicrobial Resistance in ESKAPEE Pathogens: Can Essential Oils Come to the Rescue? Fitoterapia 2020, 140, 104433. [Google Scholar] [CrossRef]
- Valerio, F.; Mezzapesa, G.N.; Ghannouchi, A.; Mondelli, D.; Logrieco, A.F.; Perrino, E.V. Characterization and Antimicrobial Properties of Essential Oils from Four Wild Taxa of Lamiaceae Family Growing in Apulia. Agronomy 2021, 11, 1431. [Google Scholar] [CrossRef]
- Khoury, M.; Stien, D.; Eparvier, V.; Ouaini, N.; El Beyrouthy, M. Report on the Medicinal Use of Eleven Lamiaceae Species in Lebanon and Rationalization of Their Antimicrobial Potential by Examination of the Chemical Composition and Antimicrobial Activity of Their Essential Oils. Evid. Based Complement. Alternat. Med. 2016, 2016, 2547169. [Google Scholar] [CrossRef] [PubMed]
- Perrino, E.V.; Valerio, F.; Jallali, S.; Trani, A.; Mezzapesa, G.N. Ecological and Biological Properties of Satureja cuneifolia Ten. and Thymus spinulosus Ten.: Two Wild Officinal Species of Conservation Concern in Apulia (Italy). A Preliminary Survey. Plants 2021, 10, 1952. [Google Scholar] [CrossRef]
- Zhang, N.; Yao, L. Anxiolytic Effect of Essential Oils and Their Constituents: A Review. J. Agric. Food Chem. 2019, 67, 13790–13808. [Google Scholar] [CrossRef] [PubMed]
- Posadzki, P.; Alotaibi, A.; Ernst, E. Adverse Effects of Aromatherapy: A Systematic Review of Case Reports and Case Series. Int. J. Risk Saf. Med. 2012, 24, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Vostinaru, O.; Heghes, S.C.; Filip, L. Safety Profile of Essential Oils. In Essential Oils-Bioactive Compounds, New Perspectives and Applications; IntechOpen: London, UK, 2020; pp. 1–13. [Google Scholar]
- Wojtunik-Kulesza, K.A. Toxicity of Selected Monoterpenes and Essential Oils Rich in These Compounds. Molecules 2022, 27, 1716. [Google Scholar] [CrossRef]
- Lanzerstorfer, P.; Sandner, G.; Pitsch, J.; Mascher, B.; Aumiller, T.; Weghuber, J. Acute, Reproductive, and Developmental Toxicity of Essential Oils Assessed with Alternative In Vitro and In Vivo Systems. Arch. Toxicol. 2021, 95, 673–691. [Google Scholar] [CrossRef]
- Dosoky, N.S.; Setzer, W.N. Maternal Reproductive Toxicity of Some Essential Oils and Their Constituents. Int. J. Mol. Sci. 2021, 22, 2380. [Google Scholar] [CrossRef]
- Saishree Anchana, R.; Smiline Girija, A.S.; Gunasekaran, S.; Vijayashree Priyadharsini, J. Detection of CsgA Gene in Carbapenem-Resistant Acinetobacter baumannii Strains and Targeting with Ocimum sanctum Biocompounds. Iran. J. Basic Med. Sci. 2021, 24, 690–698. [Google Scholar] [CrossRef]
- Amaral, S.C.; Pruski, B.B.; de Freitas, S.B.; Allend, S.O.; Ferreira, M.R.A.; Moreira, C.; Pereira, D.I.B.; Junior, A.S.V.; Hartwig, D.D. Origanum Vulgare Essential Oil: Antibacterial Activities and Synergistic Effect with Polymyxin B against Multidrug-Resistant Acinetobacter baumannii. Mol. Biol. Rep. 2020, 47, 9615–9625. [Google Scholar] [CrossRef]
- Vasconcelos, N.G.; Croda, J.; Silva, K.E.; Motta, M.L.L.; Maciel, W.G.; Limiere, L.C.; Simionatto, S. Origanum Vulgare L. Essential Oil Inhibits the Growth of Carbapenem-Resistant Gram-Negative Bacteria. Rev. Soc. Bras. Med. Trop. 2019, 52, e20180502. [Google Scholar] [CrossRef] [PubMed]
- Bekka-Hadji, F.; Bombarda, I.; Djoudi, F.; Bakour, S.; Touati, A. Chemical Composition and Synergistic Potential of Mentha pulegium L. and Artemisia herba alba Asso. Essential Oils and Antibiotic against Multi-Drug Resistant Bacteria. Molecules 2022, 27, 1095. [Google Scholar] [CrossRef] [PubMed]
- Oliva, A.; Garzoli, S.; Sabatino, M.; Tadić, V.; Costantini, S.; Ragno, R.; Božović, M. Chemical Composition and Antimicrobial Activity of Essential Oil of Helichrysum italicum (Roth) G. Don Fil. (Asteraceae) from Montenegro. Nat. Prod. Res. 2020, 34, 445–448. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, F.; Oliva, A.; Sabatino, M.; Imbriano, A.; Hanieh, P.N.; Garzoli, S.; Mastroianni, C.M.; De Angelis, M.; Miele, M.C.; Arnaut, M.; et al. Antimicrobial Essential Oil Formulation: Chitosan Coated Nanoemulsions for Nose to Brain Delivery. Pharmaceutics 2020, 12, 678. [Google Scholar] [CrossRef]
- Mahmoudi, H.; Shokoohizadeh, L.; Zare Fahim, N.; Mohamadi Bardebari, A.; Moradkhani, S.; Alikhani, M.Y. Detection of AdeABC Efllux Pump Encoding Genes and Antimicrobial Effect of Mentha Longifolia and Menthol on MICs of Imipenem and Ciprofloxacin in Clinical Isolates of Acinetobacter baumannii. BMC Complement. Med. Ther. 2020, 20, 92. [Google Scholar] [CrossRef]
- Kaskatepe, B.; Kiymaci, M.E.; Suzuk, S.; Erdem, S.A.; Cesur, S.; Yildiz, S. Antibacterial Effects of Cinnamon Oil against Carbapenem Resistant Nosocomial Acinetobacter baumannii and Pseudomonas aeruginosa Isolates. Ind. Crops Prod. 2016, 81, 191–194. [Google Scholar] [CrossRef]
- Laktib, A.; Nayme, K.; El Hamdaoui, A.; Timinouni, M.; Hassi, M.; Aitalla, A.; Msanda, F.; Bourouache, M.; El Yaagoubi, M.; Mimoun, R.; et al. Antibacterial Activity of Lavandula Mairei Humbert Essential Oil against Carbapenem-Resistant Acinetobacter baumannii. Mediterr. J. Infect. Microbes Antimicrob. 2022, 11, 1–12. [Google Scholar] [CrossRef]
- Muntean, D.; Licker, M.; Alexa, E.; Popescu, I.; Jianu, C.; Buda, V.; Dehelean, C.A.; Ghiulai, R.; Horhat, F.; Horhat, D.; et al. Evaluation of Essential Oil Obtained from Mentha × piperita L. against Multidrug-Resistant Strains. Infect. Drug Resist. 2019, 12, 2905–2914. [Google Scholar] [CrossRef]
- Patterson, J.E.; McElmeel, L.; Wiederhold, N.P. In Vitro Activity of Essential Oils against Gram-Positive and Gram-Negative Clinical Isolates, Including Carbapenem-Resistant Enterobacteriaceae. Open Forum Infect. Dis. 2019, 6, ofz502. [Google Scholar] [CrossRef]
