
Terpene Profiles Composition and Micromorphological Analysis on Two Wild Populations of Helichrysum spp. from the Tuscan Archipelago (Central Italy)

 ,
,  ,
,  , ,
, , 
 and
and
Abstract
1. Introduction
2. Results
2.1. Morphological Analysis
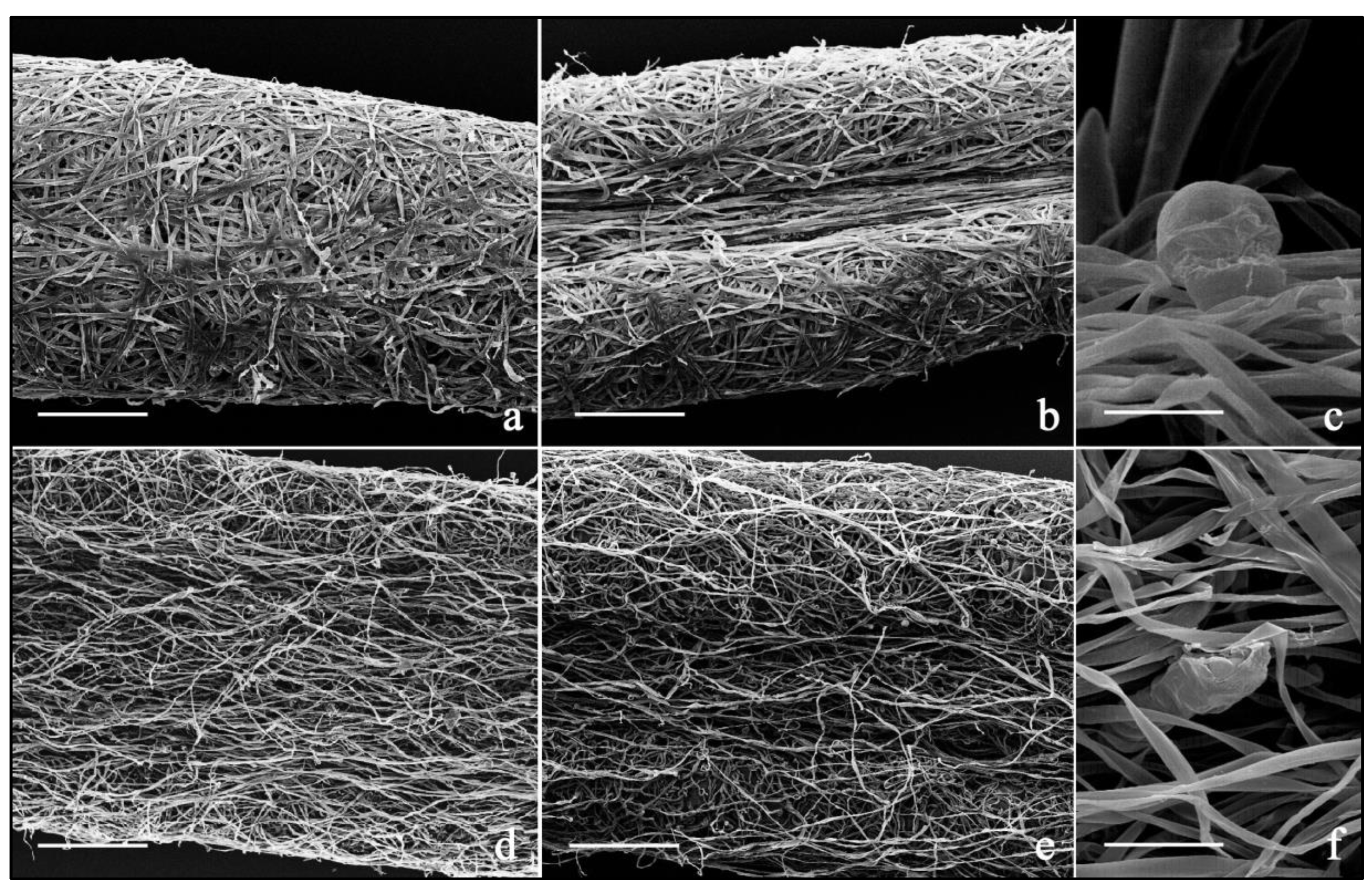
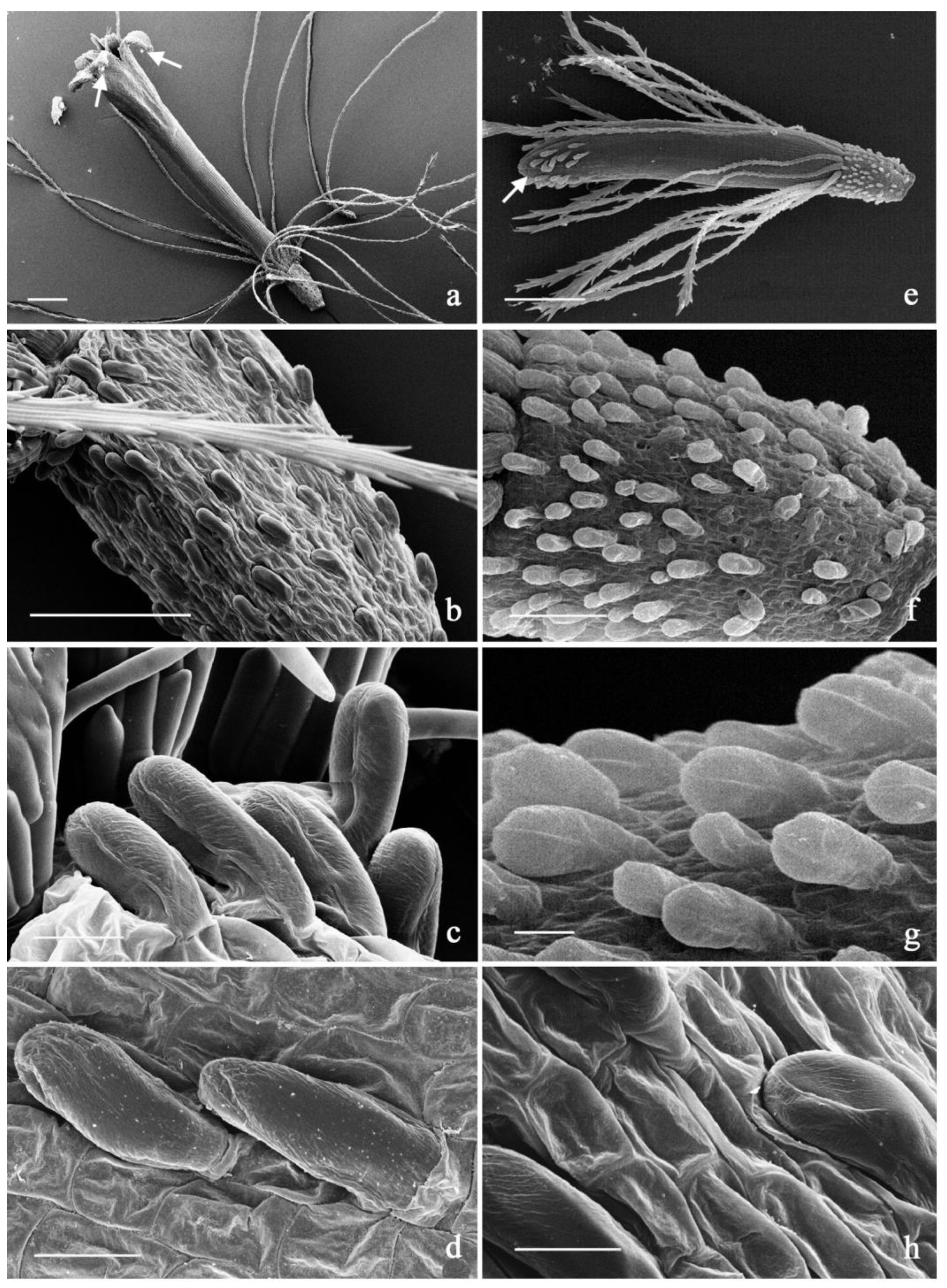
2.2. Micromorphological Analysis
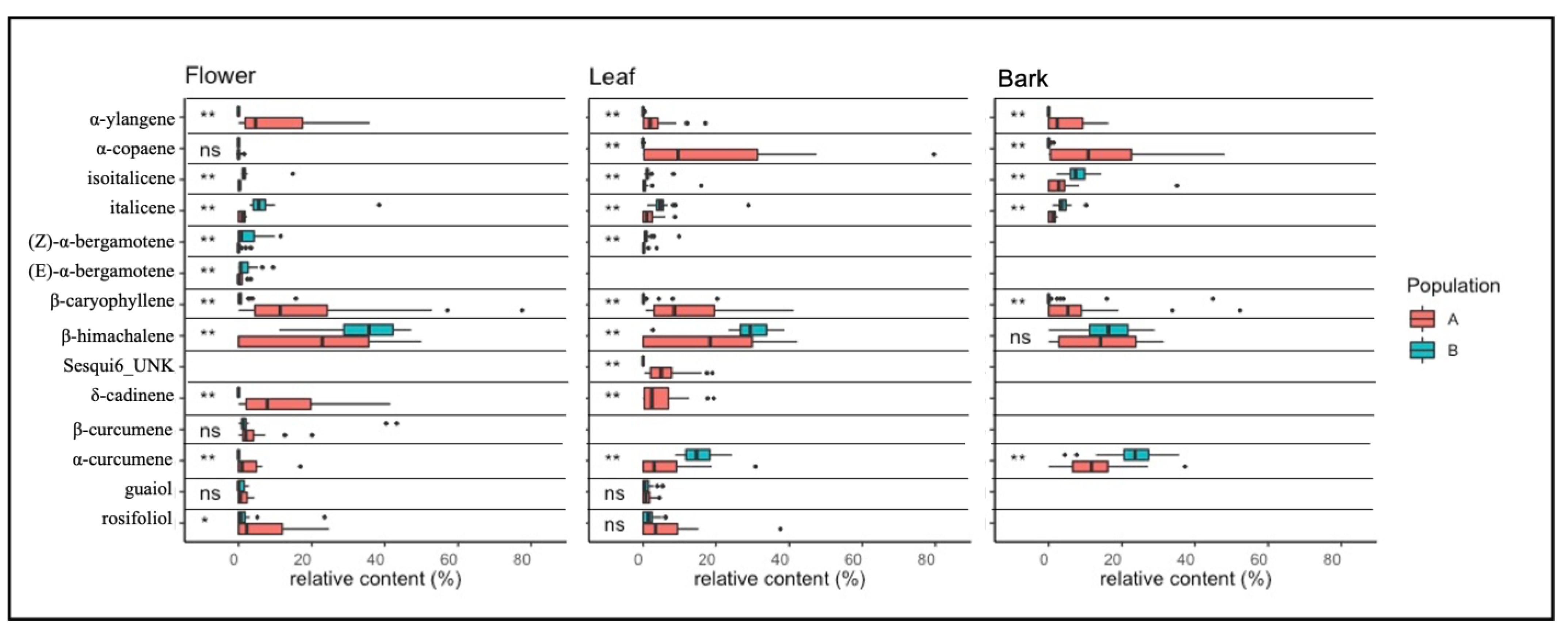
2.3. Terpene Analysis
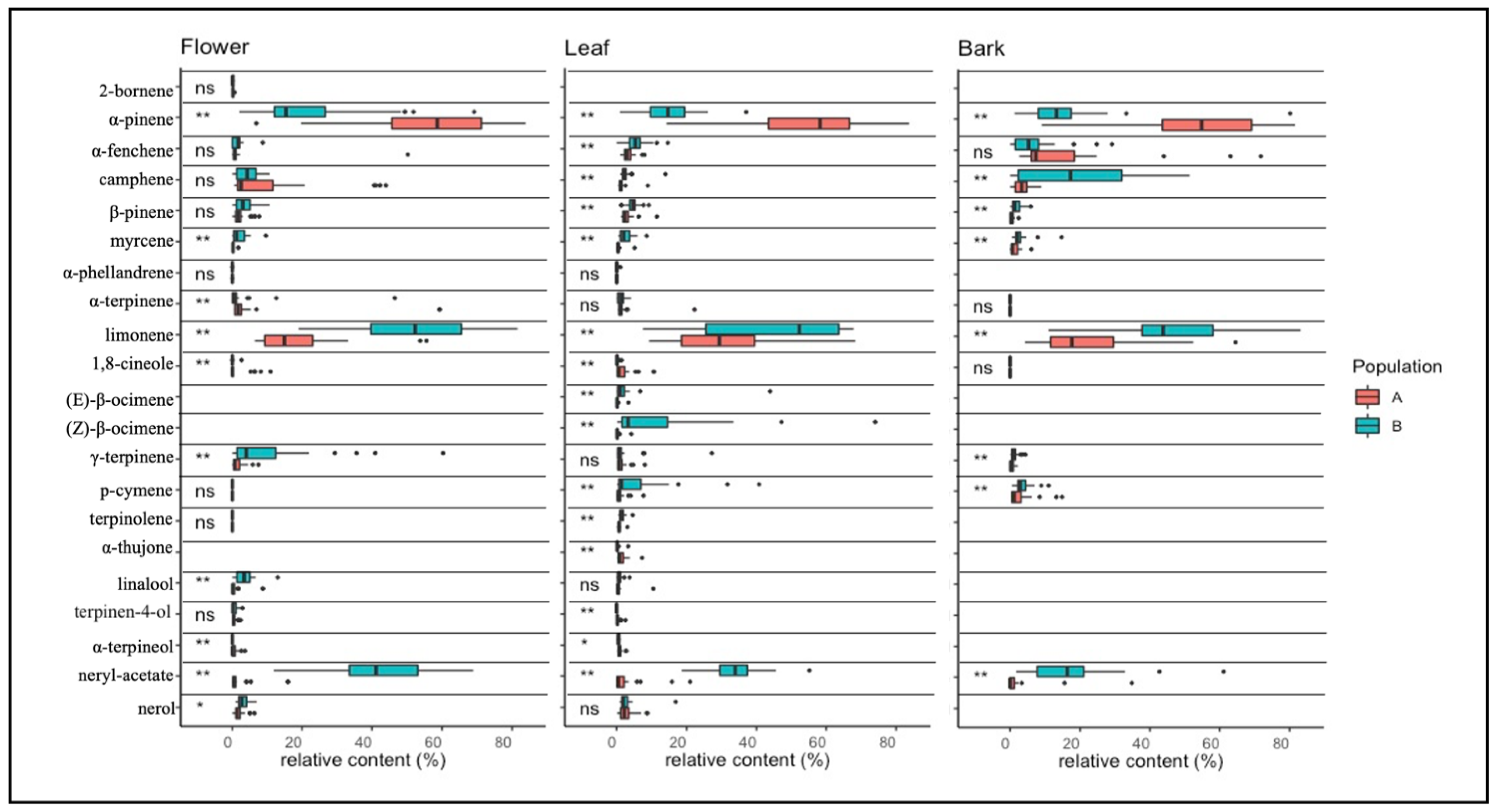
2.3.1. Monoterpenes
2.3.2. Sesquiterpenes
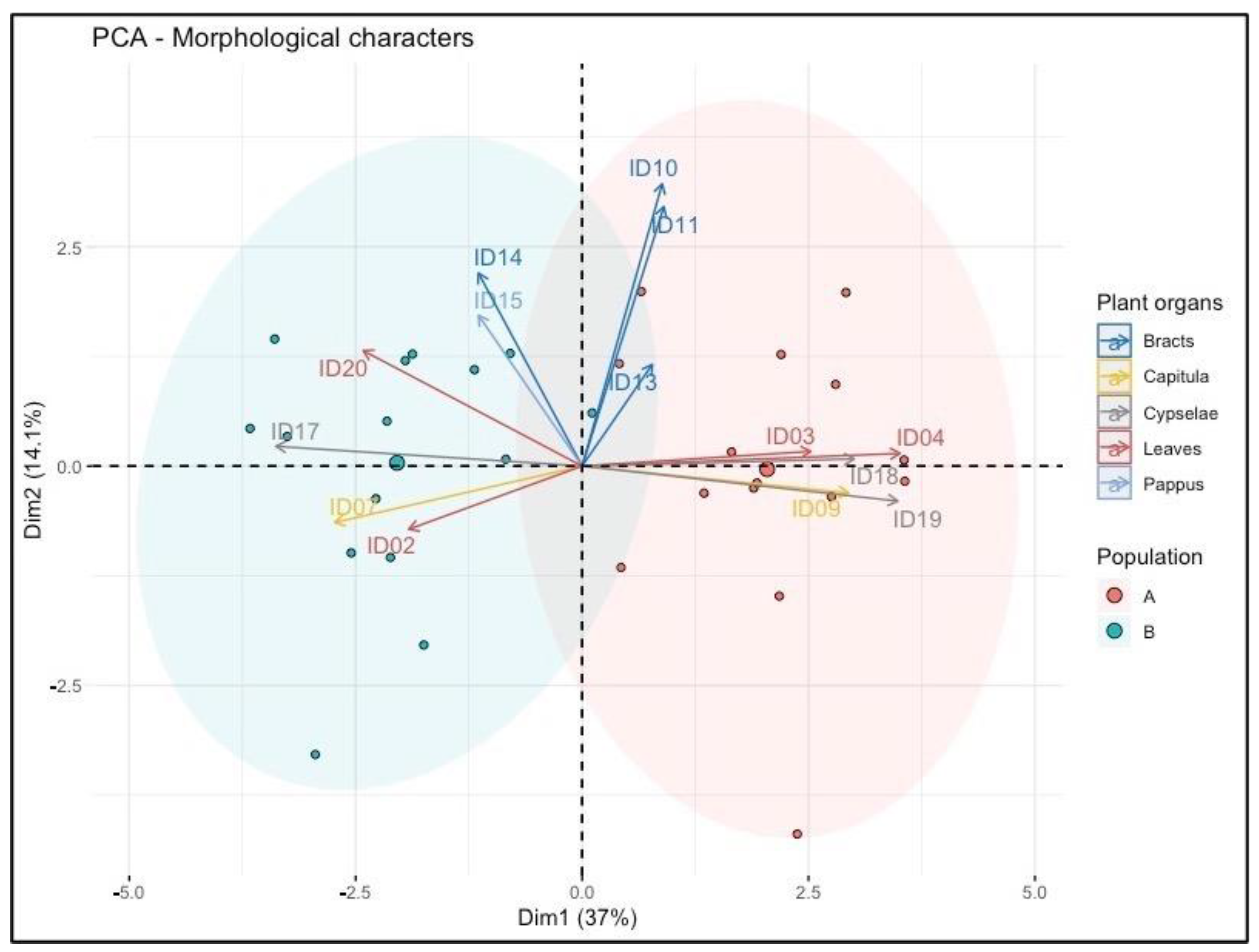
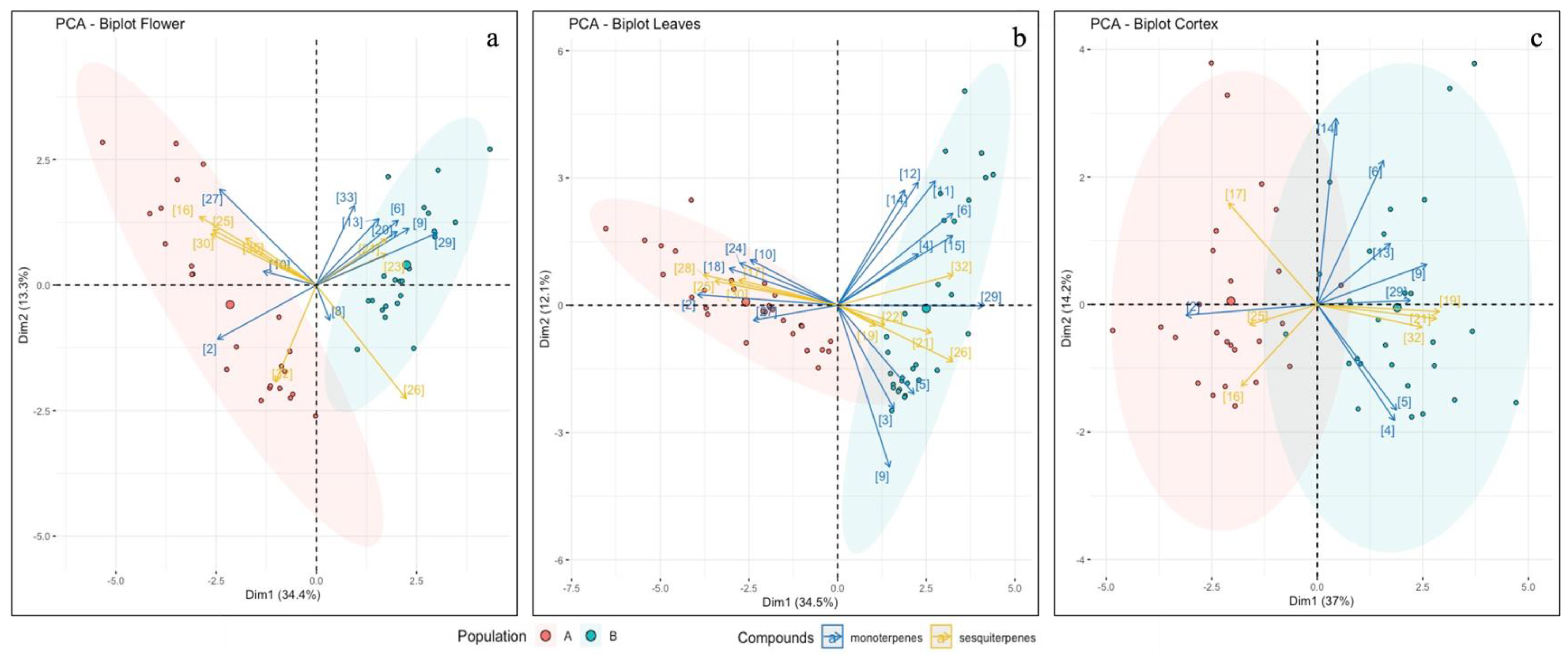
2.3.3. Multivariate Analysis of Terpene Data
3. Discussion
3.1. Morphological Analysis
3.2. Morphological Analysis
3.3. Terpene Analysis
4. Materials and Methods
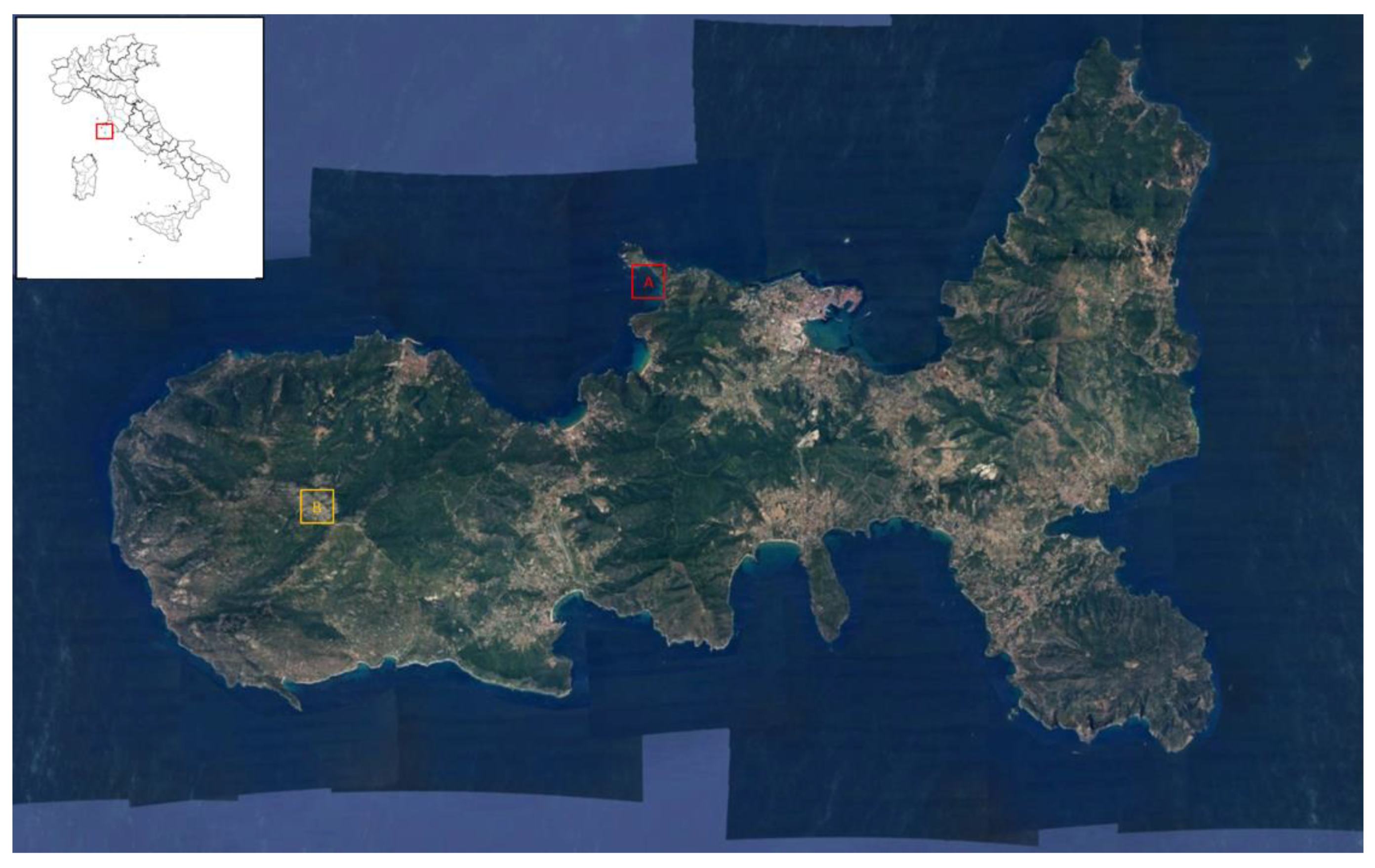
4.1. Study Area
4.2. Plant Materials
4.3. Morphological Analysis
4.4. Micromorphological Analysis
4.5. Terpene Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Galbany-Casals, M.; Garcia-Jacas, N.; Sáez, L.; Benedí, C.; Susanna, A. Phylogeny, Biogeography, and Character Evolution in Mediterranean, Asiatic, and Macaronesian Helichrysum (Asteraceae, Gnaphalieae) Inferred from Nuclear Phylogenetic Analyses. Int. J. Plant Sci. 2009, 170, 365–380. [Google Scholar] [CrossRef]
- Guinoiseau, E.; Lorenzi, V.; Luciani, A.; Muselli, A.; Costa, J.; Casanova, J.; Berti, L. Biological Properties and Resistance Reversal Effect of Helichrysum italicum (Roth) G. Don. Microb. Pathog. Strateg. Combat. Sci. Technol. Educ. 2013, 2, 1073–1080. [Google Scholar]
- Morone-Fortunato, I.; Montemurro, C.; Ruta, C.; Perrini, R.; Sabetta, W.; Blanco, A.; Lorusso, E.; Avato, P. Essential Oils, Genetic Relationships and in Vitro Establishment of Helichrysum italicum (Roth) G. Don Ssp. Italicum from Wild Mediterranean Germplasm. Ind. Crops Prod. 2010, 32, 639–649. [Google Scholar] [CrossRef]
- Galbany-Casals, M.; Sáez, L.; Benedí, C. A Taxonomic Revision of Helichrysum Sect. Stoechadina (Asteraceae, Gnaphalieae). Can. J. Bot. 2006, 84, 1203–1232. [Google Scholar] [CrossRef]
- Herrando-Moraira, S.; Blanco-Moreno, J.M.; Sáez, L.; Galbany-Casals, M. Re-Evaluation of the Helichrysum italicum Complex (Compositae: Gnaphalieae): A New Species from Majorca (Balearic Islands). Collect. Bot. 2016, 35, 5. [Google Scholar] [CrossRef][Green Version]
- Pignatti, S.; Guarino, R.; la Rosa, M. Helichrysum Mill.-Perpetuini. In Flora d’Italia; Edagricole: Milano, Italy, 2017; pp. 782–784. [Google Scholar]
