
Towards a Trait-Based Approach to Potentiate Yield under Drought in Legume-Rich Annual Forage Mixtures

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
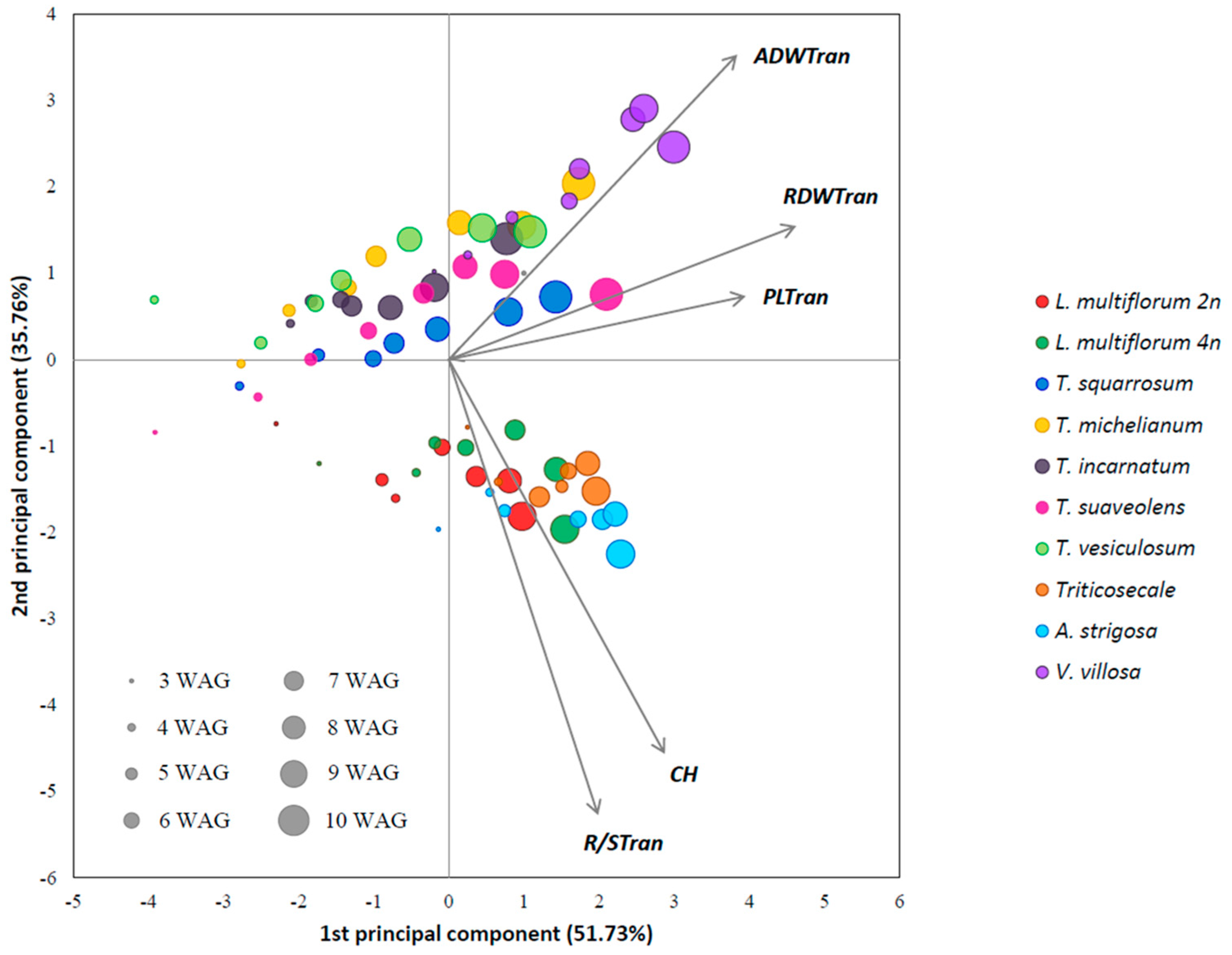
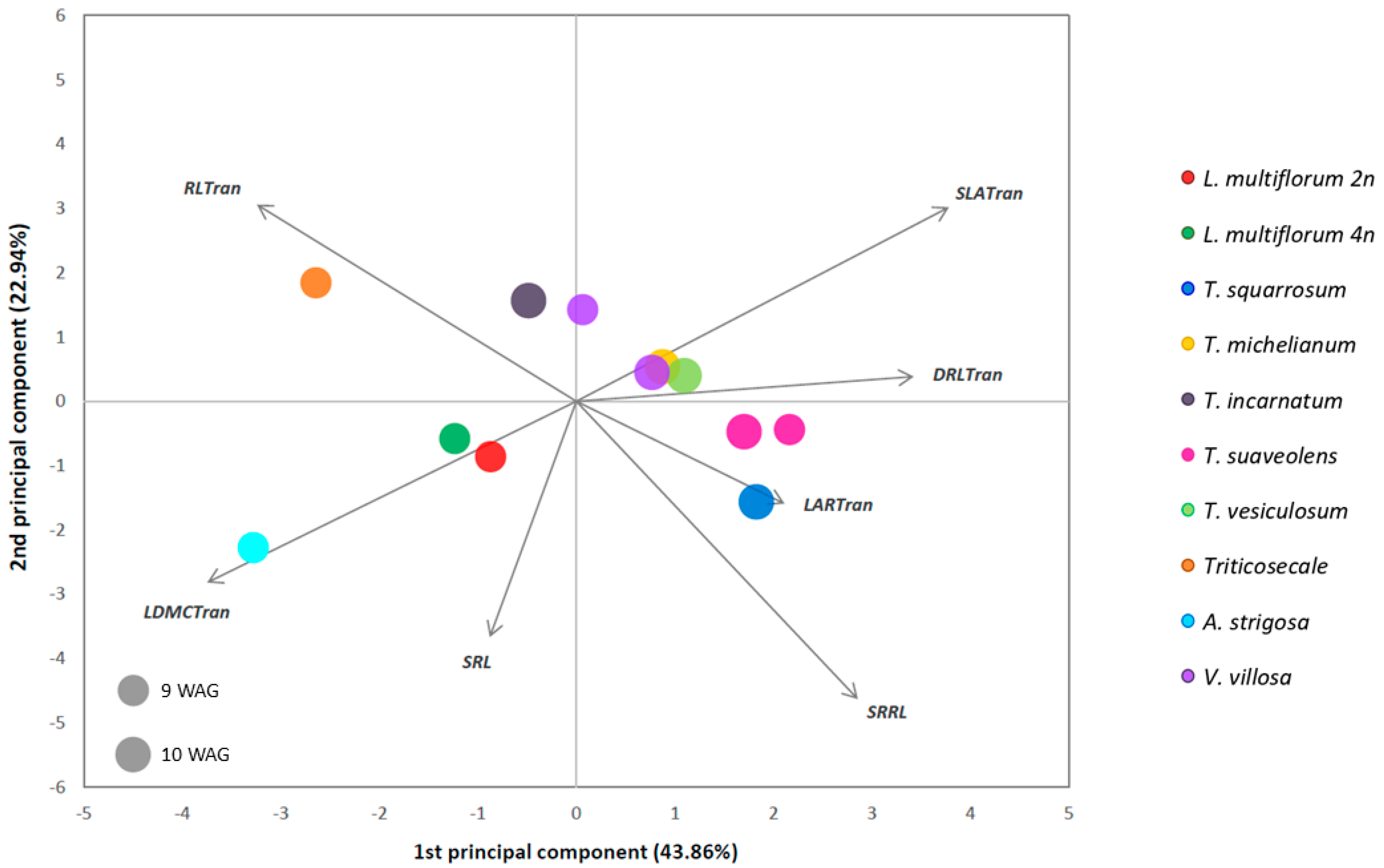
2.1. Morphological Development Characterization
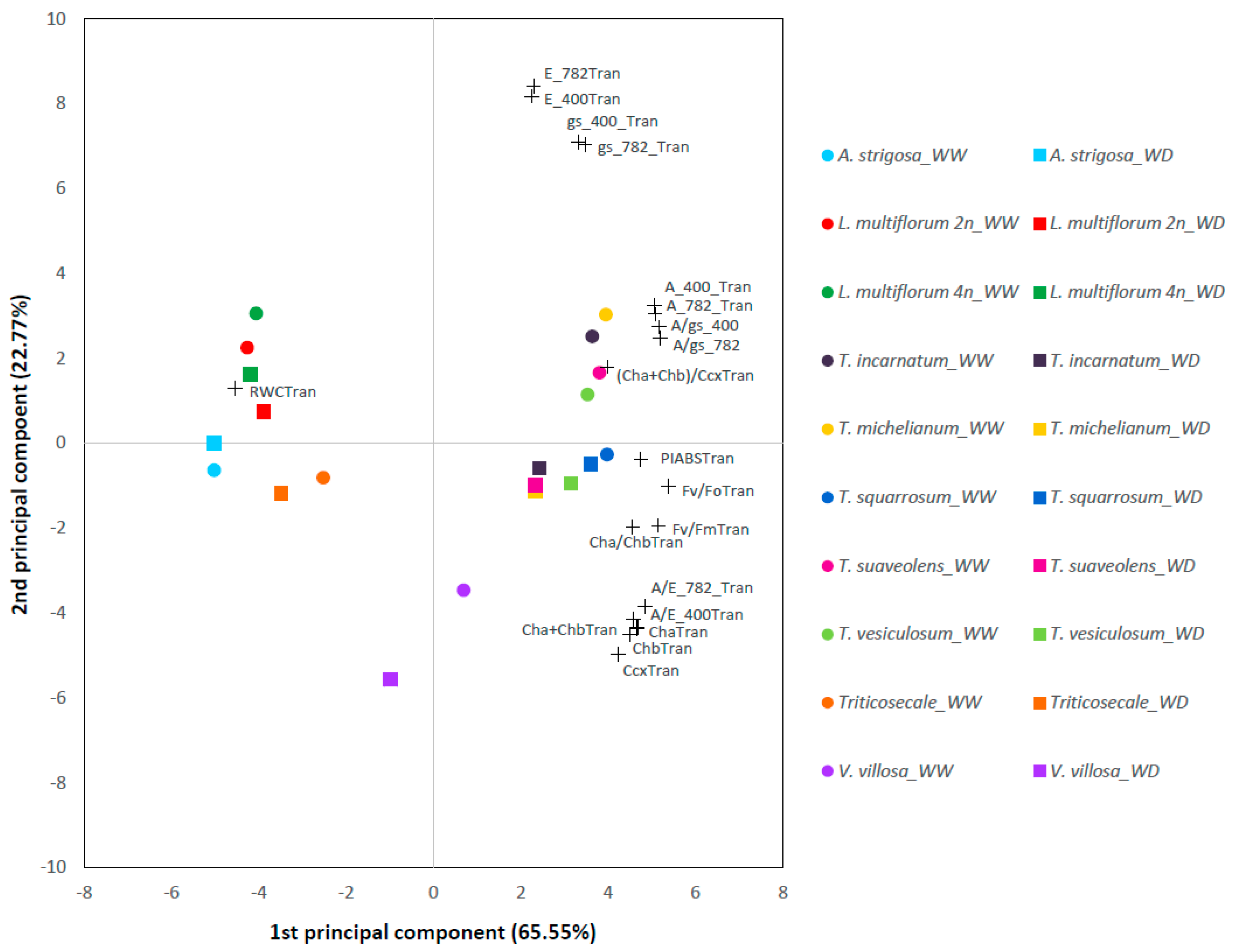
2.2. Photosynthetic Performance under Contrasting Water Regimes
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Species Morphological Development Characterization Experiment
4.3. Species Photosynthetic Performance Characterization under Contrasting Water Regimes Experiment
4.4. Morphological and Physiological Measurements
4.5. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maltoni, S.; Molle, G.; Porqueddu, C.; Connolly, J.; Brophy, C.; Decandia, M. The potential feeding value of grass-legume mixtures in dry Mediterranean conditions. In Proceedings of the Final Meeting of COST Action 852, Irdning, Austria, 7–10 May 2007; pp. 149–152. [Google Scholar]
- Lüscher, A.; Mueller-Harvey, I.; Soussana, J.F.; Rees, R.M.; Peyraud, J.L. Potential of legume-based grassland–livestock systems in Europe: A review. Grass Forage Sci. 2014, 69, 206–228. [Google Scholar] [CrossRef]
- Castro-Montoya, J.M.; Dickhoefer, U. The nutritional value of tropical legume forages fed to ruminants as affected by their growth habit and fed form: A systematic review. Anim. Feed Sci. Technol. 2020, 269, 114641. [Google Scholar] [CrossRef]
- Tuel, A.; Eltahir, E.A.B. Why Is the Mediterranean a Climate Change Hot Spot? J. Clim. 2020, 33, 5829–5843. [Google Scholar] [CrossRef]
- Cramer, W.; Guiot, J.; Fader, M.; Garrabou, J.; Gattuso, J.-P.; Iglesias, A.; Lange, M.A.; Lionello, P.; Llasat, M.C.; Paz, S.; et al. Climate change and interconnected risks to sustainable development in the Mediterranean. Nat. Clim. Chang. 2018, 8, 972–980. [Google Scholar] [CrossRef] [Green Version]
