1. Introduction
Global agriculture is unable to cope with the existing climate change scenario and to feed the worlds growing population that is projected to increase from 6.7 billion (2005) to 9.2 billion by 2050 [
1]. Among all these anthropogenic factors, drought, salinity, and climate change are the principal players behind the land degradation and desertification leading to a significant reduction in crop production and yield decline [
2,
3,
4,
5]. Due to the scarce water resources and drought episodes, the irrigation water requirement in Arabian Gulf countries is mostly fulfilled through salty ground water and treated wastewater that is recruited to irrigate a significant land area (forestry, landscaping, roadside plantation) [
5]. To meet the growing need of agriculture, date palm fruit gardens and landscaping, the Gulf countries are using desalinated water (7.2%) and groundwater (91%) to meet their requirements [
6]. In this context, appropriate crop accessions that can be well adapted to the marginalized lands and available non-conventional water resources are suitable options for long-term rehabilitation and desertification resistance [
3,
7,
8,
9,
10].
Barley is an important grain crop and ranked fourth among the cereal crops after wheat, rice, and maize [
11]. It is mainly used as food, animal fodder, and as a raw material for beer production [
12]. Several authors have demonstrated that barley can tolerate a number of environmental stresses, such as drought [
13,
14], salinity [
15], and heavy metals [
16]. However, salt tolerance within genotypes of barley under field conditions has not been evaluated intensively. Therefore, the study of genetic diversity and phenotypic plasticity should be integrated in order to evaluate and select the most tolerant genotypes within a wide range of salinity among this plant species. Furthermore, the growth, yield, and productivity of barley are highly variable in the Middle East and North Africa region because the local cultivars do not have sufficient tolerance potential against prevailing environmental constraints, especially drought and salinity. Most researchers have evaluated the variation in salinity tolerance using growth chamber or green house at a single level of salinity and there was no validation of those results under the field setting. Meanwhile, studies conducted in a controlled growth chamber generally involve the determination of salinity stress on seeding growth over a short period of time (often 1–7 days), which does not correspond to salt stress in the field that might indicate a wide variation in the growth, development, physiological, and yield traits [
17].
It has been observed that among the population of particular crop genotypes, wide variation exists at various growth and development stages for salinity tolerance. However, it was difficult to predict which salinity range will be appropriate for the screening, selection, and evaluation of genotypes that can best correlate with genetic diversity under field conditions. This kind of study is very important in order to develop efficient breeding programs and tool kits of salt tolerant crop genotypes and to assess the growth, physiological, and yield traits under field conditions [
18]. Efforts to enhance crop yields under salinity stress have also had limited success because the underlying mechanisms of salt tolerance have not been turned into useful selection criteria to evaluate a wide range of phenotypic plasticity and genotypes. Several authors have studied the salinity tolerance potential among a wide range of crop plants at the germination and seedling growth stages and showed a large genetic difference among them [
19,
20]. However, little attention has been paid to show a correlation regarding this early evaluation of salinity tolerance at germination with field condition [
21]. However, it is worthwhile to mention that these authors made significant efforts to explain the Na
+ exclusion, K
+ accumulation, and K
+/Na
+ as reliable indicators for selecting suitable genotypes that can tolerate soil and irrigation water salinity [
22]. The success of dual-purpose barley in marginal environments is subject to proper agronomic management practices along with the use of improved genotypes.
It is an urgent task of agronomists, plant physiologists, and plant breeders to identify and evaluate the genotypes and plant phenotypic plasticity using non-invasive, rapid, and reliable methods in order to screen the desired traits in a particular environment. The evaluation of the salinity tolerance potential of different genotypes and plant phenotypic attributes is highly necessary in order to understand physiological responses of the target genotypes and concerned traits associated with them [
23,
24]. The present situation can be changed through the introduction of new salt tolerant and higher yielding barley genotypes that have good yield stability and better salt tolerance potential. This will help to conserve freshwater resources as well as economic and ecological benefits for the sustainable development of salt-degraded marginal lands [
6,
25,
26]. It is important to screen, select, and evaluate the large collection of barley genotypes to check their performance (growth, yield stability, physiological characteristics) and traits are suited to salinity tolerance under field condition. In the present field study, a set of 28 genotypes from a previous trial [
27,
28,
29] were selected for elucidating the performance of different agronomical attributes (growth, number of tillers, plant biomass), yield traits (number of spikes, number of grains/spike, grain yield, harvest index), and biochemical attributes (Na
+, Cl
−, K
+), to find more suitable and tolerant genotypes under sandy marginal lands. The current study will provide a basis to promote barley cultivation on a large scale in the salt affected agro-ecosystem environment of the UAE. In addition, genotypes that showed stable yield and salt-tolerance potential will be included in the barley breeding programs for the development and release of salt-tolerant cultivars for seed multiplication and distribution among NARS for multi-location testing and large-scale cultivation.
