
Beneficial Role of Fruits, Their Juices, and Freeze-Dried Powders on Inflammatory Bowel Disease and Related Dysbiosis

 , ,
, ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Inflammatory Bowel Disease (IBD)
3. Gut Microbiota
4. Dysbiosis and IBD
5. Research Methodology
6. Fruit Intake and Potential Benefits on Gut Microbiota
6.1. Orange and Mandarin (Citrus Genus)
6.2. Cherries (Prunus Genus)
6.3. Banana (Musa L. Genus)
6.4. Apple (Malus Mill. Genus)
6.5. Blueberry and Cranberry (Vaccinium Genus)
6.6. Mulberry (Morus Genus)
6.7. Strawberry (Fragaria) and Raspberry (Rubus)
6.8. Goji (Lycium Genus)
6.9. Pomegranate (Punica Genus)
6.10. Noni Fruit (Moringa Genus)
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| AA-2βG | 2-O-β-D-glucopyranosyl-L-ascorbic acid |
| CD | Chron’s disease |
| CTX | cyclophosphamide |
| DGGE | denaturing gradient gel electrophoresis |
| DSS | dextran sulfate sodium |
| ENT | enteral nutritional therapy |
| EO-IBD | early-onset inflammatory bowel disease |
| FMT | fecal microbiota transplantation |
| GI | gastrointestinal |
| IBD | inflammatory bowel disease |
| JAK/STAT | Janus kinase/signal transducer and activator of transcription |
| LDL | low-density lipoproteins |
| miRNAs | MicroRNAs |
| n-3 PUFA | n-3 polyunsaturated fatty acids |
| n-9 PUFA | n-9 polyunsaturated fatty acids |
| SCFAs | short-chain fatty acids |
| GPR43 | G-protein-coupled receptor 43 |
| SOD | superoxide dismutase |
| TNBS | trinitrobenzene sulfonic-acid |
| TNF-α | tumor necrosis factor-alpha |
| UC | ulcerative colitis |
| VEO-IBD | very-early-onset inflammatory bowel disease |
References
- Kaplan, G.G. The global burden of IBD: From 2015 to 2025. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Loddo, I.; Romano, C. Inflammatory bowel disease: Genetics, epigenetics, and pathogenesis. Front. Immunol. 2015, 6, 551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weimers, P.; Munkholm, P. The natural history of IBD: Lessons learned. Curr. Treat. Options. Gastro. 2018, 16, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Vatn, M.H. Natural history and complications of IBD. Curr. Gastroenterol. Rep. 2009, 11, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Mulder, D.J.; Noble, A.J.; Justinich, C.J.; Duffin, J.M. A Tale of two diseases: The history of inflammatory bowel disease. J. Crohns Colitis 2014, 8, 341–348. [Google Scholar] [CrossRef] [Green Version]
- Farzaei, M.H.; Bahramsoltani, R.; Abdolghaffari, A.H.; Sodagari, H.R.; Esfahani, S.A.; Rezaei, N. A Mechanistic review on plant-derived natural compounds as dietary supplements for prevention of inflammatory bowel disease. Expert. Rev. Gastroenterol. Hepatol. 2016, 10, 745–758. [Google Scholar] [CrossRef] [PubMed]
- Larussa, T.; Imeneo, M.; Luzza, F. Potential role of nutraceutical compounds in Inflammatory Bowel Disease. World J. Gastroenterol. 2017, 23, 2483–2492. [Google Scholar] [CrossRef]
- Neurath, M.F. Current and emerging therapeutic targets for IBD. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 269–278. [Google Scholar] [CrossRef] [Green Version]
- Lucendo, A.J.; De Rezende, L.C. Importance of nutrition in inflammatory bowel disease. World J. Gastroenterol. 2009, 15, 2081–2088. [Google Scholar] [CrossRef]
- Raman, M.; Ghosh, S. Diet and nutrition in IBD—Progress and gaps. Nutrients 2019, 11, 1740. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.D.; Bushmanc, F.D.; Lewis, J.D. Diet, the human gut microbiota, and IBD. Anaerobe 2013, 24, 117–120. [Google Scholar] [CrossRef]
- Tomasello, G.; Mazzola, M.; Leone, A.; Sinagra, E.; Zummo, G.; Farina, F.; Damiani, P.; Cappello, F.; Gerges Geagea, A.; Jurjus, A.; et al. Nutrition, oxidative stress and intestinal dysbiosis: Influence of diet on gut microbiota in inflammatory bowel diseases. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czech Repub. 2016, 160, 461–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, G.G.; Windsor, J.W. The Four epidemiological stages in the global evolution of inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Ananthakrishnan, A.N.; Bernstein, C.N.; Iliopoulos, D.; Macpherson, A.; Neurath, M.F.; Ali, R.A.R.; Vavricka, S.R.; Fiocchi, C. Environmental triggers in IBD: A review of progress and evidence. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Mentella, M.C.; Scaldaferri, F.; Pizzoferrato, M.; Gasbarrini, A.; Miggiano, G.A.D. Nutrition, IBD and gut microbiota: A review. Nutrients 2020, 12, 944. [Google Scholar] [CrossRef] [Green Version]
- Guan, Q. A Comprehensive review and update on the pathogenesis of inflammatory bowel disease. J. Immunol. Res. 2019, 2019, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Khan, I.; Ullah, N.; Zha, L.; Bai, Y.; Khan, A.; Zhao, T.; Che, T.; Zhang, C. Alteration of gut microbiota in inflammatory bowel disease (IBD): Cause or consequence? IBD treatment targeting the gut microbiome. Pathogens 2019, 8, 126. [Google Scholar] [CrossRef] [Green Version]
- Chudy-Onwugaje, K.O.; Christian, K.E.; Farraye, F.A.; Cross, R.K. A state-of-the-art review of new and emerging therapies for the treatment of IBD. Inflamm. Bowel Dis. 2019, 25, 820–830. [Google Scholar] [CrossRef]