- Tilman, D.; Downing, J.A. Biodiversity and stability in grasslands. Nature 1994, 367, 363–365. [Google Scholar] [CrossRef]
- Watson, L. David Crespo takes C3 pastures the next step to boost soil carbon. Aust. Farm J. 2010, 4, 44–47. [Google Scholar]
- Jongen, M.; Förster, A.C.; Unger, S. Overwhelming effects of autumn-time drought during seedling establishment impair recovery potential in sown and semi-natural pastures in Portugal. Plant Ecol. 2019, 220, 183–197. [Google Scholar] [CrossRef]
- Li, L.; Sun, J.; Zhang, F. Intercropping with wheat leads to greater root weight density and larger below-ground space of irrigated maize at late growth stages. Soil Sci. Plant Nutr. 2011, 57, 61–67. [Google Scholar] [CrossRef]
- Li, L.; Sun, J.; Zhang, F.; Guo, T.; Bao, X.; Smith, F.A.; Smith, S.E. Root distribution and interactions between intercropped species. Oecologia 2006, 147, 280–290. [Google Scholar] [CrossRef]
- Blum, A. Effective use of water (EUW) and not water-use efficiency (WUE) is the target of crop yield improvement under drought stress. Field Crop. Res. 2009, 112, 119–123. [Google Scholar] [CrossRef]
- Ergon, Å.; Seddaiu, G.; Korhonen, P.; Virkajärvi, P.; Bellocchi, G.; Jørgensen, M.; Østrem, L.; Reheul, D.; Volaire, F. How can forage production in Nordic and Mediterranean Europe adapt to the challenges and opportunities arising from climate change? Eur. J. Agron. 2018, 92, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Qin, Y.; Chai, Q.; Feng, F.; Zhao, C.; Yu, A. Interspecies Interactions in Relation to Root Distribution Across the Rooting Profile in Wheat-Maize Intercropping Under Different Plant Densities. Front. Plant Sci. 2018, 9, 483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vries, F.T.; Brown, C.; Stevens, C.J. Grassland species root response to drought: Consequences for soil carbon and nitrogen availability. Plant Soil 2016, 409, 297–312. [Google Scholar] [CrossRef]