- Ascensao, L.; da Silva, J.A.T.; Barroso, J.G.; Figueiredo, A.C.; Pedro, L. Glandular Trichomes and Essential Oils of Helichrysum stoechas. Isr. J. Plant Sci. 2001, 49, 115–122. [Google Scholar] [CrossRef]
- Angioni, A.; Barra, A.; Arlorio, M.; Coisson, J.D.; Russo, M.T.; Pirisi, F.M.; Satta, M.; Cabras, P. Chemical Composition, Plant Genetic Differences, and Antifungal Activity of the Essential Oil of Helichrysum italicum G. Don Ssp. Microphyllum (Willd) Nym. J. Agric. Food Chem. 2003, 51, 1030–1034. [Google Scholar] [CrossRef]
- Mastelic, J.; Politeo, O.; Jerkovic, I.; Radosevic, N. Composition and Antimicrobial Activity of Helichrysum italicum Essential Oil and Its Terpene and Terpenoid Fractions. Chem. Nat. Compd. 2005, 41, 35–40. [Google Scholar] [CrossRef]
- Kladar, N.V.; Anačkov, G.T.; Rat, M.M.; Srd Strok Signenovic, B.U.; Grujic, N.N.; Šefer, E.I.; Božin, B.N. Biochemical Characterization of Helichrysum italicum (Roth) G.Don Subsp. italicum (Asteraceae) from Montenegro: Phytochemical Screening, Chemotaxonomy, and Antioxidant Properties. Chem. Biodivers. 2015, 12, 419–431. [Google Scholar] [CrossRef]
- Djihane, B.; Wafa, N.; Elkhamssa, S.; Pedro, D.H.J.; Maria, A.E.; Mohamed Mihoub, Z. Chemical Constituents of Helichrysum italicum (Roth) G. Don Essential Oil and Their Antimicrobial Activity against Gram-Positive and Gram-Negative Bacteria, Filamentous Fungi and Candida Albicans. Saudi Pharm. J. 2017, 25, 780–787. [Google Scholar] [CrossRef]
- Tzanova, M.; Grozeva, N.; Gerdzhikova, M.; Atanasov, V.; Terzieva, S.; Prodanova, R. Biochemical Composition of Essential Oil of Corsican Helichrysum italicum (Roth) G. Don, Introduced and Cultivated in South Bulgaria. Bulg. J. Agric. Sci. 2018, 24, 1071–1077. [Google Scholar]
- Dzamic, A.M.; Mileski, K.S.; Ciric, A.D.; Ristic, M.S.; Sokovic, M.D.; Marin, P.D. Essential Oil Composition, Antioxidant and Antimicrobial Properties of Essential Oil and Deodorized Extracts of Helichrysum italicum (Roth) G. Don. J. Essent. Oil-Bear. Plants 2019, 22, 493–503. [Google Scholar] [CrossRef]
- Ninčević, T.; Grdiša, M.; Šatović, Z.; Jug-Dujaković, M. Helichrysum italicum (Roth) G. Don: Taxonomy, Biological Activity, Biochemical and Genetic Diversity. Ind. Crops Prod. 2019, 138, 111487. [Google Scholar] [CrossRef]
- Motti, R. Wild Plants Used as Herbs and Spices in Italy: An Ethnobotanical Review. Plants 2021, 10, 563. [Google Scholar] [CrossRef] [PubMed]
- Galbany-Casals, M.; Garcia-Jacas, N.; Susanna, A.; Sáez, L.; Benedí, C. Phylogenetic Relationships in the Mediterranean Helichrysum (Asteraceae, Gnaphalieae) Based on Nuclear RDNA ITS Sequence Data. Aust. Syst. Bot. 2004, 17, 241–253. [Google Scholar] [CrossRef]