- Park, S.C.; Jeen, Y.T. Anti-integrin therapy for inflammatory bowel disease. World J. Gastroenterol. 2018, 24, 1868–1880. [Google Scholar] [CrossRef] [Green Version]
- Salas, A.; Hernandez-Rocha, C.; Duijvestein, M.; Faubion, W.; McGovern, D.; Vermeire, S.; Vetrano, S.; Casteele, N.V. JAK–STAT pathway targeting for the treatment of inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 323–337. [Google Scholar] [CrossRef]
- Soendergaard, C.; Bergenheim, F.H.; Bjerrum, J.T.; Nielsen, O.H. Targeting JAK-STAT signal transduction in IBD. Pharmacol. Ther. 2018, 192, 100–111. [Google Scholar] [CrossRef] [PubMed]
- Peyrin-Biroulet, L.; Christopher, R.; Behan, D.; Lassen, C. Modulation of sphin-gosine-1-phosphate in inflammatory bowel disease. Autoimmun. Rev. 2017, 16, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef] [PubMed]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proctor, L.M. The human microbiome project in 2011 and beyond. Cell Host Microbe 2011, 10, 287–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donaldson, G.P.; Lee, S.M.; Mazmanian, S.K. Gut biogeography of the bacterial microbiota. Nat. Rev. Microbiol. 2016, 14, 20–32. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Chang, E.B. Exploring gut microbes in human health and disease: Pushing the envelope. Genes Dis. 2014, 1, 132–139. [Google Scholar] [CrossRef] [Green Version]
- Umbrello, G.; Esposito, S. Microbiota and neurologic diseases: Potential effects of probiotics. J. Transl. Med. 2016, 14, 298. [Google Scholar] [CrossRef] [Green Version]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2009, 9, 313–323. [Google Scholar] [CrossRef]
- Cryan, J.F.; Dinan, T.G. Mind-altering microorganisms: The impact of the gut microbiota on brain and behaviour. Nat. Rev. Neurosci. 2012, 13, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Weiss, G.A.; Hennet, T. Mechanisms and consequences of intestinal dysbiosis. Cell. Mol. Life Sci. 2017, 74, 2959–2977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levy, M.; Kolodziejczyk, A.A.; Thaiss, C.A.; Elinav, E. Dysbiosis and the immune system. Nat. Rev. Immunol. 2017, 17, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Ajslev, T.A.; Andersen, C.S.; Gamborg, M.; Sørensen, T.I.A.; Jess, T. Childhood overweight after establishment of the gut microbiota: The role of delivery mode, pre-pregnancy weight and early administration of antibiotics. Int. J. Obes. 2011, 35, 522–529. [Google Scholar] [CrossRef] [Green Version]
- Jostins, L.; Ripke, S.; Weersma, R.K.; Duerr, R.H.; McGovern, D.P.; Hui, K.Y.; Lee, J.C.; Schumm, L.P.; Sharma, Y.; Anderson, C.A.; et al. Host-microbe interactions have shaped the genetic architecture of inflammatory bowel disease. Nature 2012, 491, 119–124. [Google Scholar] [CrossRef] [Green Version]
- Nishida, A.; Inoue, R.; Inatomi, O.; Bamba, S.; Naito, Y.; Andoh, A. Gut microbiota in the pathogenesis of inflammatory bowel disease. Clin. J. Gastroenterol. 2018, 11, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Forbes, J.D.; Chen, C.Y.; Knox, N.C.; Marrie, R.A.; El-Gabalawy, H.; de Kievit, T.; Alfa, M.; Bernstein, C.N.; Van Domselaar, G. A comparative study of the gut microbiota in immune-mediated in-flammatory diseases—Does a common dysbiosis exist? Microbiome 2018, 6, 1–15. [Google Scholar] [CrossRef]
- Zhao, L. The gut microbiota and obesity: From correlation to causality. Nat. Rev. Microbiol. 2013, 11, 639–647. [Google Scholar] [CrossRef]
- Li, W.Z.; Stirling, K.; Yang, J.J.; Zhang, L. Gut microbiota and diabetes: From correlation to causality and mechanism. World J. Diabetes 2020, 11, 293–308. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, W.D.; Wang, Y.D. The relationship between gut microbiota and inflammatory diseases: The role of macrophages. Front. Microbiol. 2020, 11, 1065. [Google Scholar] [CrossRef]
- Vinolo, M.A.; Rodrigues, H.G.; Nachbar, R.T.; Curi, R. Regulation of inflammation by short chain fatty acids. Nutrients 2011, 3, 858–876. [Google Scholar] [CrossRef] [Green Version]
- Lavelle, A.; Sokol, H. Gut microbiota-derived metabolites as key actors in inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 223–237. [Google Scholar] [CrossRef]
- Ni, J.; Wu, G.D.; Albenberg, L.; Tomov, V.T. Gut microbiota and IBD: Causation or correlation? Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 573–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barber, T.M.; Kabisch, S.; Pfeiffer, A.F.H.; Weickert, M.O. The health benefits of dietary fibre. Nutrients 2020, 12, 3209. [Google Scholar] [CrossRef]
- Liu, S.; Lee, I.-M.; Ajani, U.; Cole, S.R.; Buring, J.E.; Manson, J.E. Intake of vegetables rich in carotenoids and risk of coronary heart disease in men: The physicians’ health study. Int. J. Epidemiol. 2001, 30, 130–135. [Google Scholar] [CrossRef] [Green Version]