- Oliva, A.; Costantini, S.; De Angelis, M.; Garzoli, S.; Božović, M.; Mascellino, M.T.; Vullo, V.; Ragno, R. High Potency of Melaleuca Alternifolia Essential Oil against Multi-Drug Resistant Gram-Negative Bacteria and Methicillin-Resistant Staphylococcus aureus. Molecules 2018, 23, 2584. [Google Scholar] [CrossRef]
- Qian, W.; Liu, M.; Fu, Y.; Wang, T.; Zhang, J.; Yang, M.; Sun, Z.; Li, X.; Li, Y. Antimicrobial and Antibiofilm Activities of Citral against Carbapenem-Resistant Enterobacter Cloacae. Foodborne Pathog. Dis. 2020, 17, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Ginting, E.V.; Retnaningrum, E.; Widiasih, D.A. Antibacterial Activity of Clove (Syzygium aromaticum) and Cinnamon (Cinnamomum burmannii) Essential Oil against Extended-Spectrum β-Lactamase-Producing Bacteria. Vet. World 2021, 14, 2206–2211. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, J.; Mnayer, D.; Roointan, A.; Shahri, F.; Ayatollahi, S.A.M.; Sharifi-Rad, M.; Molaee, N. Antibacterial Activities of Essential Oils from Iranian Medicinal Plants on Extended-Spectrum Î2-Lactamase-Producing Escherichia coli. Cell. Mol. Biol. 2016, 62, 75–82. [Google Scholar] [PubMed]
- Kwiatkowski, P.; Pruss, A.; Grygorcewicz, B.; Wojciuk, B.; Dołȩgowska, B.; Giedrys-Kalemba, S.; Kochan, E.; Sienkiewicz, M. Preliminary Study on the Antibacterial Activity of Essential Oils Alone and in Combination with Gentamicin against Extended-Spectrum β-Lactamase-Producing and New Delhi Metallo-β-Lactamase-1-Producing Klebsiella pneumoniae Isolates. Microb. Drug Resist. 2018, 24, 1368–1375. [Google Scholar] [CrossRef] [PubMed]
- Benameur, Q.; Gervasi, T.; Pellizzeri, V.; Pľuchtová, M.; Tali-Maama, H.; Assaous, F.; Guettou, B.; Rahal, K.; Gruľová, D.; Dugo, G.; et al. Antibacterial Activity of Thymus Vulgaris Essential Oil Alone and in Combination with Cefotaxime against BlaESBL Producing Multidrug Resistant Enterobacteriaceae Isolates. Nat. Prod. Res. 2019, 33, 2647–2654. [Google Scholar] [CrossRef]
- Aouadhi, C.; Jouini, A.; Mechichi, D.; Boulares, M.; Hamrouni, S. Characterization of Primary Action Mode of Eight Essential Oils and Evaluation of Their Antibacterial Effect Against Extended-Spectrum β-Lactamase (ESBL)-Producing Escherichia coli Inoculated in Turkey Meat. Molecules 2022, 27, 2588. [Google Scholar] [CrossRef]
- Saliu, E.-M.; Ren, H.; Goodarzi Boroojeni, F.; Zentek, J.; Vahjen, W. The Impact of Direct-Fed Microbials and Phytogenic Feed Additives on Prevalence and Transfer of Extended-Spectrum Beta-Lactamase Genes in Broiler Chicken. Microorganisms 2020, 8, 322. [Google Scholar] [CrossRef]
- Motola, G.; Hafez, H.M.; Brüggemann-Schwarze, S. Efficacy of Six Disinfection Methods against Extended-Spectrum Beta-Lactamase (ESBL) Producing E. coli on Eggshells in Vitro. PLoS ONE 2020, 15, e0238860. [Google Scholar] [CrossRef]
- Krishnamoorthy, R.; Athinarayanan, J.; Periasamy, V.S.; Adisa, A.R.; Al-Shuniaber, M.A.; Gassem, M.A.; Alshatwi, A.A. Antimicrobial Activity of Nanoemulsion on Drug-Resistant Bacterial Pathogens. Microb. Pathog. 2018, 120, 85–96. [Google Scholar] [CrossRef]
- Tebrün, W.; Motola, G.; Hafez, M.H.; Bachmeier, J.; Schmidt, V.; Renfert, K.; Reichelt, C.; Brüggemann-Schwarze, S.; Pees, M. Preliminary Study: Health and Performance Assessment in Broiler Chicks Following Application of Six Different Hatching Egg Disinfection Protocols. PLoS ONE 2020, 15, e0232825. [Google Scholar] [CrossRef]
- Shrivastav, A.; Sharma, R.K.; Shrivastava, N.; Gautam, V.; Jain, S.K. Study of Inhibitory Potential and Percent Inhibition of Oil of Syzygium aromaticum and Leaves of Ocimum sanctum on ESBL Enzyme from Escherichia Coli in Broilers of Jabalpur. Indian J. Pharmacol. 2019, 51, 337. [Google Scholar] [CrossRef] [PubMed]
- Kaskatepe, B.; Yildiz, S.S.; Kiymaci, M.E.; Yazgan, A.N.; Cesur, S.; Erdem, S.A. Chemical Composition and Antimicrobial Activity of the Commercial Origanum onites L. Oil against Nosocomial Carbapenem Resistant Extended Spectrum Beta Lactamase Producer Escherichia coli Isolates. Acta Biol. Hung. 2017, 68, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Tadić, V.; Oliva, A.; Božović, M.; Cipolla, A.; De Angelis, M.; Vullo, V.; Garzoli, S.; Ragno, R. Chemical and Antimicrobial Analyses of Sideritis romana L. Subsp. Purpurea (Tal. Ex Benth.) Heywood, an Endemic of the Western Balkan. Molecules 2017, 22, 1395. [Google Scholar] [CrossRef] [PubMed]
- Iseppi, R.; Di Cerbo, A.; Aloisi, P.; Manelli, M.; Pellesi, V.; Provenzano, C.; Camellini, S.; Messi, P.; Sabia, C. In Vitro Activity of Essential Oils against Planktonic and Biofilm Cells of Extended-Spectrum β-Lactamase (ESBL)/Carbapenamase-Producing Gram-Negative Bacteria Involved in Human Nosocomial Infections. Antibiotics 2020, 9, 272. [Google Scholar] [CrossRef]
- Gāliņa, D.; Radenkovs, V.; Kviesis, J.; Valdovska, A. Effect of Essential Oils Supplemented with Caprylic Acid and Sodium Chloride against Faecal ESBL-Producing Escherichia coli Isolated from Pigs. Antibiotics 2022, 11, 461. [Google Scholar] [CrossRef]
- Benameur, Q.; Gervasi, T.; Pellizzeri, V.; Pľuchtová, M.; Gruľová, D.; Cicero, N.; Meriem-Hind, B. Comparison of Sensitivity to a Commercial Origanum Vulgare Essential Oil between Extended-Spectrum β-Lactamases (ESBL-) and Non-ESBL-Producing Enterobacteriaceae Isolates. Nat. Prod. Res. 2021, 36, 2830–2835. [Google Scholar] [CrossRef]
- Contreras-Moreno, B.Z.; Velasco, J.J.; Rojas, J.D.C.; Méndez, L.D.C.; Celis, M.T. Antimicrobial Activity of Essential Oil of Pimenta racemosa Var. Racemosa (Myrtaceae) Leaves. J. Pharm. Pharmacogn. Res. 2016, 4, 224–230. [Google Scholar]
- de Souza, G.H.A.; dos Santos Radai, J.A.; Vaz, M.S.M.; da Silva, K.E.; Fraga, T.L.; Barbosa, L.S.; Simionatto, S. In Vitro and In Vivo Antibacterial Activity Assays of Carvacrol: A Candidate for Development of Innovative Treatments against KPC-Producing Klebsiella pneumoniae. PLoS ONE 2021, 16, e0246003. [Google Scholar] [CrossRef]