- Aghababyan, M.; Greuter, W.; Mazzola, P.; Raimondo, F.M. On the Taxonomy and Nomenclature of Gnaphalium angustifolium Lam. and Helichrysum litoreum Guss. (Compositae). Bocconea 2009, 23, 157–163. [Google Scholar]
- Arbeiter, A.B.; Hladnik, M.; Jakše, J.; Bandelj, D. First Set of Microsatellite Markers for Immortelle (Helichrysum italicum (Roth) G. Don): A Step towards the Selection of the Most Promising Genotypes for Cultivation. Ind. Crops Prod. 2021, 162, 113298. [Google Scholar] [CrossRef]
- Hladnik, M.; Baruca Arbeiter, A.; Knap, T.; Jakše, J.; Bandelj, D. The Complete Chloroplast Genome of Helichrysum italicum (Roth) G. Don (Asteraceae). Mitochondrial DNA Part B Resour. 2019, 4, 1036–1037. [Google Scholar] [CrossRef]
- Singh, R. Chemotaxonomy: A Tool for Plant Classification. J. Med. Plants Stud. 2016, 4, 90–93. [Google Scholar]
- Otto, A.; Wilde, V. Sesqui-, Di-, and Triterpenoids as Chemosystematic Markers in Extant Conifers—A Review. Bot. Rev. 2001, 67, 141–238. [Google Scholar] [CrossRef]
- Silvestrini, E.; Michelozzi, M.; Skroppa, T.; Brancaleoni, E.; Ciccioli, P. Characterisation of Different Clones of Picea abies (L.) Karst Using Head-Space Sampling of Cortical Tissues Combined with Enantioselective Capillary Gas Chromatography for the Separation of Chiral and Non-Chiral Monoterpenes. J. Chromatogr. A 2004, 1034, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Shulha, O.; Zidorn, C. Sesquiterpene Lactones and Their Precursors as Chemosystematic Markers in the Tribe Cichorieae of the Asteraceae Revisited: An Update (2008–2017). Phytochemistry 2019, 163, 149–177. [Google Scholar] [CrossRef] [PubMed]
- Michelozzi, M.; Tognetti, R.; Maggino, F.; Radicati, M. Seasonal Variations in Monoterpene Profiles and Ecophysiological Traits in Mediterranean Pine Species of Group “Halepensis”. iForest-Biogeosciences For. 2008, 1, 65–74. [Google Scholar] [CrossRef]
- Guitton, Y.; Nicolè, F.; Jullien, F.; Caissard, J.C.; Saint-Marcoux, D.; Legendre, L.; Pasquier, B.; Moja, S. A Comparative Study of Terpene Composition in Different Clades of the Genus Lavandula. Bot. Lett. 2018, 165, 494–505. [Google Scholar] [CrossRef]
- Reimann-Philipp, U.; Speck, M.; Orser, C.; Johnson, S.; Hilyard, A.; Turner, H.; Stokes, A.J.; Small-Howard, A.L. Cannabis Chemovar Nomenclature Misrepresents Chemical and Genetic Diversity; Survey of Variations in Chemical Profiles and Genetic Markers in Nevada Medical Cannabis Samples. Cannabis Cannabinoid Res. 2020, 5, 215–230. [Google Scholar] [CrossRef]