- Bazzano, L.A.; He, J.; Ogden, L.G.; Loria, C.M.; Vupputuri, S.; Myers, L.; Whelton, P.K. Fruit and vegetable intake and risk of cardiovascular disease in US adults: The first national health and nutrition examination survey epidemiologic follow-up study. Am. J. Clin. Nutr. 2002, 76, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Bazzano, L.A.; Serdula, M.K.; Liu, S. Dietary intake of fruits and vegetables and risk of cardiovascular disease. Curr. Atheroscler. Rep. 2003, 5, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.H. Health benefits of fruit and vegetables are from additive and synergistic combinations of phytochemicals. Am. J. Clin. Nutr. 2003, 78, 517S–520S. [Google Scholar] [CrossRef]
- Cui, J.; Lian, Y.; Zhao, C.; Du, H.; Han, Y.; Gao, W.; Xiao, H.; Zheng, J. Dietary fibers from fruits and vegetables and their health benefits via modulation of gut microbiota. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1514–1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klinder, A.; Shen, Q.; Heppel, S.; Lovegrove, J.A.; Rowland, I.; Tuohy, K.M. Impact of increasing fruit and vegetables and flavonoid intake on the human gut microbiota. Food Funct. 2016, 7, 1788–1796. [Google Scholar] [CrossRef]
- Gibson, G.R. Fibre and effects on probiotics (the prebiotic concept). Clin. Nutr. Suppl. 2004, 1, 25–31. [Google Scholar] [CrossRef]
- Holscher, H.D. Dietary fiber and prebiotics and the gastrointestinal microbiota. Gut Microbes 2017, 8, 172–184. [Google Scholar] [CrossRef]
- Etxeberria, U.; Fernández-Quintela, A.; Milagro, F.I.; Aguirre, L.; Martínez, J.A.; Portillo, M.P. Impact of polyphenols and polyphenol-rich dietary sources on gut microbiota composition. J. Agric. Food Chem. 2013, 61, 9517–9533. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Xu, X.; Li, J.; Li, F. Association between gut microbiota and autism spectrum disorder: A systematic review and meta-analysis. Front. Psychiatry 2019, 10, 473. [Google Scholar] [CrossRef]
- Thumann, T.A.; Pferschy-Wenzig, E.-M.; Moissl-Eichinger, C.; Bauer, R. The role of gut microbiota for the activity of medicinal plants traditionally used in the European Union for gastrointestinal disorders. J. Ethnopharmacol. 2019, 245, 112153. [Google Scholar] [CrossRef]
- Luo, J.; Lin, X.; Bordiga, M.; Brennan, C.; Xu, B. Manipulating effects of fruits and vegetables on gut microbiota—A critical review. Int. J. Food Sci. Technol. 2021, 56, 2055–2067. [Google Scholar] [CrossRef]
- Musumeci, L.; Maugeri, A.; Cirmi, S.; Lombardo, G.E.; Russo, C.; Gangemi, S.; Calapai, G.; Navarra, M. Citrus fruits and their flavonoids in inflammatory bowel disease: An overview. Nat. Prod. Res. 2020, 34, 122–136. [Google Scholar] [CrossRef]
- Hossen, I.; Wu, H.; Luo, T.; Mehmood, A.; Jingyi, S.; Yanping, C.; Hongqing, W.; Zhipeng, G.; Kaiqi, Z.; Fang, Y.; et al. Phytochemicals and inflammatory bowel disease: A review. Crit. Rev. Food Sci. Nutr. 2019, 60, 1–25. [Google Scholar] [CrossRef]
- Somani, S.J.; Modi, K.P.; Majumdar, A.S.; Sadarani, B.N. Phytochemicals and their potential usefulness in inflammatory bowel disease. Phytother. Res. 2015, 29, 339–350. [Google Scholar] [CrossRef]
- Pei, R.; Liu, X.; Bolling, B. Flavonoids and gut health. Curr. Opin. Biotechnol. 2020, 61, 153–159. [Google Scholar] [CrossRef]
- Espín, J.C.; González-Sarrías, A.; Tomás-Barberán, F.A. The gut microbiota: A Key factor in the therapeutic effects of (poly)phenols. Biochem. Pharmacol. 2017, 139, 82–93. [Google Scholar] [CrossRef]
- Ozdal, T.; Sela, D.A.; Xiao, J.; Boyacioglu, D.; Chen, F.; Capanoglu, E. The reciprocal interactions between polyphenols and gut microbiota and effects on bioaccessibility. Nutrients 2016, 8, 78. [Google Scholar] [CrossRef] [PubMed]
- Cardona, F.; Andrés-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuño, M.I. Benefits of polyphenols on gut microbiota and implications in human health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef] [Green Version]
- Valdés, L.; Cuervo, A.; Salazar, N.; Ruas-Madiedo, P.; Gueimonde, M.; González, S. The relationship between phenolic compounds from diet and microbiota: Impact on human health. Food Funct. 2015, 6, 2424–2439. [Google Scholar] [CrossRef]
- Davinelli, S.; Scapagnini, G. Interactions between dietary polyphenols and aging gut microbiota: A review. BioFactors 2021. [Google Scholar] [CrossRef]
- Fidélix, M.; Milenkovic, D.; Sivieri, K.; Cesar, T. Microbiota modulation and effects on metabolic biomarkers by orange juice: A controlled clinical trial. Food Funct. 2020, 11, 1599–1610. [Google Scholar] [CrossRef]
- Lima, A.C.D.; Cecatti, C.; Fidélix, M.P.; Adorno, M.A.T.; Sakamoto, I.K.; Cesar, T.B.; Sivieri, K. Effect of daily consumption of orange juice on the levels of blood glucose, lipids, and gut microbiota metabolites: Controlled clinical trials. J. Med. Food. 2019, 22, 202–210. [Google Scholar] [CrossRef]
