
The Decrease of Leaf Dark Respiration during Water Stress Is Related to Leaf Non-Structural Carbohydrate Pool in Vitis vinifera L.

 ,
,  ,
, 
 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
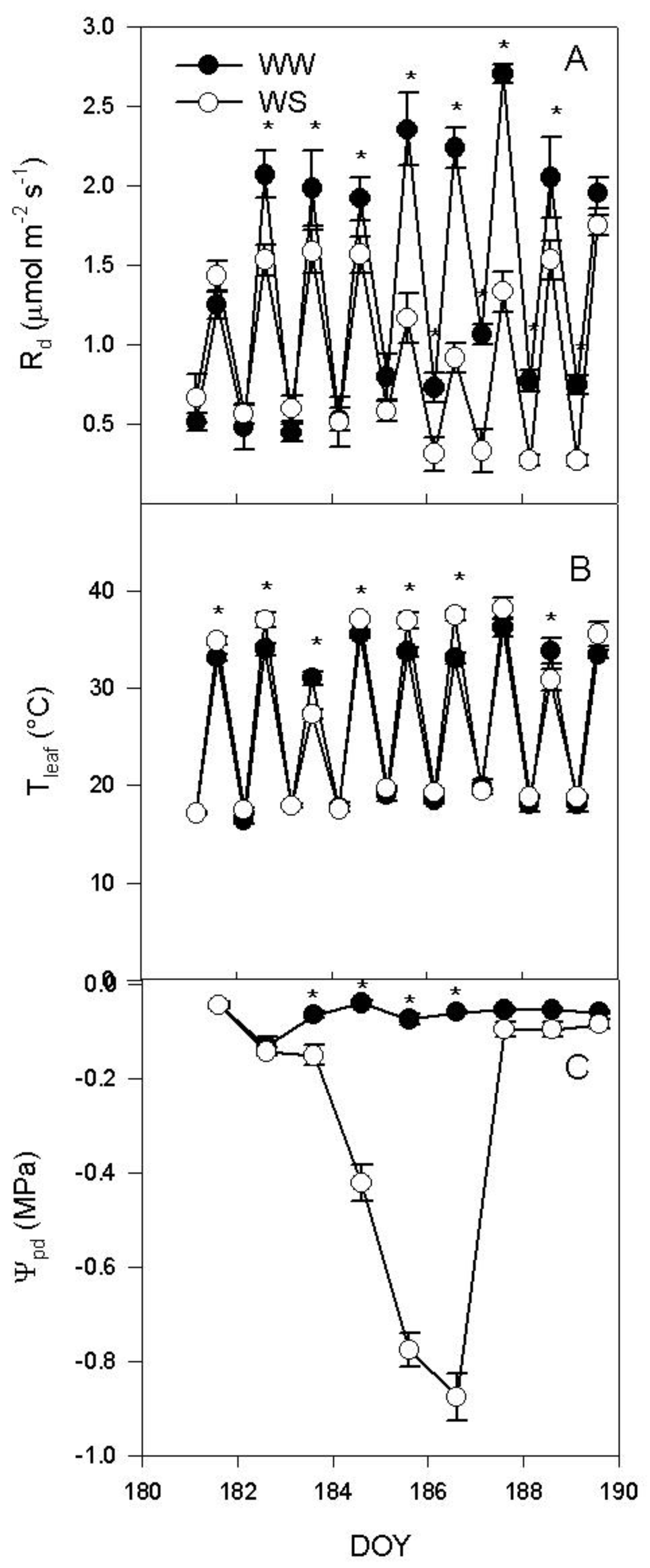
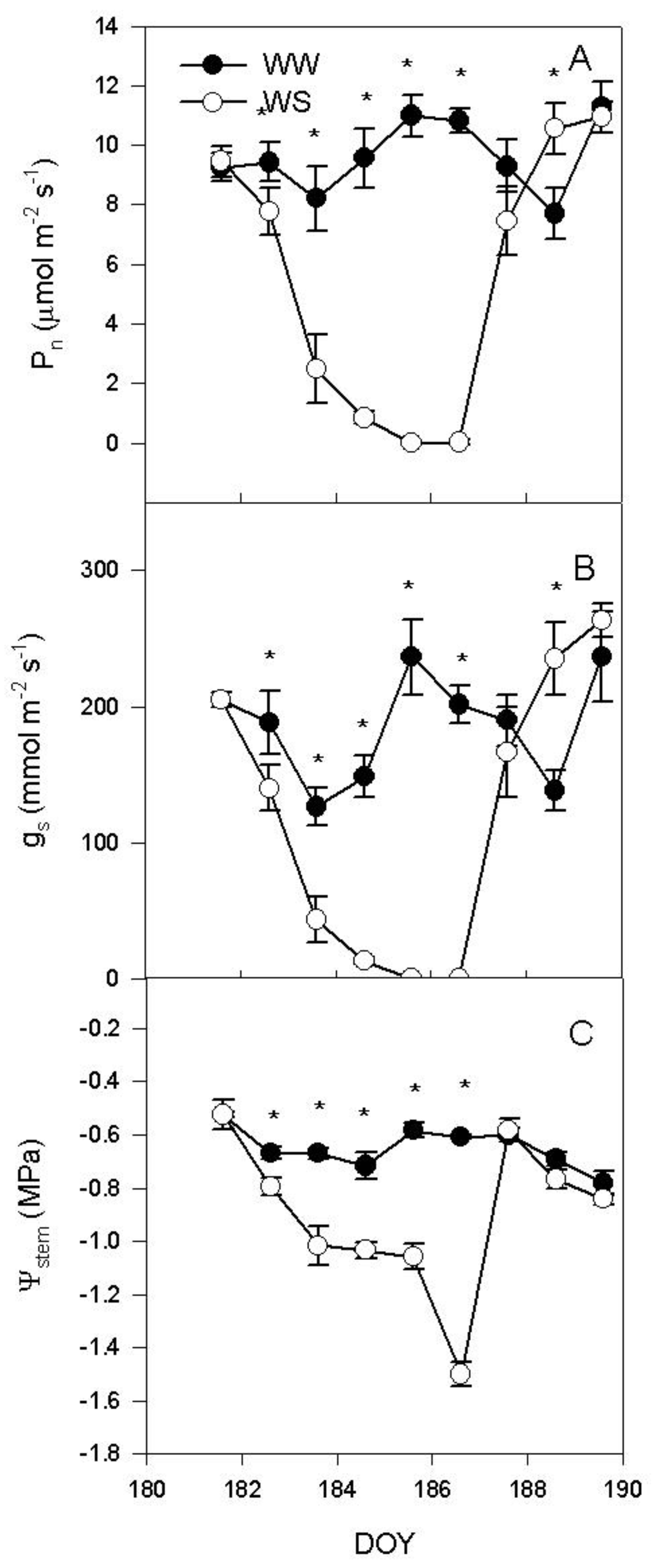
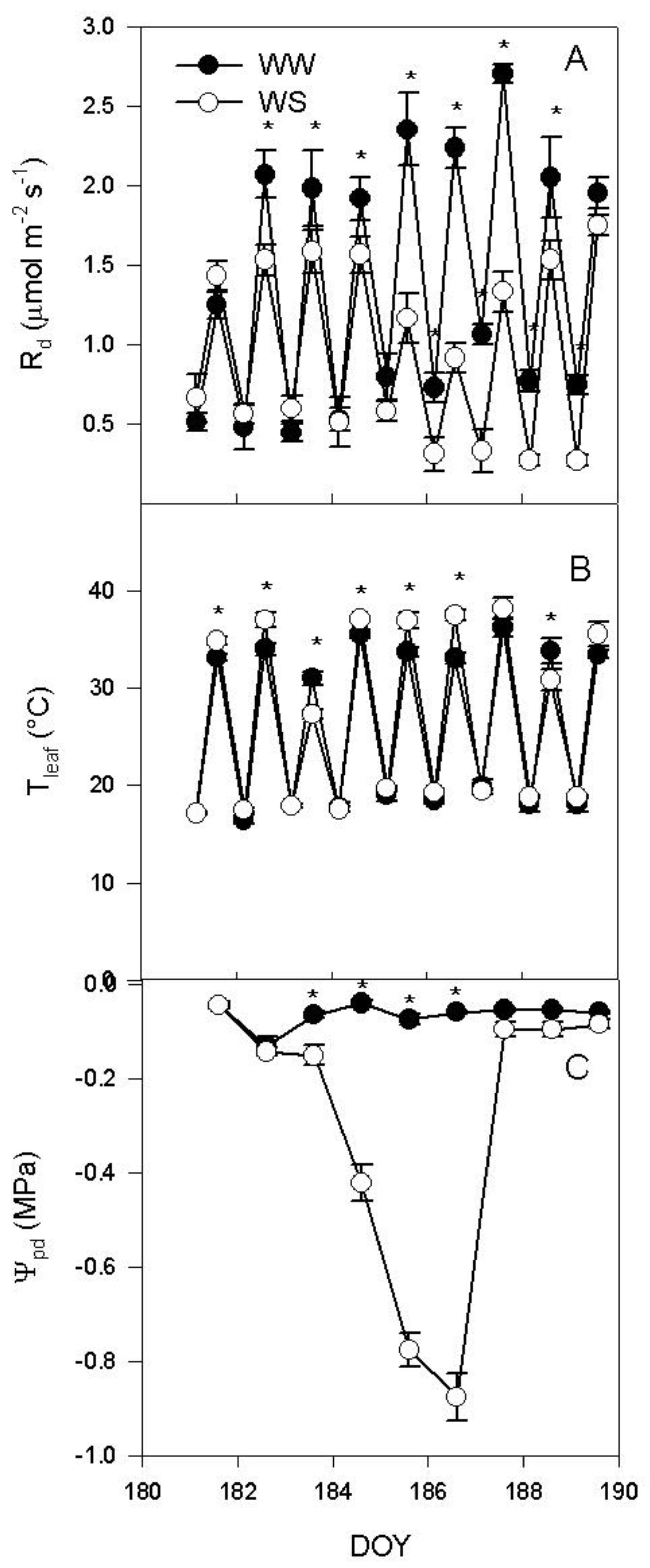
2.1. Water Deprivation and Gas Exchange over the Experiment
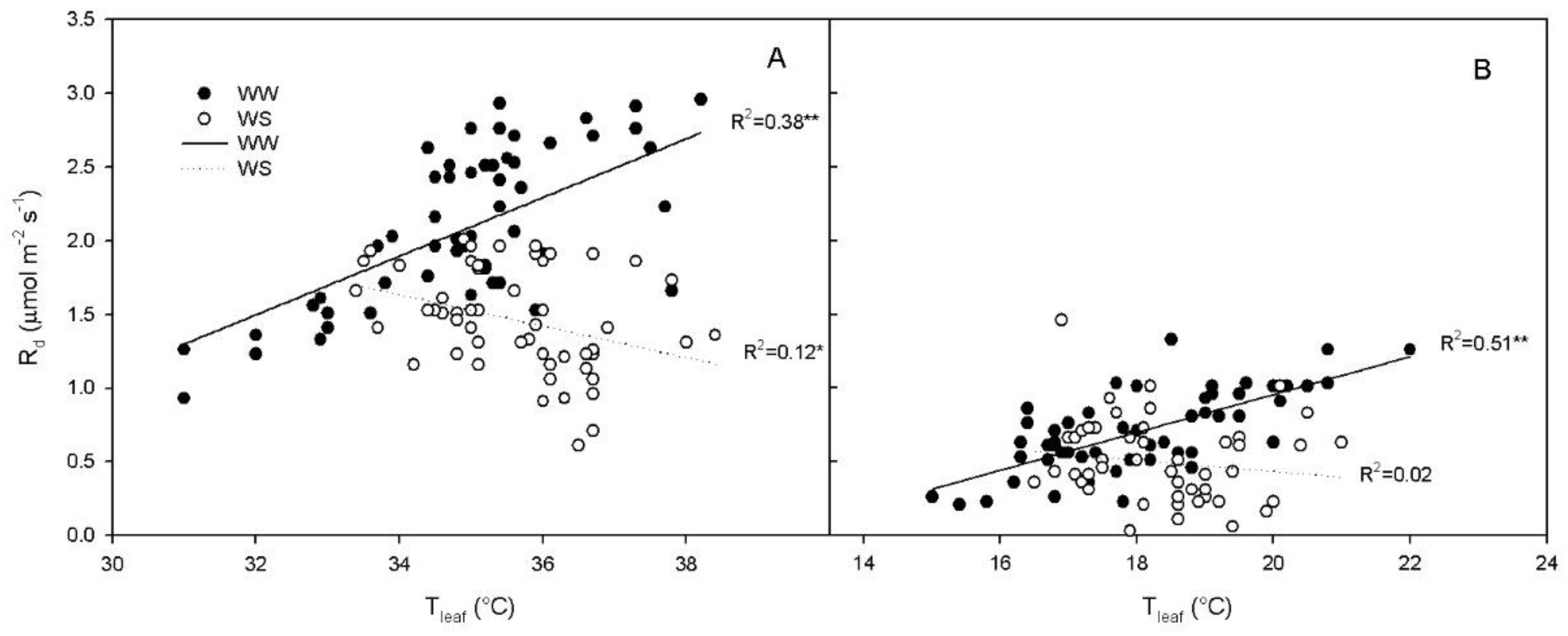
2.2. Rd Regression with Tleaf in WW Vines
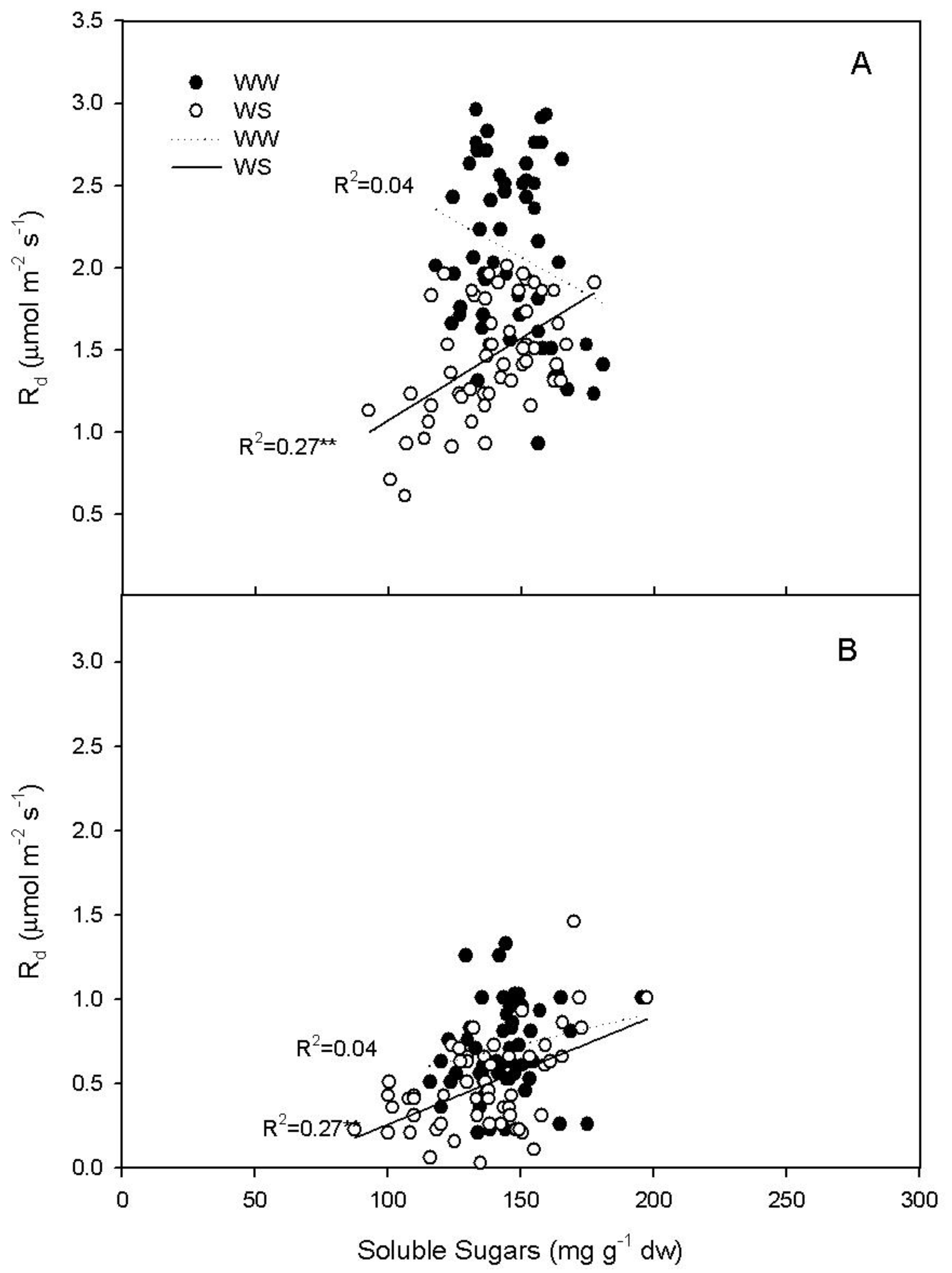
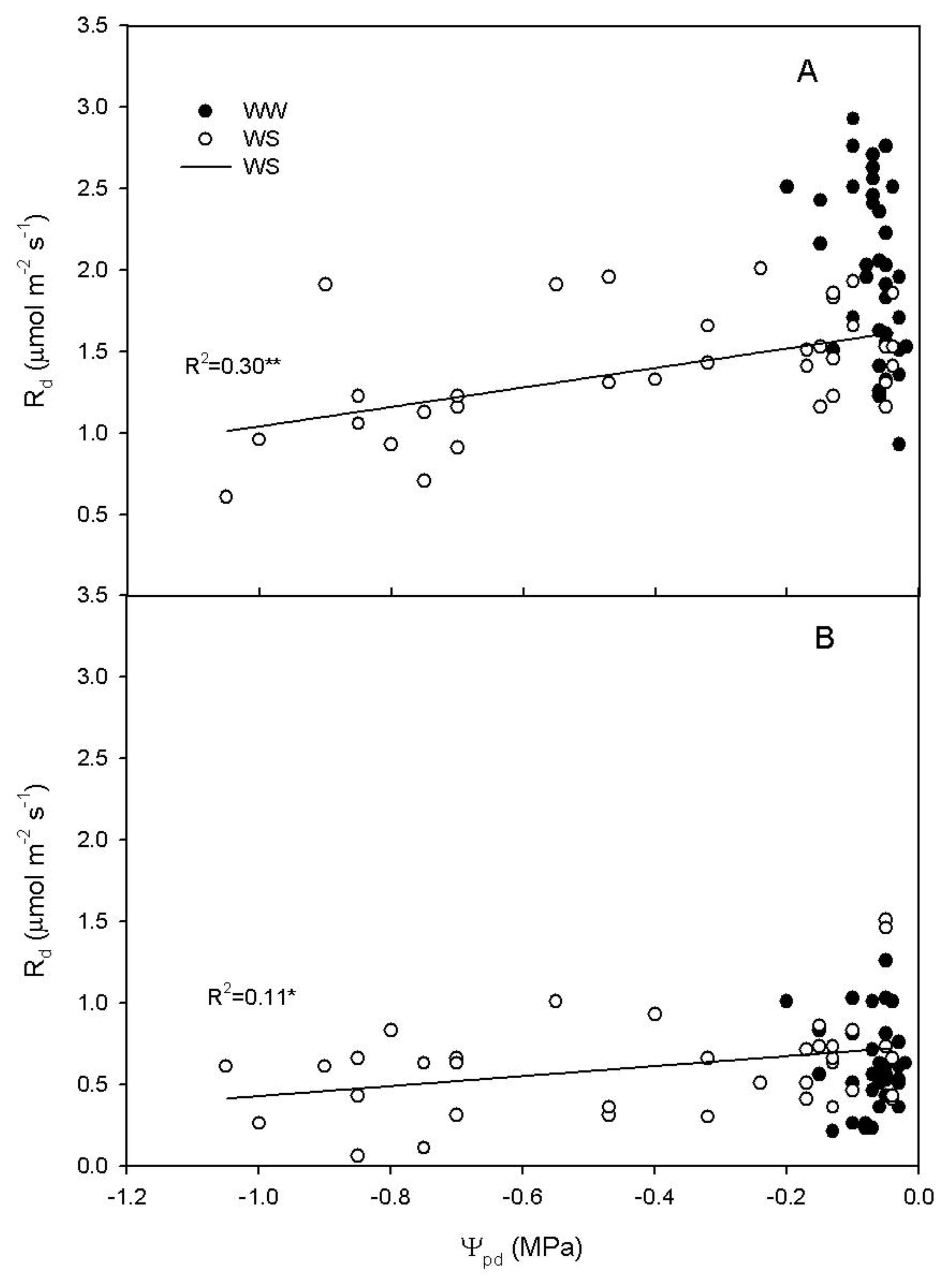
2.3. Rd Regression with Soluble Sugars in WS Vines
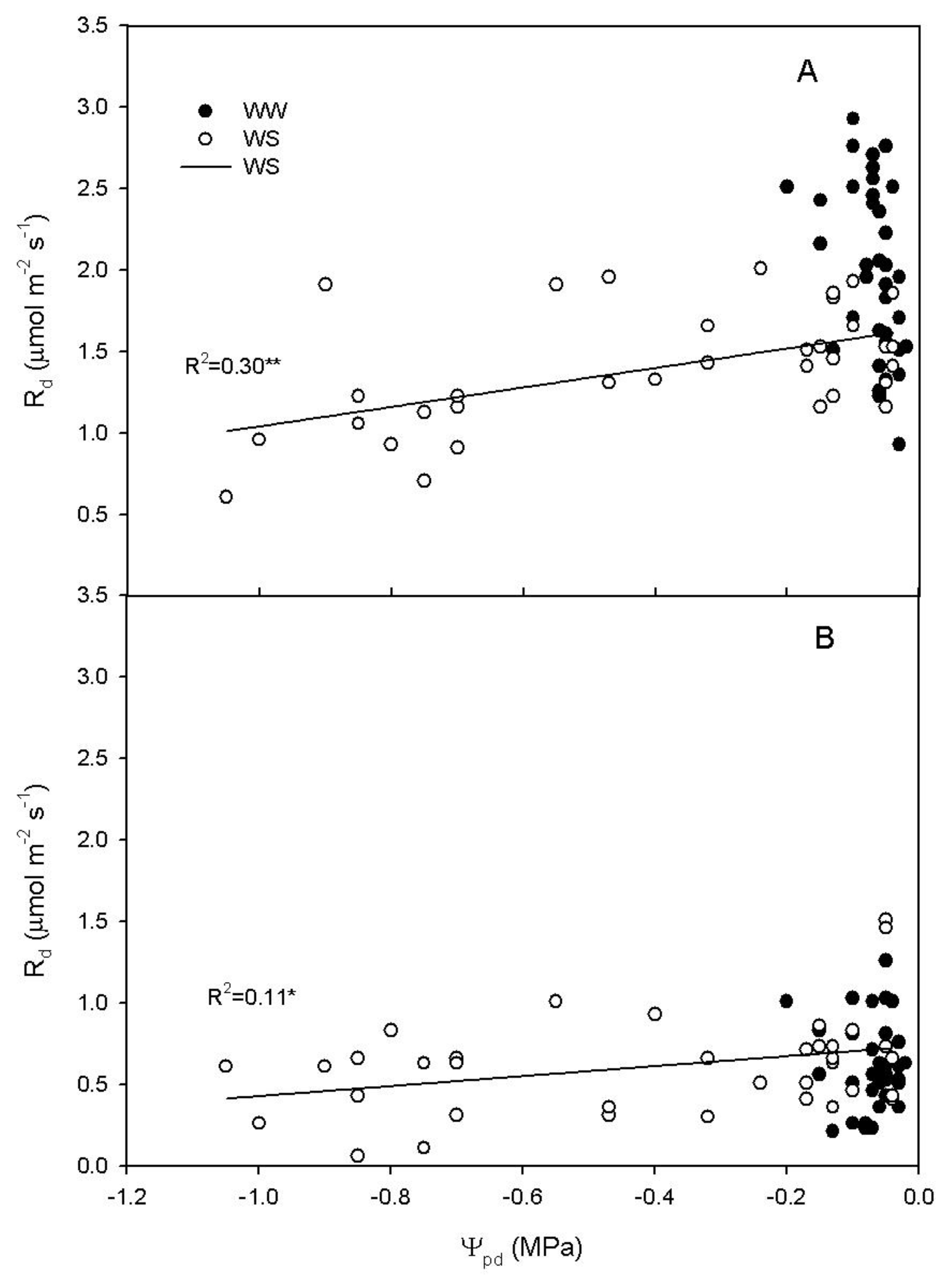
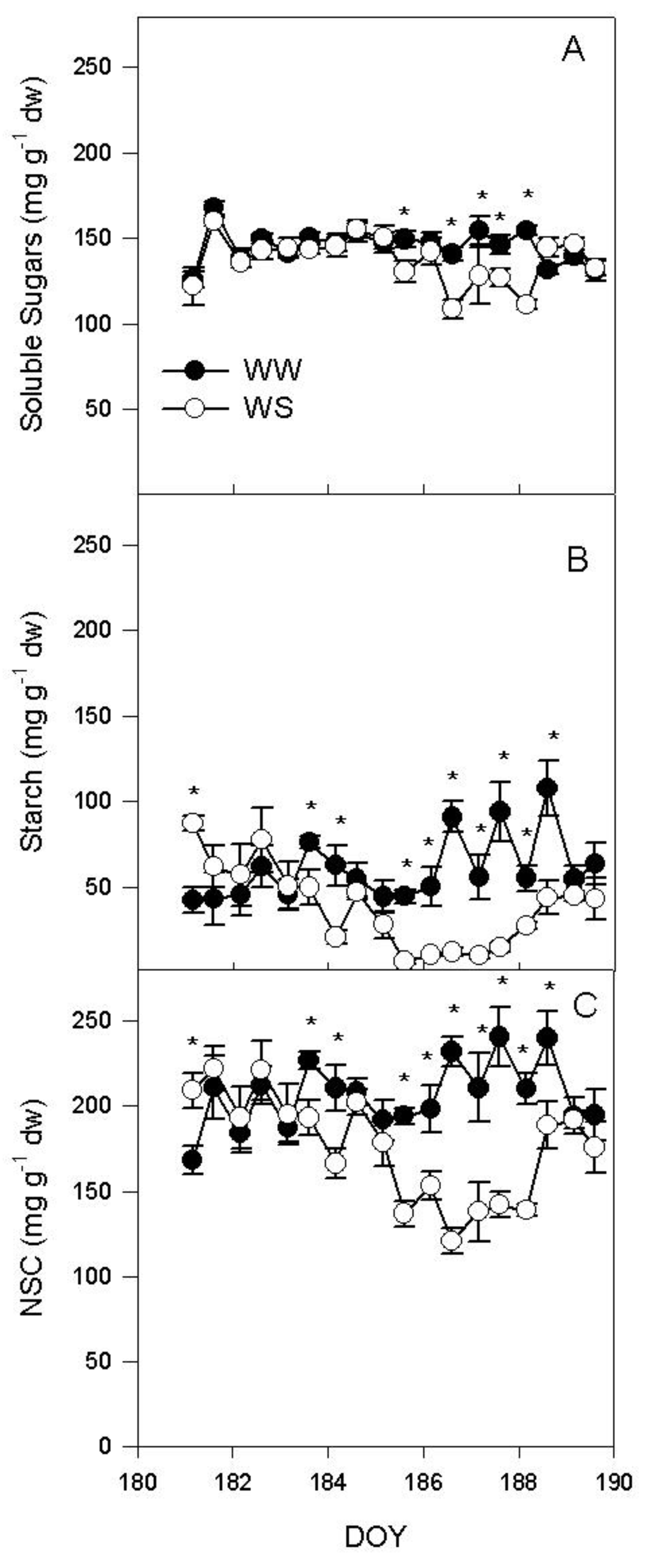
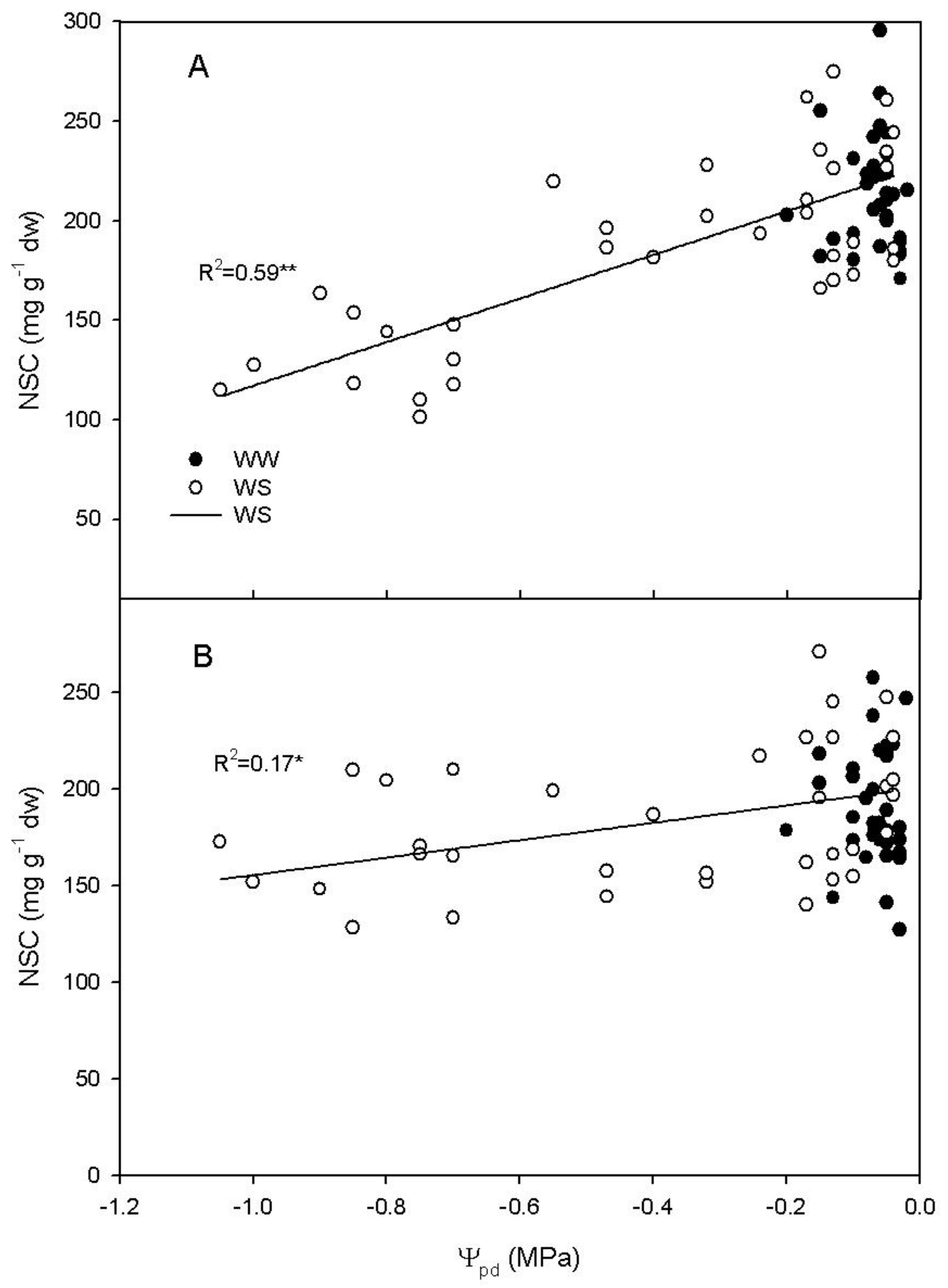
2.4. Relationship between Rd, Ψpd and Non-Structural Carbohydrates
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Gas Exchange
4.3. Water Potential Measurements
4.4. Non-Structural Carbohydrate Determination
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siedow, J.A.; Day, D.A. Respiration and Photorespiration. In Biochemistry and Molecular Biology of Plants; Buchanan, B., Gruissem, W., Jones, R., Eds.; American Society of Plant Physiologists: Rockville, MD, USA, 2000; Volume 30, pp. 676–728. [Google Scholar]
