
The Allelopathic Potential of Rosa blanda Aiton on Selected Wild-Growing Native and Cultivated Plants in Europe

 , , and
, , and
Abstract
1. Introduction
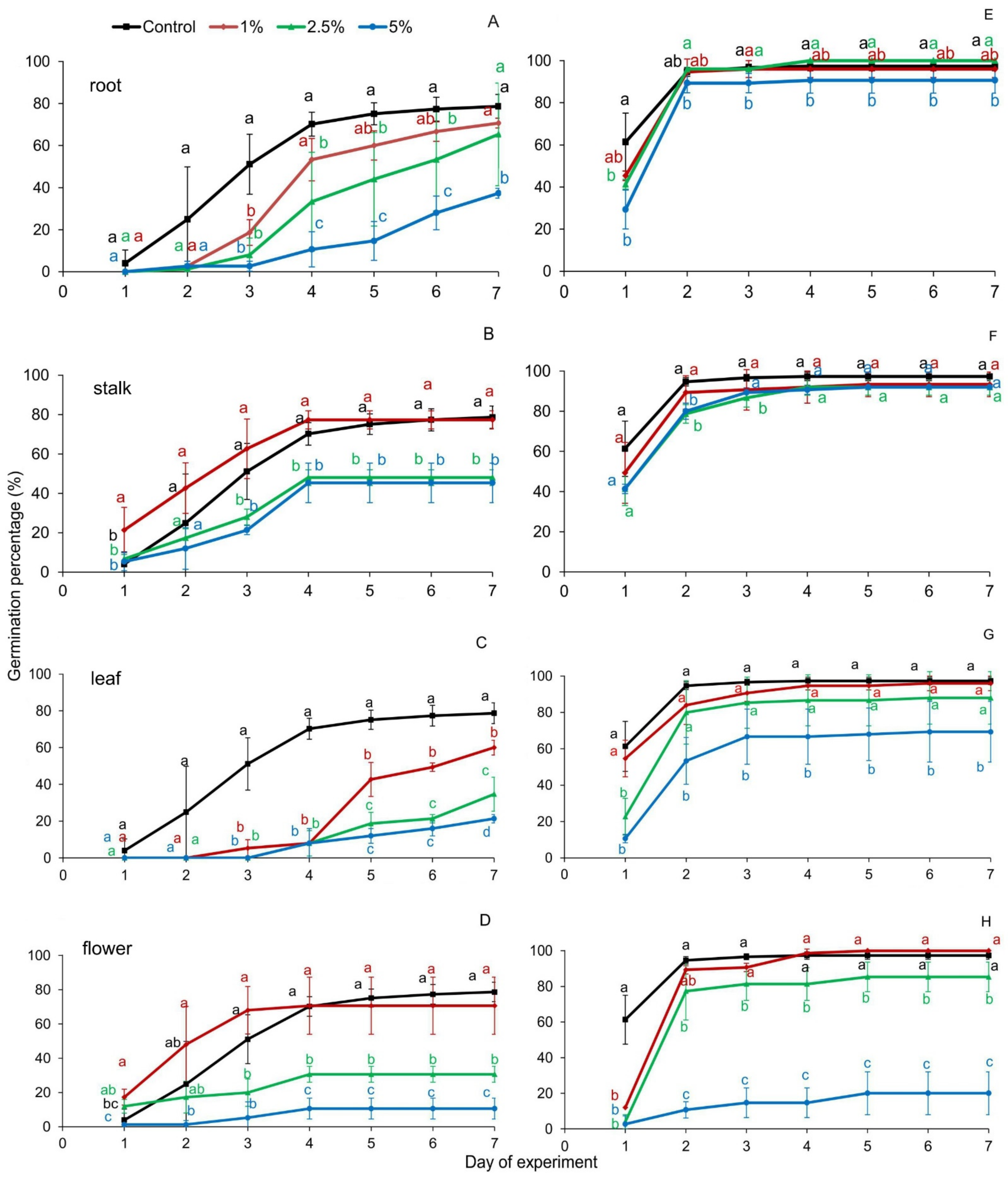
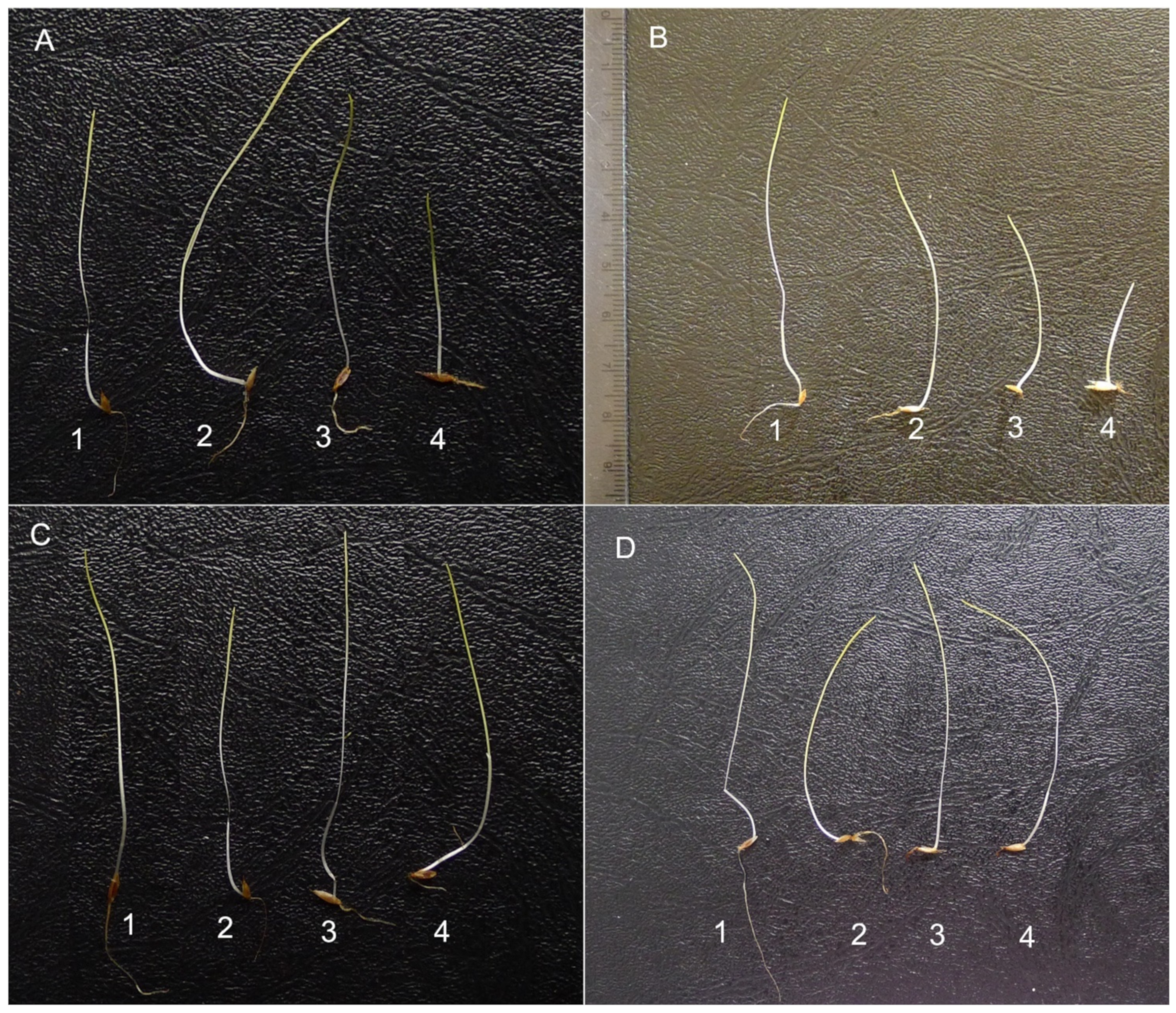
2. Results
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Extract Preparation
4.3. PH of Extracts
4.4. Seeds Preparation and Germination Conditions
4.5. Germination Indexes
D—the number of days counted from the beginning of germination, N—number of seeds that appeared on day D
n3 and n7 = number of germinated seeds on time T3 and T7, T3 and T7 = 3 and 7 days after germination
C—control germination speed, T—treatment germination speed
4.6. Seedling Elongation Growth
LE—seedling length (mm) treated with the aqueous extract, LC—seedling length (mm) treated with the distilled water (control group)
4.7. Seedlings Biomass
WC—water content, DM—dry mass, FM—fresh mass
4.8. Electrolyte Leakage
EL—electrolyte leakage, E1—EL from live seedlings, E2—EL from dead seedlings
4.9. Statistic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nilsson, M.; Wardle, D.A. Understory vegetation as a forest ecosystem driver: Evidence from the northern Swedish boreal forest. Front. Ecol. Environ. 2005, 3, 421–428. [Google Scholar] [CrossRef]
- Abhilasha, D.; Quintana, N.; Vivanco, J.; Joshi, J. Do allelopathic compounds in invasive Solidago canadensis s.l. restrain the native European flora? J. Ecol. 2008, 96, 993–1001. [Google Scholar] [CrossRef]
- Willis, R.J. The History of Allelopathy, 1st ed.; Springer: Dordrecht, The Netherlands, 2007; p. 206. [Google Scholar]
- Farooq, M.; Jabran, K.; Cheema, Z.; Wahid, A.; Siddique, H.M.K. The role of allelopathy in agricultural pest management. Pest. Manag. Sci. 2011, 67, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.H.; Tanveer, A.; Nadeem, M.A.; Javaid, M.M.; Kashif, M.S.; Andchadhar, A.R. Allelopathic effects of Rhynchosia capitata on germination and seedling growth of mung bean. Planta Daninha 2013, 31, 501–509. [Google Scholar] [CrossRef]