- Dhara, L.; Tripathi, A. Sub-Acute Toxicological and Behavioural Effects of Two Candidate Therapeutics, Cinnamaldehyde and Eugenol, for Treatment of ESBL Producing-Quinolone Resistant Pathogenic Enterobacteriaceae. Clin. Exp. Pharmacol. Physiol. 2020, 47, 977–988. [Google Scholar] [CrossRef]
- Dhara, L.; Tripathi, A. Cinnamaldehyde: A Compound with Antimicrobial and Synergistic Activity against ESBL-Producing Quinolone-Resistant Pathogenic Enterobacteriaceae. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 65–73. [Google Scholar] [CrossRef]
- Gore, M.R.; Raut, D.; Aruna, K. Antimicrobial activity of Pimenta dioica (L.) merr. leaves and its synergistic activity with ampicillin against esbl producing clinical isolates. J. Microbiol. Biotechnol. Food Sci. 2021, 11, e653. [Google Scholar] [CrossRef]
- Khan, I.; Bahuguna, A.; Kumar, P.; Bajpai, V.K.; Kang, S.C. Antimicrobial Potential of Carvacrol against Uropathogenic Escherichia coli via Membrane Disruption, Depolarization, and Reactive Oxygen Species Generation. Front. Microbiol. 2017, 8, 2421. [Google Scholar] [CrossRef] [PubMed]
- Köse, E.O. In Vitro Activity of Carvacrol in Combination with Meropenem against Carbapenem-Resistant Klebsiella pneumoniae. Folia Microbiol. 2022, 67, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.; Sun, Z.; Wang, T.; Yang, M.; Liu, M.; Zhang, J.; Li, Y. Antimicrobial Activity of Eugenol against Carbapenem-Resistant Klebsiella pneumoniae and Its Effect on Biofilms. Microb. Pathog. 2020, 139, 103924. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, G.; Rajivgandhi, G.N.; Murugan, S.; Alharbi, N.S.; Kadaikunnan, S.; Khaled, J.M.; Almanaa, T.N.; Manoharan, N.; Li, W.J. Anti-Carbapenamase Activity of Camellia Japonica Essential Oil against Isolated Carbapenem Resistant Klebsiella pneumoniae (MN396685). Saudi J. Biol. Sci. 2020, 27, 2269–2279. [Google Scholar] [CrossRef]
- Sara, M.; Yamina, B.; Ramazan, E.; Mesut, G.; Selma, A. Dietary Risk of Bla ESBL Producing Multidrug Resistant Enterobacteriaceae and Their Inhibition by Artemisia Herba-Alba and Thymus Algeriensis Essential Oils. J. Essent. Oil Bear. Plants 2021, 24, 658–670. [Google Scholar] [CrossRef]
- Vazquez, N.M.; Mariani, F.; Torres, P.S.; Moreno, S.; Galvan, E.M. Cell Death and Biomass Reduction in Biofilms of Multidrug Resistant Extended Spectrum β-Lactamase-Producing Uropathogenic Escherichia coli Isolates by 1,8-Cineole. PLoS ONE 2020, 15, e0241978. [Google Scholar] [CrossRef]
- Vázquez-López, R.; Solano-Gálvez, S.G.; Vignon-Whaley, J.J.J.; Vaamonde, J.A.A.; Alonzo, L.A.P.; Reséndiz, A.R.; Álvarez, M.M.; López, E.N.V.; Franyuti-Kelly, G.; Álvarez-Hernández, D.A.; et al. Acinetobacter baumannii Resistance: A Real Challenge for Clinicians. Antibiotics 2020, 9, 205. [Google Scholar] [CrossRef]
- Lee, C.R.; Lee, J.H.; Park, M.; Park, K.S.; Bae, I.K.; Kim, Y.B.; Cha, C.J.; Jeong, B.C.; Lee, S.H. Biology of Acinetobacter baumannii: Pathogenesis, Antibiotic Resistance Mechanisms, and Prospective Treatment Options. Front. Cell. Infect. Microbiol. 2017, 7, 55. [Google Scholar] [CrossRef]
- Ramirez, M.S.; Bonomo, R.A.; Tolmasky, M.E. Carbapenemases: Transforming Acinetobacter baumannii into a yet More Dangerous Menace. Biomolecules 2020, 10, 720. [Google Scholar] [CrossRef]
- Kyriakidis, I.; Vasileiou, E.; Pana, Z.D.; Tragiannidis, A. Acinetobacter baumannii Antibiotic Resistance Mechanisms. Pathogens 2021, 10, 373. [Google Scholar] [CrossRef]
- Cai, Y.; Chai, D.; Wang, R.; Liang, B.; Bai, N. Colistin Resistance of Acinetobacter baumannii: Clinical Reports, Mechanisms and Antimicrobial Strategies. J. Antimicrob. Chemother. 2012, 67, 1607–1615. [Google Scholar] [CrossRef]
- Ibrahim, S.; Al-Saryi, N.; Al-Kadmy, I.M.S.; Aziz, S.N. Multidrug-Resistant Acinetobacter baumannii as an Emerging Concern in Hospitals. Mol. Biol. Rep. 2021, 48, 6987–6998. [Google Scholar] [CrossRef]
- Yang, C.H.; Su, P.W.; Moi, S.H.; Chuang, L.Y. Biofilm Formation in Acinetobacter baumannii: Genotype-Phenotype Correlation. Molecules 2019, 24, 1849. [Google Scholar] [CrossRef]
- Gómara, M.; Ramón-García, S. The FICI Paradigm: Correcting Flaws in Antimicrobial in vitro Synergy Screens at Their Inception. Biochem. Pharmacol. 2019, 163, 299–307. [Google Scholar] [CrossRef]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.J.; Cheng, Z. Antibiotic Resistance in Pseudomonas aeruginosa: Mechanisms and Alternative Therapeutic Strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef]
- Stover, C.K.; Pham, X.Q.; Erwin, A.L.; Mizoguchi, S.D.; Warrener, P.; Hickey, M.J.; Brinkman, F.S.L.; Hufnagle, W.O.; Kowallk, D.J.; Lagrou, M.; et al. Complete Genome Sequence of Pseudomonas aeruginosa PAO1, an Opportunistic Pathogen. Nature 2000, 406, 959–964. [Google Scholar] [CrossRef]
- Thi, M.T.T.; Wibowo, D.; Rehm, B.H.A. Pseudomonas aeruginosa Biofilms. Int. J. Mol. Sci. 2020, 21, 8671. [Google Scholar] [CrossRef]
- Botelho, J.; Grosso, F.; Peixe, L. Antibiotic Resistance in Pseudomonas aeruginosa–Mechanisms, Epidemiology and Evolution. Drug Resist. Updat. 2019, 44, 100640. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Brinkman, F.S.L. Function of Pseudomonas Porins in Uptake and Efflux. Annu. Rev. Microbiol. 2002, 56, 17–38. [Google Scholar] [CrossRef]
- Dreier, J.; Ruggerone, P. Interaction of Antibacterial Compounds with RND Efflux Pumps in Pseudomonas aeruginosa. Front. Microbiol. 2015, 6, 660. [Google Scholar] [CrossRef]
- Langendonk, R.F.; Neill, D.R.; Fothergill, J.L. The Building Blocks of Antimicrobial Resistance in Pseudomonas aeruginosa: Implications for Current Resistance-Breaking Therapies. Front. Cell. Infect. Microbiol. 2021, 11, 665759. [Google Scholar] [CrossRef]