- Amthor, J.S. Respiration and Crop Productivity; Amthor, J.S., Ed.; Springer: New York, NY, USA, 1989. [Google Scholar]
- Escalona, J.M.; Tomas, M.; Martorell, S.; Medrano, H.; Ribas-Carbo, M.; Flexas, J. Carbon Balance in Grapevines under Different Soil Water Supply: Importance of Whole Plant Respiration. Aust. J. Grape Wine Res. 2012, 18, 308–318. [Google Scholar] [CrossRef]
- Kozlowski, T.T.; Kramer, P.J. Physiology of Woody Plants, 2nd ed.; Academic Press: New York, NY, USA, 1979. [Google Scholar]
- Vogt, K. Carbon Budgets of Temperate Forest Ecosystems. Tree Physiol. 1991, 9, 69–86. [Google Scholar] [CrossRef] [PubMed]
- Chiariello, N.R.; Mooney, H.A.; Williams, K. Growth, Carbon Allocation and Cost of Plant Tissues. In Plant Physiological Ecology; Pearcy, R.W., Ehleringer, J.R., Mooney, H.A., Rundel, P.W., Eds.; Springer: Dordrecht, The Netherlands, 1989; p. 533. [Google Scholar]
- Wright, I.J.; Reich, P.B.; Atkin, O.K.; Lusk, C.H.; Tjoelker, M.G.; Westoby, M. Irradiance, Temperature and Rainfall Influence Leaf Dark Respiration in Woody Plants: Evidence from Comparisons across 20 Sites. New Phytol. 2006, 169, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Atkin, O.K.; Evans, J.R.; Ball, M.C.; Lambers, H.; Pons, T.L. Leaf Respiration of Snow Gum in the Light and Dark. Interactions between Temperature and Irradiance. Plant Physiol. 