- Cheng, F.; Cheng, Z. Research progress on the use of plant allelopathy in agriculture and the physiological and ecological mechanisms of allelopathy. Front. Plant Sci. 2015, 6, 1020. [Google Scholar] [CrossRef]
- Mominul Islam, A.K.M.; Sabina, Y.; Qasem, J.R.S.; Juraimi, A.; Parvez, A. Allelopathy of medicinal plants: Current status and future prospects in weed management. Agric. Sci. 2018, 9, 89160. [Google Scholar] [CrossRef]
- Putnan, A.R.; Tang, C.S. The Science of Allelopathy; John Wiley Sons: New York, NY, USA, 1986; pp. 292–293. [Google Scholar] [CrossRef]
- Kroymann, J. Natural diversity and adaptation in plant secondary metabolism. Curr. Opin. Plant Biol. 2011, 14, 246–251. [Google Scholar] [CrossRef]
- Inderjit; Weston, L.A.; Duke, S.O. Challenges, achievements and opportunities in allelopathy research. J. Plant Interact. 2005, 1, 69–81. [Google Scholar] [CrossRef]
- Yoneya, K.; Takabayashi, J. Plant–plant communication mediated by airborne signals: Ecological and plant physiological perspectives. Plant Biotechnol. 2014, 31, 409–416. [Google Scholar] [CrossRef]
- Radosevich, S.R.; Holt, J.S.; Ghersa, C. Ecology of Weeds and Invasive Plants: Relationship to Agriculture and Natural Resource Management; Wiley: New York, NY, USA, 2007; p. 454. [Google Scholar]
- De Albuquerque, M.B.; Santos, R.C.; Lima, L.M.; Melo Filho, P.A.; Nogueira, R.J.M.C.; Da Câmara, C.A.G.; Ramos, A.R. Allelopathy, an alternative tool to improve cropping systems. A review. Agron. Sustain. Dev. 2011, 31, 379–395. [Google Scholar] [CrossRef]
- Eichenberg, D.; Ristok, C.; Kroeber, W.; Bruelheide, H. Plant polyphenols—Implications of different sampling, storage and sample processing in biodiversity-ecosystem functioning experiments. Chem. Ecol. 2014, 30, 676–692. [Google Scholar] [CrossRef]
- Tokarska-Guzik, B.; Dajdok, Z.; Zając, M.; Zając, A.; Urbisz, A.; Danielewicz, W.; Hołdyński, C. Plants of Foreign Origin in Poland, with Particular Emphasis on Invasive Species (Rośliny Obcego Pochodzenia w Polsce ze Szczególnym UwzglęDnieniem Gatunków Inwazyjnych); Generalna Dyrekcja Ochrony Środowiska: Warszawa, Poland, 2012; pp. 1–197.
