
Ectomycorrhizal Inoculation Enhances the Salt Tolerance of Quercus mongolica Seedlings


Abstract
1. Introduction
2. Results
2.1. External Morphology and Anatomical Structure of ECM
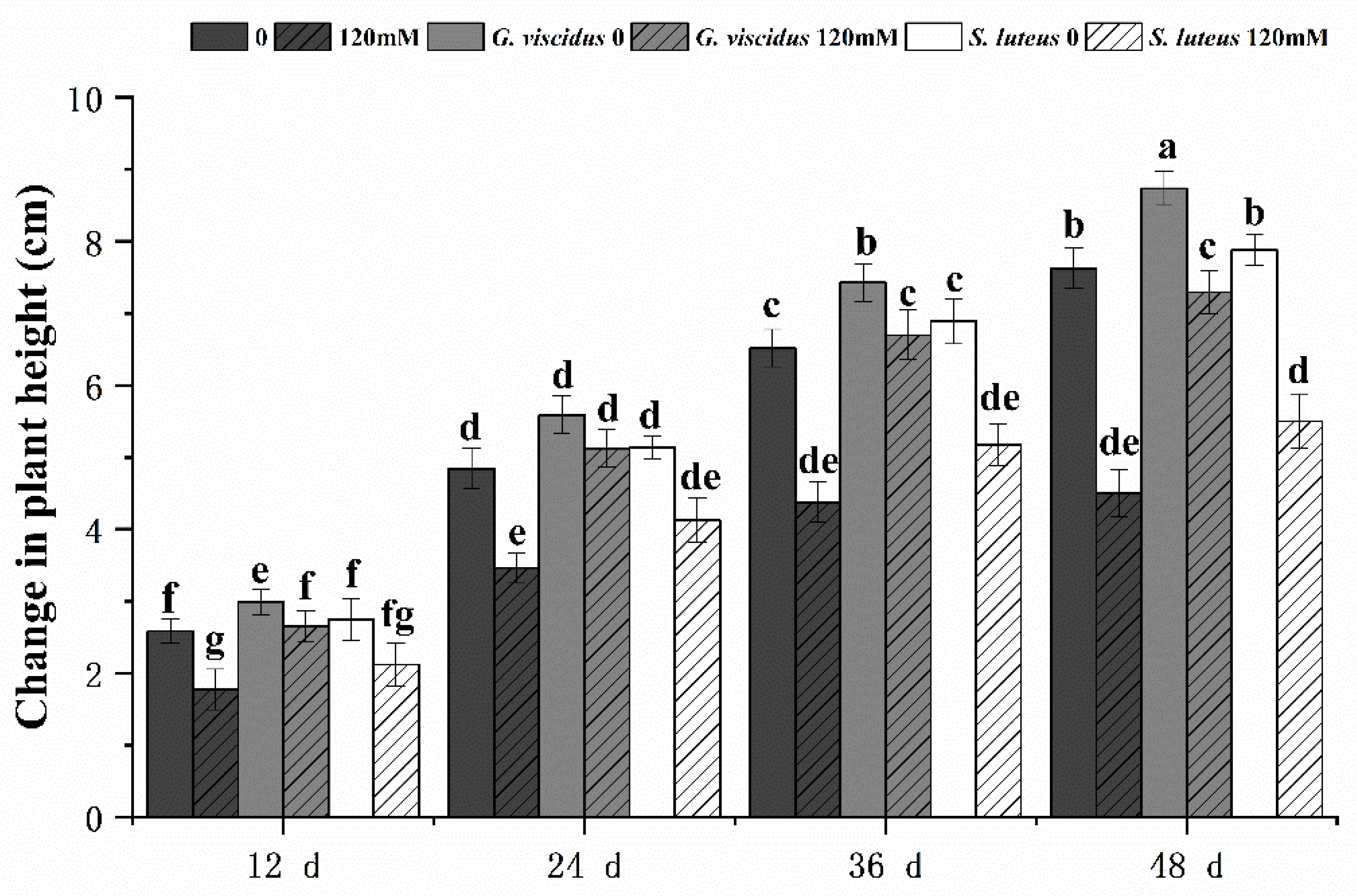
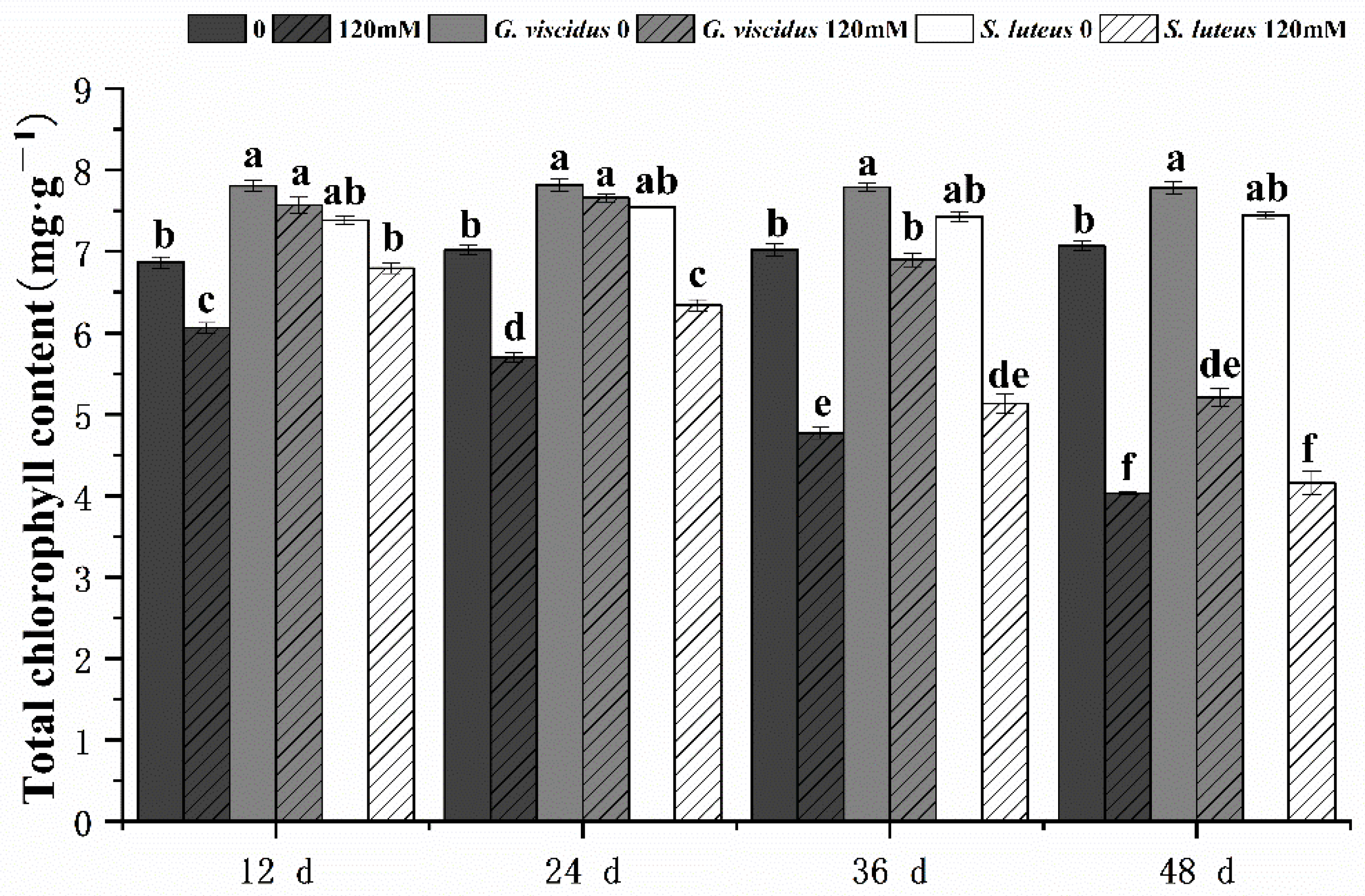
2.2. Effect of NaCl Treatment on Growth
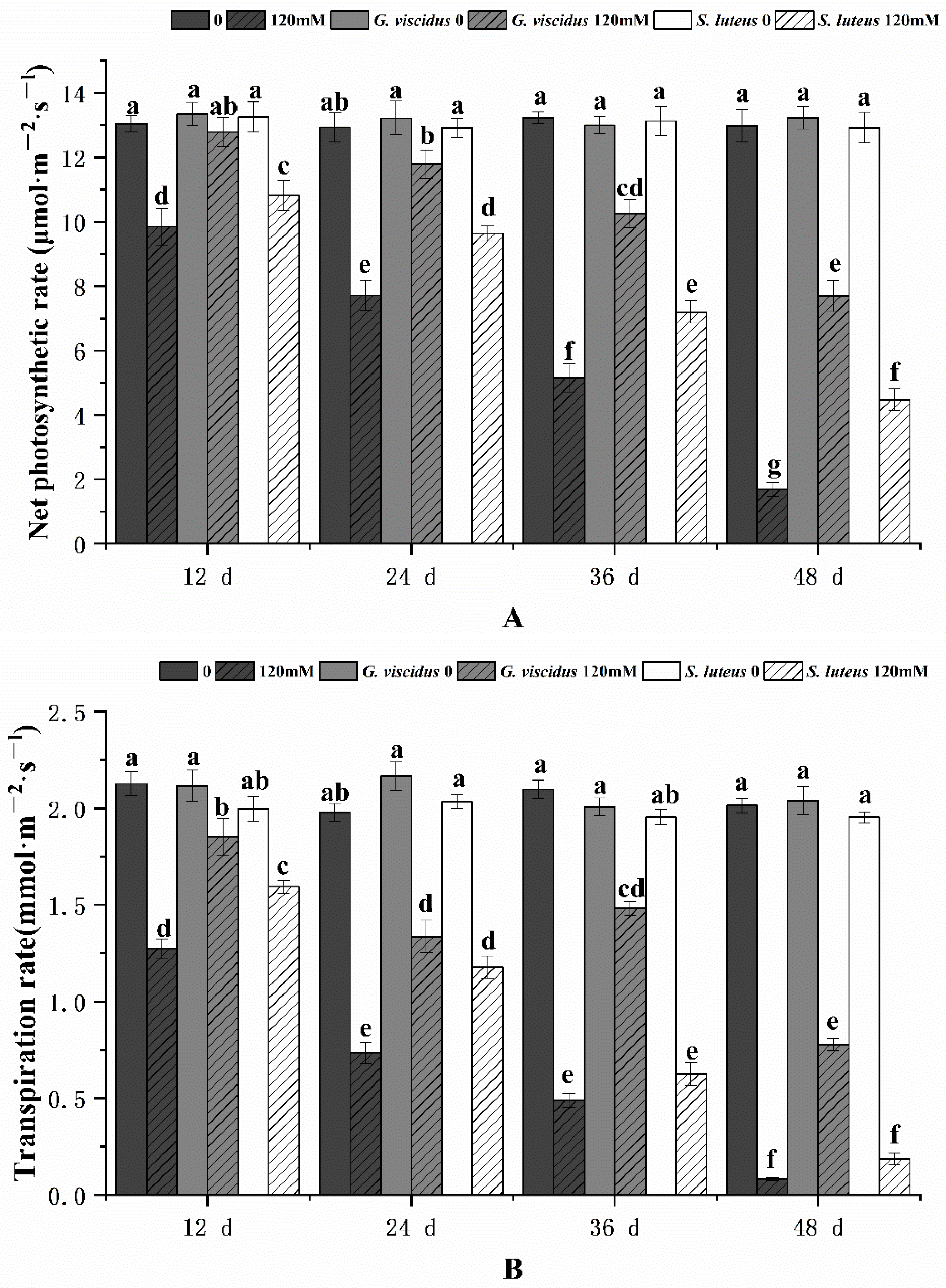
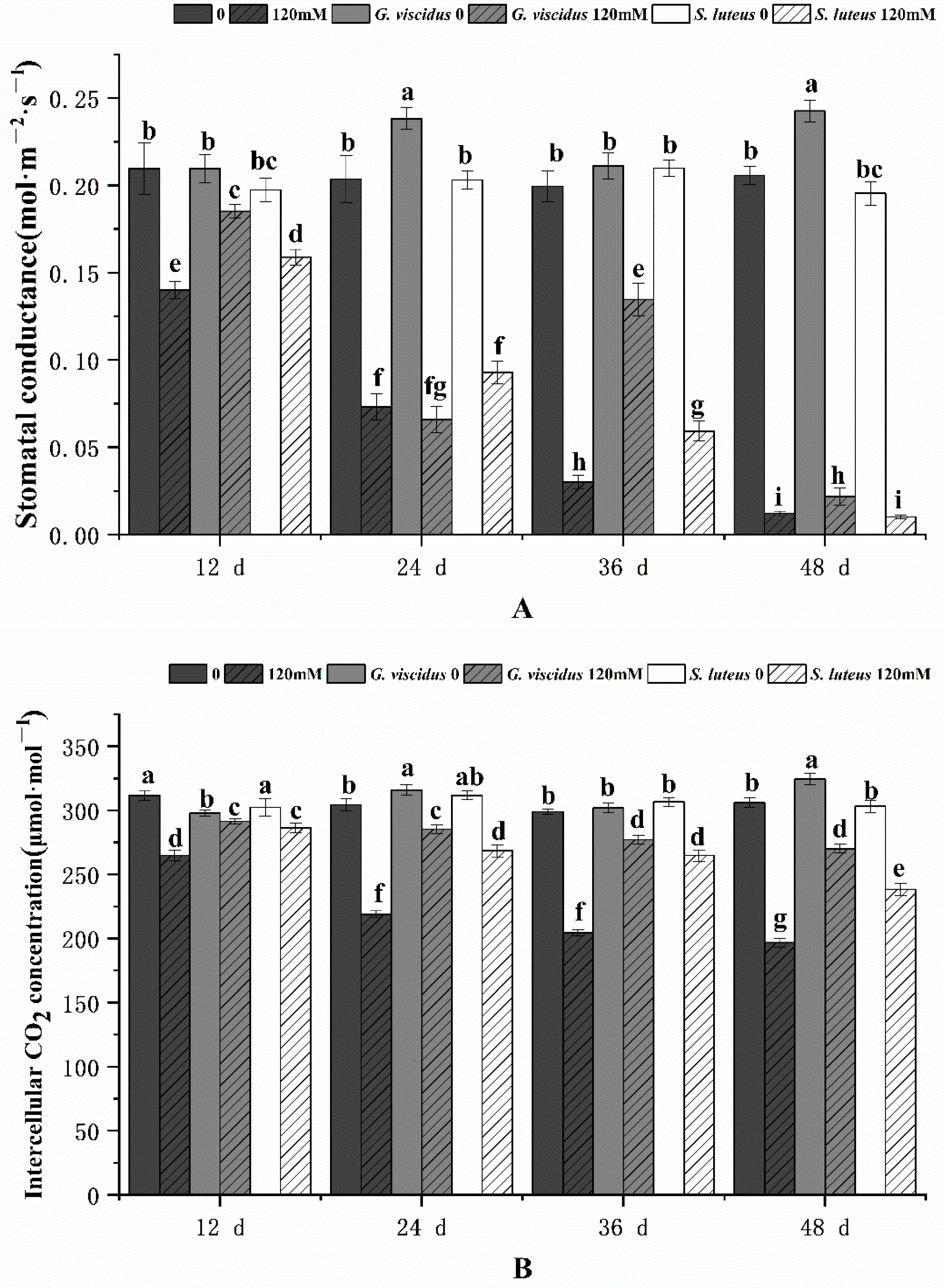
2.3. Effect of NaCl Treatment on Photosynthesis
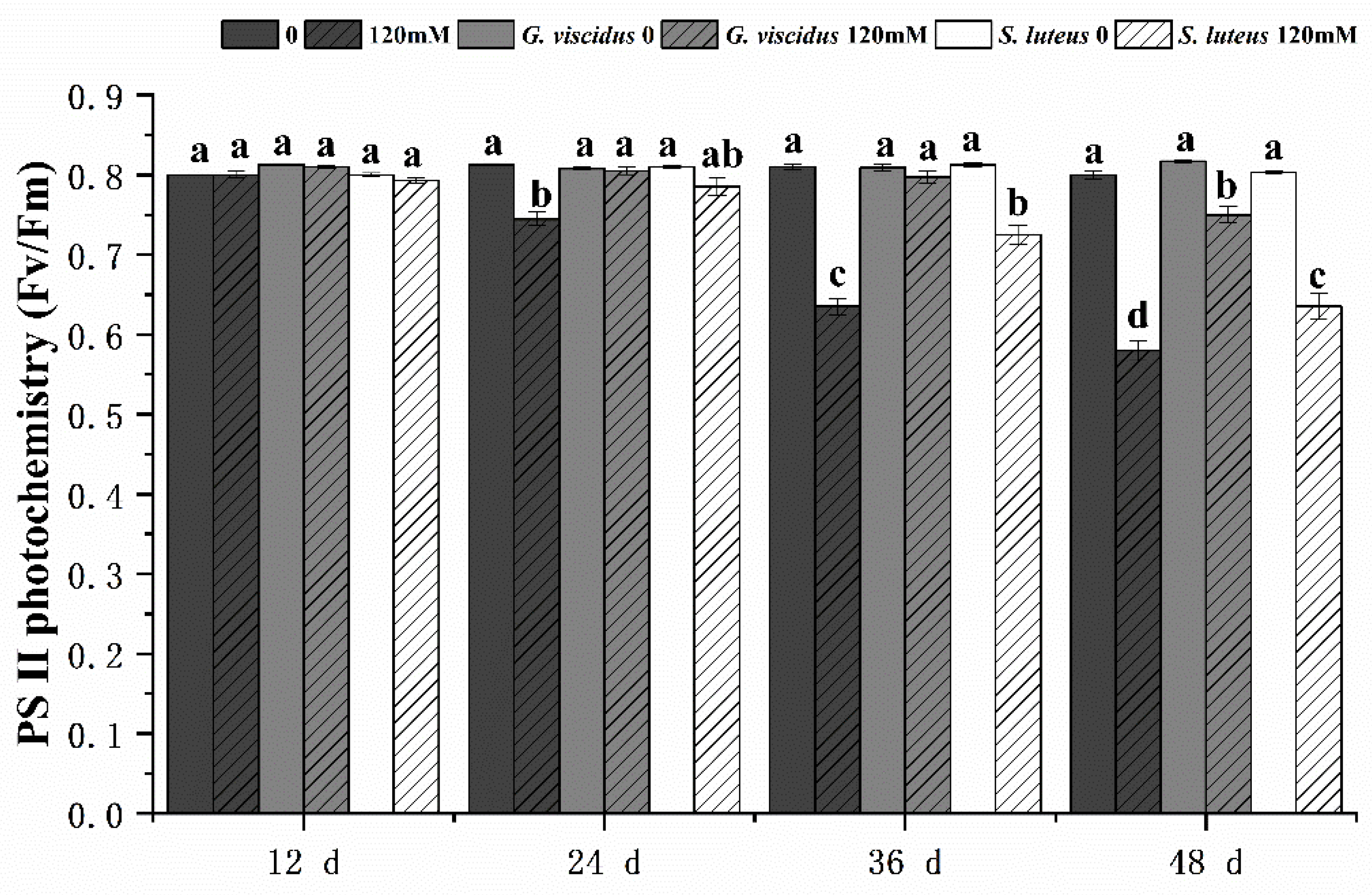
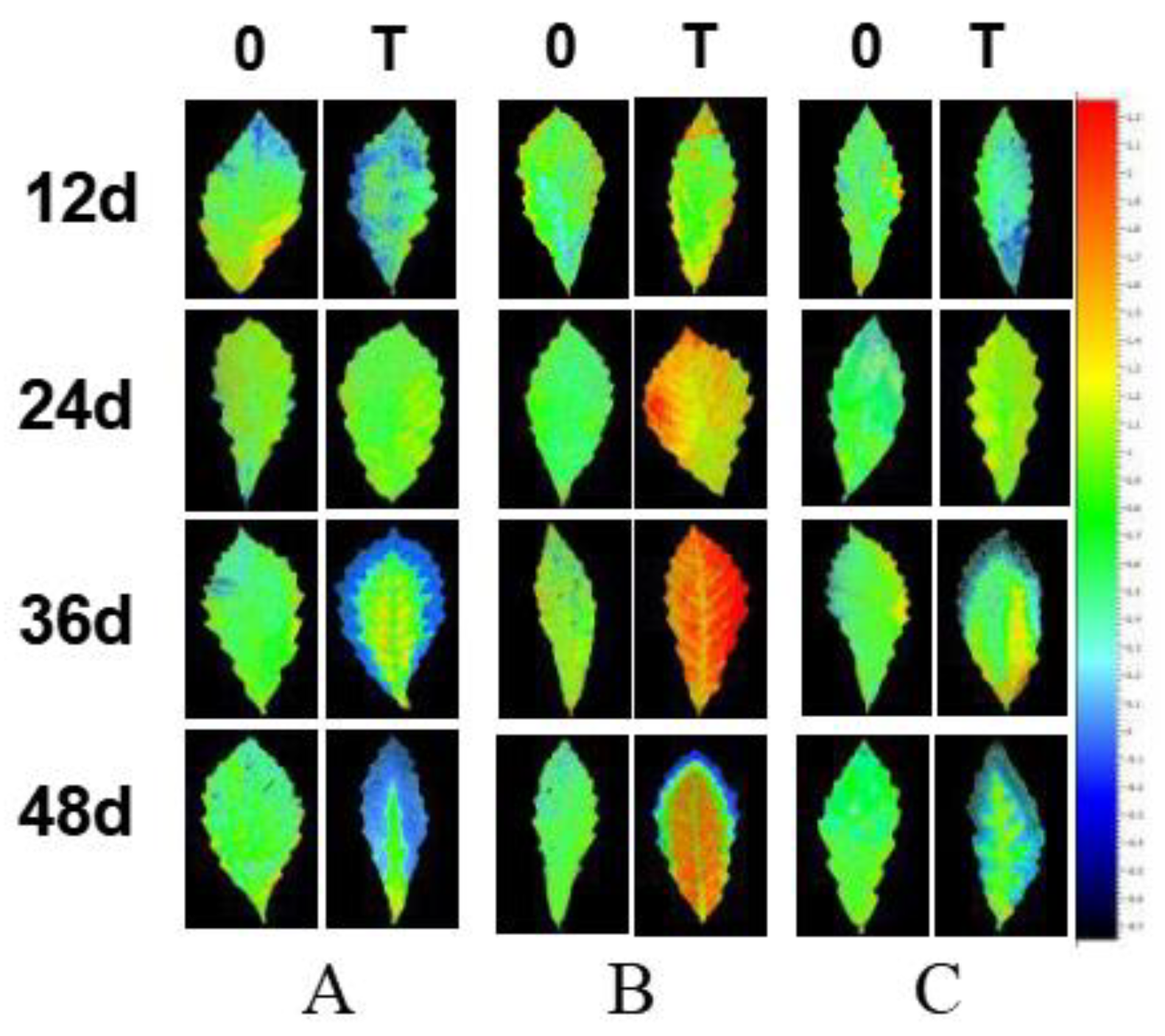
2.4. Effect of NaCl Treatment on Chlorophyll Fluorescence Parameters
2.5. Effect of NaCl Treatment on Cell Membrane Permeability
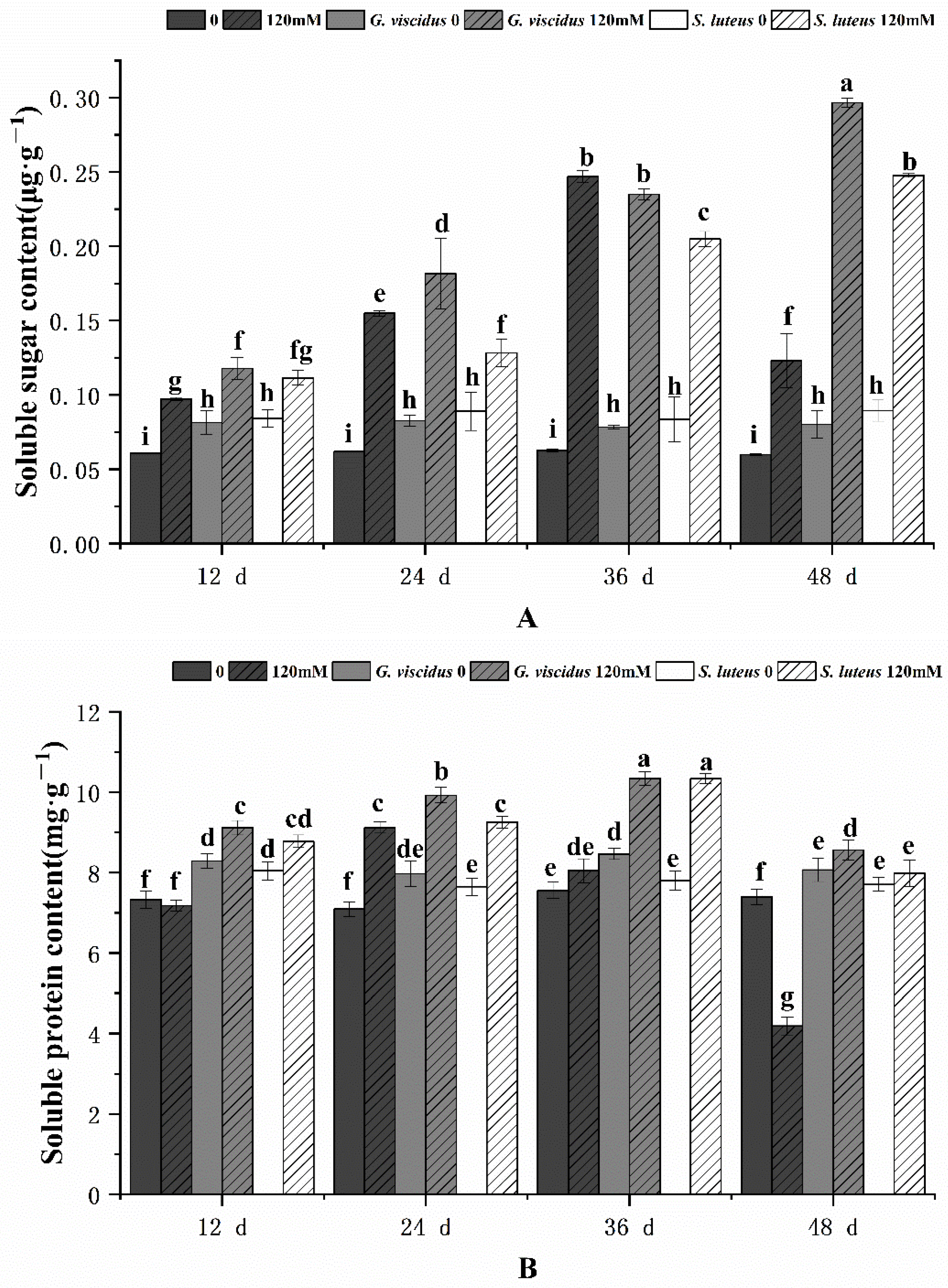
2.6. Effect of NaCl Treatment on Osmotic Adjustment Substances
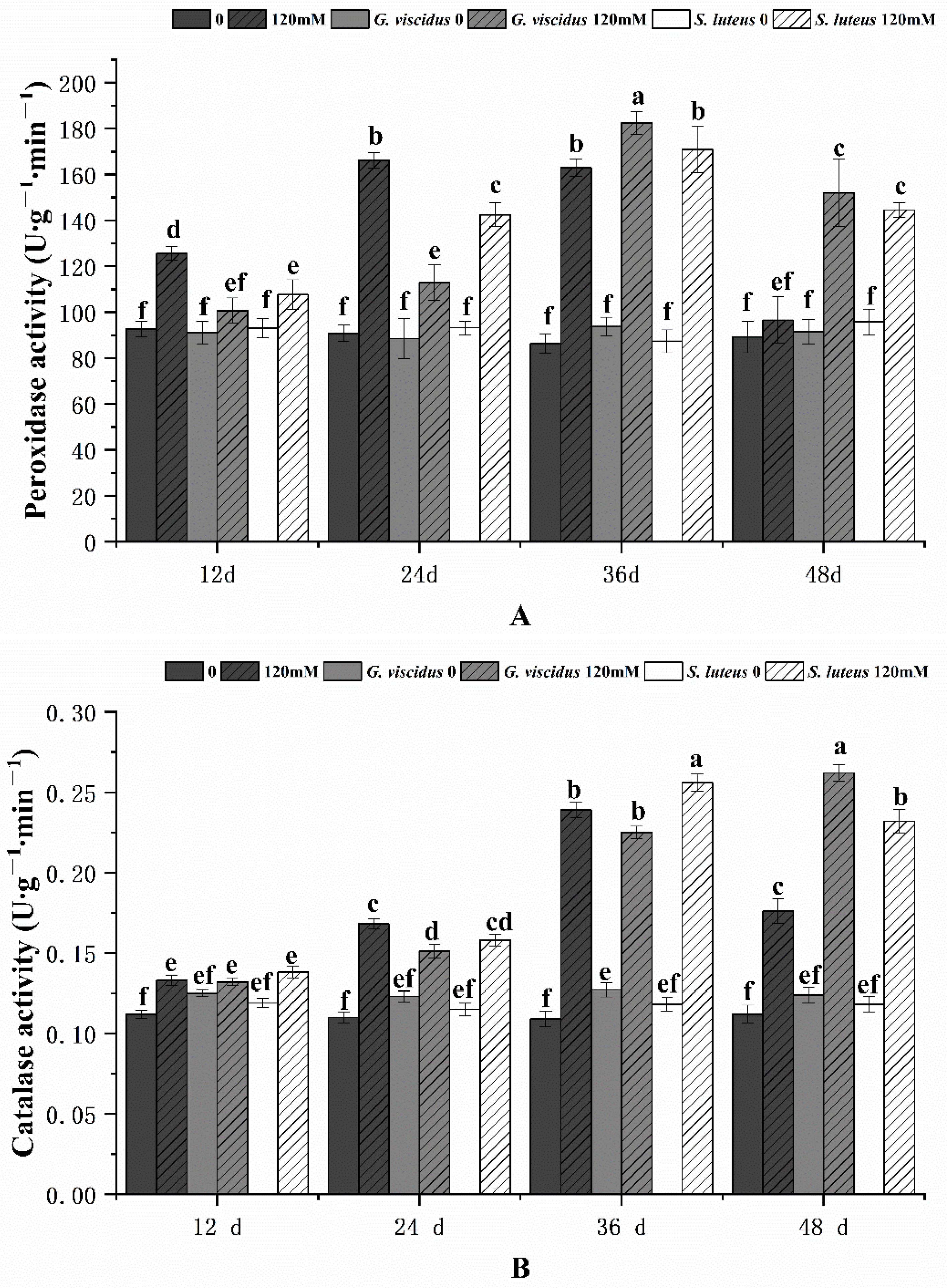
2.7. Effect of NaCl Treatment on the Antioxidant Enzyme System
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Preparation of Fungal Inoculant and Fungal Inoculation
4.3. Salinity Treatment
4.4. Morphological and Anatomical Structure Measurement
4.5. Physiological and Biochemical Indicators Measurement
4.6. Photosynthesis Index Measurements
4.7. Chlorophyll Fluorescence Parameter Measurements
4.8. Statistical Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Tavakkoli, E.; Rengasamy, P.; McDonald, G.K. High concentrations of Na+ and Cl− ions in soil solution have simultaneous detrimental effects on growth of faba bean under salinity stress. J. Exp. Bot. 2010, 61, 4449–4459. [Google Scholar] [CrossRef]
- Afzal, Z.; Howton, T.C.; Sun, Y.; Mukhtar, M.S. The roles of aquaporins in plant stress responses. J. Dev. Biol. 2016, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Verdoucq, L.; Maurel, C. Plant Aquaporins; Elsevier: Amsterdam, The Netherlands, 2018; pp. 25–56. [Google Scholar]