2000, 122, 915–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dusenge, M.E.; Duarte, A.G.; Way, D.A. Plant Carbon Metabolism and Climate Change: Elevated CO2 and Temperature Impacts on Photosynthesis, Photorespiration and Respiration. New Phytol. 2019, 221, 32–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, M.G. Effects of Climate Change on Plant Respiration. Ecol. Appl. 1991, 1, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop Production under Drought and Heat Stress: Plant Responses and Management Options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osório, J.; Osório, M.L.; Chaves, M.M.; Pereira, J.S. Water Deficits Are More Important in Delaying Growth than in Changing Patterns of Carbon Allocation in Eucalyptus Globulus. Tree Physiol. 1998, 18, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Chaves, M.M.; Pereira, J.S.; Maroco, J.; Rodrigues, M.L.; Ricardo, C.P.; Osório, M.L.; Carvalho, I.; Faria, T.; Pinheiro, C. How Plants Cope with Water Stress in the Field. Photosynthesis and Growth. Ann. Bot. 2002, 89, 907–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Del-Campo, M.; Baeza, P.; Ruiz, C.; Lissarrague, J.R. Effects of Water Stress on Dry Matter Content and Partitioning in Four Grapevine Cultivars (Vitis Vinifera L.). J. Int. Sci. Vigne Vin 2005, 39, 1–10. [Google Scholar] [CrossRef]