- Stephen, H.A. Woody Plants of the North Central Plains; University press of Kansas: Lawrence, KS, USA, 1973; pp. 268–269. [Google Scholar]
- Gleason, H.A.; Cronquist, A. Manual of Vascular Plants of Northeastern United States and Adjacent Canada, 2nd ed.; The New York Botanical Garden: New York, NY, USA, 1991; p. 910. [Google Scholar]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. Flora Europaea. Volume 2. Rosaceae to Umbelliferae; Cambridge University Press: Cambridge, UK, 1968; p. 455. [Google Scholar]
- Zieliński, J. Rosa L. In Flora of Poland; Jasiewicz, A., Ed.; Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 1987; Volume 5, p. 48. [Google Scholar]
- Kurtto, A. Rosaceae (pro parte majore). Euro + Med Plantbase—The Information Resource for Euro-Mediterranean Plant Diversity. Rosa blanda Aiton. Available online: http://ww2.bgbm.org/euroPlusMed/PTaxonDetail.asp?UUID=8447153D-554D-4BEC-98FE-16DEAEF5A989 (accessed on 15 January 2021).
- CABI. Rosa blanda. In Invasive Species Compendium; CAB International: Wallingford, UK; Available online: www.cabi.org/isc (accessed on 15 January 2021).
- GBIF Database. Available online: https://www.discoverlife.org (accessed on 16 January 2021).
- Rosa blanda Aiton. Plants of the World Online. Royal Botanic Gardens, Kew. Available online: http://plantsoftheworldonline.org/taxon/urn:lsid:ipni.org:names:222243-2 (accessed on 30 December 2019).
- Hereźniak, J. American trees and shrubs in Poland (Amerykańskie drzewa i krzewy na ziemiach polskich). In Plants of American Origin at Home in Poland (Rośliny Pochodzenia Amerykańskiego Zadomowione w Polsce); Ławrynowicz, M., Warcholińska, U.A., Eds.; Łódzkie Towarzystwo Naukowe: Łódź, Poland, 1992; pp. 97–150. [Google Scholar]
- Czarna, A. Vascular Plants of Central Wielkopolska (Rośliny naczyniowe środkowej Wielkopolski); Wydawnictwo Uniwersytetu Przyrodniczego: Poznań, Poland, 2009; p. 184. [Google Scholar]
- Sołtys-Lelek, A.; Gruszka, W. Occurrence of Rosa blanda Ait. (Rosaceae) in Poland. AUPC Stud. Nat. 2020, 5, 25–33. [Google Scholar] [CrossRef]
- List of species alien in Europe and to Europe. In Handbook of Alien Species in Europe; Invading Nature—Springer Series in Invasion Ecology; Springer: Dordrecht, The Netherlands, 2009; Volume 3. [CrossRef]
- United States Department of Agriculture. Available online: https://planthardiness.ars.usda.gov/PHZMWeb/ (accessed on 15 December 2020).
- Lewis, W.H.; Ertter, B.; Bruneau, A. Rosa. In Flora of North America North of Mexico; Oxford University Press: Oxford, NY, USA, 2014; Volume 9. [Google Scholar]
- Hobbs, R.J.; Humphries, S.E. An integrated approach to the ecology and management of plant invasions. Conserv. Biol. 1995, 9, 761–770. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P. Plant invasions: Merging the concepts of species invasiveness and community invasibility. Progr. Phys. Geogr. 2006, 30, 409–443. [Google Scholar] [CrossRef]
- Amante, M.A. An Overview of Roses. Chemical Composition of North American Roses. In a Characterization of Five Native Wisconsin Roses and Their Hips. Master’s Thesis, Horticulture and Agroecology University of Wisconsin, Madison, WI, USA, 2016; pp. 1–9. [Google Scholar]
- Kolhmunzer, S. Farmakognozja, 5th ed.; PZWL Wydawnictwo Lekarskie: Warszawa, Poland, 2012. [Google Scholar]
- Barros, L.; Carvalho, A.M.; Morais, J.S.; Ferreira, I.C.F.R. Strawberry-tree, blackthorn and rose fruits: Detailed characterisation in nutrients and phytochemicals with antioxidant properties. Food Chem. 2010, 120, 247–254. [Google Scholar] [CrossRef]
- Günes, M. Pomological and phenological characteristics of promising rose hip (Rosa) genotypes. Afr. J. Biotechnol. 2010, 9, 6301–6306. [Google Scholar] [CrossRef]
- Demir, N.; Yildiz, O.; Alpaslan, M.; Hayaloglu, A.A. Evaluation of volatiles, phenolic compounds and antioxidant activities of rose hip (Rosa L.) fruits in Turkey. LWT-Food Sci. Technol. 2014, 57, 126–133. [Google Scholar] [CrossRef]
- Czyżowska, A.; Klewicka, E.; Pogorzelski, E.; Nowak, A. Polyphenols, vitamin C and antioxidant activity in wines from Rosa canina L. and Rosa rugosa Thunb. J. Food Compos. Anal. 2015, 39, 62–68. [Google Scholar] [CrossRef]
- Yi, O.; Jovel, E.M.; Towers, G.H.N.; Wahbe, T.R.; Cho, D. Antioxidant and antimicrobial activities of native Rosa sp. from British Columbia, Canada. Int. J. Food Sci. Nutr. 2007, 58, 178–189. [Google Scholar] [CrossRef]
- Barry, R.; Sanderson, K.; Fillmore, S. Establishment of wild roses for commercial rose hip production in Atlantic Canada. Int. J. Fruit Sci. 2008, 8, 266–281. [Google Scholar] [CrossRef]
- Sanderson, K.; Fillmore, S. Evaluation of native rose selections for rose hip production in Prince Edward Island. Int. J. Fruit Sci. 2010, 10, 379–389. [Google Scholar] [CrossRef]
- Ghose, K.; McCallum, J.; Fillmore, S.; Kirby, C.; Sanderson, K.; Joly, S.; Fofana, B. Structuration of the genetic and metabolite diversity among Prince Edward Island cultivated wild rose ecotypes. Sci. Hortic. 2013, 160, 251–263. [Google Scholar] [CrossRef]
- Mikanagi, Y.; Yokoi, M.; Ueda, Y.; Saito, N. Flower flavonol and anthocyanin distribution in subgenus Rosa. Biochem. System. Ecol. 1995, 23, 183–200. [Google Scholar] [CrossRef]
- Mikanagi, Y.; Saito, N.; Yokoi, M.; Tatsuzawa, F. Anthocyanins in flowers of genus Rosa, sections Cinnamomeae (=Rosa), Chinenses, Gallicanae and some modern garden roses. Biochem. System. Ecol. 2000, 28, 887–902. [Google Scholar] [CrossRef]
- Mikanagi, Y.; Saito, N.; Yokoi, M. Flavonoid distribution in the flowers of genus Rosa, sections Synstylae, Cinnamomeae and Pimpinellifoliae. In Flavonoids in Biology and Medicine III, Current Issues in Flavonoid Research; National University of Singapore: Singapore, 1990; pp. 89–98. [Google Scholar]
- Weidema, I. NOBANIS—Invasive Alien Species Fact Sheet—Rosa rugosa—From: Online Database of the European Network on Invasive Alien Species—NOBANIS. 2006. Available online: http://www.nobanis.org (accessed on 21 May 2021).
