
Calcium and Potassium Accumulation during the Growing Season in Cabernet Sauvignon and Merlot Grape Varieties




Abstract
:1. Introduction
2. Results
Ca and K Accumulation during Berry Development
3. Discussion
4. Materials and Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Campbell, J.; Sarkhosh, A.; Habibi, F.; Gajjar, P.; Ismail, A.; Tsolova, V.; El-Sharkawy, I. Evaluation of Biochemical Juice Attributes and Color-Related Traits in Muscadine Grape Population. Foods 2021, 10, 1101. [Google Scholar] [CrossRef]
- Blatt, M.R. Cellular signalling and volume control in stomatal movements of plants. Annu. Rev. Cell Dev. Biol. 2000, 16, 221–241. [Google Scholar] [CrossRef] [PubMed]
- Zörb, C.; Senbayram, M.; Peiter, E. Potassium in agriculture—Status and perspectives. J. Plant Physiol. 2014, 171, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Demarty, M.; Morvan, C.; Thellier, M. Calcium and cell wall. Plant Cell Environ. 2006, 7, 441–448. [Google Scholar] [CrossRef]
- Martins, V.; Garcia, A.; Alhinho, A.T.; Costa, P.; Lanceros-Méndez, S.; Costa, M.M.R.; Gerós, H. Vineyard calcium sprays induce changes in grape berry skin, firmness, cell wall composition and expression of cell wall-related genes. Plant Physiol. Biochem. 2020, 150, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Martins, V.; Unlubayir, M.; Teixeira, A.; Lanoue, A.; Gerós, H. Exogenous calcium delays grape berry maturation in the White cv. Loureiro while increasing fruit firmness and flavonol content. Front. Plant Sci. 2021, 12, 742887. [Google Scholar] [CrossRef]
- Cabanne, C.; Donèche, B. Calcium accumulation and redistribution during the development of grape berry. Vitis 2003, 42, 19–21. [Google Scholar]
- Sanders, D.; Brownlee, C.; Harper, J.F. Communicating with calcium. Plant Cell 1999, 11, 691–706. [Google Scholar] [CrossRef] [Green Version]
- Hocking, B.; Tyerman, S.D.; Burton, R.A.; Gilliham, M. Fruit Calcium: Transport and Physiology. Front. Plant Sci. 2016, 7, 1–17. [Google Scholar] [CrossRef] [Green Version]
- White, P.J. The pathways of calcium movement to the xylem. J. Exp. Bot. 2001, 52, 891–899. [Google Scholar] [CrossRef]
- Ciccarese, A.; Stellacci, A.M.; Gentilesco, G.; Rubino, P. Effectiveness of pre- and post-veraison calcium applications to control decay and maintain table grape fruit quality during storage. Postharvest Biol. Technol. 2013, 75, 135–141. [Google Scholar] [CrossRef]
- Hrazdina, G.; Parsons, G.F.; Mattick, L.R. Physiological and biochemical events during development and ripening of grape berries. Am. J. Enol. Vitic. 1984, 35, 220–227. [Google Scholar]
- Possner, D.R.E.; Kliewer, W.M. The localization of acids, sugars, potassium and calcium in developing grape berries. Vitis 1985, 24, 229–240. [Google Scholar] [CrossRef]
- Creasy, G.L.; Price, S.F.; Lombard, P.B. Evidence for xylem discontinuity in Pinot Noir and Merlot: Dye uptake and mineral composition during berry maturation. Am. J. Enol. Vitic. 1993, 44, 187–192. [Google Scholar]
- Schaller, K.; Löhnertz, O.; Chikkasubbanna, V. Uptake of potassium by the grape berries of different cultivars during growth and development. Vitic. Enol. Sci. 1992, 47, 36–39. [Google Scholar]
- Nunan, K.J.; Sims, I.M.; Bacic, A.; Robinson, S.P.; Fincher, G.B. Changes in cell wall composition during ripening of grape berries. Plant Physiol. 1998, 118, 783–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogiers, S.Y.; Keller, M.; Holzapfel, B.P.; Virgona, J.M. Accumulation of potassium and calcium by ripening berries on field vines of Vitis vinifera (L.) cv. Shiraz. Aust. J. Grape Wine Res. 2000, 6, 240–243. [Google Scholar] [CrossRef]
- Liu, G.T.; Wang, J.F.; Cramer, G.; Dai, Z.W.; Duan, W.; Xu, H.G.; Wu, B.H.; Fan, P.G.; Wang, L.J.; Li, S.H. Transcriptomic analysis of grape (Vitis vinifera L.) leaves during and after recovery from heat stress. BMC Plant Biol. 2012, 12, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.M.; Huang, H.B.; Wang, H.C. Cell walls of loosening skin in post-veraison grape berries lose structural polysaccharides and calcium while accumulate structural proteins. Sci. Hort. 2005, 104, 249–263. [Google Scholar] [CrossRef]
- Chardonnet, C.; Doneche, B. Relation between calcium content and resistance to enzymatic digestion of the skin during grape ripening. Vitis 1995, 34, 95–98. Available online: https://eurekamag.com/research/002/684/002684961.php (accessed on 1 April 2022).