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Effect of salinity stress on plants and its tolerance strategies: A review. Environ. Sci. Pollut. Res. Int. 2015, 22, 4056–4075. [Google Scholar] [CrossRef] [PubMed]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Meyer, S.; Genty, B. Mapping intercellular CO2 mole fraction (Ci) in rosa rubiginosa leaves fed with abscisic acid by using chlorophyll fluorescence imaging. Significance of ci estimated from leaf gas exchange. Plant Physiol. 1998, 116, 947–957. [Google Scholar] [CrossRef] [PubMed]
- Negrão, S.; Schmöckel, S.M.; Tester, M. Evaluating physiological responses of plants to salinity stress. Ann. Bot. 2016, 119, 1–11. [Google Scholar] [CrossRef]
- Guerrero-Galan, C.; Calvo-Polanco, M.; Zimmermann, S.D. Ectomycorrhizal symbiosis helps plants to challenge salt stress conditions. Mycorrhiza 2019, 29, 291–301. [Google Scholar] [CrossRef]
- Nguyen, H.; Polanco, M.C.; Zwiazek, J.J. Gas exchange and growth responses of ectomycorrhizal Picea mariana, Picea glauca, and Pinus banksiana seedlings to NaCl and Na2SO4. Plant Biol. 2006, 8, 646–652. [Google Scholar] [CrossRef]
- Bois, G.; Bigras, F.J.; Bertrand, A.; Piché, Y.; Fung, M.Y.P.; Khasa, D.P. Ectomycorrhizal fungi affect the physiological responses of Picea glauca and Pinus banksiana seedlings exposed to an NaCl gradient. Tree Physiol. 2006, 26, 1185–1196. [Google Scholar] [CrossRef] [PubMed]
- Polanco, M.C.; Zwiazek, J.J.; Jones, M.D.; MacKinnon, M.D. Responses of mycorrhizal jack pine (Pinus banksiana) seedlings to NaCl and boron. Trees 2008, 22, 825–834. [Google Scholar] [CrossRef]
- Polanco, M.C.; Zwiazek, J.J.; Voicu, M.C. Responses of ectomycorrhizal American elm (Ulmus americana) seedlings to salinity and soil compaction. Plant Soil 2008, 308, 189–200. [Google Scholar] [CrossRef]