- Starfinger, U. Introduction and naturalization of Prunus serotina in Central Europe. In Plant Invasions: Studies from North America and Europe; Brock, J.H., Wade, M., Pysek, P., Green, D., Eds.; Backhuys Publishers: Leiden, The Netherlands, 1997; pp. 161–171. [Google Scholar]
- Cierjacks, A.; Kowarik, I.; Joshi, J.; Hempel, S.; Ristow, M.; von der Lippe, M.; Weber, E. Biological flora of the British Isles: Robinia pseudoacacia. J. Ecol. 2017, 273, 1623–1640. [Google Scholar] [CrossRef]
- Zohaib, A.; Abbas, T.; Tabassum, T. Weeds cause losses in field crops through allelopathy. Not. Sci. Biol. 2016, 8, 47–56. [Google Scholar] [CrossRef][Green Version]
- Callaway, R.M.; Ridenour, W.M. Novel weapons: Invasive success and the evolution of increased competitive ability. Front. Ecol. Environ. 2004, 2, 436–443. [Google Scholar] [CrossRef]
- Hierro, J.L.; Callaway, R.M. Allelopathy and exotic plant invasion. Plant Soil 2003, 256, 29–39. [Google Scholar] [CrossRef]
- Brown, R.T. Influence of naturally occurring compounds on germination and growth of jack pine. Ecology 1967, 48, 541–546. [Google Scholar] [CrossRef]
- Drogoszewski, B.; Barzdajn, W. Wpływ ekstraktów wodnych z tkanek Prunus serotina Ehrh. na kiełkowanie nasion Pinus silvestris L. Pr. Kom. Nauk. Rol. Kom. Nauk. Leśnych Poznańskiego Tow. Przyj. Nauk. Wydziału Nauk. Rol. Leśnych 1984, 58, 33–38. [Google Scholar]
- Vilhena, K.S.S.; Guilhon, G.M.S.P.; Zoghbi, M.G.B.; Santos, L.S.; Souza Filho, A.P.S. Chemical investigation of Cyperus distans L. and inhibitory activity of scabequinone in seed germination and seedling growth bioassays. Nat. Prod. Res. 2014, 28, 2128–2133. [Google Scholar] [CrossRef]
- Kumar, A.; Memo, M.; Mastinu, A. Plant behaviour: An evolutionary response to the environment? Plant Biol. 2020, 22, 961–970. [Google Scholar] [CrossRef] [PubMed]
- Teerarak, M.; Laosinwattana, C.; Charoenying, P. Evaluation of allelopathic, decomposition and cytogenetic activities of Jasminum officinale L. F. var. grandiflorum (L.) Kob. on bioassay plants. Biores. Technol. 2010, 101, 5677–5684. [Google Scholar] [CrossRef]
- Inderjit; Wardle, D.A.; Karban, R.; Callaway, R.M. The ecosystem and evolutionary contexts of allelopathy. Trends Ecol. Evol. 2011, 26, 655–662. [Google Scholar] [CrossRef]
- Hamayun, M.; Afzal Khan, S.; Ahmad, N. Allelopathic effects of Cyperus rotunds and Echinochloa crusgalli on seed germination and plumule and radical growth in maize (Zea mays L.). Pak. J. Weed Sci. Res. 2005, 11, 81–84. [Google Scholar]
- Hossain, M.K.; Alam, M.N. Allelopathic effects of Lantana camara leaf extract on germination and growth behavior of some agricultural and forest crops in Bangladesh. Pak. J. Weed Sci. Res. 2010, 16, 217–226. [Google Scholar]
- Khan, A.; Mehmood, S.; ullah Khan, R.; ullah Khan, S.; Khan Sherwani, S. Phytotoxic effect of Rosa bruonii extracts on germination of Cucumus sativus and Helianthus annus. IJAPR 2013, 7, 2032–2037. [Google Scholar]
- Netsere, A.; Mendesil, E. Allelopathic effects of Parthenium hysterophorus L. aqueous extracts on soybean (Glycine max L.) and haricot bean (Phaseolus vulgaris L.) seed germination, shoot and root growth and dry matter production. J. App. Bot. Food Qual. 2011, 84, 219–222. [Google Scholar]
- Mohammadkhani, N.; Zademobarak, M. Evaluation of allelopathic effect of Rosa damascena Mill Boukan and Khansar accessions extract on seed germination of wheat Zarrin cultivar. Iran. J. Seed Sci. Res. 2018, 5, 87–97. [Google Scholar] [CrossRef]
- Callaway, R.M.; Ridenour, W.M.; Laboski, T.; Weir, T.; Vivanco, J.M. Natural selection for resistance to the allelopathic effects of invasive plants. J. Ecol. 2005, 93, 576–583. [Google Scholar] [CrossRef]
- Mazur, A. The role of seed coat in the germination and early stages of growth of bean (Phaseolus vulgaris L.) in the presence of chickweed (Stellaria media (L.) Vill.). AUPC Stud. Nat. 2019, 4, 103–118. [Google Scholar] [CrossRef]