- Squillace, A.E. Analyses of Monoterpenes of Conifers by Gas-Liquid Chromatography. In Modern Methods in Forest Genetics; Springer: Berlin/Heidelberg, Germany, 1976; pp. 120–157. [Google Scholar]
- Tsoukatou, M.; Roussis, V.; Chinou, L.; Petrakis, P.V.; Ortiz, A. Chemical Composition of the Essential Oils and Headspace Samples of Two Helichrysum Species Occurring in Spain. J. Essent. Oil Res. 1999, 11, 511–516. [Google Scholar] [CrossRef]
- Leonardi, M.; Ambryszewska, K.E.; Melai, B.; Flamini, G.; Cioni, P.L.; Parri, F.; Pistelli, L. Essential-Oil Composition of Helichrysum italicum (Roth) G. Don Ssp. italicum from Elba Island (Tuscany, Italy). Chem. Biodivers. 2013, 10, 343–355. [Google Scholar] [CrossRef]
- Mashigo, M.; Combrinck, S.; Regnier, T.; du Plooy, W.; Augustyn, W.; Mokgalaka, N. Chemical Variations, Trichome Structure and Antifungal Activities of Essential Oils of Helichrysum splendidum from South Africa. S. Afr. J. Bot. 2015, 96, 78–84. [Google Scholar] [CrossRef]
- Maggio, A.; Bruno, M.; Guarino, R.; Senatore, F.; Ilardi, V. Contribution to a Taxonomic Revision of the Sicilian Helichrysum Taxa by PCA Analysis of Their Essential-Oil Compositions. Chem. Biodivers. 2016, 13, 151–159. [Google Scholar] [CrossRef]
- Maksimovic, S.; Tadic, V.; Skala, D.; Zizovic, I. Phytochemistry Separation of Phytochemicals from Helichrysum italicum: An Analysis of Different Isolation Techniques and Biological Activity of Prepared Extracts. Phytochemistry 2017, 138, 9–28. [Google Scholar] [CrossRef]
- Giuliani, C.; Lazzaro, L.; Calamassi, R.; Calamai, L.; Romoli, R.; Fico, G.; Foggi, B.; Mariotti Lippi, M. A Volatolomic Approach for Studying Plant Variability: The Case of Selected Helichrysum Species (Asteraceae). Phytochemistry 2016, 130, 128–143. [Google Scholar] [CrossRef] [PubMed]
- Najar, B.; Cervelli, C.; Ferri, B.; Cioni, P.L.; Pistelli, L. Essential Oils and Volatile Emission of Eight South African Species of Helichrysum Grown in Uniform Environmental Conditions. S. Afr. J. Bot. 2019, 124, 178–187. [Google Scholar] [CrossRef]
- Salmeri, C.; Brullo, S.; L’Episcopo, L. Variabilità Morfologica e Ultrastrutturale Delle Cipsele Delle Specie Siciliane Di Helichrysum (Asteraceae). SBI Gruppo Per La Florist. Sist. Ed Evol. 2014, 1, 35–36. [Google Scholar]
- Foggi, B.; Cartei, L.; Pignotti, L.; Signorini, M.A.; Viciani, D.; Dell’Olmo, L.; Menicagli, E. The Plant Landscape of the Elba Island (Tuscan Archipelago). Phytosociological and Carthographic Study. Fitosociologia 2006, 43, 3–95. [Google Scholar]
- Innamorati, T.F. La Flora Vascolare Dell’Isola d’Elba (Arcipelago Toscano). Parte Seconda. Webbia 1989, 43, 201–267. [Google Scholar] [CrossRef]