- Cole, M.; Wiechel, T.; Warren, M.; Holmes, R. Ensuring Optimal Grape Quality through Management Strategies for Botrytis Cinerea; Final Report to “Grape and Wine Research & Development Corporation”; Project Number: MOU 01/02; Department of Primary Industries, Victoria Monash University: Clayton, Australia, 2004; pp. 1–72. [Google Scholar]
- Chardonnet, C.; L’Hyvernay, A.; Doneche, B. Effect of calcium treatment prior to Botrytis cinereainfection on the changes in pectic composition of grape berry. Physiol. Mol. Plant Pathol. 1997, 50, 213–218. [Google Scholar] [CrossRef]
- Venios, X.; Korkas, E.; Nisiotou, A.; Banilas, G. Grapevine responses to heat stress and global warming. Plants 2020, 9, 1754. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Broadley, M.R. Calcium in plants. Ann. Bot. 2003, 92, 487–511. [Google Scholar] [CrossRef] [PubMed]
- Ruhl, E.H. Effect of potassium and nitrogen supply on the distribution of minerals and organic acids and the composition of grape juice of Sultana vines. Aust. J. Exp. Agric. 1989, 29, 133–137. [Google Scholar] [CrossRef]
- Rogiers, S.Y.; Coetzee, Z.A.; Walker, R.R.; Deloire, A.; Tyerman, S.D. Potassium in the grape (Vitis vinifera L.) Berry: Transport and function. Front. Plant Sci. 2017, 8, 1629. [Google Scholar] [CrossRef] [PubMed]
- Blanke, M.M.; Leyhe, A. Stomatal activity of the grape berry cv. Riesling, Muller-Thurgau and Ehrenfelser. J. Plant Physiol. 1987, 127, 451–460. [Google Scholar] [CrossRef]
- Marcuzzo, P.; Gaiotti, F.; Lucchetta, M.; Lovat, L.; Tomasi, D. Tuning potassium fertilization to improve ph and acidity in Glera Grapevine (Vitis vinifera L.) under a Warming Climate. Appl. Sci. 2021, 11, 11869. [Google Scholar] [CrossRef]
- Davies, C.; Shin, R.; Liu, W.; Thomas, M.R.; Schachtman, D.P. Transporters expressed during grape berry (Vitis vinifera L.) development are associated with an increase in berry size and berry potassium accumulation. J. Exp. Bot. 2006, 57, 3209–3216. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The critical role of potassium in plant stress response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [Green Version]
- Monder, H.; Maillard, M.; Chérel, I.; Zimmermann, S.D.; Paris, N.; Cuéllar, T.; Gaillard, I. Adjustment of k+ fluxes and grapevine defence in the face of climate change. Int. J. Mol. Sci. 2021, 22, 10398. [Google Scholar] [CrossRef]
- Szczerba, M.W.; Britto, D.T.; Kronzucker, H.J. K+ transport in plants: Physiology and molecular biology. J. Plant Physiol. 2009, 166, 447–466. [Google Scholar] [CrossRef] [PubMed]
- Hunter, J.J.; Ruffner, H.P.; Volschenk, C.G. Starch concentrations in grapevine leaves, berries and roots and the effect of canopy management. S. Afr. J. Enol. Vitic. 1995, 16, 35–40. [Google Scholar] [CrossRef] [Green Version]
- Véry, A.A.; Nieves-Cordones, M.; Daly, M.; Khan, I.; Fizames, C.; Sentenac, H. Molecular biology of K+ transport across the plant cell membrane: What do we learn from comparison between plant species? J. Plant Physiol. 2014, 171, 748–769. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, A.E. Accumulation of Calcium by Ripening Berries on Grape (Vitis vinifera L. cv. Asgari) Permalink. The Proceedings of the International Plant Nutrition Colloquium XVI. 2009. Available online: https://escholarship.org/uc/item/8422d8x4 (accessed on 20 May 2022).