- Lambers, H.; Chapin, F.; Pons, T. Plant Physiological Ecology; Springer Inc.: New York, NY, USA, 1998. [Google Scholar]
- Gates, D.M. Transpiration and Leaf Temperature. Annu. Rev. Plant Physiol. 1968, 19, 211–238. [Google Scholar] [CrossRef]
- Patakas, A.; Nikolaou, N.; Zioziou, E.; Radoglou, K.; Noitsakis, B. The Role of Organic Solute and Ion Accumulation in Osmotic Adjustment in Drought-Stressed Grapevines. Plant Sci. 2002, 163, 361–367. [Google Scholar] [CrossRef]
- Flexas, J.; Galmes, J.; Ribas-Carbo, M.; Medrano, H. The Effects of Water Stress on Plant Respiration. In Plant Respiration: From Cell to Ecosystem; Lambers, H., Ribas-Carbo, M., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 85–94. [Google Scholar]
- Brix, H. The Effect of Water Stress on the Rates of Photosynthesis and Respiration in Tomato and Loblolly Pine Seedlings. Physiol. Plant. 1962, 15, 10–20. [Google Scholar] [CrossRef]
- Gonzalez-Meler, M.A.; Matamala, R.; Penuelas, J. Effects of Prolonged Drought Stress and Nitrogen Deficiency on the Respiratory O2 Uptake of Bean and Pepper Leaves. Photosynthetica 1997, 34, 505–512. [Google Scholar] [CrossRef]