- Tomaszewski, M.; Thimann, K.V. Interactions of phenolic acids, metallic ions and chelating agents on auxin induced growth. Plant Physiol. 1966, 41, 1443–1454. [Google Scholar] [CrossRef]
- Muscolo, A.; Panuccio, M.; Sidari, M. The effect of phenols on respiratory enzymes in seed germination. Plant Growth Regul. 2001, 35, 31–35. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. (Eds.) Respiration and lipid metabolism. In Plant Physiology, 3rd ed.; Sinauer Associates: Sunderland, UK, 2002; pp. 223–258. [Google Scholar]
- Zohaib, A.; Ehsanullah; Tabassum, T.; Abbas, T. Influence of water soluble phenolics of Vicia sativa L. on germination and seedling growth of pulse crops. Sci. Agric. 2014, 8, 148–151. [Google Scholar] [CrossRef]
- Li, Z.H.; Wang, Q.; Ruan, X.; Pan, C.D.; Jiang, D.A. Phenolics and plant allelopathy. Molecules 2010, 15, 8933–8952. [Google Scholar] [CrossRef]
- Sisodia, S.; Siddiqui, M.B. Allelopathic effect by aqueous extracts of different parts of Croton bonplandianum Baill. on some crop and weed plants. J. Agric. Ext. Rural Dev. 2010, 2, 22–28. [Google Scholar]
- Inderjit; Duke, S.O. Ecophysiological aspects of allelopathy. Planta 2003, 217, 529–539. [Google Scholar] [CrossRef]
- Croteau, R.; Kutchan, T.M.; Lewis, N.G. Natural products (secondary metabolites). In Biochemistry and Molecular Biology of Plants; Buchanan, B., Gruissem, W., Joneas, R., Eds.; American Society of Plant Biologists: Rockville, MD, USA, 2000; pp. 1250–1268. [Google Scholar]
- Silva, C.B.; Simionatto, E.; Gebara, S.S.; Ré-Poppi, N.; Rech, K.S.; Peres, M.T.L.P.; Santos, G.O.; Dias, J.F.G.; Zanin, S.M.W.; Miguel, O.G.; et al. Sensitivity of Brachiaria decumbens and Ipomoea cordifolia to cyclic polysulfides from leaves of Microlobius foetidus. Allelop. J. 2014, 33, 213–226. [Google Scholar]
- Ma, H.; Chen, Y.; Chen, J.; Zang, J.; Zang, T.; He, H. Comparison of allelopathic effects of two typical invasive plants: Mikania micrantha and Ipomoea cairica in Hainan island. Sci. Rep. 2020, 10, 11332. [Google Scholar] [CrossRef]
- Ding, J.; Sun, Y.; Xiao, C.L.; Shi, K.; Zhou, Y.H.; Yu, J.Q. Physiological basis of different allelopathic reactions of cucumber and figf gourd plants to cinnamic acid. J. Exp. Bot. 2007, 58, 3765–3773. [Google Scholar] [CrossRef]
- Macias, F.A.; Galindo, J.C.G.; Molinillo, J.M.G.; Cutler, H.G. Allelopathy: Chemistry and Mode of Action of Allelochemicals; CRC Press: Boca Raton, FL, USA, 2003; p. 392. [Google Scholar] [CrossRef]
- Belz, R.G.; Reinhardt, C.F.; Foxcroft, L.C.; Karl, H. Residual allelopathy in Parthenium hysterophorus L. Does parthenin play a leading role? Crop Protect. 2007, 26, 237–245. [Google Scholar] [CrossRef]
- Fujii, Y.; Hiradate, S. Allelopathy: New Concepts Methodology; Science Publishers: Enfield, UK, 2007; p. 382. [Google Scholar]
- Blum, U. Allelopathic interactions involving phenolic acids. J. Nematol. 1996, 28, 259–267. [Google Scholar] [PubMed]
- Einhellig, F.A. Allelopathy—Current status and future goals. Allelopathy 1995, 582, 1–24. [Google Scholar] [CrossRef]
- Einhellig, F.A. Mechanism of action of allelochemicals in allelopathy. Allelopathy 1995, 582, 96–116. [Google Scholar] [CrossRef]
- Aldrich, R.J. (Ed.) Allelopatia w kierowaniu odchwaszczeniem. In Ekologia Chwastów w Roślinach Uprawnych. Podstawy Zwalczania Chwastów; Towarzystwo Chemii i Inżynierii Ekologicznej “Solpress”: Opolo, Poland, 1997; pp. 207–231. [Google Scholar]
- Blum, U.; Gerig, T.M.; Worsham, A.D.; King, L.D. Modification of the allelopatic effects of p-kumaric acid on morning-glory seedlings biomass by glucose, methionine and nitrate. J. Chem. Ecol. 1993, 19, 2791–2811. [Google Scholar] [CrossRef] [PubMed]
- Inderjit; Callaway, R.M. Experimental designs for the study of allelopathy. Plant Soil 2003, 256, 1–11. [Google Scholar] [CrossRef]