- Ma, X.; Sun, M.; Sa, G.; Zhang, Y.; Li, J.; Sun, J.; Shen, X.; Polle, A.; Chen, S. Ion fluxes in Paxillus involutus-inoculated roots of Populus × canescens under saline stress. Environ. Exp. Bot. 2014, 108, 99–108. [Google Scholar] [CrossRef]
- Sa, G.; Yao, J.; Deng, C.; Liu, J.; Zhang, Y.; Zhu, Z.; Zhang, Y.; Ma, X.; Zhao, R.; Lin, S.; et al. Amelioration of nitrate uptake under salt stress by ectomycorrhiza with and without a Hartig net. New Phytol. 2019, 222, 1951–1964. [Google Scholar] [CrossRef] [PubMed]
- Muhsin, T.M.; Zwiazek, J.J. Colonization with Hebeloma crustuliniforme increases water conductance and limits shoot sodium uptake in white spruce (Picea glauca) seedlings. Plant Soil 2002, 238, 217–225. [Google Scholar] [CrossRef]
- Zwiazek, J.J.; Equiza, M.A.; Karst, J.; Senorans, J.; Wartenbe, M.; Calvo-Polanco, M. Role of urban ectomycorrhizal fungi in improving the tolerance of lodgepole pine (Pinus contorta) seedlings to salt stress. Mycorrhiza 2019, 29, 303–312. [Google Scholar] [CrossRef]
- Otgonsuren, B.; Rewald, B.; Godbold, D.L.; Göransson, H. Ectomycorrhizal inoculation of Populus nigra modifies the response of absorptive root respiration and root surface enzyme activity to salinity stress. Flora 2016, 224, 123–129. [Google Scholar] [CrossRef]
- Smith, M.E.; Pfister, D.H. Tuberculate ectomycorrhizae of angiosperms: The interaction between Boletus rubropunctus (Boletaceae) and Quercus species (Fagaceae) in the United States and Mexico. Am. J. Bot. 2009, 96, 1665–1675. [Google Scholar] [CrossRef][Green Version]
- Jumpponen, A.; Jones, K.L.; Mattox, D.J.; Yaege, C. Massively parallel 454-sequencing of fungal communities in Quercus spp. ectomycorrhizas indicates seasonal dynamics in urban and rural sites. Mol. Ecol. 2010, 19 (Suppl. S1), 41–53. [Google Scholar] [CrossRef]