- Brasili, E.; Hassimotto, N.M.A.; Del Chierico, F.; Marini, F.; Quagliariello, A.; Sciubba, F.; Miccheli, A.; Putignani, L.; Lajolo, F. Daily consumption of orange juice from Citrus Sinensis L. Osbeck Cv. Cara Cara and Cv. Bahia differently affects gut microbiota profiling as unveiled by an integrated meta-omics approach. J. Agric. Food Chem. 2019, 67, 1381–1391. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Li, Y.; Liu, M.; Yu, H.; Chen, Q.; Chen, Y.; Ruan, J.; Ding, Z.; Zhang, Y.; Wang, T. Citrus Aurantium L. and its flavonoids regulate TNBS-induced inflammatory bowel disease through anti-inflammation and suppressing isolated jejunum contraction. Int. J. Mol. Sci. 2018, 19, 3057. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Cao, X.; Fang, X.; Guo, A.; Li, E. Inhibitory effects of fermented ougan (Citrus Reticulata Cv. Suavissima) juice on high-fat diet-induced obesity associated with white adipose tissue browning and gut microbiota modulation in mice. Food Funct. 2021, 12, 9300–9314. [Google Scholar] [CrossRef]
- Mayta-Apaza, A.C.; Pottgen, E.; De Bodt, J.; Papp, N.; Marasini, D.; Howard, L.; Abranko, L.; Van de Wiele, T.; Lee, S.-O.; Carbonero, F. Impact of tart cherries polyphenols on the human gut microbiota and phenolic metabolites in vitro and in vivo. J. Nutr. Biochem. 2018, 59, 160–172. [Google Scholar] [CrossRef]
- Othaim, A.A.; Marasini, D.; Carbonero, F. Impact of increasing concentration of tart and sweet cherries juices concentrates on healthy mice gut microbiota. Food Front. 2020, 1, 224–233. [Google Scholar] [CrossRef]
- Lear, R.; O’Leary, M.; O’Brien Andersen, L.; Holt, C.C.; Stensvold, C.R.; van der Giezen, M.; Bowtell, J.L. Tart cherry concentrate does not alter the gut microbiome, glycaemic control or systemic inflammation in a middle-aged population. Nutrients 2019, 11, 1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitsou, E.K.; Kougia, E.; Nomikos, T.; Yannakoulia, M.; Mountzouris, K.C.; Kyriacou, A. Effect of banana consumption on faecal microbiota: A randomised, controlled trial. Anaerobe 2011, 17, 384–387. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, G.H.; Larson, C.P.; Islam, R.; Saha, U.R.; Kabir, A. Green banana-supplemented diet in the home management of acute and prolonged diarrhoea in children: A community-based trial in rural Bangladesh. Trop. Med. Int. Health 2010, 15, 1132–1139. [Google Scholar] [CrossRef] [PubMed]
- Cassettari, V.M.G.; Machado, N.C.; de Arruda Lourenção, P.L.T.; Carvalho, M.A.; Ortolan, E.V.P.; Cassettari, V.M.G. Combinations of laxatives and green banana biomass on the treatment of functional constipation in children and adolescents: A randomized study. J. Pediatr. 2019, 95, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, K.; Ohashi, Y.; Kawasumi, K.; Terada, A.; Fujisawa, T. Effect of apple intake on fecal microbiota and metabolites in humans. Anaerobe 2010, 16, 510–515. [Google Scholar] [CrossRef] [PubMed]
- Ravn-Haren, G.; Dragsted, L.O.; Buch-Andersen, T.; Jensen, E.N.; Jensen, R.I.; Németh-Balogh, M.; Paulovicsová, B.; Bergström, A.; Wilcks, A.; Licht, T.R.; et al. Intake of whole apples or clear apple juice has contrasting effects on plasma lipids in healthy volunteers. Eur. J. Nutr. 2013, 52, 1875–1889. [Google Scholar] [CrossRef] [PubMed]
- Vendrame, S.; Guglielmetti, S.; Riso, P.; Arioli, S.; Klimis-Zacas, D.; Porrini, M. Six-week consumption of a wild blueberry powder drink increases Bifidobacteria in the human gut. J. Agric. Food Chem. 2011, 59, 12815–12820. [Google Scholar] [CrossRef] [PubMed]
- Lacombe, A.; Li, R.W.; Klimis-Zacas, D.; Kristo, A.S.; Tadepalli, S.; Krauss, E.; Young, R.; Wu, V.C.H. Lowbush wild blueberries have the potential to modify gut microbiota and xenobiotic metabolism in the rat colon. PLoS ONE 2013, 8, e67497. [Google Scholar] [CrossRef]
- Lee, S.; Keirsey, K.I.; Kirkland, R.; Grunewald, Z.I.; Fischer, J.G.; de La Serre, C.B. Blueberry supplementation influences the gut microbiota, inflammation, and insulin resistance in high-fat-diet-fed rats. J. Nutr. 2018, 148, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Paturi, G.; Mandimika, T.; Butts, C.A.; Zhu, S.; Roy, N.C.; McNabb, W.C.; Ansell, J. Influence of dietary blueberry and broccoli on cecal microbiota activity and colon morphology in Mdr1a(-/-) mice, a model of inflammatory bowel diseases. Nutrition 2012, 28, 324–330. [Google Scholar] [CrossRef]
- Song, H.; Shen, X.; Chu, Q.; Zheng, X. Vaccinium Bracteatum Thunb. fruit extract reduces high-fat diet-induced obesity with modulation of the gut microbiota in obese mice. J. Food Biochem. 2021, 45, e13808. [Google Scholar] [CrossRef]
- Cai, X.; Han, Y.; Gu, M.; Song, M.; Wu, X.; Li, Z.; Li, F.; Goulette, T.; Xiao, H. Dietary cranberry suppressed colonic inflammation and alleviated gut microbiota dysbiosis in dextran sodium sulfate-treated mice. Food Funct. 2019, 10, 6331–6341. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.; Wu, Z.; Huang, L.; Qiu, H.; Wang, L.; Li, L.; Yao, L.; Kang, K.; Qu, J.; Wu, Y.; et al. Mulberry fruit prevents LPS-induced NF-ΚB/PERK/MAPK signals in macrophages and suppresses acute colitis and colorectal tumorigenesis in mice. Sci. Rep. 2015, 5, 17348. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Hatabu, T. Mulberry juice freeze-dried powder attenuates the disease severity by the maintaining of colon mucosa in mice with DSS-induced acute colitis. Biosci. Biotechnol. Biochem. 2019, 83, 914–922. [Google Scholar] [CrossRef] [Green Version]