- Scognamiglio, M.; Fiumano, V.; D’abrosca, B.; Esposito, A.; Choi, Y.H.; Verpoorte, R.; Fiorentino, A. Chemical interactions between plants in Mediterranean vegetation: The influence of selected plant extracts on Aegilops geniculata metabolome. Phytochemistry 2014, 106, 69–85. [Google Scholar] [CrossRef]
- Kah, M.; Brown, C.D. Adsorption of ionizable pesticides in soil. Rev. Environ. Contam. Toxicol. 2006, 188, 149–217. [Google Scholar] [CrossRef]
- Weir, T.L.; Park, S.W.; Vivanco, J.M. Biochemical and physiological mechanisms mediated by allelochemicals. Curr. Opin. Plant Biol. 2004, 7, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Samedani, B.; Juraimi, A.S.; Rafii, M.Y.; Anuar, A.R.; Sheikh Awadz, S.A.; Anwar, M.P. Allelopathic effects of litter Axonopus compressus against two weedy species and its persistence in soil. Sci. World J. 2013, 2013, 695404. [Google Scholar] [CrossRef]
- Gronle, A.; Heb, J.; Böhm, H. Weed suppressive ability in sole and intercrops of pea and oat and it is interaction with ploughing and crop interference in organic farming. Org. Agric. 2015, 5, 39–51. [Google Scholar] [CrossRef]
- Mercure, M.; Bruneau, A. Hybridization between the escaped Rosa rugosa (Rosaceae) and native R. blanda in eastern North America. Syst. Phytogeogr. 2008, 95, 597–607. [Google Scholar] [CrossRef]
- Muhammad, Z.; Majeed, A. Allelopathic effects of aqueous extracts of sunflower on wheat (Triticum aestivum L.) and maize (Zea mays L.). Pak. J. Bot. 2014, 46, 1715–1718. [Google Scholar]
- Islam, A.K.M.A.; Anuar, N.; Yaakob, Z. Effect of genotypes and pre-sowing treatments on seed germination behavior of Jatropha. Asian J. Plant Sci. 2009, 8, 433–439. [Google Scholar] [CrossRef]
- Chiapusio, G.; Sanchez, A.M.; Reigosa, M.J.; Gonzalez, L.; Pellissier, F. Do germination indices adequately reflect allelochemical effects on the germination process? J. Chem. Ecol. 1997, 23, 2445–2453. [Google Scholar] [CrossRef]
- Khan, R.A.; Khan, F.; Ahmed, M.; Shah, A.S.; Khan, N.A.; Khan, M.R.; Shah, M.S. Phytotoxic and antibacterial extract of Mentha longifolia (Linn.). Afr. J. Pharm. Pharmacol. 2011, 5, 1530–1533. [Google Scholar] [CrossRef]
- Gao, X.; Li, M.; Gao, Z.; Li, C.; Sun, Z. Allelopathic effects of Hemistepta lyrate on the germination and growth of wheat, sorghum, cucumber, rape, and radish seeds. Weed Biol. Manag. 2009, 9, 243–249. [Google Scholar] [CrossRef]
- Mominul Islam, A.K.M.; Kato-Noguchi, H. Allelopathic potentiality of medicinal plant Leucas aspera. Int. J. Agric. Sustain. 2012, 4, 1–7. [Google Scholar]
- Lipniak, K.; Kliszcz, A. Allelopathic effect of goosefoot on germination and early stage growth of triticale and radish. AUPC Stud. Nat. 2020, 5, 110–128. [Google Scholar] [CrossRef]
- Szafraniec, R.; Możdżeń, K.; Barabasz-Krasny, B.; Zandi, P.; Wang, Y. Influence of volatile peppermint (Mentha × piperita L.) compounds on germination and seedling of radish (Raphanus sativus L. var. radicula Pers.) growth. Not. Bot. Horti Agrobot. 2019, 47, 1277–1284. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract Types (%) | CRG (%) | MGT (Day) | SVI (a.u.) | GI (%) | ||||
|---|---|---|---|---|---|---|---|---|
| A | B | A | B | A | B | A | B | |
| Root | ||||||||
| Control | 20.22 a ±1.24 | 23.94 a ±0.43 | 4.96 c ±0.30 | 4.18 b ±0.07 | 71.6 a ±15.61 | 129.7 a ±42.17 | 22.55 a ±6.26 | 53.70 a ±3.17 |
| 1 | 18.51 b ±0.25 | 23.53 ab ±0.05 | 5.40 b ±0.07 | 4.25 ab ±0.01 | 52.4 b ±11.07 | 108.0 a ±33.05 | 13.52 b ±0.68 | 49.40 ab ±2.35 |
| 2.5 | 17.55 b ±0.61 | 23.27 b ±0.07 | 5.70 ab ±0.20 | 4.30 a ±0.01 | 43.5 b ±9.18 | 133.2 a ±21.09 | 9.67 b ±5.38 | 49.32 ab ±0.58 |
| 5 | 17.13 b ±0.67 | 23.09 b ±0.16 | 5.84 a ±0.22 | 4.33 a ±0.03 | 24.1 c ±5.20 | 69.2 b ±14.21 | 4.46 c ±1.15 | 43.16 b ±4.00 |
| F | 10.10 | 3.80 | 20.87 | 5.22 | 36.88 | 8.64 | 8.32 | 8.45 |
| p | 0.01 * | 0.04 * | 0.000 * | 0.02 * | 0.000 * | 0.000 * | 0.002 * | 0.003 * |