- Glennon, K.L.; Cron, G. v Climate and Leaf Shape Relationships in Four Helichrysum Species from the Eastern Mountain Region of South Africa. Evol. Ecol. 2015, 29, 657–678. [Google Scholar] [CrossRef]
- Galbany-Casals, M.; Blanco-Moreno, J.M.; Garcia-Jacas, N.; Breitwieser, I.; Smissen, R.D. Genetic Variation in Mediterranean Helichrysum italicum (Asteraceae; Gnaphalieae): Do Disjunct Populations of Subsp. microphyllum Have a Common Origin? Plant Biol. 2011, 13, 678–687. [Google Scholar] [CrossRef]
- Herrando-Moraira, S.; Carnicero, P.; Blanco-Moreno, J.M.; Sáez, L.; Véla, E.; Vilatersana, R.; Galbany-Casals, M. Systematics and Phylogeography of the Mediterranean Helichrysum pendulum Complex (Compositae) Inferred from Nuclear and Chloroplast DNA and Morphometric Analyses. Taxon 2017, 66, 909–933. [Google Scholar] [CrossRef]
- Azizi, N.; Sheidai, M.; Mozaffarian, V.; Arman, M.; Noormohammadi, Z. Assessment of Relationships among and within Helichrysum Mill. (Asteraceae) Species by Using Issr Markers and Morphological Traits. Hacquetia 2019, 18, 105–118. [Google Scholar] [CrossRef]
- Costa, P.; Loureiro, J.M.; Teixeira, M.A.; Rodrigues, A.E. Extraction of Aromatic Volatiles by Hydrodistillation and Supercritical Fluid Extraction with CO2 from Helichrysum italicum Subsp. picardii Growing in Portugal. Ind. Crops Prod. 2015, 77, 680–683. [Google Scholar] [CrossRef]
- Carta, A.; Forbicioni, L.; Frangini, G.; Pierini, B.; Peruzzi, L. Supplementar Data: An Updated Inventory of the Vascular Flora of Elba Island (Tuscan Archipelago, Italy). Ital. Bot. 2018, 6, 1. [Google Scholar] [CrossRef]
- Ruberto, G.; Biondi, D.M.; Barbagallo, C.; Meli, R.; Savoca, F. Constituents of Stem and Flower Oils of Helichrysum litoreum Guss. Flavour Fragr. J. 2002, 17, 46–48. [Google Scholar] [CrossRef]
- Paolini, J.; Desjobert, J.M.; Costa, J.; Bernardini, A.F.; Castellini, C.B.; Cioni, P.L.; Flamini, G.; Morelli, I. Composition of Essential Oils of Helichysum italicum (Roth) G. Don Fil Subsp. italicum from Tuscan Archipelago Islands. Flavour Fragr. J. 2006, 21, 805–808. [Google Scholar] [CrossRef]
- Usai, M.; Foddai, M.; Bernardini, A.F.; Muselli, A.; Costa, J.; Marchetti, M. Chemical Composition and Variation of the Essential Oil of Wild Sardinian Helichrysum italicum G. Don Subsp. microphyllum (Willd.) Nym from Vegetative Period to Post-Blooming. J. Essent. Oil Res. 2010, 22, 373–380. [Google Scholar] [CrossRef]