- Cohen, S.D.; Tarara, J.M.; Kennedy, J.A. Assessing the impact of temperature on grape phenolic metabolism. Anal. Chim. Acta 2008, 621, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Chardonnet, C. Calcium de la Baie de Raisin—Relation Entre la Cohésion des Parois Cellulaires et la Sensibilité à Botrytis Cinerea (Calcium of Grape Berry: Relation between Cell Wall Cohesion and Susceptibility to Botrytis Cinerea). Ph.D. Thesis, Université Victor Segalen (U.Bordeaux 2), Bordeaux, France, 1994. [Google Scholar]
- Cabanne, C.; Donèche, B. Changes in polygalacturonase activity and calcium content during ripening of grape berries. Am. J. Enol. Vitic. 2001, 52, 331–335. [Google Scholar]
- Kidman, C.M.; Dry, P.R.; Mccarthy, M.G.; Collins, C. Effect of rootstock on nutrition, pollination and fertilisation in ‘Shiraz’ (Vitis vinifera L.). Vitis 2014, 53, 139–145. [Google Scholar]
- Keller, M.; Shrestha, P.M.; Hall, G.E.; Bondada, B.R.; Davenport, J.R. Arrested sugar accumulation and altered organic acid metabolism in grape berries affected by berry shrivel syndrome. Am. J. Enol. Vitic. 2016, 67, 398–406. [Google Scholar] [CrossRef]
- Gilliham, M.; Dayod, M.; Hocking, B.J.; Xu, B.; Conn, S.J.; Kaiser, B.N.; Leigh, R.A.; Tyerman, S.D. Calcium delivery and storage in plant leaves: Exploring the link with water flow. J. Exp. Bot. 2011, 62, 2233–2250. [Google Scholar] [CrossRef]
- Keller, M.; Zhang, Y.; Shrestha, P.M.; Biondi, M.; Bondada, B.R. Sugar demand of ripening grape berries leads to recycling of surplus phloem water via the xylem. Plant Cell Environ. 2015, 38, 1048–1059. [Google Scholar] [CrossRef]
- Fuentes, S.; Sullivan, W.; Tilbrook, J.; Tyerman, S. A novel analysis of grapevine berry tissue demonstrates a variety-dependent correlation between tissue vitality and berry shrivel. Aust. J. Grape Wine Res. 2010, 16, 327–336. [Google Scholar] [CrossRef]
- Kurt, A.; Torun, H.; Colak, N.; Seiler, G.; Ahmet, F. Nutrient profiles of the hybrid grape cultivar “Isabel” during berry maturation and ripening. J. Sci. Food Agric. 2017, 97, 2468–2479. [Google Scholar] [CrossRef] [PubMed]
- Ollat, N.; Gaudillère, J.P. Investigation of assimilate import mechanisms in berries of Vitis vinifera var. Cabernet Sauvignon. Acta Hortic. 1996, 427, 141–149. [Google Scholar] [CrossRef]
- Schaller, K.; Löhnertz, O.; Chikkasubbanna, V. Calcium absorption by the grape berries of different cultivars during growth and development. Vitic. Enol. Sci. 1992, 47, 62–65. [Google Scholar]
- Konrad, K.R.; Wudick, M.M.; Feijo, J.A. Calcium regulation of tip growth: New genes for old mechanisms. Curr. Opin. Plant Biol. 2011, 14, 721–730. [Google Scholar] [CrossRef] [PubMed]
- Inanaga, S.; Ishimoto, Y.; Nishihara, T. Ca-binding compounds in cell wall of peanut shell. Soil Sci. Plant Nutr. 1988, 34, 319–326. [Google Scholar] [CrossRef]