- Podschun, R.; Ullmann, U. Klebsiella Spp. as Nosocomial Pathogens. Clin. Microbiol. Rev. 1998, 11, 589–603. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, T.; Chen, L.; Du, H. Virulence Factors in Hypervirulent Klebsiella pneumoniae. Front. Microbiol. 2021, 12, 642484. [Google Scholar] [CrossRef]
- Lam, M.M.C.; Wick, R.R.; Watts, S.C.; Cerdeira, L.T.; Wyres, K.L.; Holt, K.E. A Genomic Surveillance Framework and Genotyping Tool for Klebsiella pneumoniae and Its Related Species Complex. Nat. Commun. 2021, 12, 4188. [Google Scholar] [CrossRef]
- Martin, R.M.; Bachman, M.A. Colonization, Infection, and the Accessory Genome of Klebsiella pneumoniae. Front. Cell. Infect. Microbiol. 2018, 8, 4. [Google Scholar] [CrossRef]
- Shon, A.S.; Bajwa, R.P.S.; Russo, T.A. Hypervirulent (Hypermucoviscous) Klebsiella pneumoniae: A New and Dangerous Breed. Virulence 2013, 4, 107–118. [Google Scholar] [CrossRef]
- Wang, G.; Zhao, G.; Chao, X.; Xie, L.; Wang, H. The Characteristic of Virulence, Biofilm and Antibiotic Resistance of Klebsiella pneumoniae. Int. J. Environ. Res. Public Health 2020, 17, 6278. [Google Scholar] [CrossRef]
- Gomez-Simmonds, A.; Uhlemann, A.C. Clinical Implications of Genomic Adaptation and Evolution of Carbapenem-Resistant Klebsiella pneumoniae. J. Infect. Dis. 2017, 215 (Suppl. 1), S18–S27. [Google Scholar] [CrossRef]
- Llobet, E.; Martínez-Moliner, V.; Moranta, D.; Dahlström, K.M.; Regueiro, V.; Tomása, A.; Cano, V.; Pérez-Gutiérrez, C.; Frank, C.G.; Fernández-Carrasco, H.; et al. Deciphering Tissue-Induced Klebsiella pneumoniae Lipid a Structure. Proc. Natl. Acad. Sci. USA 2015, 112, E6369–E6378. [Google Scholar] [CrossRef]
- Paczosa, M.K.; Mecsas, J. Klebsiella Pneumoniae: Going on the Offense with a Strong Defense. Microbiol. Mol. Biol. Rev. 2016, 80, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Nirwati, H.; Sinanjung, K.; Fahrunissa, F.; Wijaya, F.; Napitupulu, S.; Hati, V.P.; Hakim, M.S.; Meliala, A.; Aman, A.T.; Nuryastuti, T. Biofilm Formation and Antibiotic Resistance of Klebsiella pneumoniae Isolated from Clinical Samples in a Tertiary Care Hospital, Klaten, Indonesia. BMC Proc. 2019, 13 (Suppl. 11), 20. [Google Scholar] [CrossRef] [PubMed]
- Wyres, K.L.; Holt, K.E. Klebsiella pneumoniae as a Key Trafficker of Drug Resistance Genes from Environmental to Clinically Important Bacteria. Curr. Opin. Microbiol. 2018, 45, 131–139. [Google Scholar] [CrossRef]
- Srinivasan, V.B.; Venkataramaiah, M.; Mondal, A.; Vaidyanathan, V.; Govil, T.; Rajamohan, G. Functional Characterization of a Novel Outer Membrane Porin KpnO, Regulated by PhoBR Two-Component System in Klebsiella pneumoniae NTUH-K2044. PLoS ONE 2012, 7, e41505. [Google Scholar] [CrossRef]
- Peirano, G.; Ahmed-Bentley, J.; Fuller, J.; Rubin, J.E.; Pitout, J.D.D. Travel-Related Carbapenemase-Producing Gram-Negative Bacteria in Alberta, Canada: The First 3 Years. J. Clin. Microbiol. 2014, 52, 1575–1581. [Google Scholar] [CrossRef]
- Mazzariol, A.; Zuliani, J.; Cornaglia, G.; Rossolini, G.M.; Fontana, R. AcrAB Efflux System: Expression and Contribution to Fluoroquinolone Resistance in Klebsiella spp. Antimicrob. Agents Chemother. 2002, 46, 3984–3986. [Google Scholar] [CrossRef]
- Ping, Y.; Ogawa, W.; Kuroda, T.; Tsuchiya, T. Gene Cloning and Characterization of KdeA, a Multidrug Efflux Pump from Klebsiella pneumoniae. Biol. Pharm. Bull. 2007, 30, 1962–1964. [Google Scholar] [CrossRef]
- Wong, M.H.Y.; Chan, E.W.C.; Chen, S. Evolution and Dissemination of OqxAB-like Efflux Pumps, an Emerging Quinolone Resistance Determinant among Members of Enterobacteriaceae. Antimicrob. Agents Chemother. 2015, 59, 3290–3297. [Google Scholar] [CrossRef]
- Martínez-Martínez, L.; Hernández-Allés, S.; Albertí, S.; Tomás, J.M.; Benedi, V.J.; Jacoby, G.A. In Vivo Selection of Porin-Deficient Mutants of Klebsiella pneumoniae with Increased Resistance to Cefoxitin and Expanded-Spectrum Cephalosporins. Antimicrob. Agents Chemother. 1996, 40, 342–348. [Google Scholar] [CrossRef]
- Nam, Y.S.; Cho, S.Y.; Yang, H.Y.; Park, K.S.; Jang, J.H.; Kim, Y.T.; Jeong, J.W.; Suh, J.T.; Lee, H.J. Investigation of Mutation Distribution in DNA Gyrase and Topoisomerase IV Genes in Ciprofloxacin-Non-Susceptible Enterobacteriaceae Isolated from Blood Cultures in a Tertiary Care University Hospital in South Korea, 2005–2010. Int. J. Antimicrob. Agents 2013, 41, 126–129. [Google Scholar] [CrossRef]
- Clements, A.; Tull, D.; Jenney, A.W.; Farn, J.L.; Kim, S.H.; Bishop, R.E.; McPhee, J.B.; Hancock, R.E.W.; Hartland, E.L.; Pearse, M.J.; et al. Secondary Acylation of Klebsiella pneumoniae Lipopolysaccharide Contributes to Sensitivity to Antibacterial Peptides. J. Biol. Chem. 2007, 282, 15569–15577. [Google Scholar] [CrossRef] [PubMed]
- Navon-Venezia, S.; Kondratyeva, K.; Carattoli, A. Klebsiella Pneumoniae: A Major Worldwide Source and Shuttle for Antibiotic Resistance. FEMS Microbiol. Rev. 2017, 41, 252–275. [Google Scholar] [CrossRef]
- Wyres, K.L.; Lam, M.M.C.; Holt, K.E. Population Genomics of Klebsiella pneumoniae. Nat. Rev. Microbiol. 2020, 18, 344–359. [Google Scholar] [CrossRef]
- David, S.; Reuter, S.; Harris, S.R.; Glasner, C.; Feltwell, T.; Argimon, S.; Abudahab, K.; Goater, R.; Giani, T.; Errico, G.; et al. Epidemic of Carbapenem-Resistant Klebsiella pneumoniae in Europe Is Driven by Nosocomial Spread. Nat. Microbiol. 2019, 4, 1919–1929. [Google Scholar] [CrossRef]
- Reyes, J.; Aguilar, A.C.; Caicedo, A. Carbapenem-Resistant Klebsiella pneumoniae: Microbiology Key Points for Clinical Practice. Int. J. Gen. Med. 2019, 12, 437–446. [Google Scholar] [CrossRef]
- Xu, L.; Sun, X.; Ma, X. Systematic Review and Meta-Analysis of Mortality of Patients Infected with Carbapenem-Resistant Klebsiella pneumoniae. Ann. Clin. Microbiol. Antimicrob. 2017, 16, 18. [Google Scholar] [CrossRef]