- Kanodia, L.; Borgohain, M.; Das, S. Effect of fruit extract of Fragaria Vesca L. on experimentally induced inflammatory bowel disease in albino rats. Indian J. Pharmacol. 2011, 43, 18–21. [Google Scholar] [CrossRef] [PubMed]
- Montrose, D.C.; Horelik, N.A.; Madigan, J.P.; Stoner, G.D.; Wang, L.-S.; Bruno, R.S.; Park, H.J.; Giardina, C.; Rosenberg, D.W. Anti-inflammatory effects of freeze-dried black raspberry powder in ulcerative colitis. Carcinogenesis 2011, 32, 343–350. [Google Scholar] [CrossRef]
- Ding, Y.; Yan, Y.; Chen, D.; Ran, L.; Mi, J.; Lu, L.; Jing, B.; Li, X.; Zeng, X.; Cao, Y. Modulating effects of polysaccharides from the fruits of Lycium Barbarum on the immune response and gut microbiota in cyclophosphamide-treated mice. Food Funct. 2019, 10, 3671–3683. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Yan, Y.; Wan, P.; Chen, D.; Ding, Y.; Ran, L.; Mi, J.; Lu, L.; Zhang, Z.; Li, X.; et al. Gut microbiota modulation and anti-inflammatory properties of anthocyanins from the fruits of Lycium Ruthenicum Murray in dextran sodium sulfate-induced colitis in mice. Free Radic. Biol. Med. 2019, 136, 96–108. [Google Scholar] [CrossRef]
- Song, H.; Shen, X.; Chu, Q.; Zheng, X. Pomegranate fruit pulp polyphenols reduce diet-induced obesity with modulation of gut microbiota in mice. J. Sci. Food Agric. 2021. [Google Scholar] [CrossRef]
- Yang, X.; Mo, W.; Zheng, C.; Li, W.; Tang, J.; Wu, X. Alleviating effects of noni fruit polysaccharide on hepatic oxidative stress and inflammation in rats under a high-fat diet and its possible mechanisms. Food Funct. 2020, 11, 2953–2968. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Wang, F.; Lian, Y.; Xiao, H.; Zheng, J. Biosynthesis of Citrus flavonoids and their health effects. Crit. Rev. Food Sci. Nutr. 2020, 60, 566–583. [Google Scholar] [CrossRef] [PubMed]
- Stevens, Y.; Rymenant, E.V.; Grootaert, C.; Camp, J.V.; Possemiers, S.; Masclee, A.; Jonkers, D. The intestinal fate of Citrus flavanones and their effects on gastrointestinal health. Nutrients 2019, 11, 1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, J. Dietary flavonoid aglycones and their glycosides: Which show better biological significance? Crit. Rev. Food Sci. Nutr. 2017, 57, 1874–1905. [Google Scholar] [CrossRef]
- Büyüktuncel, S. Fast Determination of naringin and hesperidin in natural and commercial Citrus juices by HPLC method. Asian J. Chem. 2017, 29, 2384–2386. [Google Scholar] [CrossRef]
- Ribeiro, I.A.; Ribeiro, M.H.L. Naringin and naringenin determination and control in grapefruit juice by a validated HPLC method. Food Control. 2008, 19, 432–438. [Google Scholar] [CrossRef]
- Belajová, E.; Suhaj, M. Determination of phenolic constituents in Citrus juices: Method of high performance Liquid chromatography. Food Chem. 2004, 86, 339–343. [Google Scholar] [CrossRef]
- Williamson, G. The role of polyphenols in modern nutrition. Nutr Bull. 2017, 42, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Estruel-Amades, S.; Massot-Cladera, M.; Pérez-Cano, F.J.; Franch, À.; Castell, M.; Camps-Bossacoma, M. Hesperidin effects on gut microbiota and gut-associated lymphoid tissue in healthy rats. Nutrients 2019, 11, 324. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wu, Y.; Zhao, X.; Luo, F.; Li, X.; Zhu, H.; Sun, C.; Chen, K. Chemopreventive effect of flavonoids from ougan (Citrus Reticulata Cv. Suavissima) fruit against cancer cell proliferation and migration. J. Funct. Foods 2014, 10, 511–519. [Google Scholar] [CrossRef]
- Li, D.; Wang, P.; Luo, Y.; Zhao, M.; Chen, F. Health benefits of anthocyanins and molecular mechanisms: Update from recent decade. Crit. Rev. Food Sci. Nutr. 2017, 57, 1729–1741. [Google Scholar] [CrossRef]
- Fang, J. Classification of fruits based on anthocyanin types and relevance to their health effects. Nutrition 2015, 31, 1301–1306. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidhu, J.S.; Zafar, T.A. Bioactive compounds in banana fruits and their health benefits. Food Qual. Saf. 2018, 2, 183–188. [Google Scholar] [CrossRef]
- Moongngarm, A. chemical compositions and resistant starch content in starchy foods. Am. J. Agric. Biol. Sci. 2013, 8, 107–113. [Google Scholar] [CrossRef] [Green Version]
- Rabbani, G.H.; Albert, M.J.; Rahman, H.; Chowdhury, A.K. Short-chain fatty acids inhibit fluid and electrolyte loss induced by cholera toxin in proximal colon of rabbit in vivo. Dig. Dis. Sci. 1999, 44, 1547–1553. [Google Scholar] [CrossRef]
- Rabbani, G.H.; Teka, T.; Zaman, B.; Majid, N.; Khatun, M.; Fuchs, G.J. Clinical studies in persistent diarrhea: Dietary management with green banana or pectin in Bangladeshi children. Gastroenterology 2001, 121, 554–560. [Google Scholar] [CrossRef]
- Escarpa, A.; González, M.C. High-performance liquid chromatography with diode-array detection for the determination of phenolic compounds in peel and pulp from different apple varieties. J. Chromatogr. A 1998, 823, 331–337. [Google Scholar] [CrossRef]
- Koutsos, A.; Tuohy, K.M.; Lovegrove, J.A. Apples and cardiovascular health—Is the gut microbiota a core consideration? Nutrients 2015, 7, 3959–3998. [Google Scholar] [CrossRef]