- Etchebarne, F.; Ojeda, H.; Deloire, A. Influence of water status on mineral composition of berries in ‘Grenache Noir’ (Vitis vinifera L.). Vitis. 2009, 48, 63–68. [Google Scholar]
- Montanaro, G.; Dichio, B.; Lang, A.; Mininni, A.N.; Xiloyannis, C. Fruit calcium accumulation coupled and uncoupled from its transpiration in kiwifruit. J. Plant Physiol. 2015, 181, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Freeman, B.M.; Kliewer, W.M. Effect of irrigation, crop level and potassium fertilization on Carignane Vines. II. Grape and Wine Quality. Am. J. Enol. Vitic. 1983, 34, 197–207. [Google Scholar]
- Villette, J.; Cuéllar, T.; Verdeil, J.L.; Delrot, S.; Gaillard, I. Grapevine potassium nutrition and fruit quality in the context of climate change. Front. Plant Sci. 2020, 11, 123. [Google Scholar] [CrossRef] [Green Version]
- Klein, I.; Strime, M.; Fanberstein, L.; Mani, Y. Irrigation and fertigation effects on phosphorus and potassium nutrition of wine grapes. Vitis 2000, 39, 55–62. [Google Scholar]
- Rogiers, S.Y.; Greer, D.H.; Hatfield, J.M.; Orchard, B.A.; Keller, M. Mineral sinks within ripening grape berries (Vitis vinifera L.). Vitis 2006, 45, 115–123. [Google Scholar]
- Patrick, J.W.; Offler, C.E. Post-sieve element transport of photoassimilates in sink regions. J. Exp. Bot. 1996, 28, 736–743. [Google Scholar] [CrossRef] [PubMed]
- Fontes, N.; Côrte-Real, M.; Gerós, H. New observations on the integrity, structure, and physiology of flesh cells from fully ripened grape berry. Am. J. Enol. Vitic. 2011, 62, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Lang, A.; Thorpe, M.R. Xylem, phloem and transpiration flows in a grape—Application of a technique for measuring the volume of attached fruits to high-resolution using Archimedes principle. J. Exp. Bot. 1989, 40, 1069–1078. [Google Scholar] [CrossRef]
- Keller, M.; Shrestha, P.M. Solute accumulation differs in the vacuoles and apoplast of ripening grape berries. Planta 2014, 239, 633–642. [Google Scholar] [CrossRef]
- Boulton, R. The general relationship between potassium, sodium and pH in grape juice and wine. Am. J. Enol. Vitic. 1980, 31, 182–186. [Google Scholar]
- Mpelasoka, B.S.; Schachtman, D.P.; Treeby, M.; Thomas, M.R. Review of potassium nutrition in grapevines with special emphasis on berry accumulation. Aust. J. Grape Wine Res. 2008, 9, 154–168. [Google Scholar] [CrossRef]
- Rogiers, S.Y.; Smith, J.S.; White, R.; Keller, M.; Holzapfel, B.P.; Virgona, J.M. Vascular function in berries of Vitis vinifera (L) cv. Shiraz. Aust. J. Grape Wine Res. 2001, 7, 47–51. [Google Scholar] [CrossRef]
- Vitis International Variety Catalogue VIVC, Variety Number VIVC 11473, Species—Vitis Interspecific Crossing; Prime Name—Selektion Oppenheim 4. 2022. Available online: https://www.vivc.de/index.php?r=cultivarname%2Findex (accessed on 30 May 2022).