- Hofer, D.; Suter, M.; Buchmann, N.; Lüscher, A. Severe water deficit restricts biomass production of Lolium perenne L. and Trifolium repens L. and causes foliar nitrogen but not carbohydrate limitation. Plant Soil 2017, 421, 367–380. [Google Scholar] [CrossRef] [Green Version]
- Barot, S.; Allard, V.; Cantarel, A.; Enjalbert, J.; Gauffreteau, A.; Goldringer, I.; Lata, J.-C.; Le Roux, X.; Niboyet, A.; Porcher, E. Designing mixtures of varieties for multifunctional agriculture with the help of ecology. A review. Agron. Sustain. Dev. 2017, 37, 13. [Google Scholar] [CrossRef] [Green Version]
- Wellstein, C.; Poschlod, P.; Gohlke, A.; Chelli, S.; Campetella, G.; Rosbakh, S.; Canullo, R.; Kreyling, J.; Jentsch, A.; Beierkuhnlein, C. Effects of extreme drought on specific leaf area of grassland species: A meta-analysis of experimental studies in temperate and sub-Mediterranean systems. Glob. Chang. Biol. 2017, 23, 2473–2481. [Google Scholar] [CrossRef]
- Blumenthal, D.M.; Mueller, K.E.; Kray, J.A.; Ocheltree, T.W.; Augustine, D.J.; Wilcox, K.R. Traits link drought resistance with herbivore defence and plant economics in semi-arid grasslands: The central roles of phenology and leaf dry matter content. J. Ecol. 2020, 108, 2336–2351. [Google Scholar] [CrossRef]
- Wasaya, A.; Zhang, X.; Fang, Q.; Yan, Z. Root Phenotyping for Drought Tolerance: A Review. Agronomy 2018, 8, 241. [Google Scholar] [CrossRef] [Green Version]