- Ernst, C.M.; Braxton, J.R.; Rodriguez-Osorio, C.A.; Zagieboylo, A.P.; Li, L.; Pironti, A.; Manson, A.L.; Nair, A.V.; Benson, M.; Cummins, K.; et al. Adaptive Evolution of Virulence and Persistence in Carbapenem-Resistant Klebsiella pneumoniae. Nat. Med. 2020, 26, 705–711. [Google Scholar] [CrossRef]
- Allocati, N.; Masulli, M.; Alexeyev, M.F.; Di Ilio, C. Escherichia coli in Europe: An Overview. Int. J. Environ. Res. Public Health 2013, 10, 6235–6254. [Google Scholar] [CrossRef]
- Denamur, E.; Clermont, O.; Bonacorsi, S.; Gordon, D. The Population Genetics of Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2021, 19, 37–54. [Google Scholar] [CrossRef]
- Lee, S.Y. Systems Biology and Biotechnology of Escherichia coli; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–462. [Google Scholar] [CrossRef]
- Liu, B.; Furevi, A.; Perepelov, A.V.; Guo, X.; Cao, H.; Wang, Q.; Reeves, P.R.; Knirel, Y.A.; Wang, L.; Widmalm, G. Structure and Genetics of Escherichia coli O Antigens. FEMS Microbiol. Rev. 2020, 44, 655–683. [Google Scholar] [CrossRef]
- Sharma, G.; Sharma, S.; Sharma, P.; Chandola, D.; Dang, S.; Gupta, S.; Gabrani, R. Escherichia Coli Biofilm: Development and Therapeutic Strategies. J. Appl. Microbiol. 2016, 121, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Choi, U.; Lee, C.R. Distinct Roles of Outer Membrane Porins in Antibiotic Resistance and Membrane Integrity in Escherichia coli. Front. Microbiol. 2019, 10, 953. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Madec, J.-Y.; Lupo, A.; Schink, A.-K.; Kieffer, N.; Nordmann, P.; Schwarz, S. Antimicrobial Resistance in Escherichia coli. Microbiol. Spectr. 2018, 6, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, G.A. AmpC Β-Lactamases. Clin. Microbiol. Rev. 2009, 22, 161–182. [Google Scholar] [CrossRef] [PubMed]
- Dagher, C.; Salloum, T.; Alousi, S.; Arabaghian, H.; Araj, G.F.; Tokajian, S. Molecular Characterization of Carbapenem Resistant Escherichia coli Recovered from a Tertiary Hospital in Lebanon. PLoS ONE 2018, 13, e0203323. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Zheng, X.; Sun, Y.; Fang, R.; Zhang, S.; Zhang, X.; Lin, J.; Cao, J.; Zhou, T. Molecular Mechanisms and Epidemiology of Carbapenem-Resistant Escherichia coli Isolated from Chinese Patients during 2002–2017. Infect. Drug Resist. 2020, 13, 501–512. [Google Scholar] [CrossRef]
- Gurung, S.; Kafle, S.; Dhungel, B.; Adhikari, N.; Shrestha, U.T.; Adhikari, B.; Banjara, M.R.; Rijal, K.R.; Ghimire, P. Detection of Oxa-48 Gene in Carbapenem-Resistant Escherichia coli and Klebsiella pneumoniae from Urine Samples. Infect. Drug Resist. 2020, 13, 2311–2321. [Google Scholar] [CrossRef]
- Lombrea, A.; Antal, D.; Ardelean, F.; Avram, S.; Pavel, I.Z.; Vlaia, L.; Mut, A.M.; Diaconeasa, Z.; Dehelean, C.A.; Soica, C.; et al. A Recent Insight Regarding the Phytochemistry and Bioactivity of Origanum vulgare L. Essential Oil. Int. J. Mol. Sci. 2020, 21, 9653. [Google Scholar] [CrossRef]
- Ojeda-Sana, A.M.; van Baren, C.M.; Elechosa, M.A.; Juárez, M.A.; Moreno, S. New Insights into Antibacterial and Antioxidant Activities of Rosemary Essential Oils and Their Main Components. Food Control 2013, 31, 189–195. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. Syst. Rev. 2021, 10, 89. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Bacteria | % of Bacteria Resistant to 3rd Generation Cephalosporins | Country |
|---|---|---|
| Klebsiella pneumoniae | 79.1 | Bulgaria |
| 74.5 | Greece | |
| 67.9 | Romania | |
| 63.0 | Poland | |
| 54.7 | Cyprus | |
| 54.4 | Slovakia | |
| Escherichia coli | 41.4 | Bulgaria |
| 29.8 | Cyprus | |
| 27.1 | Slovakia | |
| 26.4 | Italy | |
| 24.1 | Latvia | |
| 21.9 | Greece |
| Bacteria | % of Bacteria Resistant to Carbapenems | Country |
|---|---|---|
| Acinetobacter spp. | 96.4 | Croatia |
| 94.6 | Greece | |
| 93.3 | Romania | |
| 91.1 | Lithuania | |
| 82.9 | Bulgaria | |
| 82.7 | Latvia | |
| Pseudomonas aeruginosa | 48.9 | Slovakia |
| 43.9 | Romania | |
| 42.9 | Bulgaria | |
| 35.7 | Greece | |
| 33.8 | Hungary | |
| 30.3 | Croatia | |
| Klebsiella pneumoniae | 66.3 | Greece |
| 48.3 | Romania | |
| 29.5 | Italy | |
| 28.1 | Bulgaria | |
| 19.8 | Cyprus | |
| 19.1 | Croatia | |
| Escherichia coli | 0.8 | Bulgaria |
| 0.7 | Romania | |
| 0.5 | Greece | |
| 0.5 | Italy | |
| 0.4 | Spain | |
| 0.2 | Portugal |
| Study Team and Year | Bacterial Strain | Essential Oil(s)/ Pure Phytocompound | Method(s) | Results | Reference |
|---|---|---|---|---|---|
| Carbapenem-Resistant Acinetobacter baumannii | |||||
| Anchana SR et al., 2021 | carbapenem-resistant A. baumanni | Ocimum sanctum L. compunds | Semi-quantitative adherent bioassay PCR amplification csgA gene sequencing Molinspiration assessment Cristal violet assay | Benzofuran from Ocimum sanctum L. EOs can be effectivein targeting csgA gene of carbapenem-resistant A. baumanni strains. Minum biofilm inhibition conc (MIC) of 50% was observed at 25 mcL and at 90%, 50 mcL. | [33] |
| Amaral SC et al., 2020 | carbapenem-resistant A. baumanni | Origanum vulgare L. EO | PCR for gyrB and blaOXA-51-like genes Flow cytometry Checkerboard assay | Carvacrol (71%) showed antibacterial effect against all Ab-MDR tested strains (MIC: 1.75–3.50 mg mL−1) Synergistic interaction with polymyxin B (16× ↓ in polymyxin B MIC) | [34] |
| Vasconcelos NG et al., 2019 | A. baumanii; K. pneumoniae; carbapenem-resistant Serratia marcescens | Origanum vulgare L. EO | Disk-diffusion Microdilution Time kill assays | MIC: 0.015% v/v for A. baumannii, MIC: 0.059% v/v for K. pneumoniae and S. marcescens Decrease in cell count: after 4 h treatment | [35] |