 —Ca skin CS;
—Ca skin CS;  —Ca skin M;
—Ca skin M;  —Ca flesh CS;
—Ca flesh CS;  —Ca flesh M).
—Ca skin CS; —Ca skin M; —Ca flesh CS; —Ca flesh M).
—Ca flesh M).
—Ca skin CS; —Ca skin M; —Ca flesh CS; —Ca flesh M).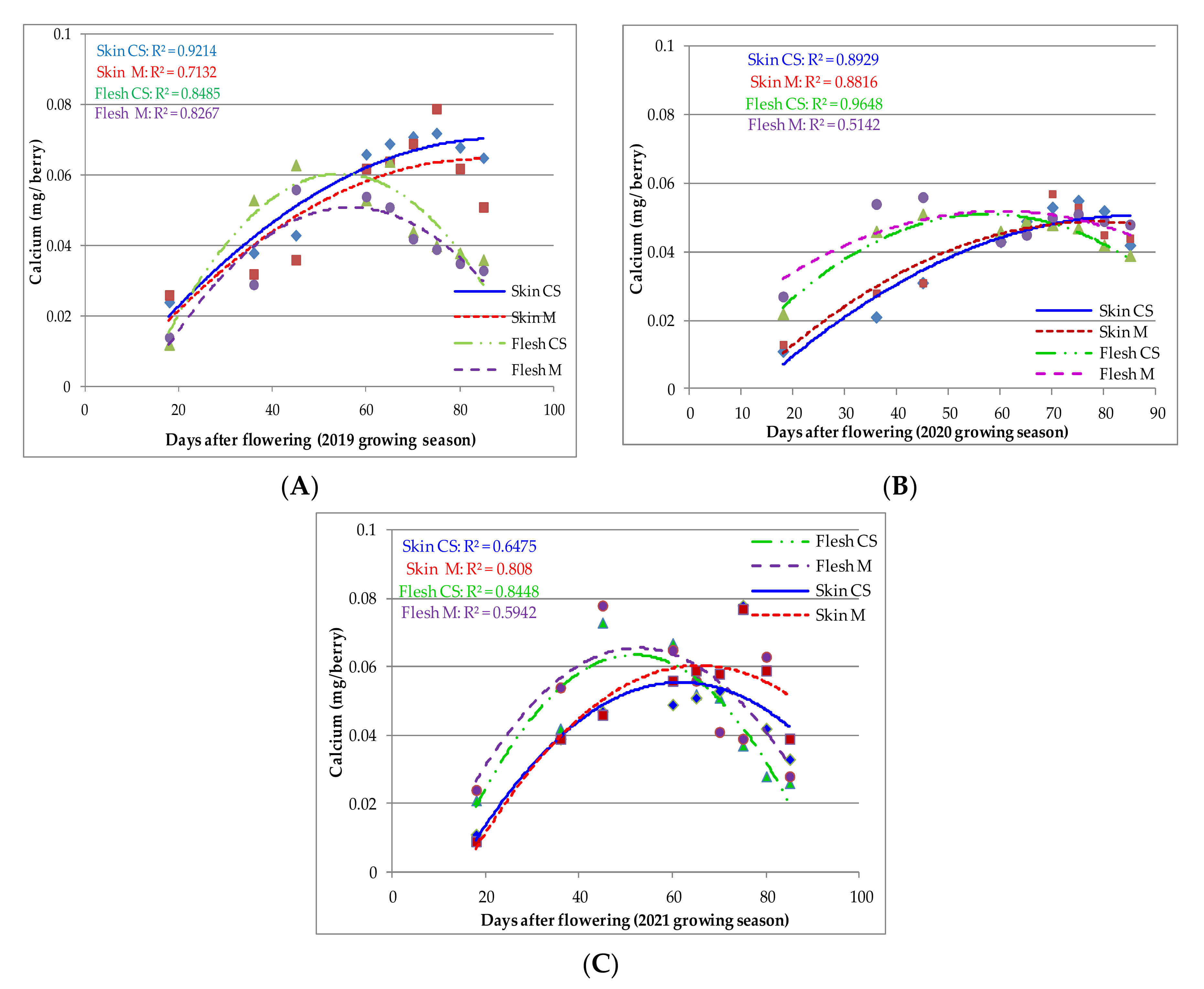 —K flesh CS; —K flesh M; —K skin CS; —K skin M).