The phenotypic plasticity, genotype variability, and agronomic adaptation of barley are extremely wide and vary significantly from hot arid to subtropical humid climates. Barley batini land races have not been characterized for salt tolerance on morphological, biochemical, ecophysiological, and isotopic bases. The main aim of the present study was the evaluation of batini barley land races and genotypes through the elucidation of salinity tolerance potential, growth performance, leaf ion homeostasis, leaf carbon and nitrogen isotope discrimination, intrinsic water use efficiency, harvest index, and yield stability attributes on six barley genotypes (113/1B, 59/3A, N1-10, N1-29, Barjouj, Alanda01) at three salinity levels (0, 7 and 14 dS m−1). For this study, it was hypothesized that batini barley land races and genotypes are genetically diverse and vary for salt tolerance potential. The evaluation of the plasticity of physiological attributes, such as number of tillers/m2, fresh biomass (FW), dry biomass (DW), grain yield, harvest index, and leaf Na+, K+, and Cl− concentration, leaf carbon and nitrogen isotope discrimination, and intrinsic water use efficiency, may help to develop a better understanding of mechanisms of salt tolerance.
4. Discussion
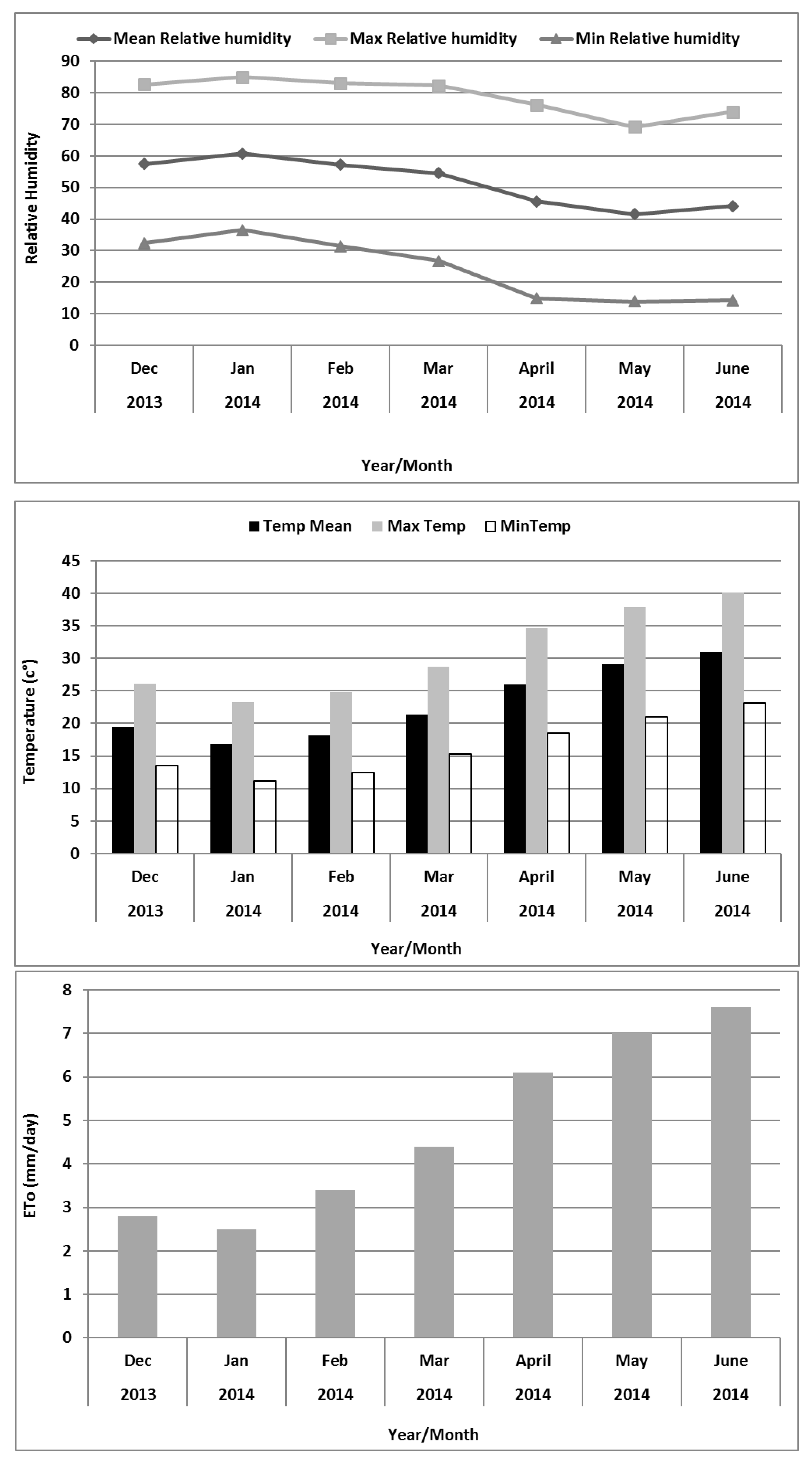
In hyper arid, salt-degraded, and marginal environments, there are several production constraints that significantly disturb growth, productivity, and crop yield stability. Under the prevailing conditions of the UAE, there is a severe lack of freshwater resources and most of it is only available for domestic purposes and other high value issues. In this situation, the management of available natural water resources (i.e., underground low-quality saline water) and nutrient poor sandy soils, and their conversion to a sustainable production system for food and feed is a most appropriate approach to the rehabilitation of these degraded lands. Soil biochemical analysis indicates that the soil is sandy with almost no organic matter content (
Table 3).
Irrigation with saline water decreased the plant dry biomass at all salinity levels, ranging from 16–31%. Meanwhile, genotype 113/1B exhibited the maximum dry biomass (116.2 t/ha) and Barjouj produced the lowest PDB (105 t ha
−1) (
Table 4). In this context, the number of tillers m