- Escalona, J.M.; Flexas, J.; Medrano, H. Stomatal and Non-Stomatal Limitations of Photosynthesis under Water Stress in Field-Grown Grapevine. Aust. J. Plant Physiol. 1999, 26, 421–433. [Google Scholar] [CrossRef]
- Haupt-Herting, S.; Klug, K.; Fock, H.P. A New Approach to Measure Gross CO2 Fluxes in Leaves. Gross CO2 Assimilation, Photorespiration, and Mitochondrial Respiration in the Light in Tomato under Drought Stress. Plant Physiol. 2001, 126, 388–396. [Google Scholar] [CrossRef] [Green Version]
- Palta, J.A.; Nobel, P. Root Respiration for Agave Deserti: Influence of Temperature, Water Status and Root Age on Daily Patterns. J. Exp. Bot. 1989, 40, 181–186. [Google Scholar] [CrossRef]
- Brown, K.W.; Thomas, J.C. The Influence of Water Stress Preconditioning on Dark Respiration. Physiol. Plant. 1980, 49, 205–209. [Google Scholar] [CrossRef]
- Ghashghaie, J.; Duranceau, M.; Badeck, F.W.; Cornic, G.; Adeline, M.T.; Deleens, E. Δ13C of CO2 Respired in the Dark in Relation to Δ13C of Leaf Metabolites: Comparison between Nicotiana Sylvestris and Heliantus Annuus under Drought. Plant. Cell Environ. 2001, 24, 505–515. [Google Scholar] [CrossRef]
- Lawlor, D.W. Water Stress Induced Changes in Photosynthesis, Photorespiration, Respiration and CO2 Compensation Concentration of Wheat. Photosynthetica 1976, 10, 378–387. [Google Scholar]