- Nardinia, A.; Salleoa, S.; Tyreeb, M.T.; Vertoveca, M. Influence of the ectomycorrhizas formed by Tuber melanosporum Vitt. on hydraulic conductance and water relations of Quercus ilex L. seedlings. Ann. For. Sci. 2000, 57, 305–312. [Google Scholar] [CrossRef]
- Rieske, L.K. Influence of symbiotic fungal colonization on oak seedling growth and suitability for insect herbivory. Environ. Entomol. 2001, 30, 849–854. [Google Scholar] [CrossRef][Green Version]
- Yan, X.; Wang, Q. Effects of ectomycorrhizal inoculation on the seedling growth of Quercus liaotungensis. Acta Phytoecol. Sin. 2002, 26, 701–707. [Google Scholar]
- Núñez, J.A.D.; Serrano, J.S.; Barreal, J.A.R.; González, J.A.S.d.O. The influence of mycorrhization with Tuber melanosporum in the afforestation of a Mediterranean site with Quercus ilex and Quercus faginea. For. Ecol. Manag. 2006, 231, 226–233. [Google Scholar] [CrossRef]
- Ling, Q. Research on symbiotical fungi species and ectomycorrhizae occurrence of Chestnut (Castanea mollissima BL.). J. Beijing Agric. Coll. 1995, 10, 71–75. [Google Scholar]
- Liu, H.; Chen, H.; Ding, G.; Li, K.; Wang, Y. Proteomic insight into the symbiotic relationship of Pinus massoniana Lamb and Suillus luteus towards developing Al-stress resistance. Life 2021, 11, 177. [Google Scholar] [CrossRef] [PubMed]
- Yin, D.; Deng, X.; Chet, I.; Song, R. Physiological responses of Pinus sylvestris var. Mongolica seedlings to the interaction between Suillus luteus and Trichoderma virens. China Acad. J. Electron. Publ. House 2014, 69, 334–342. [Google Scholar]
- Korhonen, A.; Lehto, T.; Heinonen, J.; Repo, T. Whole-plant frost hardiness of mycorrhizal (Hebeloma sp. or Suillus luteus) and non-mycorrhizal Scots pine seedlings. Tree Physiol. 2019, 39, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Dalong, M.; Luhe, W.; Guoting, Y.; Liqiang, M.; Chun, L. Growth response of Pinus densiflora seedlings inoculated with three indigenous ectomycorrhizal fungi in combination. Braz. J. Microbiol. 2011, 42, 1197–1203. [Google Scholar] [CrossRef] [PubMed]