—K flesh CS; —K flesh M; —K skin CS; —K skin M).
—K flesh CS; —K flesh M; —K skin CS; —K skin M).
—K flesh CS; —K flesh M; —K skin CS; —K skin M). —K in CS berry; —K in M berry; —Ca in CS berry; —Ca in M berry.
—K in CS berry; —K in M berry; —Ca in CS berry; —Ca in M berry.
—K in CS berry; —K in M berry; —Ca in CS berry; —Ca in M berry.
—K in CS berry; —K in M berry; —Ca in CS berry; —Ca in M berry.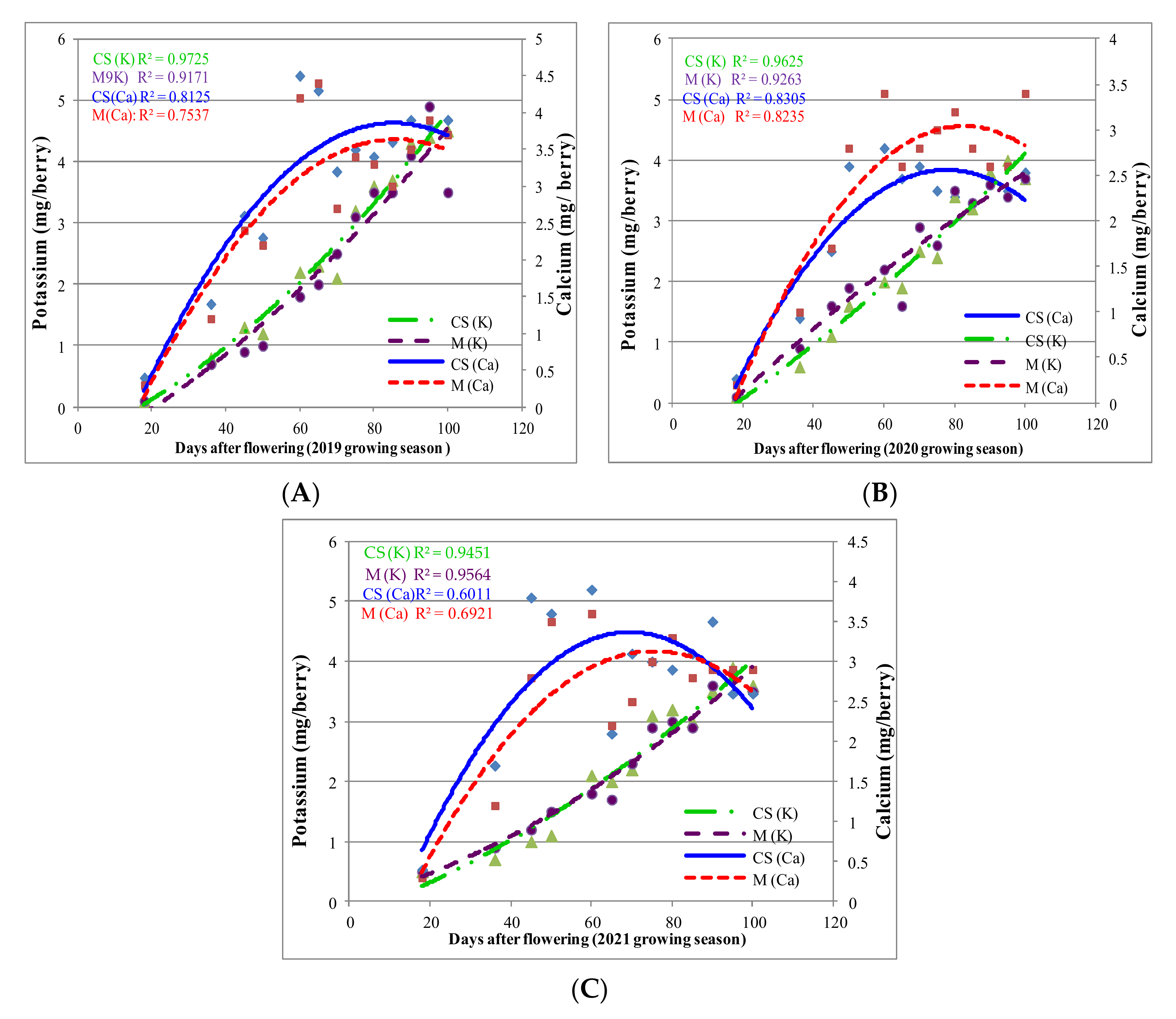 —K in CS seeds; —K in M seeds; —Ca in CS seeds; —Ca in M seeds).