- Loboda, T. Gas Exchange of Different Spring Cereal Genotypes under Normal and Drought Conditions. Photosynthetica 1993, 29, 567–572. [Google Scholar]
- Shearman, L.L.; Esatin, J.D.; Sullivan, C.Y.; Kinbacher, E.J. Carbon Dioxide Exchange in Water-Stressed Maize Sorghum. Crop Sci. 1972, 12, 406–409. [Google Scholar] [CrossRef]
- Upchurch, R.P.; Peterson, M.L.; Hagan, R.M. Effect of Soil-Moisture Content on the Rate of Photosynthesis and Respiration in Ladino Clover (Trifolium Repens L.). Plant Physiol. 1955, 30, 297–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zagdanska, B. Respiratory Energy Demand for Protein Turnover and Ion Transport in Wheat Leaves upon Water Demand. Physiol. Plant 1995, 95, 428–436. [Google Scholar] [CrossRef]
- Schultz, H.R. Water Relations and Photosynthetic Responses of Two Grapevine Cultivars of Different Geographical Origin during Water Stress. Acta Hortic. 1996, 427, 251–266. [Google Scholar] [CrossRef]
- Souza de, C.R.; Maroco, J.P.; dos Santos, T.; Rodrigues, M.; Lopes, C.; Pereira, J.; Chaves, M. Partial Rootzone Drying: Regulation of Stomatal Aperture and Carbon Assimilation in Field-Grown Grapevines (Vitis Vinifera Cv Moscatel). Funct. Plant Biol. 2003, 30, 653–662. [Google Scholar] [CrossRef]
- Zufferey, V. Leaf Respiration in Grapevine (Vitis Vinifera ’Chasselas’) in Relation to Environmental and Plant Factors. Vitis J. Grapevine Res. 2016, 55, 65–72. [Google Scholar] [CrossRef]