- Henke, C.; Jung, E.-M.; Kothe, E. Hartig’ net formation of Tricholoma vaccinum-spruce ectomycorrhiza in hydroponic cultures. Environ. Sci. Pollut. Res. 2015, 22, 19394–19399. [Google Scholar] [CrossRef]
- Krause, K.; Henke, C.; Asiimwe, T.; Ulbricht, A.; Klemmer, S.; Schachtschabel, D.; Boland, W.; Kothe, E. Biosynthesis and secretion of indole-3-acetic acid and its morphological effects on Tricholoma vaccinum-spruce ectomycorrhiza. Appl. Environ. Microbiol. 2015, 81, 7003–7011. [Google Scholar] [CrossRef]
- Navarro-RoDenas, A.; Xu, H.; Kemppainen, M.; Pardo, A.G.; Zwiazek, J.J. Laccaria bicolor aquaporin LbAQP1 is required for Hartig net development in trembling aspen (Populus tremuloides). Plant Cell Environ. 2015, 38, 2475–2486. [Google Scholar] [CrossRef] [PubMed]
- Van Zelm, E.; Zhang, Y.; Testerink, C. Salt tolerance mechanisms of plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef]
- Kohler, J.; Caravaca, F.; Roldán, A. An AM fungus and a PGPR intensify the adverse effects of salinity on the stability of rhizosphere soil aggregates of Lactuca sativa. Soil Biol. Biochem. 2010, 42, 429–434. [Google Scholar] [CrossRef]
- Wen, Z.; Zhu, X.; Liu, C. Effects of two ectomycorrhizal fungi on growth of Pinus thunbergii seedlings planted in saline soil. China Acad. J. Electron. Publ. House 2019, 39, 22–27. [Google Scholar] [CrossRef]
- Shi, L.; Xue, J.; Liu, B.; Dong, P.; Wen, Z.; Shen, Z.; Chen, Y. Hydrogen ions and organic acids secreted by ectomycorrhizal fungi, Pisolithus sp1, are involved in the efficient removal of hexavalent chromium from waste water. Ecotoxicol. Environ. Saf. 2018, 161, 430–436. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Azcon, R. Hyphal contribution to water uptake in mycorrhizal plants as affected by the fungal species and water status. Physiol. Plant. 1995, 95, 472–478. [Google Scholar] [CrossRef]