| Bekka-Hadji F et al., 2022 | imipenem-resistant A. baumannii; MRSA | Mentha pulegium L. Artemisia herba alba Asso. | Disk diffusion method Microdilution broth assay | Mentha pulegium L. EO was found to containe pulegone (74.8%) and neoisomenthol (10.0%). Artemisia herba alba Asso. EO was found to contain maily camphor (32.0%), α-thujone (13.7%), 1,8-cineole (9.8%), β-thujone (5.0%). Synergistic, antagonic or neutral effects of EOs with antibiotics were observed. The strongest: M. pulegium L. + amikacin for Imipenem-resitant A. baumannii. P. aeruginosa strains were found to be resistant to these EOs. | [36] |
| Oliva A et al., 2020 | carbapenem-resistant A. baumannii, K. pneumoniae and P. aeruginosa; MRSA; E. coli (ATCC 25922); C. albicans (ATCC 14053) | Helichrysum italicum (Roth) G. Don fil. | Dilution and disk diffusion method | The EO was analyzed in both liquid and vapour phases. Bactericidal/Fungicidal effect of EO characterized maily by β-eudesmene (21.65%) and β-bisabolene (19.90%) was observed at a concentration of 5% v/v against carbapenem- resistant A. baumannii and C. albicans. | [37] |
| Rinaldi F et al., 2020 | carbapenem-resistant A. baumannii, K. pneumoniae and P. aeruginosa; MRSA; E. coli (ATCC 25922) | Thymus vulgaris L. Syzygium aromaticum L. EOs chitosan nanoemultions | Macro dilution broth methods Disk diffusion assay Time kill assay | Both EOs nanoformulations presented a MIC/MBC for A. baumannii- and K. pneumoniae—carbapenem resistant of 0.03% v/v. Amazing concentration-dependent, antibacterial effect was observed for T. vulgaris L. nanoformulations against the two strains mentioned above. Effective intranasal formulations (promising therapeutical solutions). | [38] |
| Mahmoudi H et al., 2020 | carbapenem-resistant and fluoroquinolones-resistant A. baumannii | Mentha longifolia (L.) Hudson (Menthol) | PCR method Microbroth dilution method | MIC decreased 4× when Mentha longifolia (L.) Hudson EO was associated with ciprofloxacin and 8× when associated with imipenem. Menthol EO + imipenem reduced the resistance to imipenem up to 16× in 90% of the analyzed strains. | [39] |
| Kaskatepe B et al., 2016 | carbapenem-resistant A. baumannii and P. aeruginosa | Commercial cinnamon oil- the exact species not specified | Disk diffusion method | P. aeruginosa MIC: 0.0019 mL/mL, 21 mm zone diameter | [40] |
| Laktib A et al., 2021 | carbapenem-resistant A. baumannii | Lavandula mairei Humbert | Disk diffusion Broth microdilution method | All the tested strains showed sensitivity to the EO. MIC: 0.39–3.125 mcL/mL IDZ: 28.67–40 mm | [41] |
| Muntean D et al., 2019 | carbapenem-resistant A. baumannii, P. aeruginosa and Enterobacterales | Mentha × piperita L. | Agar disk diffusion method Microdilution method | MIC: <20 mg/mL for S. aureus, E. coli, P. mirabilis MIC: >40 mg/mL for K. pneumoniae, P. aeruginosa and A. baumannii | [42] |
| Carbapenem-Resistant Pseudomonas aeruginosa | |||||
| Patterson JE et al., 2019 | MRSA; carbapenem-resistant and ESBL-producing Enterobacterales; MDR (including carbapenem- resistant) P. aeruginosa | Cinnamomun zeylanicum L. Szygium aromaticum L. Cymbopogon flexuosus L. Origanum vulgare L. Rosmarinus officinalis L. Thymus vulgaris L. Melaleuca alternifolia (Maiden and Betche) Leptospermum scoparium J. R. et G. Forst 1 EO blend (cinnamon, clove, lemon, eucalyptus, rosemary) | Disk diffusion method | Cinnamon bark EO had the largest zone of inhibition against P. aeruginosa (18 mm). The largest inhibition’s zones for Enterobacterales spp. were observed for cinnamon bark, oregano and thyme. | [43] |
| Oliva A et al., 2018 | carbapenem-resistant P. aeruginosa, A. baumannii and K. pneumoniae; ESBL-producing and carbapenem-sensitive K. pneumoniae; MRSA | Melaleuca alternifolia (Maiden and Betche) | Macro dilution broth method Checkerboard assay Disk diffusion Vapour assay Time kill assay | Tea tree oil was reported to be an effective antimicrobial agent, used alone or in association with classical antibiotic therapy (ex: oxacillin against MRSA). Tea tree oil administered by inhalation could be an option for carbapenem— resistant A. baumannii induced pneumonia. | [44] |
| Carbapenem-Resistant Enterobacterales | |||||
| Qian W et al., 2020 | carbapenem-resistant Enterobacter cloacae | Citral (isomeric mixture of geranial and neral) | Agar dilution method Confocal laser scanning microscopy | MIC: 1000 mcg/mL. Potent antibacterial and antibiofilm activity was observed. Potential use as natural food preservative. | [45] |
| Ginting EV et al., 2021 | ESBL-producing E. coli and K. pneumoniae | Syzygium aromaticum L. Cinnamomum verum J.Presl | Disk diffusion method Broth microdilution method Scanning electron microscopy | High antibacterial activity of both EOs. Clove EO MIC: 0.078% (v/v) for all bacteria. Cinnamon EO MIC: 0.039% (v/v)–0.156 (v/v) for all bacteria | [46] |
| Sharifi-Rad J et al., 2016 | ESBL-producing E. coli | Achillea wilhelmsii C. Koch, Echinophora platyloba DC., Lallemantia royleana, Nepeta persica Boiss., Pulicaria vulgaris Gaertn., Salvia nemorosa L., Satureja intermedia C.A.Mey | Microdilution method Phenotypic disc confirmatory test Polymerase chain reaction for TEM gene | All tested EOs presented high antimicrobial activity | [47] |
| Kwiatkowski P et al., 2018 | ESBL and New Delhi metallo-β-lactamase-1 (NDM-1) producing K. pneumoniae | Carum carvi L., Foeniculum vulgare Mill., Mentha × piperita L., Pelargonium graveolens L’Hér. Ocimum basilicum L., Syzygium aromaticum L. Merrill and Perry, Thymus vulgaris L., Salvia sclarea L., Lavandula angustifolia Mill. | Broth microdilution method Checkerboard method | Peppermint oil + gentamicin induced synergistic effects against all tested strains. Caraway EO + gentamicin induced synergistic effects against ESBL-strains and gentamicin-resistant strains. Association of gentamicin + thyme, basil, fennel and clary sage induced an additive antimicrobial effect. | [48] |