−2 decreased following exposure to higher salt stress and the reduction was 10–18% at 7–14 dS m
−1 NaCl stress. According to the reports of Arif et al. [
42], sodium stress is a serious global concern for sustainable agriculture that disrupts morphological, cellular, and physiological traits, affecting plant growth and development at all stages of development. Physiological traits, e.g., number of spikes m
−2, were also decreased at each salinity stress and the highest reduction (20%) was observed at 14 dS m
−1 NaCl, respectively, compared to the non-saline treatment (
Table 3). Barley genotypes 113/1B, 59/3A, and Alanda01 exhibit the highest tillers m
−2 while N1-29 exhibits the smallest tillers m
−2. There was significant variation in the production of the number of spikes
−2. The highest number of spikes m
−2 was obtained in barley variety Alanda01 followed by 113/1B while the lowest number of spikes m
−2 was exhibited by Barjouj, respectively (
Table 4). Genotype Alanda01 exhibited the highest grain numbers plant
−1 followed by 113/1B while the lowest grains/plant was produced by Barjouj. A similar pattern of variation was obtained for number of grains spike
−1 in the corresponding barley genotypes.
Understanding the biochemical, morphological, and physiological response mechanisms that play a role in improving adaptation to saline water environments is limited and the development of even more salt tolerant barley cultivars is of vital importance [
41,
42,
43,
44,
45]. This study investigated the salinity tolerance of genetically diverse barley genotypes and landraces based on agro-morphological, biochemical, physiological, and photosynthetic carbon isotope discrimination attributes in order to identify promising genotypes for salt tolerance screening. The current study showed that salt stress reduced PDB from 16% to 31% in field plots that received highly saline water (14 dS m
−1)(
Table 3). Barley genotype 113/1B showed higher dry biomass while Barjouj exhibited the lowest PDB (
Table 4). Morpho-physiological traits varied among barley genotypes due to genotypic differences, differences in saline environment, and also genotype by environment interactions. It is critical to understand the scope of such variations in order to develop breeding strategies and improve selection methods.