- Bertoli, A.; Conti, B.; Mazzoni, V.; Meini, L.; Pistelli, L. Volatile Chemical Composition and Bioactivity of Six Essential Oils against the Stored Food Insect Sitophilus zeamais Motsch. (Coleoptera Dryophthoridae). Nat. Prod. Res. 2012, 26, 2063–2071. [Google Scholar] [CrossRef]
- Marongiu, B.; Piras, A.; Desogus, E.; Porcedda, S.; Ballero, M. Analysis of the Volatile Concentrate of the Leaves and Flowers of Helichrysum italicum (Roth) Don Ssp. microphyllum (Willd.) Nyman (Asteraceae) by Supercritical Fluid Extraction and Their Essential Oils. J. Essent. Oil Res. 2003, 15, 120–126. [Google Scholar] [CrossRef]
- Melito, S.; Petretto, G.L.; Podani, J.; Foddai, M.; Maldini, M.; Chessa, M.; Pintore, G. Altitude and Climate Influence Helichrysum italicum Subsp. microphyllum Essential Oils Composition. Ind. Crops Prod. 2016, 80, 242–250. [Google Scholar] [CrossRef]
- Andreani, S.; Uehara, A.; Blagojević, P.; Radulović, N.; Muselli, A.; Baldovini, N. Key Odorants of Industrially-Produced Helichrysum italicum Subsp. italicum Essential Oil. Ind. Crops Prod. 2019, 132, 275–282. [Google Scholar] [CrossRef]
- Schipilliti, L.; Bonaccorsi, I.L.; Ragusa, S.; Cotroneo, A.; Dugo, P. Helichrysum italicum (Roth) G. Don Fil. Subsp. italicum Oil Analysis by Gas Chromatography-Carbon Isotope Ratio Mass Spectrometry (GC-C-IRMS): A Rapid Method of Genotype Differentiation? J. Essent. Oil Res. 2016, 28, 193–201. [Google Scholar] [CrossRef]
- Roussis, V.; Tsoukatou, M.; Petrakis, P.V.; Chinou, I.; Skoula, M.; Harborne, J.B. Volatile Constituents of Four Helichrysum Species Growing in Greece. Biochem. Syst. Ecol. 2000, 28, 163–175. [Google Scholar] [CrossRef]
- Mastelič, J.; Politeo, O.; Jerkovič, I. Contribution to the Analysis of the Essential Oil of Helichrysum italicum (Roth) G. Don.-Determination of Ester Bonded Acids and Phenols. Molecules 2008, 13, 795–803. [Google Scholar] [CrossRef] [PubMed]
- Politeo, O.; Jukić, M.; Miloš, M. Chemical Composition and Antioxidant Activity of Essential Oils of Twelve Spice Plants. Croat. Chem. Acta 2006, 79, 545–552. [Google Scholar]
- Talić, S.; Odak, I.; Bevanda, A.M.; Crnjac, N.; Paštar, M. Helichrysum italicum (Roth) G. Don Subsp. italicum from Herzegovina: Volatile Composition, Variations during Seasons, Total Polyphenols, Acetylcholinesterase Inhibition and Antioxidant Activity. Croat. Chem. Acta 2019, 92, 69–77. [Google Scholar] [CrossRef]
- Schaefer, P.R.; Hanover, J.W. Taxonomic Implications of Monoterpene Compounds of Blue and Engelmann Spruces. For. Sci. 1986, 32, 725–734. [Google Scholar] [CrossRef]