- Zou, Y.N.; Srivastava, A.K.; Ni, Q.D.; Wu, Q.S. Disruption of mycorrhizal extraradical mycelium and changes in leaf water status and soil aggregate stability in rootbox-grown trifoliate orange. Front. Microbiol. 2015, 6, 203. [Google Scholar] [CrossRef]
- Alvarez, M.; Huygens, D.; Fernandez, C.; Gacitua, Y.; Olivares, E.; Saavedra, I.; Alberdi, M.; Valenzuela, E. Effect of ectomycorrhizal colonization and drought on reactive oxygen species metabolism of Nothofagus dombeyi roots. Tree Physiol. 2009, 29, 1047–1057. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Chao, L.; Zhou, M.; Hong, M.; Luo, L.; Wang, L.; Ying, W.; Cai, J.; Songjie, G.; Hong, F. Oxidative damages of maize seedlings caused by exposure to a combination of potassium deficiency and salt stress. Plant Soil 2011, 340, 443–452. [Google Scholar] [CrossRef]
- Munns, R. Genes and salt tolerance: Bringing them together. New Phytol. 2005, 167, 645–663. [Google Scholar] [CrossRef]
- Shi, L.; Wang, J.; Liu, B.; Nara, K.; Lian, C.; Shen, Z.; Xia, Y.; Chen, Y. Ectomycorrhizal fungi reduce the light compensation point and promote carbon fixation of Pinus thunbergii seedlings to adapt to shade environments. Mycorrhiza 2017, 27, 823–830. [Google Scholar] [CrossRef]
- Wang, B.; Yu, M.-K.; Sun, H.-J.; Cheng, X.-R. Photosynthetic characters of Quercus acutissima from different provenances under effects of salt stress. Chin. J. Appl. Ecol. 2009, 20, 1817–1824. [Google Scholar]
- Chen, W.; Feng, C.; Guo, W.; Shi, D.; Yang, C. Comparative effects of osmotic-, salt- and alkali stress on growth, photosynthesis, and osmotic adjustment of cotton plants. Photosynthetica 2011, 49, 417–425. [Google Scholar] [CrossRef]