—K in CS seeds; —K in M seeds; —Ca in CS seeds; —Ca in M seeds).
—K in CS seeds; —K in M seeds; —Ca in CS seeds; —Ca in M seeds).
—K in CS seeds; —K in M seeds; —Ca in CS seeds; —Ca in M seeds). —K in CS berry; —Ca in CS berry; B,D,F —K in M berry; —Ca in M berry;).
—K in CS berry; —Ca in CS berry; B,D,F —K in M berry; —Ca in M berry;).
—K in CS berry; —Ca in CS berry; B,D,F —K in M berry; —Ca in M berry;).
—K in CS berry; —Ca in CS berry; B,D,F —K in M berry; —Ca in M berry;).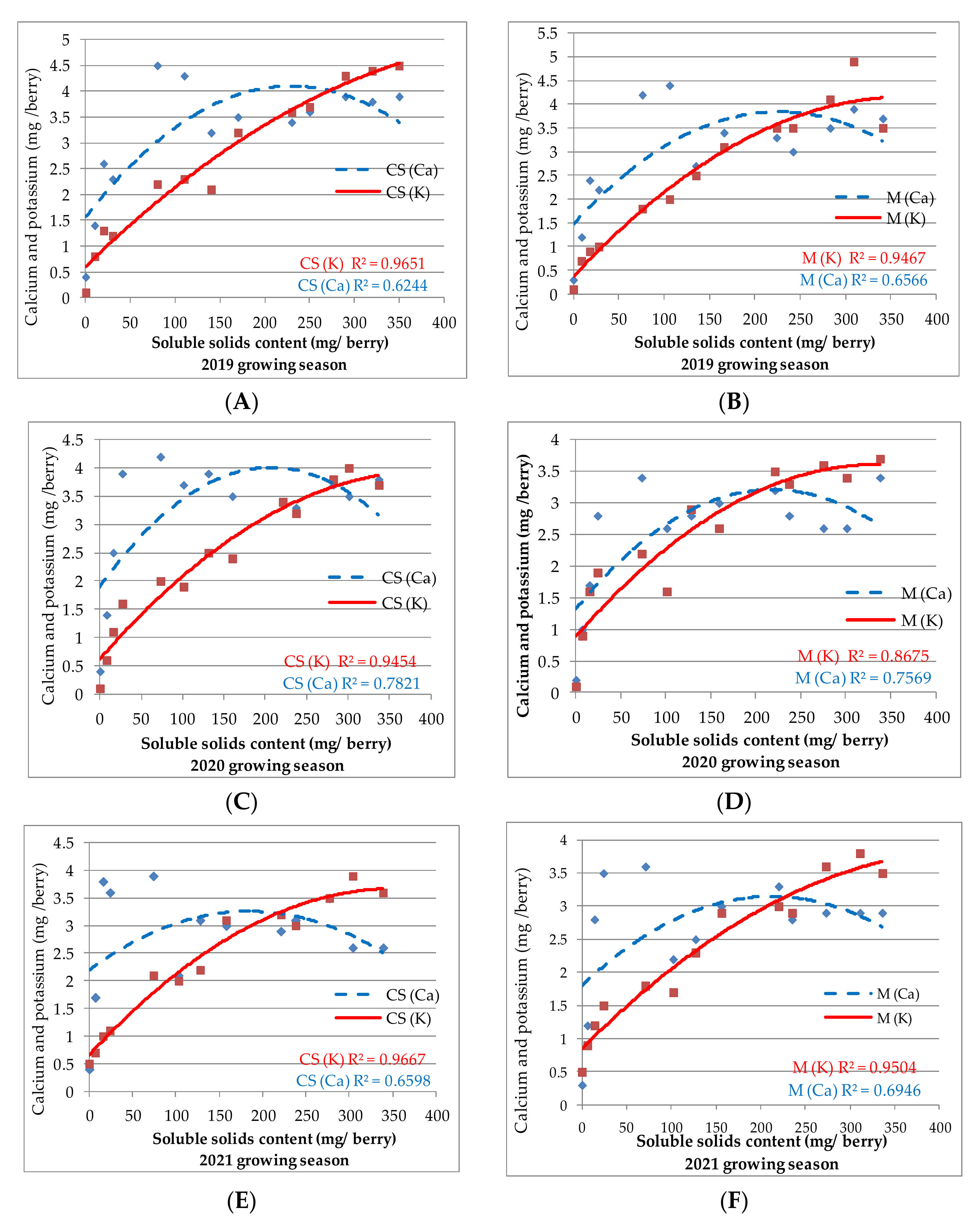

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2019 | 2020 | 2021 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Av. Max (°C) | Av. Min (°C) | Total Rainfall (mm) | Av. Max (°C) | Av. Min (°C) | Total Rainfall (mm) | Av. Max (°C) | Av. Min (°C) | Total Rainfall (mm) | |
| April | 18.30 | 7.63 | 112.10 | 16.33 | 6.40 | 22.30 | 13.17 | 5.33 | 80.70 |
| May | 19.30 | 9.67 | 259.30 | 19.29 | 10.19 | 61.80 | 19.13 | 10.39 | 97.80 |
| June | 27.52 | 16.28 | 154.70 | 23.60 | 15.33 | 196.30 | 26.90 | 15.57 | 16.70 |
| July | 27.32 | 15.38 | 113.80 | 26.16 | 16.68 | 149.20 | 32.77 | 19.29 | 95.20 |
| August | 31.13 | 17.90 | 56.20 | 29.58 | 18.64 | 56.50 | 31.00 | 17.94 | 59.00 |