| Stalk | ||||||||
| Control | 20.22 a ±1.24 | 23.94 a ±0.43 | 4.96 a ±0.30 | 4.18 b ±0.07 | 71.6 a ±15.61 | 129.7 a ±42.17 | 22.55 b ±6.26 | 53.70 a ±3.17 |
| 1 | 21.80 a ±0.81 | 23.61 a ±0.51 | 4.59 a ±0.17 | 4.24 b ±0.09 | 57.5 b ±11.75 | 115.4 a ±34.79 | 30.57 a ±6.00 | 48.69 a ±2.55 |
| 2.5 | 20.65 a ±0.19 | 23.15 b ±0.33 | 4.84 a ±0.05 | 4.32 a ±0.06 | 32.7 bc ±8.50 | 85.2 b ±21.18 | 15.28 c ±1.24 | 44.86 a ±3.26 |
| 5 | 20.23 a ±1.22 | 23.20 b ±0.12 | 4.96 a ±0.30 | 4.31 a ±0.02 | 25.0 c ±6.50 | 63.7 b ±11.23 | 13.22 c ±3.14 | 45.16 a ±0.75 |
| F | 2.63 | 3.54 | 2.07 | 4.40 | 42.25 | 10.89 | 5.94 | 7.07 |
| p | 0.09 | 0.03 * | 0.15 | 0.03 * | 0.000 * | 0.000 * | 0.008 * | 0.006 * |
| Leaf | ||||||||
| Control | 20.22 a ±1.24 | 23.94 a ±0.43 | 4.96 b ±0.30 | 4.18 c ±0.07 | 71.6 a ±15.61 | 129.7 a ±42.17 | 22.55 a ±6.26 | 53.70 a ±3.17 |
| 1 | 16.91 b ±0.53 | 23.53 ab ±0.46 | 5.92 a ±0.19 | 4.25 bc ±0.08 | 40.1 b ±9.19 | 58.5 b ±9.70 | 7.28 b ±0.69 | 49.80 a ±3.40 |
| 2.5 | 16.66 b ±0.91 | 22.75 bc ±0.70 | 6.02 a ±0.34 | 4.40 ab ±0.14 | 16.3 c ±5.19 | 51.4 b ±11.29 | 3.56 c ±0.77 | 39.34 b ±7.36 |
| 5 | 16.90 b ±0.83 | 22.15 c ±0.23 | 5.93 a ±0.29 | 4.52 a ±0.05 | 9.4 c ±2.83 | 36.6 b ±4.55 | 2.53 c ±0.72 | 27.82 c ±6.03 |
| F | 12.27 | 6.41 | 12.27 | 7.82 | 93.64 | 34.92 | 16.38 | 23.59 |
| p | 0.000 * | 0.009 * | 0.000 * | 0.004 * | 0.000 * | 0.000 * | 0.000 * | 0.000 * |
| Flower | ||||||||
| Control | 20.22 ab ±1.24 | 23.94 a ±0.43 | 4.96 ab ±0.30 | 4.18 c ±0.07 | 71.6 a ±15.61 | 129.7 a ±42.17 | 22.55 b ±6.26 | 53.70 a ±3.17 |
| 1 | 22.23 a ±0.38 | 22.23 b ±0.07 | 4.50 b ±0.08 | 4.50 b ±0.01 | 53.1 b ±8.62 | 67.3 b ±13.14 | 29.42 ab ±6.79 | 40.63 b ±0.46 |
| 2.5 | 21.92 a ±0.75 | 22.08 b ±0.14 | 4.57 b ±0.15 | 4.53 ab ±0.03 | 17.1 c ±5.51 | 10.3 c ±4.01 | 12.66 c ±3.48 | 33.40 c ±3.77 |
| 5 | 19.24 b ±1.83 | 21.22 b ±1.13 | 5.23 a ±0.47 | 4. 72 a ±0.25 | 4.5 d ±1.44 | 1.5 c ±0.37 | 2.21 d ±3.19 | 6.69 d ±4.36 |
| F | 5.95 | 14.83 | 5.95 | 11.45 | 106.9 | 65.04 | 12.44 | 109.3 |
| p | 0.008 * | 0.000 * | 0.008 * | 0.001 * | 0.00 * | 0.000 * | 0.000 * | 0.000 * |
| Extract Type (%) | Fresh Mass (g) | Dry Mass (g) | Dry Mass/Fresh Mass (a.u.) | Total Water Content (%) | ||||
|---|---|---|---|---|---|---|---|---|
| A | B | A | B | A | B | A | B | |
| Root | ||||||||
| Control | 0.0075 a ±0.001 | 0.0874 b ±0.020 | 0.0003 b ±0.0003 | 0.0049 a ±0.001 | 0.0443 b ±0.046 | 0.0560 a ±0.009 | 95.57 a ±4.61 | 94.40 b ±0.87 |
| 1 | 0.0072 a ±0.002 | 0.1219 a ±0.032 | 0.0008 a ±0.0005 | 0.0047 a ±0.001 | 0.1124 a ±0.073 | 0.0396 b ±0.009 | 88.76 b ±7.29 | 96.04 a ±0.95 |
| 2.5 | 0.0074 a ±0.002 | 0.1145 ab ±0.032 | 0.0008 a ±0.0004 | 0.0058 a ±0.001 | 0.1139 a ±0.065 | 0.0519 a ±0.010 | 88.61 b ±6.50 | 94.81 b ±1.01 |
| 5 | 0.0062 a ±0.001 | 0.1027 ab ±0.030 | 0.0006 ab ±0.0003 | 0.0051 a ±0.002 | 0.0994 a ±0.046 | 0.0489 a ±0.009 | 90.06 b ±4.60 | 95.11 b ±0.95 |
| F | 1.74 | 2.65 | 3.68 | 0.94 | 3.12 | 5.10 | 3.12 | 5.10 |
| p | 0.18 | 0.06 | 0.021 * | 0.43 | 0.038 * | 0.005 * | 0.038 * | 0.005 * |
| Stalk | ||||||||
| Control | 0.0075 a ±0.001 | 0.0874 c ±0.020 | 0.00003 b ±0.0003 | 0.0049 c ±0.001 | 0.0443 b ±0.046 | 0.0560 bc ±0.009 | 95.57 a ±4.61 | 94.40 ab ±0.87 |
| 1 | 0.0074 a ±0.001 | 0.1280 a ±0.037 | 0.0005 ab ±0.003 | 0.0061 bc ±0.001 | 0.0692 ab ±0.044 | 0.0490 c ±0.009 | 93.08 ab ±4.40 | 95.10 a ±0.93 |
| 2.5 | 0.0072 a ±0.002 | 0.1011 bc ±0.018 | 0.0005 ab ±0.0004 | 0.0067 ab ±0.001 | 0.0747 ab ±0.056 | 0.0667 a ±0.013 | 92.53 ab ±5.59 | 93.32 c ±1.27 |
| 5 | 0.0074 a ±0.002 | 0.1200 ab ±0.031 | 0.0008 a ±0.0003 | 0.0075 a ±0.001 | 0.0980 a ±0.039 | 0.0640 ab ±0.011 | 90.20 b ±3.91 | 93.60 bc ±1.12 |
| F | 0.14 | 3.84 | 2.70 | 6.04 | 2.23 | 5.05 | 2.23 | 5.05 |
| p | 0.94 | 0.018 * | 0.01 * | 0.002 * | 0.02 * | 0.005 * | 0.02 * | 0.005 * |
| Leaf | ||||||||