- Foxx, A.J.; Fort, F. Root and shoot competition lead to contrasting competitive outcomes under water stress: A systematic review and meta-analysis. PLoS ONE 2019, 14, e0220674. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.R. Improving Photosynthesis. Plant Physiol. 2013, 162, 1780–1793. [Google Scholar] [CrossRef] [Green Version]
- Islam, M.A.; Ashilenje, D.S. Diversified Forage Cropping Systems and Their Implications on Resilience and Productivity. Sustainability 2018, 10, 3920. [Google Scholar] [CrossRef] [Green Version]
- Bolinder, M.A.; Angers, D.; Bélanger, G.; Michaud, R.; Laverdière, M. Root biomass and shoot to root ratios of perennial forage crops in eastern Canada. Can. J. Plant Sci. 2002, 82, 731–737. [Google Scholar] [CrossRef] [Green Version]
- Hughes, A.R.; Stachowicz, J.J. Genetic diversity enhances the resistance of a seagrass ecosystem to disturbance. Proc. Natl. Acad. Sci. USA 2004, 101, 8998–9002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook-Patton, S.C.; McArt, S.H.; Parachnowitsch, A.L.; Thaler, J.S.; Agrawal, A.A. A direct comparison of the consequences of plant genotypic and species diversity on communities and ecosystem function. Ecology 2011, 92, 915–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuppinger-Dingley, D.; Schmid, B.; Petermann, J.S.; Yadav, V.; De Deyn, G.B.; Flynn, D.F. Selection for niche differentiation in plant communities increases biodiversity effects. Nature 2014, 515, 108–111. [Google Scholar] [CrossRef]
- Craine, J.M.; Dybzinski, R. Mechanisms of plant competition for nutrients, water and light. Funct. Ecol. 2013, 27, 833–840. [Google Scholar] [CrossRef]
- Weih, M. Trade-offs in plants and the prospects for breeding using modern biotechnology. New Phytol. 2003, 158, 7–9. [Google Scholar] [CrossRef]
- Wintermans, J.E.G.; De Mots, A. Spectrophotometric characteristics of chlorophyll a and b and their phaeophytins in ehanol. Biochim. Biophys. Acta 1965, 109, 448–453. [Google Scholar] [CrossRef]