| Benameur Q et al., 2019 | blaESBL-producing Enterobacterales strains | Thymus vulgaris L. alone or associated with cefotaxime | Disc diffusion assay PCR Checkerboard test | Thymus vulgaris L. EO showed increased antimicrobial activity against all tested strains (MIC: 24–40 mm/10 mcL for MDR strains and 2.87–11.5 mcg/mL for blaESBL producing strains). Synergistic activity was observed when Thymus vulgaris L. EO was associated with cefotaxime for blaESBL-producing E. coli strains, as well as an additive activity for ESBL-producing Enterobacter cloacae | [49] |
| Aouadhi C et al., 2022 | ESBL-producing E. coli | Eucalyptus globulus L., Eucalyptus camaldulensis Dehnh., Artemisia absinthium L., Myrtuscommunis L., Mentha pulegium L., Trachyspermum ammi (L.) Sprague, Cymbopogon citratus (DC.) Stapf, Thymbra capitata (L.) Cav. | Disc-diffusion assay Broth dilution method Time-kill study Bacteriolysis | Thymbra capitata (L.) Cav. EOs induced the most potent inhibitory effect against E. coli strains (MIC: 0.02–0.78%) by reducing its viability, NaCl tolerance, enhancing the loss of 260 nm-absorbing material and sensitive to autolysis. Thymbra capitata (L.) Cav. EO was proposed as food preservative (especially in meat). | [50] |
| Saliu EM et al., 2020 | ESBL-producing E. coli; Salmonella typhimurium inoculated in cecal contents of 2 weeks old broilers | Carvacrol Cinnamaldehyde Eugenol Lactobacillus agilis LA73 Lactobacillus salivarius LS1 used as feed additives (alone or in combination) | Inoculation Aerobic co-incubation | The experimental diet highlighted a lower ESBL-prevalence in the study group of animals (broilers) when EOs and Lactobacillus spp. were added. Horizontal gene transfer effect was less obvious. | [51] |
| Motola G et al., 2020 | ESBL producing E. coli | Essential oils used as spray or cold mist disinfectant method | - | The essential oil cold fog disinfecting method managed to partly reach the expected efficacy threshold in reducing ESBL-producing E. coli contamination on the hatching egg. | [52] |
| Krishnamoorthy R et al., 2018 | ESBL-producing E. coli, K. pneumoniae and P. aeruginosa; MRSA | Cleome viscosa L. essential oil nanoemulsion | Agar well diffusion method Broth dilution method | The nanoemultion was effective against the tested strains by inhibiting the drug efflux mechanism. Nanofromulation from Cleome viscosa L. proved to induce a broad-spectrum antimicrobial activity against the tested strains. Gram-negative ESBL-producing strains: MIC: 30 mcg/mL; MBC: 40 mcg/mL; MRSA MIC: 25 mcg/mL; MBC: 40 mcg/mL, including ATCC control strain, the nanoemulsion being more effective against Gram-positive bacteria pathogens. | [53] |
| Tebrün W et al., 2020 | ESBL-producing E. coli | Essential oil spray (a mixture of Origanum vulgare L. oil and alcohol) | - | Decreased hatchability was observed for essential oil spray application on egg broiler chicks, leading it to be inappropriate in daily practice, compared to the application of hydrogen peroxide as disinfectant protocol. | [54] |
| Shrivastav A et al., 2019 | ESBL-producing E. coli | Syzygium aromaticum L. EO and Ocimum sanctum L. fresh jouice | Colorimetric assay | Higher inhibitory activity observed for S. aromaticum L. EO on beta-lactamase enzyme of cecal samples of healthy broilers. | [55] |
| Kaskatepe B et al., 2017 | ESBL-producing E. coli | Origanum onites L. | Disc diffusion Agar dilution method Micro-dilution method | Origanum onites L. EO presented antimicrobial activity against all tested strains and also inhibited microbial growth on ESBL-producing E. coli strains. MIC: 1.56–25 mcL/mL; 3.12–25 mcL/mL | [56] |
| Tadić V et al., 2017 | carbapenem-resistant K. pneumoniae; E. coli (ATCC 25922); MRSA | Sideritis romana L. subsp. purpurea (Tal. ex Benth.) Heywood | Mueller Hinton broth | Good antimicrobial effect against MRSA with MIC: 0.076 mg/mL and MBC: 0.153 mg/mL. No effect on carbapenem resistant K. pneumoniae or E. coli. | [57] |
| Iseppi R et al., 2020 | ESBL-producing E. coli and K. pneumoniae; Carbapenamase-producing K. pneumoniae; metallo-beta-lactamase-producing P. aeruginosa | Melaleuca alternifolia (Maiden and Betche), Eucalyptus globulus L., Mentha × piperita L., Thymus vulgaris L. | Agar disk diffusion assay | Melaleuca alternifolia (Maiden and Betche) and Thymus vulgaris L. EOs were the most effective on the tested strains, the first EO being effective even at low concentrations. Melaleuca alternifolia (Maiden and Betche) MIC: 0.5–16 mcg/mL. Thymus vulgaris L. MIC: 1–16 mcg/mL. Both EOs presented also a good anti-biofilm activity, making them good candidates for infections caused by these pathogens even in association with classical antibiotics. | [58] |
| Gāliņa D et al., 2022 | ESBL-producing E. coli | Thymus serpyllum L. Thymus vulgaris L. Satureja montana L. alone or in combination with caprylic acid and sodium chloride | Broth microdilution method | The antimicrobial activity of tested EOs was superior when sodium chloride or caprylic acid was added. All tested EOs showed good antimicrobial activity against ESBL and non-ESBL-producing, as well as MDR and non-MDR E. coli strains. | [59] |
| Benameur Q et al., 2021 | ESBL-producing Enterobacterales | Origanum vulgare L. | Disc diffusion assay Twofold serial dilution method | High antibacterial effect against all studied strains MIC: 0.31–5 mcL/mL, ESBL-producing strains being more responsive to EO. | [60] |
| Contreras-Moreno BL et al., 2016 | ESBL-producing E. coli and Enterobacter cloacae; MRSA | Pimenta racemosa var. racemose (Mill.) of different densities | Disc diffusion agar method | Antibacterial effect on all the tested strains of nosocomial provenience. MIC: 20–400 mcL/mL. | [61] |