- Larrosa, M.; Luceri, C.; Vivoli, E.; Pagliuca, C.; Lodovici, M.; Moneti, G.; Dolara, P. Polyphenol metabolites from colonic microbiota exert anti-inflammatory activity on different inflammation models. Mol. Nutr. Food Res. 2009, 53, 1044–1054. [Google Scholar] [CrossRef]
- Monagas, M.; Khan, N.; Andrés-Lacueva, C.; Urpí-Sardá, M.; Vázquez-Agell, M.; Lamuela-Raventós, R.M.; Estruch, R. Dihydroxylated phenolic acids derived from microbial metabolism reduce lipopolysaccharide-stimulated cytokine secretion by human peripheral blood mononuclear cells. Br. J. Nutr. 2009, 102, 201–206. [Google Scholar] [CrossRef] [Green Version]
- Selma, M.V.; Espin, J.C.; Tomas-Barberan, F.A. Interaction between phenolics and gut microbiota: Role in human health. J. Agric. Food Chem. 2009, 57, 6485–6501. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, M.; Oruna-Concha, M.J.; Kolida, S.; Walton, G.E.; Kallithraka, S.; Spencer, J.P.; de Pascual-Teresa, S. Metabolism of anthocyanins by human gut microflora and their influence on gut bacterial growth. J. Agric. Food Chem. 2012, 60, 3882–3890. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Fischer, J.; Wicker, L. Intermolecular binding of blueberry pectin-rich fractions and anthocyanin. Food Chem. 2016, 194, 986–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, M.; Li, T.; Li, Y.; Qian, H.; Zhang, H.; Rao, Z.; Wang, L. Vaccinium Bracteatum Thunb. as a promising resource of bioactive compounds with health benefits: An updated review. Food Chem. 2021, 356, 129738. [Google Scholar] [CrossRef] [PubMed]
- De Pádua Lúcio, K.; Rabelo, A.C.S.; Araújo, C.M.; Brandão, G.C.; de Souza, G.H.B.; da Silva, R.G.; de Souza, D.M.S.; Talvani, A.; Bezerra, F.S.; Cruz Calsavara, A.J.; et al. Anti-inflammatory and antioxidant properties of black mulberry (Morus Nigra L.) in a model of LPS-induced sepsis. Oxid. Med. Cell Longev. 2018, 2018, 5048031. [Google Scholar] [CrossRef] [Green Version]
- Crespo, I.; San-Miguel, B.; Mauriz, J.L.; Ortiz de Urbina, J.J.; Almar, M.; Tuñón, M.J.; González-Gallego, J. Protective effect of protocatechuic acid on TNBS-induced coli-tis in mice is associated with modulation of the SphK/S1P signaling pathway. Nutrients 2017, 9, 288. [Google Scholar] [CrossRef]
- Zhang, Z.; Wu, X.; Cao, S.; Cromie, M.; Shen, Y.; Feng, Y.; Yang, H.; Li, L. Chlorogenic acid ameliorates experimental colitis by promoting growth of Akkermansia in mice. Nutrients 2017, 9, 677. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Hu, M.; Zhang, L.; Gao, Y.; Ma, L.; Yan, X.; Xu, Q. Dietary taxifolin potently protects against dextran sulfate sodium-induced colitis via NF-κB signaling, enhancing intestinal barrier and modulating gut microbiota. Front. Immunol. 2020, 11, 3915. [Google Scholar]
- Dong, Y.; Lei, J.; Zhang, B. Dietary quercetin alleviated DSS-induced colitis in mice through several possible pathways by transcriptome analysis. Curr. Pharm. Biotechnol. 2020, 21, 1666–1673. [Google Scholar] [CrossRef]
- Abdel-Raheem, I.T. Gastroprotective effect of rutin against indomethacin-induced ulcers in rats. Basic Clin. Pharmacol. Toxicol. 2010, 107, 742–750. [Google Scholar] [CrossRef]
- Zhang, W.; Han, F.; He, J.; Duan, C. HPLC-DAD-ESI-MS/MS analysis and antioxidant activities of nonanthocyanin phenolics in mulberry (Morus Alba L.). J. Food Sci. 2008, 73, C512–C518. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Chung, M.S. Antiviral Activities of mulberry (Morus Alba) juice and seed against influenza viruses. Evid Based Complement. Alternat. Med. 2018, 2018, e2606583. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shao, S.; Guo, C.; Zhang, S.; Li, M.; Ding, K. The Homogenous polysaccharide SY01-23 purified from leaf of Morus Alba L. has bioactivity on human gut Bacteroides ovatus and Bacteroides cellulosilyticus. Int. J. Biol. Macromol. 2020, 158, 698–707. [Google Scholar] [CrossRef]
- Chen, T.; Shi, N.; Afzali, A. Chemopreventive effects of strawberry and black raspberry on colorectal cancer in inflammatory bowel disease. Nutrients 2019, 11, 1261. [Google Scholar] [CrossRef] [Green Version]
- Huang, K.; Dong, W.; Liu, W.; Yan, Y.; Wan, P.; Peng, Y.; Xu, Y.; Zeng, X.; Cao, Y. 2-O-β-D-Glucopyranosyl-L-ascorbic acid, an ascorbic acid derivative isolated from the fruits of Lycium Barbarum L., modulates gut microbiota and palliates colitis in dextran sodium sulfate-induced colitis in mice. J. Agric. Food Chem. 2019, 67, 11408–11419. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Yan, Y.; Chen, D.; Zhao, Y.; Dong, W.; Zeng, X.; Cao, Y. Ascorbic acid derivative 2-O-β-D-Glucopyranosyl-L-ascorbic acid from the fruit of Lycium Barbarum modulates microbiota in the small intestine and colon and exerts an immunomodulatory effect on cyclophosphamide-treated BALB/c Mice. J. Agric. Food Chem. 2020, 68, 11128–11143. [Google Scholar] [CrossRef] [PubMed]
- Jurenka, J. Therapeutic applications of pomegranate (Punica Granatum L.): A review. Altern. Med. Rev. 2008, 13, 128–144. [Google Scholar]