- Kiær, L.P.; Scherber, C.; Weih, M.; Rubiales, D.; Tavoletti, S.; Adam, E.; Vaz Patto, M.C.; Leitão, S.T.; Schmutz, A.; Schöb, C.; et al. Handbook of Protocols to Assess Traits in Plant Teams, Developed by the EU-H2020 Project DIVERSify (‘Designing Innovative Plant Teams for Ecosystem Resilience and Agricultural Sustainability’), Funded by the European Union’s Horizon 2020 Research and Innovation programme under Grant Agreement Number 727824. Handbook for Trait Assessment in Agricultural Plant Teams. Deliverable Report D2.2 (D17). 2020. Available online: https://orgprints.org/id/eprint/38654/ (accessed on 22 August 2021).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leitão, S.T.; Alves, M.L.; Pereira, P.; Zerrouk, A.; Godinho, B.; Barradas, A.; Vaz Patto, M.C. Towards a Trait-Based Approach to Potentiate Yield under Drought in Legume-Rich Annual Forage Mixtures. Plants 2021, 10, 1763. https://doi.org/10.3390/plants10091763
Leitão ST, Alves ML, Pereira P, Zerrouk A, Godinho B, Barradas A, Vaz Patto MC. Towards a Trait-Based Approach to Potentiate Yield under Drought in Legume-Rich Annual Forage Mixtures. Plants. 2021; 10(9):1763. https://doi.org/10.3390/plants10091763
Chicago/Turabian StyleLeitão, Susana T., Mara Lisa Alves, Priscila Pereira, Aziza Zerrouk, Bruno Godinho, Ana Barradas, and Maria Carlota Vaz Patto. 2021. "Towards a Trait-Based Approach to Potentiate Yield under Drought in Legume-Rich Annual Forage Mixtures" Plants 10, no. 9: 1763. https://doi.org/10.3390/plants10091763
APA StyleLeitão, S. T., Alves, M. L., Pereira, P., Zerrouk, A., Godinho, B., Barradas, A., & Vaz Patto, M. C. (2021). Towards a Trait-Based Approach to Potentiate Yield under Drought in Legume-Rich Annual Forage Mixtures. Plants, 10(9), 1763. https://doi.org/10.3390/plants10091763