- Zhu, L.; Fu, Z.; Zhang, J.; Wang, J. Effects of mycorrhizal fungi on photosynthetic characteristics of Zelkova serrata Thunb. China Acad. J. Electron. Publ. House 2018, 42, 121–127. [Google Scholar]
- Ke, Y.; Zhou, J.; Lu, N.; Zhang, X. Effects of salinity on photosynthetic physiology and chlorophyll fluorescence characteristics of mulberry (Moms alba) seedlings. For. Res. 2009, 22, 200–206. [Google Scholar] [CrossRef]
- Wu, N.; Li, Z.; Liu, H.; Tang, M. Influence of arbuscular mycorrhiza on photosynthesis and water status of Populus cathayana Rehder males and females under salt stress. Acta Physiol. Plant. 2015, 37, 183. [Google Scholar] [CrossRef]
- Koca, H.; Bor, M.; Özdemir, F.; Türkan, İ. The effect of salt stress on lipid peroxidation, antioxidative enzymes and proline content of sesame cultivars. Environ. Exp. Bot. 2007, 60, 344–351. [Google Scholar] [CrossRef]
- Jia, X.M.; Zhu, Y.F.; Wang, H. Study on physiological response of Malus halliana to saline-alkali stress. Acta Ecol. Sin. 2019, 39, 6349–6361. [Google Scholar]
- Sharifi, M.; Ghorbanli, M.; Ebrahimzadeh, H. Improved growth of salinity-stressed soybean after inoculation with salt pre-treated mycorrhizal fungi. J. Plant Physiol. 2007, 164, 1144–1151. [Google Scholar] [CrossRef]
- HAO, H.; Cao, L.; Chen, W. Effects of salt stress on the ion balance and physiological-biochemical characteristics of Quercus dentata seedlings. Acta Ecol. Sin. 2020, 40, 6897–6904. [Google Scholar]
- Ghorbanli, M.; Ebrahimzadeh, H.; Sharifi, M. Effects of NaCl and mycorrhizal fungi on antioxidative enzymes in soybean. Biol. Plant. 2004, 48, 575–581. [Google Scholar] [CrossRef]
- Lu, W.; Li, Y. Experimental Course of Plant Physiology; China Forestry Publishing House: Beijing, China, 2012; Volume 1. [Google Scholar]