- Shaygannia, E.; Bahmani, M.; Zamanzad, B.; Rafieian-Kopaei, M. A review study on Punica Granatum L. J. Evid. Based Complementary Altern. Med. 2016, 21, 221–227. [Google Scholar] [CrossRef] [Green Version]
- Chan-Blanco, Y.; Vaillant, F.; Mercedes Perez, A.; Reynes, M.; Brillouet, J.-M.; Brat, P. The noni fruit (Morinda Citrifolia L.): A review of agricultural research, nutritional and therapeutic properties. J. Food Compost. Anal. 2006, 19, 645–654. [Google Scholar] [CrossRef]
- West, B.J.; Jensen, C.J.; Westendorf, J.; White, L.D. A safety review of noni fruit juice. J. Food Sci. 2006, 71, R100–R106. [Google Scholar] [CrossRef]
- Potterat, O.; Hamburger, M. Morinda Citrifolia (noni) fruit—Phytochemistry, pharmacology, safety. Planta Med. 2007, 73, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Graf, D.; Di Cagno, R.; Fåk, F.; Flint, H.J.; Nyman, M.; Saarela, M.; Watzl, B. Contribution of diet to the composition of the human gut microbiota. Microb. Ecol. Health Dis. 2015, 26, 26164. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Liu, H.; Qin, N.; Ren, X.; Zhu, B.; Xia, X. Impact of food additives on the composition and function of gut microbiota: A review. Trends Food Sci. Technol. 2020, 99, 295–310. [Google Scholar] [CrossRef]
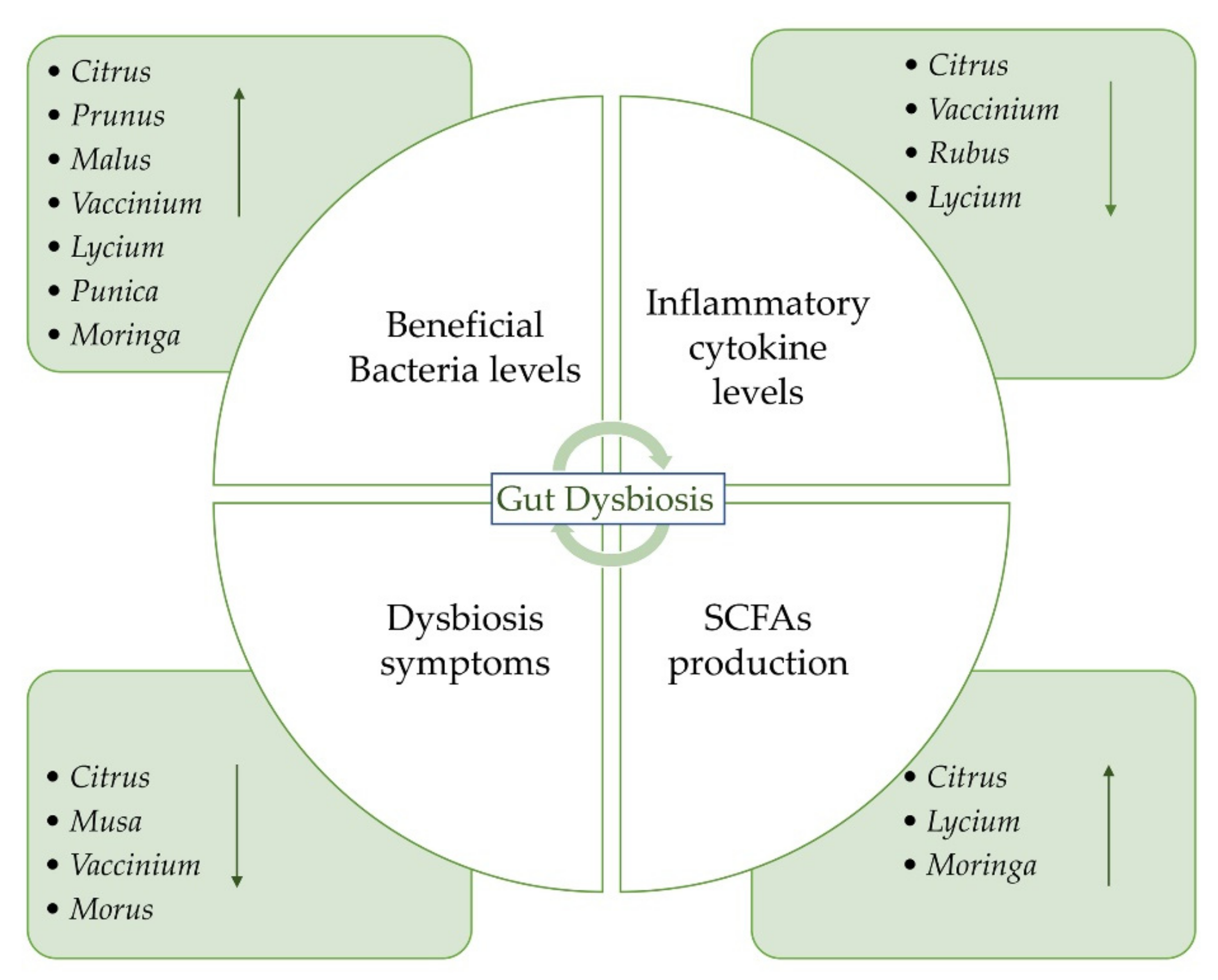
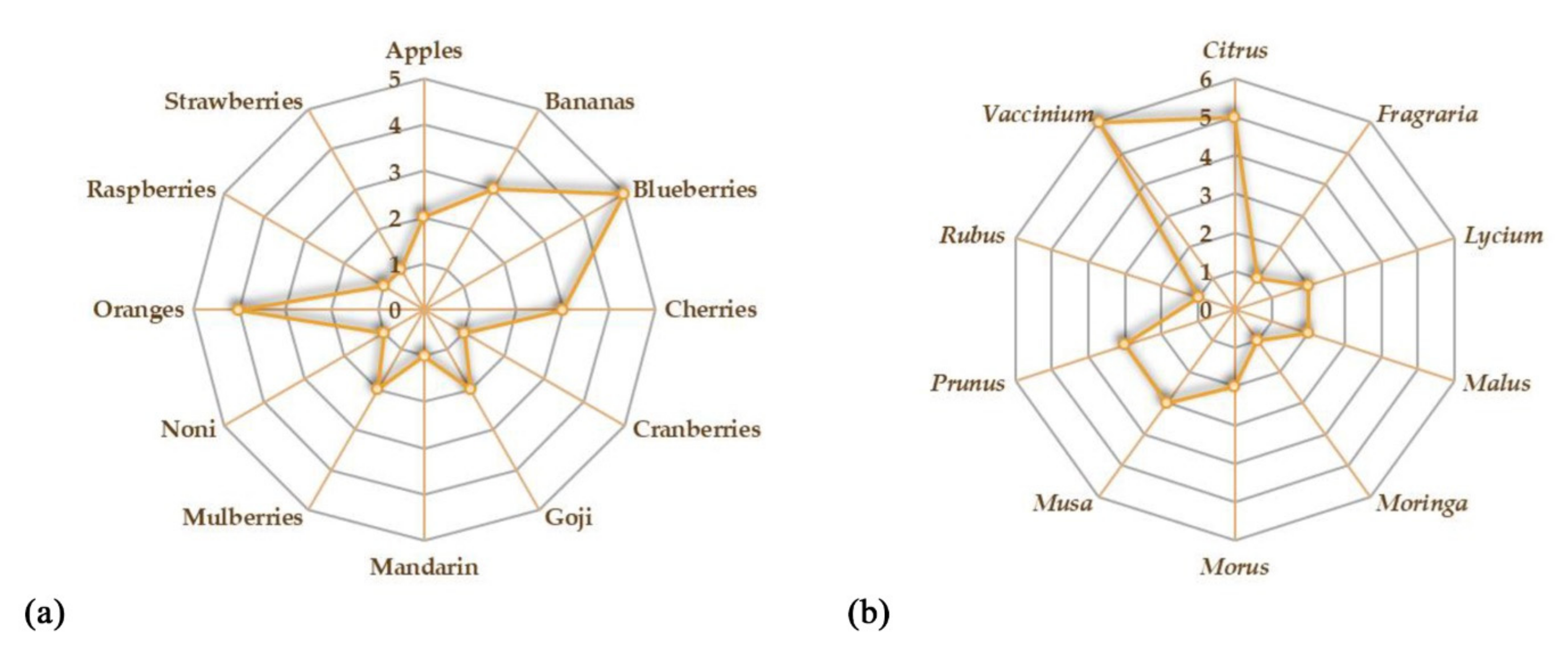


{kind=link}
{kind=link}
{kind=link}
| Plant Genus | Plant Species | Sample/Extract | Study | Animal Model/Participants | Treatment | Results | Ref |
|---|---|---|---|---|---|---|---|
| Citrus | Orange (Citrus sinensis L.) | Juice (‘Pera Rio’ variety) | In vivo | 10 women | Consumption of 300 mL/d for 60 days | Increased levels of Lactobacillus, Akkermasia, and Ruminococcus spp. | [65] |
| Juice | In vivo | 10 healthy women | 2 months of commercial pasteurized orange juice consumption | Increased fecal composition of Bifidobacterium and Lactobacillus spp. | [66] | ||
| Juice (‘Cara Cara’ and ‘Bahia’) | In vivo | 21 healthy volunteers (18–45 years) | Daily supplementation of two orange juices with different flavanone content for 7 days | Increased abundance of Clostridia operational taxonomic units from Mogibacteriaceae, Tissierellaceae, Veillonellaceae, Odoribacteraceae, and Ruminococcaceae families | [67] | ||
| Bitter orange (C. aurantium L.) | Dried fruits | In vivo | Trinitrobenzene sulfonic-acid (TNBS)-induced IBD in rats | From 125 to 500 mg/kg in rats with IBD | Decreased colitis inflammatory cell infiltration and inflammatory cytokine levels and ameliorated weight loss and diarrhea symptoms | [68] | |
| ‘Ougan’ mandarin (C. reticulata cv. suavissima) | Lyophilized juice | In vivo | Fifty high-fat diet (HFD)-fed C57Bl/6J male mice | 20 mL kg−1 daily | Reduced abundance of phylum Erysipelatoclostridiaceae and remarkably increase in SCFA-producing bacteria Blautia | [69] | |
| Prunus | Tart cherry (Prunus cerasus L.) | Juice | In vitro | - | - | Increase in Bacteroides and Bifidobacterium | [70] |
| Juice | In vivo | 10 young, healthy participants (5 males, 5 females) | 8 oz. of juice daily for 5 days | Increase in Ruminococcus, Lachnospiraceae, and Collinsella in high-Bacteroides individuals. Increase in Bacteroides in low-Bacteroides | [70] | ||
| Juice | In vivo | 45 mice | juice added to drinking water for 23 days | Increase in Barnesiella and Akkermansia | [71] | ||
| Concentrate | In vivo | 28 participants (40–60 years) | 60 mL per day for 4 weeks | Supplementation did not alter gut microbiome | [72] | ||
| Prunus | Sweet cherry (Prunus avium L.) | Juice | In vivo | 45 mice | Increased concentration of juice added to drinking water for 23 days | Increase in Barnesiella and Akkermansia | [71] |
| Musa | Banana | Fruit and flavored drink | In vivo | 34 healthy women (19–45 years) | Pre-meal snack (fruit or banana-flavored drink) twice a day | Not statistically significant increase in Bifidobacterium levels after 30 and 60 days of intervention | [73] |
| Cooked green banana mixed with rice flour | In vivo | 62 children | 250 g/L of cooked green banana for 7 days | Reduced vomiting, stool, and diarrheal duration and reduced need for oral rehydration solution | [74] | ||
| Cooked fruits | In vivo | 80 children and adolescents | - | Green banana biomass resulted advantageous as an adjunct therapy on functional constipation, mainly for reducing doses of laxatives | [75] | ||
| Malus | Apple | Fresh fruit | In vivo | 8 healthy male volunteers (21–60 years) | 2 apples per day for 2 weeks | Increased fecal Bifidobacteria; reduced Enterobacteriaceae and lecithinase-positive Clostridia, including C. perfringens | [76] |
| Apples, apple pomace, and juice | In vivo | 23 healthy volunteers | 5 × 4 weeks dietary crossover study: whole apples (550 g/day), apple pomace (22 g/day), apple juices (500 mL/day) | A modulation of the gut microbiota population was not confirmed | [77] | ||
| Vaccinium | Blueberry (V. angustifolium Ait.) | Wild blueberry drink | In vivo | 20 healthy male volunteers | Wild blueberry drink (25 g of wild blueberry powder in 250 mL of water) for 6 weeks and daily placebo drink for further 6 weeks | Increased Bifidobacterium spp. | [78] |