- Perez-Martin, A.; Flexas, J.; Ribas-Carbò, M.; Bota, J.; Tomàs, M.; Infante, J.M.; Diaz-Espejo, A. Interactive Effects of Soil Water Deficit and Air Vapour Pressure Deficit on Mesophyll Conductance to CO2 in Vitis Vinifera and Olea Europaea. J. Exp. Bot. 2009, 60, 2391–2405. [Google Scholar] [CrossRef]
- Azcon-Bieto, J.; Osmond, C.B. Relationship between Photosynthesis and Respiration. Plant Physiol. 1983, 71, 574–581. [Google Scholar] [CrossRef] [Green Version]
- Dayer, S.; Prieto, J.; Galat, E.; JP, P. Leaf Carbohydrate Metabolism in Malbec. Aust. J. Grape Wine Res. 2016, 22, 115–123. [Google Scholar] [CrossRef]
- Ayub, G.; Smith, R.A.; Tissue, D.T.; Atkin, O.K. Impacts of Drought on Leaf Respiration in Darkness and Light in Eucalyptus Saligna Exposed to Industrial-Age Atmospheric CO2 and Growth Temperature. New Phytol. 2011, 190, 1003–1018. [Google Scholar] [CrossRef]
- Loewus, F.A. Improvement in Anthrone Method for Determination of Carbohydrates. Anal. Chem. 1952, 24, 219. [Google Scholar] [CrossRef]
- Morris, D.L. Quantitative Determination of Carbohydrates with Drewood’s Anthrone Reagent. Science 1948, 107, 254–255. [Google Scholar] [CrossRef]