- Zeneli, G.; Tsitsimpikou, C.; Petrakis, P.V.; Naxakis, G.; Habili, D.; Roussis, V. Foliar and Cortex Oleoresin Variability of Silver Fir (Abies alba Mill.) in Albania. Z. Fur Nat.-Sect. C J. Biosci. 2001, 56, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Bianchini, A.; Tomi, P.; Bernardini, A.F.; Morelli, I.; Flamini, G.; Cioni, P.L.; Usaï, M.; Marchetti, M. A Comparative Study of Volatile Constituents of Two Helichrysum italicum (Roth) Guss. Don Fil Subspecies Growing in Corsica (France), Tuscany and Sardinia (Italy). Flavour Fragr. J. 2003, 18, 487–491. [Google Scholar] [CrossRef]
- Talić, S.; Odak, I.; Lukic, T.; Brkljaca, M.; Martinović Bevanda, A.; Lasic, A. Chemodiversity Oh Helichrysum italicum (Roth) G. Don Subsp. italicum Essential Oil from Bosnia and Herzegovina. Fresenius Environ. Bull. 2021, 30, 2492–2502. [Google Scholar]
- Derita, M.; Zacchino, S. Chemotaxonomic Importance of Sesquiterpenes and Flavonoids in Five Argentinian Species of Polygonum Genus. J. Essent. Oil Res. 2011, 23, 11–14. [Google Scholar] [CrossRef]
- Kubelka, W.; Kastner, U.; Glasl, S.; Saukel, J.; Jurenitsch, J. Chemotaxonomic Relevance of Sesquiterpenes within the Achillea millefolium Group. Biochem. Syst. Ecol. 1999, 27, 437–444. [Google Scholar] [CrossRef]
- Perrini, R.; Morone-Fortunato, I.; Lorusso, E.; Avato, P. Glands, Essential Oils and in Vitro Establishment of Helichrysum italicum (Roth) G. Don Ssp. microphyllum (Willd.) Nyman. Ind. Crops Prod. 2009, 29, 395–403. [Google Scholar] [CrossRef]
- Bianchini, A.; Tomi, P.; Costa, J.; Bernardini, A.F. Composition Of Helichrysum italicum (Roth) G. Don Fil. Subsp. italicum Essential Oils from Corsica (France). Flavour Fragr. J. 2001, 16, 30–34. [Google Scholar] [CrossRef]
- Melito, S.; Sias, A.; Petretto, G.L.; Chessa, M.; Pintore, G.; Porceddu, A. Genetic and Metabolite Diversity of Sardinian Populations of Helichrysum italicum. PLoS ONE 2013, 8, e79043. [Google Scholar] [CrossRef] [PubMed]
- Meteoam.it. Servizio Meteorologico Aeronautica Militare. 2022. Available online: http://www.meteoam.it/dati_in_tempo_reale (accessed on 15 June 2021).
- Puglia, G.; Grimaldi, S.; Pavone, P.; Spampinato, G. Genetic and Morphological Variability Analysis Revealed a Complex Network in South-Eastern Sicilian Helichrysum Occurrences. Plant Biosyst. 2018, 152, 142–151. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development Environment for R 2021; RStudio, Inc.: Boston, MA, USA, 2016. [Google Scholar]
- Alboukadel, K.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. R Package Version. 1.0.5. 2017. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 23 January 2022).