| Blueberry powder | In vivo | Sprague-Dawley rats | Control diet + 8% w/w blueberry powdersubstituting for dextrose | Increased relative abundance of Bifidobacteriaceae and Coriobacteriaceae family | [79] | ||
| Blueberry powder | In vivo | 24 male Wistar rats | 10 g freeze-dried blueberry powder/100 g diets for 8 weeks | Increased Gammaproteobacteria abundance | [80] | ||
| Blueberry (V. corymbosum L.) | Fruit (hybrid “Marimba,” “Misty”, “O’Neal”) | In vivo | IBD mouse model | 200 g/kg for 21 weeks | Altered composition and metabolism of the cecal microbiota and colon morphology | [81] | |
| Oriental blueberry (V. bracteatum Thunb) | Polyphenol-rich fruit extract | In vivo | High-fat diet (HFD)-induced obese mice | 150 mg/kg body weight for 14 weeks | Reduced HFD induced body weight gain and insulin resistance index; increased abundances of Verrucomicrobia and Bacteroidetes, and decreased Proteobacteria and Firmicutes | [82] | |
| Cranberry (V. macrocarpon Aiton) | Freeze-dried whole cranberry powder | In vivo | Dextran sulfate sodium (DSS)-induced acute colitis in mice | Diet + 1.5% (w/w) freeze-dried whole cranberry powder | Decrease in the severity of colitis and of proinflammatory cytokines levels. Increase in the abundance Lactobacillus and Bifidobacterium and decrease in Sutterella and Bilophila | [83] | |
| Morus | Mulberry (Morus alba L.) | Mulberry fruits extracts | In vivo | DSS-induced acute colitis in mice | 5% or 10% mulberry extracts for 3 months | Amelioration of colitis and prevention of severe injuries in colon crypts | [84] |
| Mulberry juice freeze-dried powder | In vivo | DSS-induced acute colitis in BALB/c mice | 28 days | Mitigation of DSS-induced acute colitis by changing the gut microbial flora and by improving mucosal conditions | [85] | ||
| Fragaria | Strawberry (Fragaria vesca L.) | Ethanolic extract | In vivo | Acetic acid-induced IBD in rats | 500 mg/kg for 5 days | Improved colon architecture and reduced tissue oxidative stress, with a significant improvement of superoxide dismutase and catalase tissue levels | [86] |
| Rubus | Raspberry | Freeze-dried black raspberry powder | In vivo | DSS-induced ulcerative colitis in C57BL/6J mice | 5% or 10% raspberry sample for 7–14 days | Anti-inflammatory activity, with reduction in colonic shortening and ulceration and suppression of different proinflammatory cytokines | [87] |
| Lycium | Goji (L. barbarum L.) | Extract from dried fruits | In vivo | Cyclophosphamide (CTX)-induced mice | 50, 100, and 200 mg/kg d−1 by intragastric gavage for 9 days | Promoted production of short-chain fatty acids. Increase in Bacteroidaceae, Lactobacillaceae, Prevotellaceae and Verrucomicrobiaceae | [88] |
| Black goji (L. ruthenicum Murray) | Anthocyanins extract from fruits | In vivo | DSS-induced colitis in C57BL/6 mice | 200 mg/kg/d for 8 days | Decrease in the expression of proinflammatory cytokines; reversal of DSS-induced decreases in relative abundances of Porphyromonadaceae, Rikenellaceae and Prevotellaceae | [89] | |
| Punica | Pomegranate (P. granatum L.) | Polyphenol extract from fresh fruits | In vivo | C57BL/6J mice | 200 mg/kg daily by intragastric administration for 14 weeks | Changes in gut microbiota composition. Increased abundance of Akkermansia muciniphila, Bacteroides acidifaciens, Mucispirillum schaedleri, and other species | [90] |
| Moringa | Noni (M. citrifolia L.) | Freeze-dried polysaccharide extract from dehydrated fruits | In vivo | Sprague-Dawley rats | 100 mg per kg bw for 5 weeks | Improved cecal short-chain fatty acid (SCFA) production; improvement in the levels of Lactobacillus, Ruminococcaceae, and Parasutterella, and decrement in the levels of Prevotella_9, Collinsella, and Bacteroides | [91] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perri, M.R.; Romano, C.; Marrelli, M.; Zicarelli, L.; Toma, C.-C.; Basta, D.; Conforti, F.; Statti, G. Beneficial Role of Fruits, Their Juices, and Freeze-Dried Powders on Inflammatory Bowel Disease and Related Dysbiosis. Plants 2022, 11, 4. https://doi.org/10.3390/plants11010004
Perri MR, Romano C, Marrelli M, Zicarelli L, Toma C-C, Basta D, Conforti F, Statti G. Beneficial Role of Fruits, Their Juices, and Freeze-Dried Powders on Inflammatory Bowel Disease and Related Dysbiosis. Plants. 2022; 11(1):4. https://doi.org/10.3390/plants11010004
Chicago/Turabian StylePerri, Maria Rosaria, Carmen Romano, Mariangela Marrelli, Ludovica Zicarelli, Claudia-Crina Toma, Daniele Basta, Filomena Conforti, and Giancarlo Statti. 2022. "Beneficial Role of Fruits, Their Juices, and Freeze-Dried Powders on Inflammatory Bowel Disease and Related Dysbiosis" Plants 11, no. 1: 4. https://doi.org/10.3390/plants11010004
APA StylePerri, M. R., Romano, C., Marrelli, M., Zicarelli, L., Toma, C.-C., Basta, D., Conforti, F., & Statti, G. (2022). Beneficial Role of Fruits, Their Juices, and Freeze-Dried Powders on Inflammatory Bowel Disease and Related Dysbiosis. Plants, 11(1), 4. https://doi.org/10.3390/plants11010004