| Control | 0.0075 a ±0.001 | 0.0874 a ±0.020 | 0.0003 a ±0.0003 | 0.0049 b ±0.001 | 0.0443 b ±0.046 | 0.0560 b ±0.009 | 95.57 a ±4.61 | 94.40 a ±0.87 |
| 1 | 0.0065 b ±0.001 | 0.0467 b ±0.013 | 0.0006 a ±0.0003 | 0.0058 ab ±0.002 | 0.0839 b ±0.051 | 0.1239 a ±0.017 | 91.61 a ±5.08 | 87.61 b ±1.74 |
| 2.5 | 0.0064 b ±0.001 | 0.0530 b ±0.008 | 0.0005 a ±0.0004 | 0.0061 ab ±0.001 | 0.0732 b ±0.070 | 0.1175 a ±0.031 | 92.68 a ±7.01 | 88.25 b ±3.11 |
| 5 | 0.0037 c ±0.001 | 0.0561 b ±0.011 | 0.0006 a ±0.0004 | 0.0063 a ±0.001 | 0.1669 a ±0.099 | 0.1156 a ±0.031 | 83.31 b ±9.94 | 88.44 b ±3.09 |
| F | 2.27 | 1.28 | 1.26 | 1.92 | 5.68 | 1.58 | 5.68 | 1.58 |
| p | 0.000 * | 0.000 * | 0.32 | 0.15 | 0.003 * | 0.000 * | 0.003 * | 0.000 * |
| Flower | ||||||||
| Control | 0.0075 a ±0.001 | 0.0874 b ±0.020 | 0.0003 b ±0.0003 | 0.0049 b ±0.001 | 0.0443 b ±0.046 | 0.0560 c ±0.009 | 95.57 a ±4.61 | 94.40 a ±0.87 |
| 1 | 0.0071 a ±0.001 | 0.1041 a ±0.015 | 0.0005 ab ±0.0003 | 0.0063 b ±0.002 | 0.0746 ab ±0.037 | 0.0617 c ±0.018 | 92.54 ab ±3.71 | 93.83 a ±1.77 |
| 2.5 | 0.0078 a ±0.001 | 0.0670 c ±0.010 | 0.0007 a ±0.0002 | 0.0094 a ±0.002 | 0.0875 a ±0.030 | 0.1434 b ±0.037 | 91.25 b ±2.95 | 85.66 b ±3.66 |
| 5 | 0.0070 a ±0.002 | 0.0294 d ±0.011 | 0.0004 b ±0.0003 | 0.0085 a ±0.003 | 0.0579 ab ±0.040 | 0.2980 a ±0.071 | 94.21 a ±4.03 | 70.20 c ±7.06 |
| F | 0.65 | 4.92 | 3.01 | 9.63 | 2.38 | 6.36 | 2.38 | 6.36 |
| p | 0.59 | 0.000 * | 0.04 * | 0.000 * | 0.03 * | 0.000 * | 0.03 * | 0.000 * |
| Extracts (%) | Root | Stalk | Leaf | Flower | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parameters | 1 | 2.5 | 5 | 1 | 2.5 | 5 | 1 | 2.5 | 5 | 1 | 2.5 | 5 | |
| GP | |||||||||||||
| CRG | |||||||||||||
| MGT | |||||||||||||
| SVI | |||||||||||||
| GI | |||||||||||||
| RI | |||||||||||||
| Length (whole seedling) | |||||||||||||
| FM | |||||||||||||
| DM | |||||||||||||
| DM/FM | |||||||||||||
| TWC | |||||||||||||
| EC | |||||||||||||
| Extracts (%) | Root | Stalk | Leaf | Flower | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parameters | 1 | 2.5 | 5 | 1 | 2.5 | 5 | 1 | 2.5 | 5 | 1 | 2.5 | 5 | |
| GP | |||||||||||||
| CRG | |||||||||||||
| MGT | |||||||||||||
| SVI | |||||||||||||
| GI | |||||||||||||
| RI | |||||||||||||
| Length (whole seedling) | |||||||||||||
| Fresh mass | |||||||||||||
| Dry mass | |||||||||||||
| DM/FM | |||||||||||||
| TWC | |||||||||||||
| EC | |||||||||||||
| Parts of Rosa | Root | Stalk | Leaves | Flowers | |
|---|---|---|---|---|---|
| Extracts [%] | |||||
| 1 | 4.99 a ±0.02 | 4.56 b ±0.01 | 4.12 c ±0.03 | 4.85 b ±0.01 | |
| 2.5 | 4.12 c ±0.01 | 4.47 c ±0.02 | 4.55 b ±0.01 | 4.86 b ±0.01 | |
| 5 | 4.55 b ±0.01 | 4.62 a ±0.01 | 4.86 a ±0.07 | 5.04 a ±0.01 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Możdżeń, K.; Tatoj, A.; Barabasz-Krasny, B.; Sołtys-Lelek, A.; Gruszka, W.; Zandi, P. The Allelopathic Potential of Rosa blanda Aiton on Selected Wild-Growing Native and Cultivated Plants in Europe. Plants 2021, 10, 1806. https://doi.org/10.3390/plants10091806
Możdżeń K, Tatoj A, Barabasz-Krasny B, Sołtys-Lelek A, Gruszka W, Zandi P. The Allelopathic Potential of Rosa blanda Aiton on Selected Wild-Growing Native and Cultivated Plants in Europe. Plants. 2021; 10(9):1806. https://doi.org/10.3390/plants10091806
Chicago/Turabian StyleMożdżeń, Katarzyna, Agnieszka Tatoj, Beata Barabasz-Krasny, Anna Sołtys-Lelek, Wojciech Gruszka, and Peiman Zandi. 2021. "The Allelopathic Potential of Rosa blanda Aiton on Selected Wild-Growing Native and Cultivated Plants in Europe" Plants 10, no. 9: 1806. https://doi.org/10.3390/plants10091806
APA StyleMożdżeń, K., Tatoj, A., Barabasz-Krasny, B., Sołtys-Lelek, A., Gruszka, W., & Zandi, P. (2021). The Allelopathic Potential of Rosa blanda Aiton on Selected Wild-Growing Native and Cultivated Plants in Europe. Plants, 10(9), 1806. https://doi.org/10.3390/plants10091806