| de Souza et al., 2021 | carbapenem-resistant K. pneumoniae | Carvacrol | Broth microdilution method Time-kill assay Mouse model of infection | Antibacterial effect on all the tested strains with eradication of all bacterial cells within 4 h. MICs/MBCs: 130–260 mg/L. The in vivo effect of carvacrol determined through a mouse model of infection induced an increased survival and a decreased bacterial load in the peritoneal lavage. Moreover, leucopenia was observed, as well as increased number of platelets when compared with the control group. | [62] |
| Dhara L et al., 2020 | ESBL-producing and quinolone-resistant Enterobacterales | Cinnamaldehyde Eugenol | Broth microdilution method Murine model toxicity level | E. coli MIC: 7.28 mcg/mL (cinnamaldehyde); 7.34 mcg/mL (eugenol). K. pneumoniae MIC: 0.91 mcg/mL (cinnamaldehyde); 3.67 mcg/mL (eugenol). Good safety profiles manifested by both EOs in a murine model (Swiss albino mice). | [63] |
| Dhara L, Tripathi A et al., 2020 | ESBL-producing and quinolone-resistant Enterobacterales | Cinnamaldehyde alone or in combination with cefotaxime/ciprofloxacin | Broth microdilution method Checkerboard assay Isobologram analysis Time-kill assay | Synergism of cinnamaldehyde with cefotaxime was observed. Cinnamaldehyde managed to decrease MIC of cefotaxime and ciprofloxacin up to 1024×, the bactericidal and synergistic effect being observed after 24 h. | [64] |
| Gore MR et al., 2021 | ESBL-producing Enterobacterales | Pimenta dioica (L.) Merr. extracts | Agar dilution method | Chloroform extract induced the maximal antibacterial activity, with MBC: 2–5 mg/mL. Ampicillin MBC decreased from 10 mg/mL to 300–500 mcg/mL in the presence of Pimenta dioica (L.) Merr. Chloroform extracts and thus it induced a synergistic action. Eugenol was observed to be the major compound of the extract. EO induced a deformation in the bacterial cell membrane (stress and cellular damage). | [65] |
| Khan I et al., 2017 | ESBL-producing E. coli | Carvacrol | Broth microdilution method | MIC: 450 mcg/mL, time-dependent effect (after 2 h it managed to completely diminish the growth in E. coli strains). The antibacterial effect of carvacrol on the tested strains was induced by high level of reactive oxygen species and bacterial cell membrane depolarization, leading to its disruption and release of cellular material. | [66] |
| Kose EO et al., 2021 | carbapenem-resistant K. pneumoniae | Carvacrol + meropenem | Broth microdilution method Checkerboard assay Time-kill assay | Carvacrol + meropenem MIC: 32–128 mcg/mL, with a synergy between the 2 substances observed in 8 of the 25 tested strains. Cell membrane damage was observed. | [67] |
| Qian W et al., 2020 | carbapenem-resistant K. pneumoniae | Eugenol | Agar dilution method | Eugenol MICs: 0.2 mg/mL. Strong inhibitory effects on biofilm formation and biofilm- associated gene expression. | [68] |
| Ramachandran G et al., 2020 | carbapenem-resistant K. pneumoniae | Camellia japonica L. | Disc diffusion Agar well method Micro broth dilution method | MIC: 50 mcg/mL, concentration dependent effect on membrane destruction. | [69] |
| Sara M et al., 2021 | ESBL-producing Enterobacterales | Artemesia herba-alba Asso. Thymus algeriensis Boiss and Reut. | Disc diffusion Microdilution methods | K. pneumoiae SB6 had the highest zones of inhibition. Both EOs MIC: 1.56–12.5 mg/mL. A. herba-alba MBC: 6.25–25 mg/m. T. algeriensis MBC: 12.5–25 mg/mL. Possible new natural preservative solution for dairy products, although in vivo studies are needed in order to determine their safety profile as well as their acceptability of the aroma and flavor of EOs by the consumer. | [70] |
| Vasquez NM et al., 2020 | ESBL-producing E. coli | Rosmarinus officinalis L. | Broth microdilution method | 1,8-cineole (eucalyptol) presents antimicrobial [MIC: 0.8 (v/v)] and antibiofilm activities against uropathogenic E. coli ESBL-producing strains. The antibiofilm activity has been reported to be concentration-dependent as well as time-dependent over pre-formed biofilm, suggesting it to be a good candidate for E. coli biofilm infections. | [71] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Badescu, B.; Buda, V.; Romanescu, M.; Lombrea, A.; Danciu, C.; Dalleur, O.; Dohou, A.M.; Dumitrascu, V.; Cretu, O.; Licker, M.; et al. Current State of Knowledge Regarding WHO Critical Priority Pathogens: Mechanisms of Resistance and Proposed Solutions through Candidates Such as Essential Oils. Plants 2022, 11, 1789. https://doi.org/10.3390/plants11141789
Badescu B, Buda V, Romanescu M, Lombrea A, Danciu C, Dalleur O, Dohou AM, Dumitrascu V, Cretu O, Licker M, et al. Current State of Knowledge Regarding WHO Critical Priority Pathogens: Mechanisms of Resistance and Proposed Solutions through Candidates Such as Essential Oils. Plants. 2022; 11(14):1789. https://doi.org/10.3390/plants11141789
Chicago/Turabian StyleBadescu, Bianca, Valentina Buda, Mirabela Romanescu, Adelina Lombrea, Corina Danciu, Olivia Dalleur, Angele Modupe Dohou, Victor Dumitrascu, Octavian Cretu, Monica Licker, and et al. 2022. "Current State of Knowledge Regarding WHO Critical Priority Pathogens: Mechanisms of Resistance and Proposed Solutions through Candidates Such as Essential Oils" Plants 11, no. 14: 1789. https://doi.org/10.3390/plants11141789
APA StyleBadescu, B., Buda, V., Romanescu, M., Lombrea, A., Danciu, C., Dalleur, O., Dohou, A. M., Dumitrascu, V., Cretu, O., Licker, M., & Muntean, D. (2022). Current State of Knowledge Regarding WHO Critical Priority Pathogens: Mechanisms of Resistance and Proposed Solutions through Candidates Such as Essential Oils. Plants, 11(14), 1789. https://doi.org/10.3390/plants11141789