| September | 25.33 | 13.20 | 54.20 | 26.10 | 15.30 | 43.00 | 26.07 | 13.30 | 3.20 |
| October | 20.94 | 15.82 | 27.80 | 17.42 | 10.74 | 117.70 | 16.52 | 7.65 | 10.07 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nistor, E.; Dobrei, A.G.; Mattii, G.B.; Dobrei, A. Calcium and Potassium Accumulation during the Growing Season in Cabernet Sauvignon and Merlot Grape Varieties. Plants 2022, 11, 1536. https://doi.org/10.3390/plants11121536
Nistor E, Dobrei AG, Mattii GB, Dobrei A. Calcium and Potassium Accumulation during the Growing Season in Cabernet Sauvignon and Merlot Grape Varieties. Plants. 2022; 11(12):1536. https://doi.org/10.3390/plants11121536
Chicago/Turabian StyleNistor, Eleonora, Alina Georgeta Dobrei, Giovan Battista Mattii, and Alin Dobrei. 2022. "Calcium and Potassium Accumulation during the Growing Season in Cabernet Sauvignon and Merlot Grape Varieties" Plants 11, no. 12: 1536. https://doi.org/10.3390/plants11121536
APA StyleNistor, E., Dobrei, A. G., Mattii, G. B., & Dobrei, A. (2022). Calcium and Potassium Accumulation during the Growing Season in Cabernet Sauvignon and Merlot Grape Varieties. Plants, 11(12), 1536. https://doi.org/10.3390/plants11121536