- Shi, S.; Ya, Y.; Wei, L. Experimental Guidance of Plant Physiology; China Forestry Publishing House: Beijing, China, 2011; Volume 4. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Root Radius (μm) | Vascular Bundle Radius (μm) | Vascular Bundl Radius/Root Radius | Cortical Cells Thickness (μm) | Fungus Sheath Thickness (μm) | |
|---|---|---|---|---|---|---|
| Control group | 10 d | 42.46 e | 22.96 f | 0.54 b | ─ | ─ |
| 20 d | 44.74 e | 24.08 e | 0.54 b | ─ | ─ | |
| 30 d | 52.20 d | 40.19 a | 0.77 a | ─ | ─ | |
| G. viscidus group | 10 d | 70.34 c | 24.91 e | 0.35 c | 47.58 c | 16.43 a |
| 20 d | 75.98 b | 26.56 d | 0.35 c | 50.52 b | 16.98 a | |
| 30 d | 81.27 a | 27.42 cd | 0.34 d | 54.48 a | 17.17 a | |
| S. luteus group | 10 d | 77.37 b | 26.37 d | 0.34 cd | 50.94 b | 14.31 b |
| 20 d | 79.99 ab | 28.41 c | 0.36 c | 51.62 b | 17.81 a | |
| 30 d | 81.88 a | 30.93 b | 0.38 c | 51.23 b | 17.70 a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, X.-N.; Hao, H.; Hu, Z.-H.; Leng, P.-S. Ectomycorrhizal Inoculation Enhances the Salt Tolerance of Quercus mongolica Seedlings. Plants 2021, 10, 1790. https://doi.org/10.3390/plants10091790
Bai X-N, Hao H, Hu Z-H, Leng P-S. Ectomycorrhizal Inoculation Enhances the Salt Tolerance of Quercus mongolica Seedlings. Plants. 2021; 10(9):1790. https://doi.org/10.3390/plants10091790
Chicago/Turabian StyleBai, Xiao-Ning, Han Hao, Zeng-Hui Hu, and Ping-Sheng Leng. 2021. "Ectomycorrhizal Inoculation Enhances the Salt Tolerance of Quercus mongolica Seedlings" Plants 10, no. 9: 1790. https://doi.org/10.3390/plants10091790
APA StyleBai, X.-N., Hao, H., Hu, Z.-H., & Leng, P.-S. (2021). Ectomycorrhizal Inoculation Enhances the Salt Tolerance of Quercus mongolica Seedlings. Plants, 10(9), 1790. https://doi.org/10.3390/plants10091790