Salinity stress can cause inhibition of the photosynthetic process and hence agricultural productivity, yield stability, and environmental sustainability. Plants’ ability to become photosynthetically active in adverse saline conditions, on the other hand, is largely untapped. Salt stress has been shown to reduce barley yield by interfering with reproductive development and grain filling [
46,
47]. In barley, both successful seed setting and grain filling processes are critical for determining final grain yield. During the growth, reproductive, and grain filling periods, barley genotypes were exposed to salt stress (14 dS m
−1), with an average of number of spikes m
−2. However, 113/1B, 59/3A, and Alanda01 showed a greater number of tillers m
−2 as compared to other genotypes, while genotype N1-29 displayed the lowest tillers m
−2. We observed a significant reduction in grains per spike and grain weight across genotypes grown under saline conditions, resulting in a reduction in grain yield of 23% on average when compared to non-saline conditions (
Table 4). Genotype Alanda01 revealed highest grain numbers plant
−1 (527.9) followed by 113/1B (508.4) while Barjouj (480.5) produced the lowest grains plant
−1. Meanwhile, severe salinity stress during the grain filling stage may have an impact on other yield components, such as grain filling duration and grain filling process, and hence can cause significant effects in lowering grain weight and yield in barley [
46,
47,
48].
In response to salt stress, Na
+ and Cl
− levels were significantly higher in the barley leaves while the K
+/Na
+ ratio in the leaf tissues increased consistently. The K
+ content was higher with saline water, while the rest of the elements did not show any changes. The K
+ levels were consistent with K
+ availability, even under saline environment, and they could also be linked to the physiological changes seen in barley. Plant exposure to a saline environment can cause higher Na
+ absorption via roots, which leads to the development of osmotic and water stress [
48,
49,
50]. In comparison to the control, increased salinity levels resulted in an increase in tissue sodium and chloride content. Under severe salt stress, the increase in tissue sodium affects cell wall integrity and cell expansion, in addition to oxidative damage [
51]. In this context, Na
+ stress confines the absorption of other essential nutrient elements (K
+, Ca
2+, P, N) [
48,
52] that trigger the disturbance in the ion homeostasis, physiological, and biochemical cell activities [
53].
Genotypes Brjouj and N1-29 showed the highest grain yield among the salinity treatments as compared to control and at the same time also accumulated higher K
+ levels. It was also noticed that these genotypes have a substantial amount of Na
+ in the leaf tissue that might counterbalance the toxicity effect through the accumulation of K
+ ions. Potassium concentrations varied widely, 2.6-fold, ranging from 599.4 to 639.2 mg/100 g DW. Similar genotypic variation for salinity stress tolerance was demonstrated in barley [
54]. Such genotypic variation for salt tolerance might be due to the presence of a discrepancy among physiological traits, such as photosynthetic capacity, ion uptake, and maintenance of plant water status or antioxidant potential [
54]. Other researchers also demonstrated that barley exhibits tolerance to medium salinity [
55,
56]. Our results showed that N concentration increased after salinity treatments. Barley cultivars 113/1B and Barjouj showed highest N% and it was significantly higher than all other genotypes. The C% was higher in three barley genotype, 113/1B, N1-10, and Barjouj, respectively. Several researchers demonstrated that salt stress impedes the plant growth, physiological attributes, and yield contributing factors, such as the number of fertile tillers, grain weight, yield per square meter, and finally grain yield. The carbon metabolism, plant growth, and nutritional deficiency due to excess sodium accumulation in soil and plant tissues will lead to oxidative disorders and lower crop yield [
6,
7,
10,
57,
58,
59].