- Lê, S.; Josse, J.; Rennes, A.; Husson, F. FactoMineR: An R Package for Multivariate Analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID. | Morphological Variables | A | B | p-Value | Sig. |
|---|---|---|---|---|---|
| 01 | Axillary leaf fascicles (vegetative stem) a | 1.82 ± 0.39 | 1.94 ± 0.23 | 0.85 | ns |
| 02 | Leaf margin undulation a | 1.08 ± 0.27 | 1.63 ± 0.19 | 0.00 | ** |
| 03 | Caulinar leaf length b | 21.53 ± 4.50 | 16.82 ± 4.51 | 0.00 | ** |
| 04 | Caulinar leaf width b | 0.91 ± 0.20 | 0.63 ± 0.15 | 0.00 | ** |
| 05 | Glandular indumentum of leaf abaxial side (vegetative stem) c | 12.24 ± 4.84 | 12.31 ± 7.44 | 0.77 | ns |
| 06 | Synflorescence length c | 23.73 ± 5.27 | 26.69 ± 7.40 | 0.99 | ns |
| 07 | Number of capitula per synflorescence c | 17.53 ± 6.55 | 33.04 ± 13.43 | 0.02 | * |
| 08 | Capitulum length c | 4.26 ± 0.51 | 4.16 ± 0.34 | 0.86 | ns |
| 09 | Capitulum width | 3.09 ± 0.24 | 2.68 ± 0.37 | 0.00 | ** |
| 10 | Outermost involucral bract lenght | 1.16 ± 0.07 | 0.97 ± 0.06 | 0.00 | ** |
| 11 | Outermost involucral bract width b | 0.57 ± 0.12 | 0.45 ± 0.16 | 0.03 | * |
| 12 | Innermost involucral bract length b | 3.66 ± 0.34 | 3.59 ± 0.71 | 0.52 | ns |
| 13 | Innermost involucral bract width | 0.64 ± 0.09 | 0.59 ± 0.09 | 0.01 | * |
| 14 | Glandular indumendum of Innermost involucral bract | 44.44 ± 9.11 | 27.43 ± 7.76 | 0.00 | ** |
| 15 | Pappus setae length b | 2.89 ± 0.30 | 3.07 ± 0.24 | 0.00 | ** |
| 16 | Shape of pappus apical cells a | 1.03 ± 0.16 | 1.16 ± 0.37 | 0.46 | ns |
| 17 | Cypsela duplex hair density c | 1.53 ± 0.56 | 4.42 ± 1.72 | 0.02 | * |
| 18 | Cypsela length | 0.73 ± 0.10 | 0.58 ± 0.09 | 0.00 | ** |
| 19 | Cypsela width b | 0.37 ± 0.07 | 0.23 ± 0.07 | 0.00 | ** |
| 20 | Glandular indumendum of leaf abaxial side (floral stem) c | 15.10 ± 5.81 | 26.81 ± 13.13 | 0.02 | * |
| 21 | Axillary leaf fascicles (floral stem) a | 1.35 ± 0.48 | 1.25 ± 0.44 | 0.40 | ns |
| 22 | Caulinar leaf length/caulinar leaf width c | 24.20 ± 5.25 | 27.44 ± 7.10 | 0.01 | * |
| 23 | Capitulum length/capitulum width b | 1.41 ± 0.22 | 1.57 ± 0.20 | 0.29 | ns |
| 24 | Outermost involucral bract length/ Outermost involucral bract width | 2.05 ± 0.49 | 2.51 ± 1.71 | 0.57 | ns |
| 25 | Innermost involucral bract length/ Innermost involucral bract width b | 5.80 ± 0.90 | 6.22 ± 1.28 | 0.07 | ns |
| 26 | Cypsela length/Cyspela width c | 2.00 ± 0.34 | 2.75 ± 0.38 | 0.02 | * |
| 27 | Synflorence length/ Number of capitula per synflorescence b | 1.47 ± 0.37 | 0.92 ± 0.16 | 0.00 | ** |
| ID. | Morphological Characters | Unit of Measure | References |
|---|---|---|---|
| 01 | Axillary leaf fascicles (vegetative stem) | 1 absence; 2 presence | [5,41] |
| 02 | Leaf margin undulation | 1 most without; 2 some leaves, 3 most leaves | [5] |
| 03 | Caulinar leaf length | mm | [4,5] |
| 04 | Caulinar leaf width | mm | [4,66] |
| 05 | Glandular indumentum of leaf abaxial side (floral stem) | n/0.64 mm2 | [4,5] |
| 06 | Synflorence length | mm | [5] |
| 07 | Number of capitula per synflorescence | mm | [4,5] |
| 08 | Capitulum length | mm | [4,5] |
| 09 | Capitulum width | mm | [4,5] |
| 10 | Outermost involucral bract lenght | mm | [4,66] |
| 11 | Outermost involucral bract width | mm | [4,66] |
| 12 | Innermost involucral bract length | mm | [40,66] |
| 13 | Innermost involucral bract width | mm | [40,66] |
| 14 | Glandular indumendum of Innermost involucral bract | n/0.8 mm2 | [40,66] |
| 15 | Pappus setae length | mm | [5] |
| 16 | Shape of pappus apical cells | 1 acute; 2 obtuse | [4] |
| 17 | Cypsela duplex hair density | n/0.04 mm2 | [4] |
| 18 | Cypsela length | mm | [5] |
| 19 | Cypsela width | mm | [5] |
| 20 | Glandular indumendum of leaf abaxial side (floral stem) | n/0.64 mm2 | This work |
| 21 | Axillary leaf fascicles (floral stem) | 1 absence; 2 presence | This work |
| 22 | Caulinar leaf length/caulinar leaf width | Ratio: 1/2 | [4,5] |
| 23 | Capitulum length/capitulum width | Ratio: 6/7 | [4,5] |
| 24 | Outermost involucral bract length/ Outermost involucral bract width | Ratio: 8/9 | [5] |
| 25 | Innermost involucral bract length/ Innermost involucral bract width | Ratio: 10/11 | [5] |
| 26 | Cypsela length/Cyspela width | Ratio: 15/17 | [5] |
| 27 | Synflorence length/ Number of capitula per synflorescence | Ratio: 4/5 | This work |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marini, L.; Palchetti, E.; Brilli, L.; Fico, G.; Giuliani, C.; Michelozzi, M.; Cencetti, G.; Foggi, B.; Bruschi, P. Terpene Profiles Composition and Micromorphological Analysis on Two Wild Populations of Helichrysum spp. from the Tuscan Archipelago (Central Italy). Plants 2022, 11, 1719. https://doi.org/10.3390/plants11131719
Marini L, Palchetti E, Brilli L, Fico G, Giuliani C, Michelozzi M, Cencetti G, Foggi B, Bruschi P. Terpene Profiles Composition and Micromorphological Analysis on Two Wild Populations of Helichrysum spp. from the Tuscan Archipelago (Central Italy). Plants. 2022; 11(13):1719. https://doi.org/10.3390/plants11131719
Chicago/Turabian StyleMarini, Lorenzo, Enrico Palchetti, Lorenzo Brilli, Gelsomina Fico, Claudia Giuliani, Marco Michelozzi, Gabriele Cencetti, Bruno Foggi, and Piero Bruschi. 2022. "Terpene Profiles Composition and Micromorphological Analysis on Two Wild Populations of Helichrysum spp. from the Tuscan Archipelago (Central Italy)" Plants 11, no. 13: 1719. https://doi.org/10.3390/plants11131719
APA StyleMarini, L., Palchetti, E., Brilli, L., Fico, G., Giuliani, C., Michelozzi, M., Cencetti, G., Foggi, B., & Bruschi, P. (2022). Terpene Profiles Composition and Micromorphological Analysis on Two Wild Populations of Helichrysum spp. from the Tuscan Archipelago (Central Italy). Plants, 11(13), 1719. https://doi.org/10.3390/plants11131719