- Chow, P.S.; Landhäusser, S.M. A Method for Routine Measurements of Total Sugar and Starch Content in Woody Plant Tissues. Tree Physiol. 2004, 24, 1129–1136. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tombesi, S.; Frioni, T.; Grisafi, F.; Sabbatini, P.; Poni, S.; Palliotti, A. The Decrease of Leaf Dark Respiration during Water Stress Is Related to Leaf Non-Structural Carbohydrate Pool in Vitis vinifera L. Plants 2022, 11, 36. https://doi.org/10.3390/plants11010036
Tombesi S, Frioni T, Grisafi F, Sabbatini P, Poni S, Palliotti A. The Decrease of Leaf Dark Respiration during Water Stress Is Related to Leaf Non-Structural Carbohydrate Pool in Vitis vinifera L. Plants. 2022; 11(1):36. https://doi.org/10.3390/plants11010036
Chicago/Turabian StyleTombesi, Sergio, Tommaso Frioni, Francesca Grisafi, Paolo Sabbatini, Stefano Poni, and Alberto Palliotti. 2022. "The Decrease of Leaf Dark Respiration during Water Stress Is Related to Leaf Non-Structural Carbohydrate Pool in Vitis vinifera L." Plants 11, no. 1: 36. https://doi.org/10.3390/plants11010036
APA StyleTombesi, S., Frioni, T., Grisafi, F., Sabbatini, P., Poni, S., & Palliotti, A. (2022). The Decrease of Leaf Dark Respiration during Water Stress Is Related to Leaf Non-Structural Carbohydrate Pool in Vitis vinifera L. Plants, 11(1), 36. https://doi.org/10.3390/plants11010036