Effect of Irrigation Water Salinity, and Genotype on Grain Yield, Stable Isotope Composition of Carbon and Nitrogen
The assessment of stable isotopes of carbon and nitrogen (δ
13C and δ
15N) provides a very useful parameter that can help to analyze the impact of the surrounding environment in which the plants are growing. Meanwhile, carbon isotope discrimination can provide an integrated assessment of the stomatal regulation of internal CO
2 content as well as elaborate C
3 plant species’ long-term photosynthetic carbon [
32,
33]. Leaf growth and area development, photosynthesis, and nitrogen use are all closely related to crop yield. Salinity inhibits leaf growth, limiting grain yield and yield characteristics [
60]. The current findings show that when salinity increased from 7 to 14 dS m
−1, grain yield fell, ranging from 24% to 42.10%. Meanwhile,
Ci/
Ca was much lower, indicating that the stomata had closed (
Table 8). Stomatal closure can reduce CO
2 supply to carboxylation sites, lowering the activity of Ribulose-1,5-bisphosphate carboxylase oxygenase (RuBisCO), carbon synthesis, and translocation [
32,
33]. Higher Δ
13C is caused by a higher
Ci/
Ca ratio mainly due to a larger stomatal conductance, which can lead to a higher photosynthetic rate and hence a higher yield, i.e., positive relationship between Δ
13C and yield. Genotypes Barjouj and Alanda01 exhibit higher HI (36.6%, 36.2%), followed by N1-10 (26.8%), while the lowest HI was observed in 59/3A (20.8%) (
Table 8). When different barley genotypes were tested for salinity tolerance, they demonstrated better
Ci/
Ca and yield potential, indicating their adaption to the Dubai climate. There was a substantial difference in seed yield and harvest index between different genotypes which could be attributable to genetic differences. Such variances are much more pronounced in genotypes Barjouj and Alanda01, and 59/3A which had grain yield variation of 1.6%. HI (%) was decreased by 14% and 9.86% at 20 and 10 dS m
−1 salinity, respectively, as compared to control (
Table 7). This is due to some genotypes’ superior tolerance to the UAE’s agro-climatic conditions. Genotypes Alanda01 and Barjouj had the maximum photosynthetic CO
2 rate (
Ci/
Ca), yield, and productivity and were the most suited and well-adapted genotypes for the Dubai marginal soil environment. N1-10 and N-29 had the lowest rates (37% and 30% lower
Ci/
Ca than Barjouj), indicating that they were the least adapted. The
Ci/
Ca ratio of intercellular to ambient CO
2 concentrations did not differ significantly between the remaining genotypes (113/1B, 59/3A, Barjouj, Alanda01).
Although variation in plant N isotopic composition (
15N) does not offer a measure of NUE, it can be used to follow N mobility and infer N sources and/or N cycle dynamics in vegetation at the local, community, and landscape scales. The diffusion gradient for CO
2 into the leaf through the stomata is linked to both the efficiency of water usage (carbon (C) fixed per unit water transpired) and the efficiency of N use (C fixed per unit N absorbed). Plants need the majority of their water to support photosynthesis through transpiration. Photosynthesis accounts for more than half of total leaf N [
61], and total leaf N content and photosynthetic capability are frequently associated [
62]. If the CO
2 diffusion gradient steepens, reductions in stomatal conductance (gs) or higher investments in foliar N can result in higher water-use efficiency (WUE), while lower intercellular CO
2 concentrations can diminish N-use efficiency (NUE) by reducing rates of C fixation per cell. For salinity treatment, phenotypic differences for
15N characteristics were also investigated, and they were found to be larger in treated plants (4.5 and 4.8) than in control treatments (3.3). In terms of nitrogen isotope composition, most genotypes were in the range of 4.4–4.6, while Alanda01 had low
15N (3.13) values when compared to other genotypes. The leaf N concentration has a substantial G x T interaction, and tolerant genotypes’
15N was lowered to a greater extent than sensitive genotypes under all salinity stress conditions, resulting in a significant GxT interaction (
Table 8). Carbon isotope discrimination (Δ
13C), the difference in
13C/
12C composition between plant C and environmental CO
2, has frequently been used to estimate WUE. Previous studies have demonstrated negative correlations between Δ
13C and WUE under a CO
2 in various species, such as barley, cowpea, and wheat [
63,
64,
65,
66]. Following salinity treatment, the intrinsic water use efficiency (iWUE) values dramatically rise. The genotype N1-29 exhibited highest iWUE values, followed by N1-10, while Barjouj demonstrated the lowest iWUE values (
Table 8).


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}